Vol. 0022-538X/82/091038-08$02.00/0
Copyright ©1982, AmericanSocietyforMicrobiology
Identification
of
Ecotropic
Proviral
Sequences in High-
and
Low-Ecotropic-Virus-Producing
Mouse
Strains
JANETL. MOORE1tANDHARDYW. CHAN2t*
Laboratory ofViralDiseases'andLaboratoryof Molecular Microbiology,2 National Institute of Allergy and
InfectiousDiseases, Bethesda,Maryland20205
Received 18February 1982/Accepted4 June 1982
The arrangement ofendogenous ecotropic retroviruses in selected high- and
low-ecotropic-virus-producing mouse strains was examined by Southern blot
hybridization analysis, usingan ecotropic retrovirus-specific DNA probe.
High-ecotropic-virus-producingmousestrains of the AKRfamily displayed
heterogene-ity withrespect tothe number ofcopies and the sites of insertion of endogenous ecotropic specificDNA. Thisdiversity was seen evenamong individuals ofthe same AKR subline. Contrastingly, individuals within the same
low-ecotropic-retrovirus-producing mouse strain showed no evidence of variability in their endogenous ecotropic proviral sequences. These results favored the hypothesis that germ line proviral reinsertion was responsible for the proviral sequence
heterogeneity observed inhigh-ecotropic-virus-producing mousestrains.
The endogenous type C viralgenomes in the
mouse are acomplexgroupof DNAs,
represent-ing a multicopy gene family. Some of the
se-quences are potentially infectious genomes of ecotropic andxenotropicmurineleukemia
virus-es (MuLVs), but the majority appear to be defective. The inducible, infectious viruses have been extensively characterized with regard to
theirbiological properties (19), but biochemical characterization of the endogenous chromo-somal sequenceshasbeen limited. Studies done with liquid hybridization between in vitro
syn-thesizedcDNAand mousecellular DNAs show
that partsof the MuLV genome are present in many copies, whereas other portions of the genome, namely, those that code for
type-spe-cific properties, are presentin smaller numbers
(4). Ecotropic MuLV-specific sequences fell
into threecategories with respect to copy
num-ber in the DNA of different strains of mice.
High-virus-producing mice such as AKR
showed 3 to 4copies per haploid genome,
low-virus-producingmice such as BALB/c showed 1 to 2, and ecotropic virus-negative strains
showed none. Xenotropic MuLV-specific
probesshowedlarger copy numbers (3 to 10 per
haploidgenome inall mice), but the number did not appear to correlate with the ease of
induc-tionof virus(14).
Studies involving blot hybridization
tech-tPresent address: Barbara Kopp Research Center,
Au-burn,NY13021, and Upstate Medical Center, State Universi-tyof New York, Syracuse, NY 13210.
tPresentaddress:SyntexCorp.,PaloAlto, CA 94304.
niques have confirmed and extended these
re-sults (1-3,22, 23). Probesprepared from whole viralgenomesshowallmouseDNAstocontain many copies of viral sequences, whereas more
type-specific probes prepared by absorption or
molecular cloning of subgenomic portions ofthe
virus have shown the env genedeterminants of ecotropic virusestobepresent in much smaller numbers (1, 3, 4, 17). In addition to providing
more precise quantitation, the blot analyses
al-lowed the identification of single-copy
provi-ruses in terms of the restriction sites in their
flanking sequences, the organization of the se-quences,and characterization of thediversity of the endogenous sequences (7).
We previously have presented preliminary results ofour studies ofendogenous ecotropic
MuLV sequences based on blot hybridization with a molecularly cloned env gene segment fromAKR ecotropic virus (1). In thisreport, we
extend these studiesto furthercharacterize the
differences inendogenous ecotropic proviruses
within and among various strains of laboratory
mice.
MATERIALS AND METHODS
Mice.AKR/N, AKR/J, BALB/c, C3H/He, C57BR,
CBA,C57BL/6, SEA/J, and DBA mice were obtained from the National Institutes of Health and the Jackson
Laboratory. The AKR/Boy mice were a kind gift of E.A.Boyse. The AKV congenic NFS mice were bred inthe laboratory of W. Rowe.
DNA extraction and purification. High-molecular-weight mouse liver DNAs were extracted and purified fromfresh tissue as previously described (1).
