Copyright 0 1975 AmericanSociety forMicrobiology Printedin U.S.A.
Intermediate
in
Adenovirus Type
2
Replication
GEORGE D. PEARSON
DepartmentofBiochemistryandBiophysics, Oregon State University,Corvallis, Oregon97331 Receivedforpublication27September 1974
Replicating chromosomes, called intermediateDNA,have been extracted from theadenovirus replication complex. Comparedtomaturemolecules, intermedi-ate DNA had a greater buoyant density in CsCl gradients and ethidium
bromide-cesium chloride gradients. Digestion of intermediate DNA with Si endonuclease, butnotwithRNase, abolished thedifference indensities. These properties suggest that replicating molecules contain extensive regions of
parental single strands. Although intermediate DNA sedimented faster than
marker viralDNA in neutral sucrose gradients, singlestrands longer thanunit
lengthcouldnotbe detectedafter alkalinedenaturation. Integralsizeclassesof nascent chains in intermediate DNA suggest a relationship between units of
replication and thenucleoproteinstructureof the viruschromosome.Adenovirus
DNA was replicated at a rate of 0.7 x 106 daltons/min. Although newly
synthesized molecules had the same sedimentation coefficient and buoyant densityasmaturechromosomes, theystill containedsingle-strandinterruptions.
Complete joiningofdaughterstrandsrequired an additional 15 to 20min.
Adenoviruses replicate in the nuclei of in-fectedcells.Viral DNA mostlikelyexists inthe
nucleus as a nucleoprotein complex. Pearson
and Hanawalt (13) separated nascent viral
DNA, as a replication complex, from finished molecules. The kinetics of labeling indicated
that the adenovirus complexwas an
intermedi-ate in replication. Studies with three com-plementationgroups of type 31adenovirus
tem-perature-sensitive (ts)mutants, alldefectivein
the initiation of viral DNA synthesis, showed that the replication complex does not form at nonpermissive temperatures(19, 27). Yamashita and Green (34) recently isolated from adeno-virus-infected cells a nuclear membrane com-plex which contains two virus-specified DNA
binding proteins (30, 31), as well as endo-nuclease and DNA polymerase activities (T. Yamashita, M. Arens, and M. Green, personal communication).Nevertheless,theintranuclear
site of viral replication hasnot yetbeen iden-tified. Electron microscopeautoradiography
in-dicates that viral DNA synthesis occurs in the nucleoplasm, notinassociation with the nuclear envelope (20, 21).
There is as yet little information about the composition ofthe adenovirus complex. I show in this paper that the complex contains repli-cating adenovirus chromosomes which differ in physical properties from mature molecules. These resultsconfirmandextend earlier studies onadenovirusreplication. In addition, an
anal-ysis of nascent chains in replicating molecules reveals sizeclasses thatareintegralmultiplesof a unit 1,750nucleotides long
('ho
ofan adeno-virus strand). A possible relationship betweenreplicationunits and the nucleoprotein organi-zation of the adenovirus chromosome is dis-cussed (J. Corden, H. M. Engelking, and G.
Pearson, manuscriptinpreparation).Moreover,
evidence for a maturation step in adenovirus replication isprovided.
(A preliminary report of this work was pre-sented at the Fourth Tumor Virus Meeting at ColdSpringHarbor, N.Y., inAugust 1972.)
MATERIALS AND METHODS
Coil culture and synchronization. HeLa S,cells were grown in suspension culture using medium F-13 (Grand Island Biological Co.) supplemented with 7% fetal calf serum. Cells were synchronized with respect to DNA synthesis by two exposures for 20 h to 2 mM thymidine separated bya period of 12 h (13). Often cellular DNA was uniformly labeled with
["4C]thymidine
(3 pM, 15 sCi/4smol) during this period.Adenovirus infection. Inocula for all experiments consisted oftype 2 adenovirus purified in CsCl density gradients according to Doerfler (5). Cells were in-fected with 104particles/cell as previously described
(13).
Isolation ofthe replication complex. The isola-tionofthe adenovirus replication complex has been described in detail (13). In brief, nuclei from infected cells weredigestedwithPronase and sodium dodecyl sulfate. After shearing, the lysate was layered on a 17
on November 10, 2019 by guest
http://jvi.asm.org/
sucrose shelf gradient at 2 C. The detergent crystal-lizes in the cold. The gradient was centrifuged at 25,000rpm for 135 min at 0Cin theSW27 rotor. The replication complex collects as a turbid detergent band on the 60% sucrose shelf.
DNAextraction.Cells(107/ml)weredigested with 1 mg ofPronase/ml (previously incubated for 1 h at 37C at 5mg/ml), 0.5% sodium dodecyl sulfate, and 0.01 M EDTA for 1 h at 37C. DNA was extracted with an equal volume ofchloroform-isoamyl alcohol (24:1, vol/vol) by blending in a Vortex mixer for 1 minat room temperature.Thephaseswereseparated by centrifugation at 12,000 x g for 10 min, and the. aqueousphasewasretained.
CsCldensity gradient centrifugation. Gradients with an initialdensityof 1.710 were constructed by dissolving 7.90 g of CsCl in 6.5 g of liquid. To minimize the loss of DNA by adsorption to walls, gradients contained 0.1% sarkosyl and were cen-trifugedinpolyallomer tubes.Centrifugationwasfor 36 h at 37,000 rpm at 20 C in the 5OTi angle rotor. Fractions (0.3ml) were collectedvolumetricallyfrom a pin hole in the bottom of the tube by pumping mineral oil in at the top. CsCl gradients containing 300ug of ethidium bromide per ml were constructed by dissolving6.70g ofCsCl in 7.25 g ofliquidtogive aninitialdensityof 1.550.
