0022-538X/96/$04.0010
Copyrightq1996, American Society for Microbiology
A Short Peptide Eluted from the H-2K
b
Molecule of a
Polyomavirus-Positive Tumor Corresponds to Polyomavirus Large T Antigen
Peptide at Amino Acids 578 to 585 and Induces
Polyomavirus-Specific Immunity
ZSOFIA BERKE,
1* SARAH PALMER,
1† TOMAS BERGMAN,
2DANIELE WESTER,
1JONATHAN SVEDMYR,
1STIG LINDER,
3HANS JO
¨ RNVALL,
2AND
TINA DALIANIS
1Division of Clinical Virology, Karolinska Institutet, Huddinge University Hospital, F67, S-141 86 Huddinge,
1and
Department of Medical Biochemistry and Biophysics, Karolinska Institutet,
2and Department of Experimental
Oncology, Karolinska Hospital,
3S-171 77 Stockholm, Sweden
Received 4 December 1995/Accepted 20 February 1996
A short peptide in complex with the H-2K
bmolecule on PyRMA, a polyomavirus transfectant of the mouse
lymphoma cell line RMA, was identified as a polyomavirus tumor-specific transplantation antigen. The peptide
was obtained by affinity chromatography, acidic extraction, and reverse-phase high-pressure liquid
chroma-tography (HPLC). In one HPLC fraction, a peptide sequence in which 5 of 8 amino acids, GKxGLxxA,
corresponded to residues 578 to 585 of polyomavirus large T antigen was identified. In tumor rejection assays,
we therefore tested three related synthetic peptides, corresponding to the octapeptide LT 578–585,
GKTG-LAAA; the nonapeptide LT 578–586, GKTGLAAAL; and the decapeptide LT 578–587, GKTGLAAALI. The
octapeptide was found to give the most effective immunization against the outgrowth of the polyomavirus
DNA-positive PyRMA tumor. However, none of the three peptides immunized against the original
polyoma-virus-negative RMA line.
Murine polyomavirus, a potentially oncogenic, small DNA
virus, does not induce tumors in its natural host under normal
conditions because of T-cell-dependent immunity (2, 18). The
antigens recognized by the immune system are now suggested
to be peptide antigens derived from the three polyomavirus T
antigens, large T antigen (LT), middle T antigen, and small T
antigen (8, 24, 26).
Previously, it had been shown that immunization of mice
with irradiated polyomavirus tumor cells, wild-type
polyoma-virus, deletion mutants of polyomapolyoma-virus, or vaccinia virus
re-combinant mutants could prevent the outgrowth of
polyoma-virus tumors (9, 10, 17, 29). Later, we demonstrated that
purified middle or small T antigen and short synthetic peptides
derived from the amino acid sequences of the three T antigens
could evoke polyomavirus-specific immunity (23, 24, 26). Since
none of the three T antigens was detected on the outside of the
cell surface, it was suggested that the three T antigens were
processed intracellularly into peptides (24). These peptide
products could then be presented to the immune system
to-gether with determinants of the major histocompatibility
com-plex (MHC) (24, 26). This had been reported previously for the
influenza virus nucleoprotein by Townsend et al. (31) and later
by Ro
¨tzschke et al. (27) and for the vesicular stomatitis virus
nucleocapsid protein by Van Bleek et al. (32).
The aim of this study was to examine if polyomavirus
T-antigen-derived peptides could indeed be presented by MHC
class I molecules on the surfaces of polyomavirus tumor cells
and also if such peptides were immunogenic. The murine
lym-phoma cell line RMA and its polyomavirus transfectant
PyRMA were used for these studies. H-2K
bMHC class
I-bound peptides were eluted from both PyRMA and RMA cells
by affinity chromatography under acidic conditions. The
differ-ent peptides were then separated by reverse-phase
high-pres-sure liquid chromatography (HPLC). In one of the fractions, a
unique PyRMA peptide in which 5 of 8 amino acid residues
(GKxGLxxA) corresponded to amino acid residues 578 to 585
of polyomavirus LT was identified. In tumor rejection tests,
this LT 578–585 octapeptide, GKTGLAAA, was shown to
immunize efficiently against PyRMA cells, but not against
RMA cells.