Restriction endonuclease digestion and gel
electropho-1038
on November 10, 2019 by guest
http://jvi.asm.org/
ENDOGENOUS ECOTROPIC MuLV SEQUENCES 1039
resis. All restriction enzymes used in these experi-ments wereobtained from New England Biolabs and used as specified by the supplier. Completeness of digestion of cellular DNAs was monitored by adding lambda DNA to a portion of the restriction digestion mixture and evaluating its cleavage by gel electropho-resis. Multiple fragments of digested cellular DNA were resolved byelectrophoresis in 0.7 or
1.0%1
neu-tralagarose gels as described previously (11, 12) and transferred to nitrocellulose sheets as outlined bySouthern (21).
Preparation of hybridization probe. The construction of the recombinant plasmid clone consisting of the 400-base-pair ecotropic-specific env segment from a cloned infectious AKR ecotropic MuLV (15) has been
previously described (1). The DNA was labeled by
nicktranslation (16) and hadspecific activities of 8 x
1 to12x 106cpm/gg.
Nuclekacidhybridization. Preincubation of nitrocel-lulose sheets was carried out asdescribedpreviously (5), and the filters were hybridized in sealed plastic
bags containing10 ml of 3x SSC, pH 7.4(Ix SSC is 0.15 M NaCl plus 0.015 M sodium citrate), lOx
Denhardt solution (1x Denhardt is 0.02% each of bovine serum albumin, Ficoll, and
polyvinylpyrrol-idone)(5),0.1%sodiumdodecyl sulfate, sheared salm-onsperm DNA (50g/ml),andlabeled DNA (3 x 106 cpm persheet) at 60°C for 16 to 36 h. Nitrocellulose membranes were washed as previouslydescribed(13), airdried, andexposed topreflashedKodakXR-2 film.
RESULTS
Because of the extensive polynucleotide se-quencehomology among ecotropic, xenotropic, and dualtropic MuLVs, we have prepared
enve-lope-specificprobes (1) that specifically
hybrid-ize to different classes of proviral DNA and
permitanexamination ofendogenous retroviral sequences. In aprevious report that described the construction and potential usesforan
eco-tropic envelope probe,wedemonstratedthatthe
copy number of endogenous ecotropic
provi-rusesand the arrangement of cellular sequences that flank integrated ecotropic proviral DNA
could bereadily ascertained (1). Suchanalyses
weregreatly facilitated by thediscoverythat the
restrictionenzyme mapsofecotropicproviruses
were virtually identical (3, 22; unpublished
data). Thus,the strategy outlinedpreviouslyof
using restriction endonucleases which cleave
ecotropicproviralDNAsasingletime(eitherto
the 5' or the 3' side of the 400-base-pair
eco-tropic envelope-specific segment) in
combina-tion withspecific MuLVprobestoevaluate the
organization of endogenous ecotropic
provi-ruses in AKR orBALB/c mice could be
safely
extended to all mouse strains because of thenearly completeconservation of restriction en-zyme sites.Intheexperiments tobedescribed,
mouseliver DNAs weredigestedwithHindIII,
Sacl,orXbaI,which cleavethe
ecotropic
provi-ral DNA3.1,3.7,and 7.8kilobases(kb)from the
5' terminus,
respectively.
Since HindIII andSacl cut the provirus to the 5' side of the
ecotropic env-specific segment (5.7 to 5.1 kb from the 3' terminus), fragments that hybridize to theprobe will contain viral sequences includ-ing the 3' long terminal repeat (LTR) and flank-ingcellular sequences (Fig. 1). Conversely, cel-lular DNA fragments which hybridize to the ecotropic envelope-specific probe after XbaI digestion will contain 7.8
kb.