Sucrose gradient centrifugation. Sucrose gradi-ents(5 to20%, wt/vol; 30 ml)were formed inSW27 tubes. Gradientswerecentrifugedat25,000rpm and 20C.The duration of the runs and the compositions ofthe individualgradientsaredetailed in the appro-priatefigure legends. Fractions (1 ml) were collected volumetrically asdescribed above.
Radioactivity determinations. 8H and "4C were analyzedasdescribedpreviously (13).Isotope overlap calculations were computed on a Hewlett-Packard 9821A calculator.
Materials. Pronase (free ofnucleases), ethidium bromide,andAquacideIIwerepurchasedfrom Calbi-ochem; pancreatic RNase (five times crystallized) and thymidine came from Schwarz/Mann; and [methyl- 14C
Ithymidine
was from New England Nu-clear. The single-strand-specific S1 endonuclease fromAspergillusoryzae waspurifiedfromEnzopharm powder(EnzymeDevelopmentCorp.) andassayedas described by Sutton(28).RESULTS
Specificlabelingof adenovirus DNA. HeLa cellsweresynchronizedbytwosuccessive expo-sures to 2mMthymidineand infectedwithtype 2 adenovirus at the beginning of S phase.
Infected cells proceeded normally through S phase, but did not enter mitosisnor initiate a subsequent round of cellular DNA synthesis (13, 21). Since cellular replication had ceased
by 10 h after infection, adenovirus molecules could belabeled exclusivelybetween 13and 16 h after infection, the period of maximal viral DNAreplication. Allexperimentsreportedhere were started 13 h afterinfection.
Demonstration ofintermediate DNA in the replication complex. Figure 1A shows the dis-tribution ofa6-minpulseof [3H
]thymidine
ina sucrose shelfgradient. About 60 to 70% of the pulse label (newlyreplicated DNA) collectedon the shelfas adetergent band (13). Lessthan5% of the [4C]cellular
DNA sedimented onto the shelf; the bulk remained at the top of the gradient. The detergent band (labeled b) and the combined top fractions (marked c) were centrifuged separately to equilibrium in CsCl density gradients. Nascent viral DNA isolated from thedetergent band had a buoyant density of 1.725 (Fig. 1B), greater than the density of marker adenovirus DNA (1.715; Fig. 1E).[14C]HeLa
DNA banded at 1.700. Denaturedadenovirus DNA has adensity of 1.730(25). Ten percentof the 3H label appeared as a shoulder tailing through the position of marker viral DNA. On the other hand, Fig. 1C shows that [3H
]DNA
from thetop fractionsbanded atthe expected density of 1.715. No pulse label was found in the region of cellular DNA in either gradient.Newly replicated molecules exhibiting in-creased buoyant density can be extracted di-rectly from infected cells after digestion with Pronase and detergent (Fig. 1D). DNA labeled for 6 min with [3H]thymidine was broadly distributed with a peak at 1.725 and a promi-nentshoulderat1.715. Viral DNA labeledfor 1 h bandedatthepositionofmarker DNA witha shoulderat 1.725 (notshown). No radioactivity was detected at 1.725 when a 6-min pulsewas chased for 90 min with 10-4 M unlabeled
thymidine (notshown).
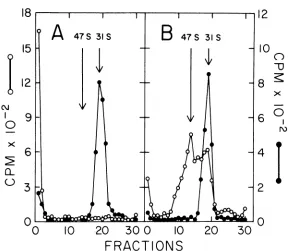
Isolation of intermediate DNA in cesium chloride-ethidium bromide density gradi-ents. Infected cells were labeled with [3H ]-thymidine for 6 min. After extraction, the DNA was banded in a CsCl density gradient containing ethidium bromide(Fig. 2A). A major peakwasobserved at adensityof 1.605.Aminor peak at 1.585 coincided with the band of [14C]HeLa DNA.Doerfleret al. (6, 7) have also shown that during infection some ofthe intra-cellular adenovirus DNA can be isolated as a dense bandinpropidium diiodide-cesium chlo-ride gradients. The dense band (indicated by the bar in Fig. 2A) was further analyzed by velocity sedimentation in a neutral sucrose gradient(Fig. 2B). Over85%of theradioactivity sedimented faster than31S, therateformarker adenovirusDNA. Someofthepulse sedimented faster than 60S (fractions marked as a). The rest of the pulse sedimented from 31 to 50S (labeled b). For comparison, the calculated
on November 10, 2019 by guest
http://jvi.asm.org/
REPLICATINGADENOVIRUSDNA 19
24
20
16 ro
° 12 x
08
4
0,
_ 0
l(
A, O
0
W
L
l l
C)
u
I
rX
0 5 10 15 -o 10 20 30
FRACTIONS
FRACTIONSFIG. 1. Identification ofintermediateDNA in theadenovirusreplication complex.Synchronized, infected cells were concentrated to 1.6 x 101 cells/ml and labeledfor 6 minwith [3H]thymidine (10 iCi/ml at 20 Ci/mmol). Symbols: 0, [14CJHeLa DNA; 0, [sHjadenovirus DNA. (A) Cells(8 x 106) were converted to a nuclearlysateandcentrifugedon asucroseshelfgradientaspreviouslydescribed (13). Sedimentation isfrom righttoleftinallfigures. Theshelffraction (b)and thefractionsatthe topofthegradient (c) were extracted withchloroform-isoamylalcohol andconcentratedby dialysis againstsolidAquacide II. (B) Centrifugation of theshelf fraction(b)in a CsCIdensity gradientwithaninitial p = 1.710.Densityincreasesfrom righttoleftin allfigures. (C) CsCI density gradient analysis of fractionsatthe topofthesucroseshelfgradient (c). (D) Cells(4 x 10') described above were extracted with chloroform-isoamyl alcohol after digestion with Pronase and sodium dodecyl sulfate. Thewhole-cell extract wascentrifuged toequilibrium ina CsCIgradient. (E) ["1C]-HeLaDNAfromuninfectedcellswasmixed with [3HJadenovirusDNA extractedfrom purified viral particles andcentrifugedtoequilibriuminaCsCIgradient.
sedimentation coefficients for linear dimer and trimer molecules are 40 and 45S, respectively
(22). Fractions a and b both had a buoyant
density of1.725in CsCl (notshown).