MATERIALS AND METHODS
Tumor cells.The RMA cell line was derived from the Rauscher leukemia virus-induced mouse T-cell lymphoma RBL-5 of the H-2bhaplotype (19). The PyRMA cell line was obtained by transfecting RMA cells by electroporation (960
mF, 250 V) with plasmid construct Pywt-neo, a modified version of p1023 con-taining the polyomavirus genome cloned into the pBR322 plasmid at the BamHI sites (21). Transfected clones were selected on G418 and were identified by a polyomavirus-specific PCR (Fig. 1), as described previously (5). Expression of LT was confirmed by Northern (RNA) blotting (data not shown). No infectious virus is produced by using this protocol. All cells were grown in tissue culture (RPMI medium supplemented with 5% fetal calf serum,L-glutamine, penicillin, and streptomycin) or as ascites tumors in (C57BL/63A.SW)F1mice.
Peptide extraction.Cells (108) from ascites tumors were lysed in 10 ml of
phosphate-buffered saline (PBS) containing 0.5% Nonidet P-40, 0.1% aprotinin (Boehringer Mannheim Scandinavia, Bromma, Sweden), and 0.1 mM AEBSF (Calbiochem, La Jolla, Calif.) and stirred for 30 min at 48C. Lysates were centrifuged for 30 min at 13,0003g. Supernatants were then subjected to affinity
prepurification on a Prosep A column coated with normal mouse serum and thereafter passed through a Prosep A column coated with anti-H-2Kb
antibodies (AF6-88.5.3; American Type Culture Collection, Rockville, Md.). Bound mate-rial was eluted from the column with citrate buffer at pH 3. Supernatants were centrifuged in Centriprep ultrafiltration tubes stepwise at 100, 30, and 10 kDa according to the manufacturer’s (Amicon; Grace AB, Helsingborg, Sweden) instructions. The filtrate volume was reduced to 3 ml by vacuum evaporation. The separation of peptides contained in the filtrate was performed by reverse-phase HPLC (16).
Reverse-phase HPLC.HPLC analysis were carried out with a Gilson Medical Electronics (Villiers-le-Bel, France) instrument equipped with a dual-wavelength monitor, set at A214and A280. All integrations were performed by using RAININ
* Corresponding author.
† Present address: Department of Virology, Swedish Institute for Infectious Disease Control, S-105 21 Stockholm, Sweden.
3093
on November 9, 2019 by guest
http://jvi.asm.org/
Dynamax HPLC method manager software. Sample volumes of 500 ml were injected onto the HPLC column. Peptides were separated on a Super Pac Pep-S (5mm; 250 by 4 mm) mixed-bed C18/C2column (Pharmacia LKB Biotechnology,
Uppsala, Sweden) by using a gradient of 0 to 60% acetonitrile in 0.1% triflu-oracetic acid for elution, at a constant flow of 1.0 ml/min. For amino acid analysis and peptide sequencing, 1-min fractions were collected.
Peptide analysis.The amino acid compositions of peptides were determined after hydrolysis with 6 M HCl–0.5% phenol in evacuated tubes for 24 h at 1108C. The released amino acids were analyzed by reverse-phase HPLC as phenylthio-carbamyl derivatives (3).
Sequence analysis was carried out with an Applied Biosystems 470A instru-ment. Phenylthiohydantoin derivatives from degradations were analyzed by re-verse-phase HPLC essentially as described by Bergman and Jo¨rnvall (4).
Synthetic peptides corresponding to polyomavirus LT 578–585 (GKTG LAAA), LT 578–586 (GKTGLAAAL), and LT 578–587 (GKTGLAAALI) were obtained from Scandinavian Peptide Synthesis AB, Ko¨ping, Sweden.
Mice.Inbred C57BL/6 (H-2b) and (C57BL/63A.SW)F
1(H-2b3H-2s) mice
were used for tumor rejection tests.
Immunization with polyomavirus, synthetic peptides, and tumor cells.Mice were immunized four times at weekly intervals intraperitoneally with either 53
106PFU of large-plaque strain PA2 of polyomavirus (14) or 6mg of peptide
dissolved in 100ml of PBS or subcutaneously (s.c.) with 23106irradiated
(10,000 R) RMA cells.