ofproviral DNA including the 5' LTR and adjacent cellular DNA sequences. Mouse liver DNAs were also hybrid-ized to the ecotropic MuLV-specific probe after digestion with EcoRI which does not cleave within the ecotropic provirus. Although diges-tion with EcoRI should also indicate the number of ecotropic proviruses present in a given cellu-lar DNA preparation, the very high-molecucellu-lar- high-molecular-weight cleavage productsgenerated were often difficult to resolve in the agarose gel systems employed.Characterization of the ecotropic proviral DNAspresent in three colonies of AKR mice. The number of ecotropic proviruses in AKRJN,
AKR/J, and AKRIBoy mice was initially deter-mined by digesting preparations of mouse liver DNAwith HindIII (Fig. 2A). Cleavage of intact endogenous ecotropic provirus with HindIII generated a restriction fragment at least 5.7 kb in size (Fig. 1) that contained the 3' LTR and adjacent flanking cellular sequences. Digestion of mouse liver DNA prepared from an NFS mouse congenic for the Akv-1 locus with HindlIl yielded asingle 5.8-kbfragmentwhich
hybridized to the ecotropic env-specific probe (Fig. 2A, lane 9). Liver DNAs obtained from three different AKR/N (Fig.2A, lanes 1, 2, and
4),four differentAKR/J(Fig. 2A, lanes5 to8),
andtwo different AKR/Boy (Fig. 2A, lanes 10
and 11) mice andcleavedwith HindIII all
con-tained the 5.8-kb DNA fragment characteristic
oftheAkv-1 locus. Hindllldigestion of cellular
DNApreparedfromanNFSmousecongenicfor
the Akv-2 locus generated a 7.0-kb fragment
whichannealedtotheecotropicenvDNAprobe (Fig. 2A, lane 3). This same 7.0-kb cleavage productwas present in the three AKR/N (Fig.
2A, lanes 1, 2, and 4) DNAs and in the two
AKR/Boy (Fig. 2A, lanes 10 and
11)
DNAs.However, a band comigrating with the 7.0-kb
HindlIl Akv-2fragmentwasabsent inoneAKR/ Nmousesampleandwasdetected inonlytwoof
fourAKR/J DNA
samples
(Fig. 2A, lanes 5 to8).ForthetwoAKR/Boy (Fig. 2A,lanes 10 and
11)DNA, oneHindIII
fragment
appeared
tobeshorter than 5.7 kb. This fragment probably represented apartially deleted
proviral
genome (i.e., PstIdigestionsof thetwoDNAsgeneratedaweakreactiveband ofabout 3.3kb,in addition to an8.2-kb bandwhich istypicalofacomplete
ecotropic viral sequence
[data
notshown]). VOL. 43,1982on November 10, 2019 by guest
http://jvi.asm.org/
Hind1II
-~~Si.7kb -i< .1 kb
78kb
[image:3.501.104.392.83.137.2]Xba I
FIG. 1. Restriction enzymecleavagemap ofintegratedAKRecotropicMuLV. Thestraight line together with theclosed boxes(representingthe LTRsequence)is indicative of thelengthof the AKRecotropicviral genome. The openbox represents theportionof theenvgene thatwas subcloned inpBR322togenerate theecotropic MuLV-specificprobe (1). indicates theadjacentmousecellularsequence.Thehorizontalarrowsrepresentthe distance (inkb)between the enzymecleavagesites and the viral-cellular DNAjunctions.
Saclcan also be usedinconjunctionwith the ecotropic env probe to evaluate endogenous
ecotropic proviruses and cellular sequences which flank the 3' LTR sequence (Fig. 1). To
furtherinvestigatethepossibleabsenceor
alter-ation of theAkv-2locus insomeAKR/Janimals suggestedfrom the HindIIIdigestions (Fig. 2A),
mouseliver DNAswere cleaved with Sacland annealedto theecotropicenvprobeafter
trans-fertonitrocellular membranes. None of thefou.
AKR/J DNAs examined (Fig. 2B, lanes 3 to 6)
containedthe 6.2-kb SacIfragment
characteris-tic of the Akv-2 (Fig. 2B, lane 9) locus. On the otherhand,twoAKR/Boy(Fig. 2B, lanes 1 and 2) and one AKR/N (Fig. 2B, lane 7) DNAs containedthis DNA segment. In agreementwith the results obtained afterHindIII cleavage,
rep-resentatives of all three AKR colonies contained
1 2 3 4 5 6 7 8 9 10
23.5- W_e OW;}44
9.6-6.6-.
4.4-A
1 2 3 4 5 6 7 8 9
*11e < 1FV .I
:
:.
s.P -A2-FIG. 2. EcotropicMuLV proviralDNAsequencesinindividualAKRmouse.(A)AKR/N (no. 7) (lane1),
AKR/N(no. 78) (lane2),NFS.Akv-2(lane 3), AKRIN(no. 79)(lane4),AKR/J(no. 66) (lane5),AKR/J(no.67)
(lane 6),AKR/J(no.42)(lane 7),AKRIN (no. 55) (lane 8),NFS.Akv-1 (lane9),AKR/Boy(no. 49)(lane10), and
AKR/Boy (no. 50) (lane 11)mouse liverDNAswerecleaved withHindIIIand analyzed by hybridization to
labeled ecotropic env MuLV DNA probe as described in the text. Arrows indicate the DNA fragments
correspondingtoAkv-land Akv-2genesthatarecharacteristic ofthe analogousgeneinNFS congenic mice(see
lanes 3and 9). The sizes(in kb) of HindlIl-cleavedXDNAfragmentsareindicatedatthe left.(B)AKR/Boy(no.