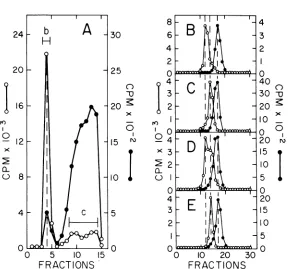
Figure3A shows that after labeling intracel-lular adenovirus DNA for15min twoprominent peaks appearedatdensities of1.605 and 1.585.
[14C
]HeLa DNA again coincided with the lighter band (not shown). When centrifuged in CsCl gradients after removing the ethidiumbromide, the densepeak (designated b)banded at adensityof 1.725 (Fig. 3B) and the light peak (marked c) bandedat 1.715(Fig.3C). Figure3D shows that twopeakswith theexpected densi-ties appeared when b and c were mixed. The dense band markede, intermediate DNA, was broadly distributed in a neutral sucrose gradi-ent (Fig. 3E). Only about one-halfoftheDNA sedimented faster than 31S, although some of the molecules still sedimented as fast as 60S.
The differencebetween gradient profiles in Fig. 2B and 3E most likely can be attributed to shearing duetotheextrapurification step.The small shoulder at 26S corresponds to duplex molecules one-half the molecular weight of adenovirus DNA. The light CsCl band, pooled asf, sedimentedat31S withaslight shoulderat 26S (Fig. 3F). Figure 3G demonstrates that a fast sedimenting peakat 42S appeared when a mixture of e and f were centrifuged together. Intermediate DNA sedimenting between 35 to 40S(fractions markedh inFig. 3E) bandedat a densityof1.725 (Fig. 3H). Intermediated DNA sedimentingnear31S(fractions labeledi inFig.
3E) bandedat aslightlylowerdensity,spanning the region between 1.715 and 1.725 (Fig. 3I). Mature31SDNA(peak jinFig. 3F), previously isolated at a density of 1.715, remained at the samedensity (Fig. 3J).
Kinetics of labeling intermediate DNA. VOL.16,1975
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.498.96.388.64.333.2]0 10 20 30
FRACT
ION S
3
0~75
a_ 2
0-0 10 20 30
FRACTIONS
FIG. 2. Isolationofintermediate DNA by equilibrium centrifugation incesium chloride-ethidium bromide density gradients.Synchronized, infected cellswereconcentratedto1.7x 10"cells/ml and labeled for 6 min with [3H]thymidine (10 MCi/ml at6.7Ci/mmol). (A) Whole cells wereextractedas described inFig.1 and
centrifuged to equilibrium in a CsCl density gradient (initial p = 1.550) containing 300 isg of ethidium
bromide/ml.Symbols: 0, [14C]HeLaDNA; 0, ['H]adenovirusDNA. Thefractions indicated by the barwere
pooled, extractedwith isopropanoltoremovethe ethidiumbromide,anddialyzedtoremovetheisopropanol and CsCl. (B) Velocity sedimentation ofintermediate DNA in a neutral sucrose gradient. The gradient
contained 1.0 MNaCl, 0.005 MEDTA, and 0.01 M Tris, pH 8. Centrifugationwasfor 7 hat25,000rpm.The arrowindicates theposition ofmarkerviralDNA inaseparategradient (not shown). Fractions labeledaand b
werepooled separately and further analyzedonCsCIdensity gradients (not shown).
Replicating molecules should be labeled prefer-entially during intervals shorter than the time to synthesize completed daughter molecules. Figure 4A displays the distribution in cesium chloride-ethidium bromide gradients of DNA labeled with [3H]thymidine for various
pe-riods of time. During the first minute, label appeared exclusively in intermediate DNA with
a density of 1.605. The shortest pulsewas 20s
(not shown). Incorporation into DNA banding at 1.585required 3to5min.The total incorpo-ration in both peaks, normalized for the
recov-ery of [14C]HeLa DNA in each gradient, is presented in Fig. 4Bas afunction of the labeling
time. Radioactivity waslocated predominantly in intermediate DNA during the first 10 min. After a short delay (extrapolated back to 3 min), label accumulated linearly in finished molecules. Figure 4B also shows that about 50% of thepulse label could be chased from interme-diate DNA intocompleted viral molecules after adding a 1,000-fold excessofunlabeled thymi-dine at10 min. The chasewaseffective within
10min (Fig. 4B, inset). The kinetics of labeling
and the chase confirmaprecursor-product
rela-tionship. Adenovirus DNA is replicated in 17 min, the time required to label intermediate DNA andcompleted molecules equally (i.e., the intersection inFig. 4B).