Transplantation rejection tests.Living RMA or PyRMA tumor cells (53103
) were inoculated s.c. 1 week after the last immunization in different groups of control or immunized mice. Each group contained five mice. At least three separate experiments were performed for each tumor. In order to exclude non-specific boosting of the immune system by polyomavirus, animals were subjected to 400 R of whole-body irradiation prior to tumor cell inoculation (9). Animals were checked for tumor growth, and each tumor diameter was measured every second or third day for 3 weeks. When tumor diameters were approximately 15 mm, animals were sacrificed in order to avoid dissemination of the tumor and further suffering of animals. The mean tumor load (MTL) was calculated for each group of animals by adding the individual tumor diameters (d11d21dn) and dividing the sum by the total number of animals within the group (n), as described previously (26). The significance of a decrease in MTL was calculated according to the Wilcoxon-Mann-Whitney test (28).
RESULTS
Identification of a polyomavirus-specific H-2K
b-restricted
peptide.
Short peptides bound to the H-2K
bMHC class I
molecule were extracted from PyRMA and RMA cells under
acidic conditions. The extracted material was separated by
reverse-phase HPLC. Comparison of the chromatograms from
RMA and PyRMA cell extracts revealed one PyRMA-specific
peak (compare Fig. 2A and B). Corresponding fractions,
frac-tions 22, from both RMA and PyRMA extracts were subjected
to amino acid sequence and compositional analysis and
pep-tide sequencing. The results of analysis indicated the following
specific amino acid sequence in PyRMA fraction 22: GKxGLxxA,
with 5 of 8 amino acids common to residues 578 to 585 of
polyomavirus LT (30). The amino acid sequence identified in
RMA fraction 22 was ANEQNEQG. Since the exact length of
the peptide was not determined and since H-2K
b-restricted
peptides have been suggested to be 8 or 9 residues long (for
review, see references 11 and 12), three peptides
correspond-ing to this part of the polyomavirus LT were synthesized. These
peptides were 8, 9, and 10 amino acids long with a common N
terminus at residue 578 of LT and different C termini: LT
578–585, GKTGLAAA; LT 578–586, GKTGLAAAL; and LT
578–587, GKTGLAAALI. These peptides were then analyzed
by HPLC, using the same protocol as that for cellular eluates.
Of the three peptides tested, the synthetic nonamer, LT 578–
586, was the one observed to coelute with the specific PyRMA
peak in fraction 22 (Fig. 2C).
A synthetic octamer peptide corresponding to polyomavirus
LT 578–585 immunizes against PyRMA cells, not against
RMA cells.
Groups of C57BL/6 or (C57BL/6
3
A.SW)F
1mice
were immunized with either peptide LT 578–585, LT 578–586,
or LT 578–587, polyomavirus, or irradiated RMA cells and
challenged with living PyRMA or RMA tumor cells. Three
separate experiments were performed for each tumor. In each
experiment, the outgrowth of tumors was monitored and
tu-mor diameters were measured. The MTL was calculated. The
cell number inoculated was determined as the lowest dose
required for 100% tumor outgrowth in control animals and was
5
3
10
3cells per inoculum.
Figure 3 illustrates the outgrowth of PyRMA in immunized
and control mice in one experiment. Immunization with
wild-type polyomavirus or the synthetic octapeptide inhibited
growth of the PyRMA tumor efficiently (Fig. 3). Tumor
out-growth was observed in nonimmunized controls and in mice
inoculated with the nonapeptide (Fig. 3). In this particular
experiment, immunization with the decapeptide provided some
protection, but tumor outgrowth was still extensive (Fig. 3).
Figure 4 summarizes the results of three PyRMA tumor
rejection tests performed in C57BL/6 or (C57BL/6
3
A.SW)F
1mice. The MTLs of PyRMA in different groups of mice given
are those at the time when the MTLs in the controls of all three
experiments were about 5 (Fig. 4A) or 11 (Fig. 4B) mm.