49) (lane 1),AKR/Boy(no. 50) (lane2), AKR/J(no. 66) (lane 3),AKR/J(no. 67) (lane 4), AKR/J(no.42) (lane5), AKR/J (no.55) (lane6),AKR/N(no. 7)(lane7), NFS.Akv-1 (lane8), and NFS.Akv-2 (lane9) DNAsweretreated exactly thesame asin (A)exceptfor digestionwithSacl.
-*-.A--1
J. VIROL.
1040 MOORE AND CHAN
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.501.125.368.315.586.2]ENDOGENOUS ECOTROPIC MuLV SEQUENCES 1041
the8.8-kbSaclfragmentcharacteristic of
Akv-1
(Fig. 2B).
Liver DNAs from AKR mice were digested withXbaI and hybridized tothe ecotropic env
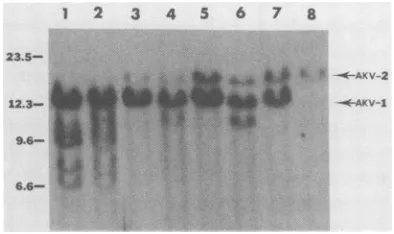
probe to evaluate the organization of cellular sequences flanking the 5' LTR sequence. One
AKR/N (Fig. 3, lane 7), four AKR/J (Fig. 3,
lanes3 and 6), and twoAKR/Boy (Fig. 3, lanes 1
and2) DNA preparationscontainedthe 12.5-kb
XbaI fragment associated with the
Akv-1
locus.The 16-kb XbaI cleavage product characteristic
ofAkv-2 (Fig. 3, lane 8) was present in AKR/N
DNA (Fig.3, lane 7), clearlyabsent in twoAKR/ BoyDNAs (Fig. 3, lanes 1 and 2), and detected
intwooffourAKR/J DNAs(Fig. 3, lanes 3 to 6).
Digestion ofmouse liver DNAs with EcoRI,
whichdoes not cleave withinthe ecotropic pro-virus (23), followed by hybridization to the
eco-tropic envelope DNA probe, was the third
ap-proach used to examine the organization of
endogenous ecotropic proviral DNA in AKR
mice. AKR/N (Fig. 4, lanes 5 and 6), AKRIJ
(Fig. 4, lanes 3 and 4), and AKR/Boy DNAs
contained the 32-kbAkv-1EcoRIfragment. Con-versely, the 25-kb EcoRI fragment associated
with the Akv-2 locus (Fig. 4, lane 2) was only
detected inAKR/N (Fig. 4, lanes 5 and 6) DNA
preparations; itwas not seenin samplesof AKR/ J(Fig. 4, lanes 3 and 4)orAKR/Boy (Fig.4,lane
1) liver DNAs.
Ananalysis of proviral DNAfragments char-acteristic of the Akv-1 and Akv-2 loci andpresent
in liver DNAs from three AKR colonies is
summarized in Table 1. Mice from all three
coloniesinvariablycontained the Akv-1locus,as
1 2 3 4 5 6 7 8
23.S-"l 'A ---rAKV-2
12.3- v vAKf-1
9.6-FIG. 3. Hybridizationofendogenous MuLV-relat-ed DNA sequences to an ecotropic MuLV-specific
DNA probe. AKRIBoy (no. 50) (lane 1), AKR/Boy (no. 49)(lane 2),AKR/J (no. 55) (lane3),AKR/J(no.
42) (lane 4), AKR/J(no. 67)(lane5), AKR/J(no. 66) (lane 6),AKRIN(no.7)(lane 7),andNFS.Akv-2 (lane 8) DNAswerecleaved withXbaIandanalyzed byblot hybridization as described in the text. The arrows correspond to ecotropic-specific DNA fragments
found inNFS.Akv-1 and NFS.Akv-2 congenic mice.
Thesizes(in kb)ofHindIII-cleavedA DNAfragments areindicatedattheleft.