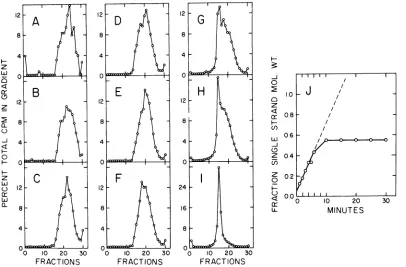
IntermediateDNAandcompletedviral mole-culesfrom each time point were also analyzed by velocity sedimentation in alkaline sucrose
gradients (Fig. 5A through I). In agreement with Horwitz (9), no singlestrands longerthan unit length (34S) were everdetected. After1 min of labeling, strands of intermediate DNA (Fig. 5A) sedimented in the region oflOS (1,750 nucleo-tides) to 14S (3,500 nucleotides) with faster sedimenting shoulders at 19S (9,000 nucleo-tides) and 26S (17,500 nucleotides). Chains as
shortas400nucleotidescould also be detected. Vlak et al. (33) and E. Winnacker (personal communication) have also found 9 to 11S strands asintermediates in adenovirus
replica-tion. SincethelOSchaincorrespondsto'hoofan
adenovirusstrand, Ihave defined this unitas a
"faceful" (i.e., equivalenttooneof the 20 faces of the icosahedral virus particle). The signifi-6
rK)
40
0
2
0
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.498.127.409.71.293.2]REPLICATINGADENOVIRUS DNA 21 canceoffacefulswillbe discussedbelow. After 2
(Fig. 5B) or 3 min (Fig.5C)oflabeling, strands ofintermediate DNA sedimented at 14, 18, 23, and 26S, which correspond to 2, 4, 8, and 10 facefuls. Longer strands of 30 to 32S (16 to 18 facefuls) appearby4 (Fig. 5D) and 5 min (Fig. 5E). The size of nascent chains reached an
equilibrium distribution of26S within 10 min (Fig. 5F). Unit-length single strands were also present atthis time. Further experiments to size nascent strands by agarose gel electrophoresis are in progress.
At all times, completed molecules sedi-mented primarily as unit-length single strands.
Some newly finished molecules evidently have transient single-strand interruptions. Finished
molecules isolatedafter 10 (Fig. 5G) or15min
(Fig. 5H) oflabelingcontain shorter fragments
sedimenting at18, 23, 26, and 31S (i.e., 1/4, 1/3, 1/2, and 3/4 fractional lengths). After a chase withunlabeled thymidine for 80 min (Fig. 5I), all strandswere unitlength. Thisalsoexcludes
the possiblity that ethidium bromide intro-duced strand breaks during the separation of intermediate DNA from finished molecules.
The center of mass of each sucrose gradient profile of intermediate DNA was plotted in Fig. 5J as a function of labeling time. Interestingly, the graph extrapolated to a molecular weight of 5.8 x 10' (or 1,750 nucleotides) for an infinitely shortpulse. The initial rate of chain elongation was 0.7 x 106 daltons/min. The time to com-plete unit-length single strands was calculated to be 16 min, in close agreement with the estimate in Fig. 4B.
Properties of intermediate DNA. Either
single-strandedDNAorRNA could increase the
buoyant densityofintermediate DNA. Petter-sson (14) showed that the density difference
between replicating and mature adenovirus DNA was eliminated after digestion with the single-strand-specific nuclease from Neuro-spora crassa, but not after digestion with
RNase. The following experiments confirm
0 l0 20 30
FRACTIONS
0-U
olorlI ws~
0 10 20 30
FRACTIONS
0
x
l0 20 30
FRACTIONS FiG. 3. Characterization of intermediate DNA and completed viral DNA. A sample from the culture describedinFig.2wasremovedafter labeling for15min. (A) Equilibrium centrifugation ofextracted DNA in an ethidium bromide-cesium chloride density gradient. Fractions labeled b and c werepooled separately,
extracted withisopropanol,anddialyzed. ["4C]HeLaDNA(not shown)co-banded with thelightband(c). (B) Rebanding of fractionband["4CJHeLaDNA(not shown)inaneutral CsCIdensity gradient (initialp= 1.710).
Fractions labeled e were pooled and dialyzed. The vertical dashed lines represent, from left to right: intermediateDNA, adenovirus DNA, and HeLaDNA.(C) Rebanding of fractioncand["4ClHeLa (not shown)
inaneutralCsCI density gradient. Fractions labeled fwerepooledanddialyzed. (D) Rebanding ofamixtureof
b and c in a neutral CsCI density gradient. Closed circles represent marker ["4C]HeLa DNA. (E) Neutral sucrosegradient velocity sedimentation of fraction e. Sucrosegradients (containing 1.0 MNaCI, 0.005M
EDTA, and 0.01 M Tris, pH 8) werecentrifuged for7hat25,000rpm.Fractions labeledhandiwerepooled
separately and dialyzed. (F) Neutralsucrosegradient velocitysedimentationof fraction f. The fractionlabeledj wasretained anddialyzed. (G) Neutralsucrosegradient velocity sedimentation ofamixtureofeandf. The arrowmarks theposition of 42S. (H) Rebanding of fraction h inaneutral CsCIdensity gradient. Symbols: *,
[CJC]HeLaDNA. The verticaldashed lines represent,from lefttoright:intermediateDNA, adenovirus DNA, andHeLa DNA.(1) Rebanding offraction i and["4CJHeLa DNA(not shown) inaneutralCsCIdensity gradient.
(J) Rebanding of fraction j and ["C JHeLa DNA (not shown)inaneutral CsCIdensity gradient.