Im-munization with polyomavirus induced a significant reduction
in tumor progression of PyRMA (Fig. 4). In the groups of mice
immunized with polyomavirus, decreases in the MTLs of 90 (P
#
0.0015) and 80% (P
#
0.0015) were observed when the
MTLs of the controls were 5 (Fig. 4A) and 12 (Fig. 4B) mm,
respectively. Immunization with the octapeptide LT 578–585
also inhibited tumor progression of PyRMA. In the groups of
mice immunized with the octapeptide LT 578–585, significant
reductions in the MTLs, 67 (P
#
0.002) and 68% (P
#
0.0035),
were observed when the MTLs of the controls were 5 (Fig. 4A)
and 12 (Fig. 4B) mm, respectively. Both the nonapeptide LT
578–586 and the decapeptide LT 578–587 were less efficient in
their inhibition of tumor outgrowth. The nonapeptide LT 578–
586 induced nonsignificant reductions in the MTLs, 30% when
the MTL of controls was 5 mm (Fig. 4A) and 45% (P
#
0.03)
when the MTL of controls was 12 mm (Fig. 4B). The
decapep-tide LT 578-587 induced reductions in the MTL of 55% (P
#
0.0075) when that of the controls was 5 mm (Fig. 4A) and of 45%
(P
#
0.03) when that of the controls was 12 mm (Fig. 4B).
Outgrowth of the RMA tumor was inhibited efficiently only
in the group of mice immunized with RMA cells (Fig. 5).
Polyomavirus and the three LT peptides had no significant
effect (Fig. 5).
[image:2.612.104.245.71.246.2]Figure 6 illustrates the results of three RMA tumor rejection
tests at the time point when the MTLs of the three control
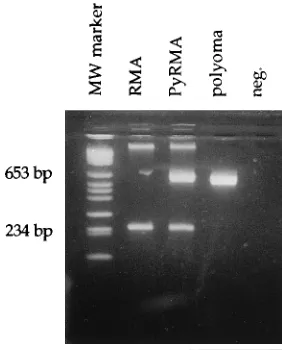
FIG. 1. Detection of polyomavirus DNA in the polyomavirus-transfected PyRMA cell line, not in the RMA cell line. The size of the polyomavirus-specific PCR product was 650 bp, and that of the myogenin-specific product was 245 bp (5). MW, molecular weight; neg., negative.
on November 9, 2019 by guest
http://jvi.asm.org/
groups were around 14 mm. Only immunization with
irradi-ated RMA cells reduced the MTL significantly, 90% (P
#
0.005) (Fig. 6). Immunization with polyomavirus or any of the
polyomavirus LT peptides had no significant (
,
23%)
inhibi-tory effect on RMA tumor outgrowth (Fig. 6).
DISCUSSION
Naturally occurring peptides associated with the H-2K
bMHC class I molecule were eluted off a
polyomavirus-trans-fected RMA (H-2
b) tumor, PyRMA. In one HPLC fraction, a
peptide sequence in which 5 of 8 amino acids (GKxGLxxA)
corresponded to residues 578 to 585 of polyomavirus LT was
identified. Although it is not absolute evidence, this indicates
that a peptide presented on the cell surface of a
polyomavirus-positive tumor in association with the H-2K
bMHC class I
molecule may originate from polyomavirus LT. It is likely that
FIG. 3. In vivo growth of PyRMA in different groups of immunized and control mice after inoculation with an s.c. inoculum of 53103
cells per mouse. MTLs in mice immunized with polyomavirus, LT 578–585, LT 578–586, and LT578–587 and in controls are indicated. The MTL was calculated for five mice at different time points after tumor cell inoculation.
FIG. 2. Separation of peptides by reverse-phase HPLC. Peptides were puri-fied by affinity chromatography on an anti-H-2Kb
-coated Prosep A column from 108
RMA (A) and PyRMA (B) cells. (C) Three synthetic polyomavirus LT peptides, octapeptide LT 578–585, nonapeptide LT 578–586, and decapeptide LT 578–587.
on November 9, 2019 by guest
http://jvi.asm.org/
the peptide presented on the surface of PyRMA is a
nonapep-tide, since this peptide coeluted with the PyRMA-specific
HPLC fraction in which the sequence was identified (Fig. 2B
and C). Furthermore, it has a leucine at position 9, which is in
line with previous reports suggesting that peptides associated
with the H-2K
bmolecule are 8 or 9 amino acids long with an
anchoring leucine at the last position (11, 12).