1 2 3 4 5 6
--*'-AKV-2
23.S;- * v ; ;
12.3- i _ _
9.6-FIG. 4. Reactivity of endogenous ecotropic provi-ral sequence with an ecotropic MuLV-specific DNA probe. AKR/Boy (no. 49) (lane 1), NFS.Akv-2 (lane2), AKR/J (no. 67) (lane 3), AKR/J (no. 66) (lane 4), AKR/ N(no. 7) (lane 5), and AKR/N (no. 78) (lane 6) DNAs were digested withEcoRI. Southemblot hybridization was performed as described in the text.
judged by thepresence of characteristicHindIII, Sacl, XbaI, and EcoRI fragments. The three
AKR/N animals examined also contained the
Akv-2 locus. On the other hand, AKR/J and
AKR/Boy animalsexhibited an inconsistent pat-tern with respect to the presence of authentic
Akv-2 cleavage products. For example, four
AKR/J liverDNA preparations did not contain
the Sacl or EcoRI fragment characteristic of
Akv-2,yetthreeof the four contained fragments
that comigrated with HindlIl orXbaI cleavage
products associated with the Akv-2 locus.
One possible explanation forthe inconsisten-cyis that the 16-kbXbaIfragments observed in
thetwo AKR/J animalswere in fact of slightly different molecular weights than the Akv-2 se-quence. This is a likely interpretation since
molecularweight resolution byagarosegel
elec-trophoresisis relativelypoor in this size range. Athirdlocus(ecotropicEnvlocus-3,Table1) containingsequencesreactivewith theecotropic
envelope probe wasconsistently detected in all
three AKR mouse colonies. This endogenous
proviral DNA segment was associated with
characteristic HindIII (22.5 kb; Fig. 2A), SacI
(5.3 kb; Fig. 2B), XbaI (12.3 kb; Fig. 3), and
EcoRI (13 kb; Fig. 4) fragments most readily
seen inAKR/N DNA which contains onlytwo
other(Akv-1 andAkv-2)reactive loci. A second
consistentfeature of thisanalysis wasthe exis-tenceofareactive6.0-kbHindlIl fragmentin all
AKR/J andAKRIBoyDNAs(Fig. 2A,lanes5 to
8, 10, and 11). It should benoted that the data
shown inFig. 2-4arecompatiblewith the pres-ence of3, 4or 5, and 7 copies ofendogenous ecotropic proviruses in AKR/N, ADR/J, and
AKR/Boymice, respectively. In allcases, PstI
digestionofAKRmouseliver DNAgenerateda
single 8.2-kb cleavage product which reacted
with theecotropic envprobe (datanotshown).
Evaluation of theendogenous ecotropic provi-VOL.43, 1982
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.501.273.448.81.183.2] [image:4.501.56.253.456.573.2]1042 MOORE AND CHAN
I II +++++++ ++
+++ ++++ ++
+++ ++++ ++
+ ++ ++
+ + +I + + + +
+ tl
+ +1 +1 + + +
I I +
.+ I++ I++I ++
.+ ++ ++ + +
.+ +++++++ ++
++++ ++
.+ ++ ++ + ++ + +
-4
0-I+
C D + +
LXC
.0
4z _~+ +
re 4 +
c-*
.0
be ++
++0
C) +
N.0
++
ta00 +
ce
P".0
S.4 ++
lao +
0is
a- 1
._
U
2
0
0e
2
4) cU
._ ._
z
0o
4)
s-J
0_
I-0 4)
0D
4)
0n
4)
0
2 2
J-F,
z
4)
U,
0 2
4)
C--,
z-0
J
U,
< 4)0Z
S CU
4)
2e_ 0
C
e.a C |
4)
t+
0
V).0
0 2 2 V)
on November 10, 2019 by guest
http://jvi.asm.org/
ENDOGENOUS ECOTROPIC MuLV SEQUENCES
ral DNA present in mouse strains expressing
ecotropic MuLVs at a low frequency. The ap-proach outlined in Fig. 1 and used to examine
AKRmouse strains was alsoemployedtostudy
ecotropic proviruses in low-virus-expressing mice. Incontrasttothepatterns observed with
AKR animals, most low-virus-expressing mice
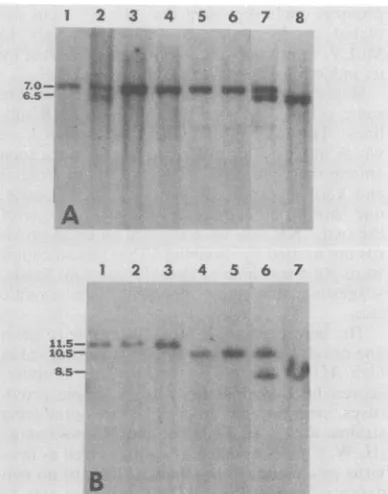