0 X
cL
re)
0
0-(-)I VOL.16,1975
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.498.47.438.320.497.2]8
r'l)
0
X 6
A-q
0
X
CL
0 10
20
30
020
40
60
80
100
FRACTIONS
MINUTES
FIG. 4. Kinetics of labeling intermediate DNA and finished adenovirus DNA. Infected cells were concen-trated to 4 x 106 cells/ml and labeled with [9H]thymidine (10 tiCi/ml at 10 Ci/mmol). At 10 min the incorporationof[3H]thymidine was quenched by adding 10-4 M unlabeled thymidine. Samples of 4x 106 cells wereremoved at the indicated times, extracted, and centrifugedto equilibrium incesium chloride-ethidium bromidegradients. (A) Profilesofcesium chloride-ethidium bromide densitygradients. The peakof[14C]HeLa DNA (not shown) is indicated by the arrow. The [9H]thymidine incorporated into adenovirus DNA was normalized to the recovery ofHeLa DNA in eachgradient (45,000 counts/min of 14C). Symbols: 0, 1 min; 0, 3 min; A, 5 min; 0, 10 min. (B) Radioactivity in the peak fractions ofthe cesium chloride-ethidium bromide gradientswas normalizedfor recovery ofHeLa DNA in eachgradientandplottedasafunction of the time oflabeling. The chase with 10-4 Mthymidine wasstarted 10minafteraddingthe [3H]thymidine. Symbols: 0, 3Hin intermediateDNA; 0, 3H in completedadenovirus DNA. (Insert) Total incorporation of.
['H]thymidinein eachgradientas afunction oftime. Thedashed line indicates the initialrateofincorporation.
Thearrowindicates thebeginning ofthe chase.
those observations. Purified intermediateDNA,
labeled for 5 min with ['H ]thymidine, was mixed with [14C
]adenovirus
DNA anddigestedwith pancreatic RNase or the
single-strand-specific Sl endonuclease from A. oryzae(1, 28). Intermediate DNA had a buoyant density of 1.725 (Fig. 6A). After treatment with Si endo-nuclease (Fig. 6B), intermediate DNA banded at the density of marker DNA, although less than 1% of 3H and '4C labelswere solubilized. Under the same conditions, more than 95% of heat-denatured viral DNA could be digested. Figure6C shows thatRNase digestion inbuffer containing 0.2 M NaCl had no effect on the density of intermediate DNA. RNase
hydro-lyzed betterthan 98% of
['H
]HeLarRNAunder the same conditions. These experimentsstrongly suggest that the increased buoyant
densityof intermediate DNA is dueto
substan-tial single strands of parental DNA. In this regard, replicating adenovirus chromosomes ap-pear in electron micrographs as branched and unbranched linear molecules with extensive single-stranded regions (8, 23, 26, 29). Thus, adenovirus might replicate by displacement synthesis, a mechanism proposed for mitochon-drial DNAreplication (3, 16, 17).
In spite of the above conclusions, hydrogen-bondedRNA mayplayaroleinmaintainingthe structure ofintermediate DNA.Figure6D dem-onstrates that RNase disrupted intermediate DNA in 0.02 M NaCl, conditionswhere RNase attacks RNA in RNA-DNA hybrids. In three experiments an average of 9% (range, 6 to 12%) ofthe 3H labelbandedattheposition of marker DNA. The rest of the 3H activity floatedatthe meniscus ofthe CsCl gradient (the total recov-ery of 3H was better than 70%). An
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.498.75.460.56.338.2]REPLICATING ADENOVIRUSDNA 23 tion is that nascent strands in intermediate
DNA arereleasedas alow density complex. The
complex must be insensitive to detergents, Pronase, and concentrated salt solutions. Pet-tersson (14) also reported that RNase digestion in buffer lacking salt reduces the buoyant density of replicating adenovirus DNA, but the magnitude of the change was less than reported here. Furthermore, intermediate DNA can be labeledwith radioactive uridine (7; G. D. Pear-son, unpublished data).
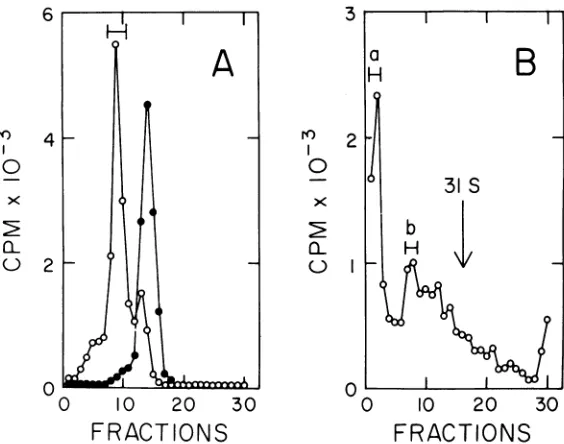
Thesedimentation rate of intermediate DNA is markedly altered by changes in ionic strength. Figure 7A demonstrates that
interme-diateDNA formed a fast sedimenting aggregate (greaterthan100S) when the ionic strength was loweredto 0.01 M NaCl. At salt concentrations greater than 0.1 M NaCl, intermediate DNA sedimented as a broad zone with peaks at 47 and 31S (Fig. 7B; compare with Fig. 2B and
l0 20 30
FRACTIONS
0 10 20 30
FRACTIONS
3E).Thesedimentationrateof marker adenovi-rus DNA was not affected within the range of ionic strengths used in these experiments. It is difficult to attribute this behavior to single-stranded regions in intermediate DNA. Single
DNA chainsassume an extended conformation in low salt due to electrostatic repulsion by negatively charged phosphates. The sedimenta-tion rate decreases as a consequence (22). Al-though the basis for the aggregation is not known, it has been used to separate finished molecules from replicating molecules to con-struct a replication map of adenovirus (G. D.
Pearson, manuscript in preparation) by the method of Danna and Nathans (4).