Since the exact length of the peptide was not determined
after amino acid analysis, three synthetic peptides, an
octapep-tide, a nonapepoctapep-tide, and a decapeptide with the N terminus at
amino acid position 578 of LT, were synthesized and analyzed
further. The octapeptide, GKTGLAAA, was shown to
immu-nize efficiently against a polyomavirus-positive tumor cell
(PyRMA) inoculum in vivo (Fig. 3 and 4). The other two
peptides were not as immunogenic as was the octapeptide. The
immune response to PyRMA cells after peptide immunization
seemed to be polyomavirus specific, since none of the synthetic
LT peptides immunized significantly against the control RMA
tumor (Fig. 5 and 6).
The fact that immunization with the octapeptide resulted in
better protection against the outgrowth of a PyRMA inoculum
than did immunization with the nonapeptide or the
decapep-tide provides an additional aspect to consider when using
syn-thetic peptides as immunogens in vaccine trials. It indicates
that an abundant naturally presented peptide may not be the
most immunogenic. Existing T cells may be triggered more
efficiently by modified versions of the naturally presented
pep-tide. Potentially, by identifying the sequence of naturally
oc-curring peptides and modifying them, we may be able to
im-FIG. 4. In vivo growth of PyRMA in different groups of immunized and control mice after inoculation with an s.c. inoculum of 53103
cells per mouse. The total combined MTLs in different groups of immunized mice and controls from three experiments are given at the time when the MTL was 5 (A) and 12 (B) mm in the controls.
FIG. 5. In vivo growth of RMA in different groups of immunized and control mice after inoculation with an s.c. inoculum of 53103
cells per mouse. MTLs in mice immunized with irradiated RMA cells, polyomavirus, LT 578–585, LT 578–586, and LT 578–587 and in controls are indicated. The MTL was calculated for four mice at different time points after tumor cell inoculation.
FIG. 6. In vivo growth of RMA in different groups of immunized and control mice after inoculation with an s.c. inoculum of 53103
cells per mouse. The total combined MTLs in different groups of immunized mice and controls from three experiments are given at the time when the MTL was 14.2 mm in the controls.
on November 9, 2019 by guest
http://jvi.asm.org/
prove the immune response of the host to certain tumors. This
strategy is an improvement over those of earlier trials, in which
peptide immunization in vivo was less efficient than was
im-munization with wild-type virus at inducing the rejection of
polyomavirus tumors (26). This may have been due to the fact
that the peptides used previously were not the naturally
occur-ring ones or were the wrong size. Similar findings have been
reported earlier by others (25, 27, 32).
Even though the in vivo protective effect of certain
polyoma-virus-specific peptides against polyomavirus tumor challenge is
clear, the exact mechanisms of immune recognition and tumor
rejection are still not understood. The immune response
against polyomavirus tumors is believed to be mediated by T
cells (2, 18, 22, 33). In fact, we have previously shown that both
CD4
1and CD8
1T cells are important for the rejection of
polyomavirus tumor cells (18). So far, only two reports have
indicated the presence of polyomavirus-specific cytotoxic T
lymphocytes in mice (15, 20). However, in vitro cytotoxicity has
been demonstrated in several other systems, not only against
virus-infected cells (1, 27, 32, 34) but also against
virus-trans-formed cells (7, 13).
Antiviral and antitumor immunities may demand a wide
spectrum of immune mechanisms, but the observation that
immunity to polyomavirus protects against polyomavirus
tu-mor growth suggests a link between the two. It is also likely
that certain MHC class I-presented antigenic peptides are
shared by polyomavirus-infected cells and
polyomavirus-in-duced tumor cells. Still, there must be some differences. Both
CD4
1and CD8
1cells are necessary for polyomavirus tumor
rejection (19). However, a permanent polyomavirus infection
is eliminated in both CD4
2/2and CD8
2/2single knockout
mice, not in CD4
2/2CD8
2/2double negative mice (6).
In conclusion, in vivo protection against a polyomavirus
tu-mor cell inoculum can be obtained after immunization with a
synthetic polyomavirus LT 578–585 peptide. This sequence was
identified by amino acid analysis of naturally occurring
pep-tides on the H-2K
bMHC class I molecule on a polyomavirus
tumor cell. However, it seems that the most efficient
immuno-gen was a shorter version of the naturally occurring peptide.