contained only a single ecotropic env reactive DNA segment. As shown in Fig. 5A, the
eco-tropic envelope probe hybridized to a 7.0-kb
HindIII fragment present in restricted C3H/He
(lane 1), C57BR (lane 2), CBA (lane 3), C57BL
(lane 4), BALB/c (lanes 5 and 6), and SEA/J
(lane 7) DNAs. DBA mouse liver DNA
con-tained a single 6.5-kb HindIII ecotropic env
reactive(Fig.5A,lane 8). C57BR and SEA mice were theonlymembersofthis group containing
asecondcopy ofendogenous ecotropic proviral DNA after digestion with HindIII. A similar
simple blot hybridization patternwas observed
aftercleavageoflow-ecotropic-virus-expressing
mouseDNA withXbaI(Fig.SB).Inthis experi-ment, however, the ecotropic provirus present in C57 andBALB/c micecould bedistinguished; theecotropicenv probehybridizedto an11.5-kb XbaI fragment from C57BR (lane 1), C57BL/6 (lane 2), and C57BL/10 (lane 3) DNAs and to a
10.5-kb fragment from BALB/c (lanes 4 and 5) DNAs. As was the case after HindIIIcleavage, XbaIdigestion of SEA (lane6)DNA generated
tworeactive fragments;onecomigratedwith the
8.5-kb XbaI fragment found in the DBA (lane7) mouse,and the othercomigrated with the single endogenous ecotropicprovirus bandpresent in
BALB/canimals.
1 23 4 56 78
A
2 3 4 5 6 7
11.5- -*
8.5-.*:.;. 0
am1. _f
B
FIG. 5. Ecotropic MuLVsequences in inbred mice with lowincidence ofleukemia. The experiments were
performed in the same manner as described in the legends to Fig. 2-4. (A) HindlIl digestion of DNAs fromC3H/He (lane 1), C57BR (lane 2), CBA (lane 3), C57BL (lane 4),BALB/c (lanes 5 and 6),SEA/J(lane 7), and DBA (lane 8) mice. (B) XbaI-cleaved DNAs from C57BR (lane 1), C57BL (no. 6) (lane 2), C57BL (no. 10)(lane 3),BALB/c (lanes 4 and 5), SEA (lane 6), and DBA (lane 7) mice.
DISCUSSION
The most striking observation in the studies
presentedhereis the extentofheterogeneity in the number and location of endogenous
eco-tropic MuLV proviruses among different AKR
mice,notonlyamongdifferentsublinesbut also
among individuals of a single colony. These resultsareconsistentwiththosepreviously
pub-lishedby otherinvestigators (8, 17, 18, 25). Datafrom ourlaboratory showed that AKR/N mice
generally contained three ecotropic
proviruses,
AKR/J
generally containedfour,
andAKR/Boy
contained aboutseven.AllAKRmicecontained
a provirus
corresponding
to that in the Akv-1congenic mice, but onlythe AKR/N mice
con-tainedaprovirus with all the characteristics of the Akv-2 locus, as was
pointed
outpreviously
(20).
The question of whether AKR/J mice carry
Akv-2, meaning the chromosome 16 locus de-fined by the NFS congenics, is not clear from these studies. Some AKR/J mice showed
eco-tropic
env-specific
probe-reactive
HindIII or XbaI orboth restrictionfragments
comigrating
with those of Akv-2, but otherAKR/J mice did
not.Inthose DNAs with both HindIII andXbaI fragments like thoseofAkv-2, it is notpossible
to say if they are from the same or different
proviruses,becauseof themultiplicityof bands. Since thetwofragmentsare notuniformly pres-ent together, it seems most likely that they
representdifferentprovirusesand that their sim-ilarity in size to Akv-2-containing fragments is
fortuitous. Further Mendelian and biochemical studies arein progress to clarifythis
point.