DISCUSSION
Nascent DNA contained in the adenovirus replication complex, called intermediate DNA, wasshown to differphysicallyfrommatureviral
0 l0 20 30
FRACTIONS
H
-J
0 C)
z
4
ci)
(5
zI
z
0
H)
0
4-.0
//
0.8 /
06 /
04 02
0.0 11I I11
0 10 20
MINUTES
30
FIG. 5. Alkaline sucrosegradient velocitysedimentationofintermediate DNA and completedadenovirus
DNA isolatedfrom the cesium chloride-ethidium bromidegradientsdescribed inFig.4. Peakfractionswere
pooled separately,extracted withisopropanol, anddialyzed.Thefractionswerethen denaturedby adjustingto
0.2 M NaOH andcentrifuged for7 h at25,000rpmin alkalinesucrosegradients (containing1.0 MNa+).At
least2,000counts/min of3Hwerein eachgradient. (A)IntermediateDNA,1-min pulse; (B)intermediateDNA,
2-min pulse; (C) intermediateDNA, 3-min pulse; (D)intermediateDNA,4-min pulse; (E)intermediateDNA, 5-min pulse; (F)intermediateDNA,10-min pulse; (G) completedviralDNA, 10-min pulse; (H) completedviral
DNA,10-min pulse plus 5-min chase; (I) completedviralDNA, 10-min pulse plus 80-min chase; (J) plot ofthe
centerofmassofintermediateDNA in eachgradientas afunction ofthelabelingtime. The centerofmass was
calculatedfrom the relationship .,f1CIC,, wheref, = fraction number and Cl = countsperminute in that
fraction. Thiswasconverted tosingle-stranded molecularweight (22). z
111 ES
cr
J 4
(5
z
a-4 I
0
z Lu
LuJ
a-16,1975
I..
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.498.41.447.294.561.2]16
AC
8
184
o
I
0
01
o
cL8~~~~~~~~~~~~~~
N00
10
20
30
0
10
20
3Q0
FRACTIONS
FI G. 6. Digestion ofintermediate DNA withSlendonuclease orribonuclease. ['H]intermediateDNA was purifiedasdescribed inFig.2,ForSl endonuclease,the reactioncontainedper 0.3 ml: 25Ml of['HJintermediate
DNA; 10 MI of ['4C]adenovirusDNA; 7.5Mgofheat-denaturedcalf thymus DNA;25 MIofenzymein 0.07 M NaCl,0.03M sodiumcitrate, 0.001 MZnCI2, pH4.5. The mixture wasincubated for1h at 50C, stoppedby adding0.5%sarkosyl, andcentrifugedin CsCIgradients containing0.5%sarkosyl. RNasewasfirst heatedto 80C for 10 min. Reactions in high salt contained per 1.0 ml: 25
Al
of [(H]intermediate DNA, 10Al of['4C]adenovirusDNA,and 100Mgof pancreaticRNase in 0.2MNaCI,0.01MEDTA,0.01 MTris, pH8. AfterI h at 37C,thereactions werestopped by adding0.5%sarkosylandcentrifugedasabove.Reactionsinlow salt wereidentical except that thebuffercontained: 0.02 MNaCI,0.01MEDTA,0.01 MTris,pH 8. Symbols:0, [3H]intermediateDNA; 0, ['4C]adenovirusDNA. (A) NoSlendonuclease(control). Thecontrols inhighand low saltwithout RNase gave similarprofiles (not shown); (B) with Slendonuclease;(C)with pancreatic RNase in 0.2 MNaCI; (D) with pancreaticRNase in 0.02 MNaCI.
molecules. Forexample, intermediate DNAhad an increased buoyant density (1.725
compared
to1.715) andagreatersedimentation coefficient (upto 100Scompared to31S). Sincetreatmentwith thesingle-strand-specificSi endonuclease,
but not with RNase, lowered thebuoyant den-sity of intermediate DNA without loss of the pulse label, a substantial fraction of the paren-tal strands must be single stranded (also see
14).Branched and unbranched linear molecules containing extensive single-stranded regions
have been visualizedinthe electronmicroscope (8, 23, 26, 29). Theseproperties agree with the properties of replicating adenovirus DNA
re-portedbyotherlaboratories(2, 8, 23-26, 29,32). An improved methodforisolating intermediate DNA has been developed. Ethidium bromide-cesium chloride density gradients provide in-creased separation between intermediate DNA (1.605) and mature viral DNA (1.585). Experi-ments reported here carefully document the identitiesof thesebands.
When labeled for 1 min or less, nascent
strands from intermediate DNA sedimented
primarilyatlOS,correspondingto achain 1,750 nucleotides long. No strands longer than an intact viral strand were ever detected. Vlak et al. (33) and E. Winnacker (personal communi-cation) have shown that9 to11Snascentchains are complementary to both viral strands throughout the entire genome. Remarkably, othernascentstrandsatthis andlonger times of labelingwereintegral multiplesof 1,750 nucleo-tides. Thisreplication unit has been defined as a faceful, since it is exactly '/2 of a finished adenovirus strand. Recentexperimentssuggest arelationship between replicationunitsand the nucleoprotein structure of the virus chromo-some. Double-stranded equivalentsofthe face-ful (i.e., 1,750 base pairs) appearas the initial product when disrupted adenovirus particles are digested with staphylococcal nuclease. Longer digestiontimesyield homogeneous frag-ments of about 150 base pairs. We postulate that a regular arrangement of core proteins protect discrete regions of viral DNA from nuclease attack. A complete model with sup-porting evidencewillbepublishedelsewhere(J.