These results further support the feasibility of using antiviral or
antitumor vaccines based on synthetic peptides.
ACKNOWLEDGMENTS
This work was supported by the Karolinska Institute, Swedish Med-ical Research Council project no. 13x-10832 (T.B. and H.J.), and Swedish Cancer Society project no. 1806 (T.B. and H.J.) and 1753 (T.D.).
REFERENCES
1. Aichele, P., H. Hengartner, R. M. Zinkernagel, and M. Schulz. 1990. Anti-viral cytotoxic T cell response induced by in vivo priming with a free synthetic peptide. J. Exp. Med. 171:1815–1820.
2. Allison, A. C. 1980. Immune responses to polyoma virus and polyoma virus induced tumors, p. 481–487. In G. Klein (ed.), Viral oncology. Raven Press, New York. 3. Bergman, T., M. Carlquist, and H. Jo¨rnvall.1986. Amino acid analysis by
high performance liquid chromatography of phenylthiocarbamyl derivatives, p. 45–55. In B. Wittmann-Liebold, J. Salnikow, and V. A. Erdmann (ed.), Advanced methods in protein microsequence analysis. Springer, Berlin. 4. Bergman, T., and H. Jo¨rnvall.1987. Electroblotting of individual
polypep-tides from SDS-polyacrylamide gels for direct sequence analysis. Eur. J. Biochem. 169:9–12.
5. Berke, Z., and T. Dalianis. 1993. Persistence of polyomavirus in mice in-fected as adults differs from that observed in mice inin-fected as newborns. J. Virol. 67:4369–4371.
6. Berke, Z., T. Wen, S. Jin, G. Klein, and T. Dalianis. 1995. Polyomavirus persists in CD42/282/2double knockout, but not in CD42/2or CD82/2 single knockout mice. Virology 212:268–271.
7. Bright, R. K., M. H. Shearer, and R. C. Kennedy. 1994. Immunization of Balb/c mice with recombinant simian virus 40 large tumor antigen induces antibody-dependent cell-mediated cytotoxicity against simian virus 40-trans-formed cells. J. Immunol. 153:2064–2071.
8. Dalianis, T. 1990. Studies on the polyoma virus tumour-specific transplan-tation antigen (TSTA). Adv. Cancer Res. 55:57–85.
9. Dalianis, T., G. Magnusson, Y. Ito, and G. Klein. 1982. Immunization against the polyoma virus-induced tumor-specific transplantation antigen by early region mutants of the virus. J. Virol. 43:772–777.
10. Dalianis, T., T. Ramqvist, and G. Klein. 1984. Studies on the polyoma-virus-induced tumor specific transplantation antigen (TSTA)—does middle or large T-antigen play a role? Int. J. Cancer 34:403–406.
11. Engelhard, V. H. 1994. Structure of peptides associated with MHC class I molecules. Curr. Opin. Immunol. 6:13–17.
12. Falk, K., O. Ro¨tzschke, S. Stevanovic, G. Jung, and H.-G. Rammensee.1991. Allele-specific motifs revealed by sequencing of self-peptides eluted from MHC molecules. Nature (London) 351:290–296.
13. Feltkamp, M. C. W., H. L. Smiths, M. P. M. Vierboom, R. P. Minnaar, B. M.
de Jongh, J. W. Drijfhout, J. ter Schegget, C. J. M. Melief, and W. M. Kast.
1993. Vaccination with cytotoxic T lymphocyte epitope-containing peptide protects against a tumor induced by human papillomavirus type 16 trans-formed cells. Eur. J. Immunol. 23:2242–2249.
14. Fried, M., B. E. Griffin, E. Lund, and D. L. Robberson. 1974. A study of wild type mutant and defective DNAs. Cold Spring Harbor Symp. Quant. Biol.
39:45–52.
15. Greene, M., L. Perry, E. Kinney-Thomas, and T. Benjamin. 1982. Specific thymus-derived (T) cell recognition of papova virus transformed cells. J. Immunol. 128:732–736.
16. Joyce, S., and S. G. Nathenson. 1994. Methods to study peptides associated with MHC class I molecules. Curr. Opin. Immunol. 6:24–29.