In addition to Akv-1 and Akv-2
proviruses,
AKR/N DNAsconsistently showed athird eco-tropic-reactive provirus; its flanking sequence parameters with the various enzymescould be reliably inferred sincethe only otheruniformly
presentbandwasAkv-1.This thirdprovirus
was present in all mice of all three AKRcolonies,
andmustrepresentanoninducibleprovirus
that,
like Akv-l, was present in AKR before theseparation ofAKR/J and AKR/N in the 1940s.
(Bothlineswerederivedfrom AKR stockatthe
Rockefeller Institute in the
1940s,
the N linebeing carried initially by
Lloyd
Law[6].)
ThisVOL.43, 1982 1043
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.501.264.458.80.327.2]1044 MOORE AND CHAN
provirus couldrepresentthe "Akv-3" locus de-tected by its ability to induce antibody to MuLV,orpossiblytheAkvlp locus,detectedby its induction ofgs andGixantigens (9, 10).
Heterogeneity between individuals of the same colony wasseenwith all three AKR sub-lines. This was most evident with AKRIJ, in whichatleast four different patterns were seen among the eight mice tested with both HindIII
and XbaIdigestion. Ofthe three AKR/N mice, one showed an additional provirus, and one of thetwoAKR/Boy mice showedanextra provi-rusnotcarried by its sibling. These extracopies
wereoften less intense than theconstantbands, suggesting that they represent heterozygotic loci.
The heterogeneity ismost likely due togerm lineproviral reinsertions previouslyobservedin high Akv-1 congenic mice (20). This is
under-scored by the multiplicity of ecotropic provi-ruses seen in the other high-virus-producing strains, e.g., C3H/Fg, C58, and M. molossinus
(H. W. Chan, unpublished data),aswellas low-virus-producing strains showing little or no evi-dence of reinsertions. In fact, we were able to show that in related low-ecotropic-virus-produc-ingmousestrains, despite separation of perhaps 50years or300 generations, the identical provi-ralintegrations arestill retained.
Ifgermline proviral reinsertions indeedoccur in high-virus-producing mouse strains, the fre-quency ofreinsertion orthe ability to maintain the inserted proviral sequences must vary
among individual strains. For instance, AKR/N mice generally showed three ecotropic provi-ruses,whereasAKR/Boymice had aboutseven.
Thus, other hostgenesarelikely tobe involved in the control ofgermline proviral reinsertions.
Althoughgermline proviral reinsertion is one
possibleexplanation forourfinding, ourresults
can also be seen as evidence that retroviral
genomes undergo frequent genetic
rearrange-mentsuchastranslocation and amplification. In
many regards, MuLV proviral DNA is
remark-ably similar to the structure of bacterial and eucaryotic transposable elements(24). Whether
endogenous viral genomes can indeed
translo-cate to new genomic loci via transposition is difficulttoevaluate empirically. Suffice ittosay,
however, that such postulated ability to
trans-pose must have been lost among
low-virus-producingmouse strains.
ACKNOWLEDGMENTS
We thank MalcolmA.Martin andWallaceP. Rowe fortheir
interest andsupportof this work. LITERATURECITED
1. Chan,H. W., T. Bryan, J.L. Moore, S.P.Staal,W.P.
Rowe,andM. A. Martin. 1980. Identificationof ecotropic
proviralsequencesin inbredmousestrainswithacloned
subgenomic DNA fragment. Proc. Natl. Acad. Sci. U.S.A. 77:5779-5783.
2. Chattopadhyay, S. K., M. R. Lander, S. Gupta, E. Rands, andD. R. Lowy. 1981. Origin of mink cytopathic focus-forming (MCF) viruses: comparison with ecotropic and xenotropic murine leukemia virus genomes. Virology 113:465-483.
3. Chattopadhyay, S.K., M. R. Lander, E. Rands, and D. R. Lowy. 1980. Structure ofendogenous murine leukemia virus DNA in mouse genomes. Proc. Natl. Acad. Sci. U.S.A.77:5774-5778.
4. Chattopadhyay, S. K., D. R. Lowy, N. M. Teich, A. S. Levine, and W. P. Rowe. 1974. Qualitative and quantita-tive studies of AKR-type murine leukemia virus se-quences in mouse DNA. Cold Spring Harbor Symp. Quant. Biol.39:1085-1101.
5. Denhardt, D. T. 1966. A member filtertechnique for the detection of complementary DNA. Biochem. Biophys. Res. Commun. 23:641-646.
6. Furth, J. 1978. The creation of the AKR strain whose DNAcontains the genome ofaleukemia virus. In H. C. Morse III(ed.), Origins of inbred mice. Academic Press, Inc., New York.