on November 10, 2019 by guest
http://jvi.asm.org/
REPLICATING ADENOVIRUS DNA 25
18
15
I12
9
0
6
03
0'-0
10
20
30 0
10
20
30
12
10
n
u
8 Z
X
6 0
r%)
4t
4
FRACTIONS
FIG. 7. Effect of ionicstrengthonthesedimentationrateofintermediate DNA. ['H]intermediateDNAwas mixedwith ["ICfadenovirusDNA andsedimentedfor5hat25,000rpm inneutralsucrosegradients containing 0.01 MNaCI,0.Ou MEDTA,and 0.01 MTris, pH8 (A),orU.1MNaCI,0.001MEDTA,and0.01MTris, pH8 (B). Theprorfueo0 intermediate DNAingradientscontaining1.0MNaCI,0.001MEDTA,and 0.01 MTris,pH 8,resembledtheprofilein(B). Symbols: 0, ['H]intermediateDNA;*,
[(1C
adenovirus DNA.Corden, H. M. Engelking, and G. Pearson,
manuscript in preparation). The average time required for adenovirus replication was
calcu-latedtobe16 to17'min,correspondingto a rate of 0.7 x 106
daltons/min
for chain elongation.This rate compares favorably with 0.5 x 106
daltons/min for chicken embryo lethal orphan virus, an avian adenovirus (2), 0.6 x 106 dal-tons/minfor simianvirus 40(12),and 0.8 x 106
daltons/min
for the L strand ofmitochondrialDNA(3). Pearson and Hanawalt (13)previously overestimated the replication rate because
excessive shear wasused toisolate the
replica-tion complex.
Although newly finished molecules sedi-mentedat31Sand hadadensityof1.715, they
stillcontainedsingle-strand interruptions. Ma-ture chromosomes donot. Complete joining of
daughterstrandsrequiredatleastanadditional 15 to 20 min (unpublished observations). Un-joined strands were approximately 0.25, 0.5, and 0.75 fractional lengths. Interestingly, an adenovirus replication map (Pearson, manu-script in preparation), constructed by the method ofDanna and Nathans (4), locates an origin in Eco RI fragment F or in Eco RI fragment B veryneartofragmentFandatleast
one otherorigin withinthe Eco RI fragmentA in aregion definedbytheHpa I fragment F (E. Winnacker, personal communication). The sizes (15) and order (11) ofthe Eco RI
endonu-clease fragments of adenovirus DNA have been
reported. Thus, tworeplicativeorigins are posi-tioned about25%fromeither end of the chromo-some. Horwitz (10) also has demonstrated
originsinbothhalves ofthe adenovirusgenome using a similar approach. Unbranched mole-cules with manysingle-strandedgapshavebeen
visualizedbyelectronmicroscopy (8, 26, 29). It is tempting to speculate thatunjoined strands aresealed at thejunctionsbetween origins and termini.
ACKNOWLEDGMENTS
This research wassupportedbygrants(NP-67, NP-67A)
from theAmerican Cancer Society anda Biomedical Sciences
SupportGrant (5 S05RR07079-06)administered throughthe
OregonStateUniversityResearchCouncil. Part of this work
wasdone as a DernhamJunior Fellow (J-126) of the Califor-niaDivision of the AmericanCancer Society.
I thankPhilip Hanawalt, John Kiger,ErnstWinnacker, DavidDressler,and JohnWolfson for many valuable sugges-tionsand discussions. I also thank John Sussenbach for a copy of his paper priortopublication. Gail Foster provided devoted technical assistance.
VOL.16,1975
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.498.94.390.69.320.2]LITERATURE CITED
1. Ando, T. 1966. A nuclease specific forheat-denatured DNA isolated from a product of Aspergillus oryzae. Biochem. Biophys. Acta 114:158-168.
2. Bellett, A. J. D., and H. B. Younghusband. 1972. Replication of the DNA of chick embryo lethal orphan virus. J. Mol. Biol. 72:691-709.
3. Berk, A. J., and D. A. Clayton. 1974. Mechanism of mitochondrial DNA replication in mouse L-cells; asyn-chronous replication ofstrands, segregation of circular daughter molecules, aspects of topology and turnover of aninitiation sequence. J. Mol. Biol.86:801-824. 4. Danna, K. J., and D. Nathans. 1972. Bidirectional
replication of simian virus 40 DNA. Proc. Natl. Acad. Sci.U.S.A. 69:3097-3100.
5. Doerfler, W. 1969. Nonproductive infection of baby hamster kidney cells(BHK21) with adenovirus type 12. Virology 38:587-606.
6. Doerfler, W., U. Lundholm, and M. Hirsch-Kauffmann. 1972. Intracellular forms of adenovirus deoxyribonu-cleic acid. I. Evidence for a deoxyribonudeoxyribonu-cleic acid protein complex inbaby hamster kidney cells infected with adenovirus type 12. J. Virol. 9:297-308. 7. Doerfler, W., U. Lundholm, U. Rensing, and L. Philipson.
1973.Intracellular forms of adenovirus DNA. II. Isola-tion indye-buoyant density gradients of a DNA-RNA
complexfromKB cells infected with adenovirustype
2.J. Virol. 12:793-807.
8. Ellens, D. J., J. S. Sussenbach, and H. S. Jansz. 1974. Studies on the mechanism of replication of adenovirus DNA. III. Electron microscopy of replicating DNA. Virology 61:427-442.
9. Horwitz, M. S. 1971. Intermediates in the synthesis of type 2 adenovirus deoxyribonucleic acid. J. Virol. 8:675-683.
10. Horwitz, M. S. 1974. Location of the origin of DNA replication in adenovirus type 2. J. Virol. 13:1046-1054. 11. Mulder, C., P. A. Sharp, H. Delius, and U. Pettersson. 1974. Specific fragmentation of DNA of adenovirus serotypes 3, 5, 7, and 12, and adeno-simian virus 40
hybrid virus Ad2+ND1 by restriction endonuclease
R-EcoRI. J. Virol. 14:68-77.