17. Lathe, R., M. P. Kieny, P. Gerlinger, P. Clertant, I. Guizani, F. Cuzin, and
P. Chambon.1987. Tumor prevention and rejection with recombinant vac-cinia. Nature (London) 326:378–380.
18. Ljunggren, G., H.-G. Ljunggren, and T. Dalianis. 1994. T cell subsets involved in immunity against polyoma virus-induced tumors. Virology 198:714–716. 19. Ljunggren, H.-G., and K. Ka¨rre.1985. Host resistance directed selectively
against H-2 deficient lymphoma variants. Analysis of the mechanism. J. Exp. Med. 162:1745–1759.
20. Lukacher, A. E., Y. Ma, J. P. Carroll, S. R. Abromson-Leman, J. C. Laning,
M. E. Dorf, and T. Benjamin.1995. Susceptibility to tumors induced by polyoma virus is conferred by an endogenous mouse mammary tumor virus superantigen. J. Exp. Med. 181:1683–1692.
21. Luthman, H., M.-G. Nilsson, and G. Magnusson. 1982. Non-contiguous segments of the polyoma genome required in cis for DNA replication. J. Mol. Biol. 161:533–550.
22. Miller, J. F. A. P., R. C. Ting, and L. W. Law. 1964. Influence of thymectomy on tumor induction by polyoma virus in C57BL mice. Proc. Soc. Exp. Biol. Med. 116:323–328.
23. Ramqvist, T., D. O. Pallas, J. DeAnda, L. A¨hrlund-Richter, G. Reinholdsson,
T. Roberts, B. S. Schaffhausen, and T. Dalianis.1988. Immunisation against the polyoma tumor specific transplantation antigen (TSTA) with polyoma T-antigens. Int. J. Cancer 42:123–128.
24. Ramqvist, T., G. Reinholdsson, M. Carlquist, T. Bergman, and T. Dalianis. 1989. A single peptide derived from the sequence common to polyoma small-and middle-T antigen induces immunity against polyoma tumors. Virology
172:359–362.
25. Reddehase, M. J., J. B. Rothbard, and U. H. Koszinowski. 1989. A pen-tapeptide as minimal antigenic determinant for MHC class I-restricted T lymphocytes. Nature (London) 337:651–653.
26. Reinholdsson-Ljunggren, G., T. Ramqvist, L. A¨hrlund-Richter, and T. Dalianis.1992. Immunization against polyoma tumors with synthetic pep-tides derived from the sequences of middle- and large-T antigens. Int. J. Cancer 50:142–146.
27. Ro¨tzschke, O., K. Falk, K. Deres, H. Schild, M. Norda, J. Metzger, G. Jung, and H.-G. Rammensee.1990. Isolation and analysis of naturally processed viral peptides as recognised by cytotoxic T cells. Nature (London) 348:252–254. 28. Siegel, S., and N. J. Castellan. 1988. Nonparametric statistics for the
behav-ioral sciences, 2nd ed., p. 128. McGraw-Hill, Singapore.
29. Sjo¨gren, H. O.1964. Studies on specific transplantation resistance to poly-omavirus-induced tumors. I. Transplantation resistance induced by polyoma-virus infection. J. Natl. Cancer Inst. 32:361–374.
30. Soeda, E., J. R. Arrand, N. Smolar, J. E. Walsh, and B. E. Griffin. 1980. Coding potential and regulatory signals of the polyoma virus genome. Nature (London) 283:445–453.
31. Townsend, A. R. M., J. Rothbard, F. M. Gotch, G. Bahadur, D. Wraith, and
A. J. McMichael.1986. The epitopes of influenza nucleoprotein recognized by cytotoxic T lymphocytes can be defined with short synthetic peptides. Cell
44:959–968.
32. Van Bleek, G. M., and S. G. Nathenson. 1990. Isolation of an endogenously processed immunodominant viral peptide from the class I H-2Kbmolecule.
Nature (London) 348:213–216.
33. Vandeputte, M. 1968. The effects of heterologous antilymphocytic serum on the oncogenic activity of polyoma virus. Life Sci. 7:855–865.
34. Zhou, X., L. Berg, U. M. Abdel Motal, and M. Jondal. 1992. In vivo primary induction of virus-specific CTL by immunisation with 9-mer synthetic pep-tides. J. Immunol. Methods 153:193–200.