7. Groner, B.,and N.E.Hynes.1980. Number and location of mouse mammary tumor virusproviral DNA in mouse DNAof normal tissue andof mammarytumors.J. Virol. 33:1013-1025.
8. Herr, W.,and W.Gilbert. 1982. Germ-line MuLV reinte-grations in AKR/J mice. Nature(London)296:865-867. 9. IhIe, J. N., and D. R. Joseph. 1979. Serological and
virological analysis of NIH (NIH XAKR)mice: evidence for three AKR murine leukemia virus loci. Virology 87:287-297.
10. Ikeda, H.,W. P.Rowe,E. A.Boyse, E.Stockert,H. Sato, and S. Jacobs. 1976. Relationship of infectious murine leukemia virus andvirus-relatedantigensingenetic cross-esbetween AKR and Fv-1 compatible strains C57L. J. Exp. Med. 143:32-46.
11. Israel, M. A., H. W. Chan, W. P. Rowe,and M. A. Martin. 1979.Molecularcloning of polyoma virus DNA in Escherichia coli:plasmidvectorsystem.Science 203:883-887.
12. Israel,M.A.,D. F.Vanderryn, D. F.Meltzer, and M. A. Martin. 1980. Characterization of polyoma viral DNA sequencesin polyoma induced hamster tumor cell lines. J.
Biol.Chem. 255:3798-3805.
13. Jeffreys, A. J., and R. A. Flavell. 1977. Aphysical map of the DNA regions flanking rabbit B globin gene. Cell 12:429-439.
14. Lowy, D. R., S. K.Chattopadhyay, N. M. Teich, W. P. Row, and A. S. Levine. 1974. AKRmurine leukemia virus genome:frequencyof sequences in DNA of high-, low-, andnonvirus yielding mouse strains. Proc. Natl. Acad. Sci.U.S.A. 71:3555-3559.
15. Lowy, D. R., E. Rands, S. K. Chattopadhyay, C. F. Garron, and G. L. Hager. 1980. Molecular cloning of infectious integrated murine leukemia virus DNA from infected mouse cells. Proc. NatI. Acad. Sci. U.S.A. 77:614-618.
16. Maniatis, T., A.Jeffreys, and D. G. Kleld. 1975. Nucleo-tide sequence of therightward operator of phage X. Proc. Natl. Acad. Sci. U.S.A.72:1184-1188.
17. Quint, W., W. Quax, H. van der Putten, and A. Berns. 1981. Characterization of AKR murine leukemia virus sequences in AKR mouse substrains and structure of integrated recombinant genomes in tumor tissues. J. Virol. 39:1-10.
18. Quint, W., H. van der Putten, F.Janssen, and A. Berns. 1982.Mobility of endogenous ecotropicmurineleukemia viral genomes within mouse chromosomal DNA and
integrationof a mink cell focus-forming virus-type recom-binant provirus in the germ line. J. Virol. 41:901-908. 19. Rowe, W. P. 1978. Leukemia virus genomes in the
chro-mosomal DNA of the mouse. Harvey Lect. 71:173-192. 20. Rowe, W. P., and C. A. Kojak. 1980. Germ-line
on November 10, 2019 by guest
http://jvi.asm.org/
ENDOGENOUS ECOTROPIC MuLV SEQUENCES 1045
tions of AKV-1 congenic mice. Proc. Natl. Acad. Sci. U.S.A. 77:4871-4874.
21. Southern, E. M. 1975. Detection ofspecific sequences amongDNAfragments separated by gel electrophoresis. J. Mol. Biol. 98:503-517.
22. Steffen, D., S. Bird, W. P. Rowe, and R. A. Weinberg. 1979. Identification of DNA fragments carrying ecotropic proviruses of AKR mice. Proc. Natl. Acad. Sci. U.S.A.
76:4554-4558.
23. Steffen, D., and R. A. Weinberg. 1978. The integrated
genomeof murine leukemia virus. Cell 15:1003-1010. 24. Temin,H. M. 1980. Origin of retroviruses from cellular
moveablegenetic elements. Cell 21:599-600.
25. Yoshimura, F. K., and M. Breda. 1981. Lack of AKR ecotropic provirus amplification in AKR leukemic
thy-muses.J.Virol. 39:808-815. VOL. 43, 1982
on November 10, 2019 by guest
http://jvi.asm.org/