12. Nathans, D., and K.J. Danna. 1972.Specific origin in
SV40 DNA replication. Nature (London) New Biol.
236:200-202.
13. Pearson, G. D., and P. C.Hanawalt. 1971. Isolation of,
DNAreplicationcomplexesfromuninfected and
ade-novirus-infectedHeLa cells. J. Mol. Biol. 62:65-80.
14. Pettersson, U. 1973. Some unusualpropertiesof
replicat-ing adenovirus type2DNA. J. Mol.Biol. 81:521-527. 15. Pettersson,U.,C.Mulder,H.Delius,and P.Sharp.1973.
Cleavage of adenovirus type2 DNA into six unique
fragments by endonuclease R-RI. Proc. Natl. Acad.
Sci.U.S.A. 70:200-204.
16. Robberson,D.L.,and D. A.Clayton.1972.Replicationof
mitochondrial DNAin mouseLcells and their
thymi-dine kinasederivatives:displacementreplication on a
covalently-closedcirculartemplate.Proc.Natl. Acad.
Sci. U.S.A. 69:3810-3814.
17. Robberson, D. L., and D. A. Clayton. 1973. Pulse-labeled componentsinthereplication of mitochondrial deoxy-ribonucleic acid. J. Biol.Chem. 248:4512-4514. 18. Robinson, A. J., H. B. Younghusband, and A. J. D.
Bellett. 1973. A circular DNA-protein complex from adenoviruses. Virology 56:54-69.
19. Shiroki, K., and H. Shimojo. 1974.Analysis of adenovirus 12 temperature-sensitive mutants defective in viral DNAreplication. Virology 61:474-485.
20. Shiroki, K., H.Shimojo, and K. Yamaguchi. 1974. The viral DNAreplication complex of adenovirus 12. Virol-ogy 60:192-199.
21. Simmons, T., P. Heywood, and L. D. Hodge. 1974.
Intranuclear site of replication of adenovirus DNA. J. Mol. Biol. 89:423-433.
22. Studier, F. W. 1965. Sedimentation studies of the size andshape of DNA. J. Mol. Biol. 11:373-390. 23. Sussenbach, J. S., D. J. Ellens, and H. S. Jansz. 1973.
Studies on the mechanism of replication of adenovirus DNA.J.Virol. 12:1131-1138.
24. Sussenbach, J.S., and P. C.van der Vliet. 1972. Viral
DNA synthesis in isolated nuclei from adenovirus-infected KB cells.FEBS Lett. 21:7-10.
25. Sussenbach,J.S.,and P. C.vander Vliet. 1973.Studies
onthe mechanism ofreplication of adenovirus DNA.I. The effect ofhydroxyurea. Virology 54:299-303. 26. Sussenbach, J. S., P. C. van der Vliet, D. J. Ellens, and
H.S. Jansz. 1972. Linear intermediatesinthe
replica-tion ofadenovirus DNA. Nature (London) New Biol. 239:47-49.
27. Suzuki, E., and H. Shimojo. 1974. Temperature-sensitive formation of the DNAreplicationcomplexin adenovi-rus31-infected cells. J. Virol.13:538-540.
28. Sutton, W. D. 1971. A crude nuclease preparation suita-ble for useinDNA reassociationexperiments.Biochim. Biophys. Acta 240:522-531.
29. vanderEb, A. J. 1973. Intermediates in type 5 adenovirus DNAreplication. Virology 51:11-23.
30. vanderVliet,P.C.,and A. J. Levine. 1972.DNA-binding
proteins specific for cells infectedby adenovirus.
Na-ture(London)New Biol.246:170-174.
31. vanderVliet, P.C.,A. J. Levine, M. J. Ensinger, and H.
S.Ginsberg.1975.Thermolabile DNAbindingproteins
from cells infected with a temperature-sensitive
mu-tantofadenovirusdefectiveinviral DNAsynthesis.J.
Virol.15:348-354.
32. van derVliet, P. C., and J. S. Sussenbach. 1972. The mechanism of adenovirus-DNA synthesis in isolated nuclei. Eur. J. Biochem. 30:584-592.
33. Vlak, J. M., T. H. Rozijn, and J. S. Sussenbach. 1975. Studiesonthe mechanismofreplicationofadenovirus DNA. IV. Discontinuous DNAchain propagation. Vi-rology 63:168-175.
34. Yamashita, T., and M. Green. 1974. Adenovirus DNA
replication. I. Requirement for protein synthesis and isolation of nuclear membrane fractions containing newly synthesized viral DNA and proteins. J. Virol.
14:412-420.
on November 10, 2019 by guest
http://jvi.asm.org/
![FiG. 3.EDTA,andseparatelyFractionssucrosewasarrowdescribedanextractedRebanding(J)inintermediateb[CJC]HeLa and a Characterization of intermediate DNA and completed viral DNA](https://thumb-us.123doks.com/thumbv2/123dok_us/1571085.109699/5.498.47.438.320.497/edta-andseparatelyfractionssucrosewasarrowdescribedanextractedrebanding-inintermediateb-hela-characterization-intermediate-completed-viral.webp)
![FIG. 4.Symbols:DNAnormalizedthebromidetrated0,incorporationwerebromideThe['H]thymidine 3 Kinetics of labeling intermediate DNA and finished adenovirus DNA](https://thumb-us.123doks.com/thumbv2/123dok_us/1571085.109699/6.498.75.460.56.338/symbols-dnanormalizedthebromidetrated-incorporationwerebromidethe-thymidine-kinetics-labeling-intermediate-adenovirus.webp)