DLR Rheinpfalz, ABt. Phytomedizin, Labor f. Biotechnologischen Pflanzenschutz, Breitenweg 71, 67435 Neustadt/Wstr., Germany
What do we (need to) know about low-susceptibility of codling moth against
Cydia pomonella granulovirus (CpGV)!
Was wir über die Minderempfindlichkeit des Apfelwicklers gegenüber dem Cydia
pomonella Granulovirus (CpGV) wissen (müssen)!
Johannes A. Jehle, Samy Sayed, Britta Wahl-Ermel
Abstract
In 2005, the first codling moth (CM) populations with a reduced susceptibility to Cydia pomonella granulovirus (CpGV) products have been observed. This phenomenon might be an indicator of an emerging resistance of CM to CpGV. This paper summarizes some aspects of CM CpGV interaction and discusses potential mechanisms in the host virus interaction, which could contribute to the observed low susceptibility. In order to develop and implement successful resistance management strategies, much more knowledge about the plasticity of CM CpGV interaction and the diversity of CM populations is essential.
Keywords: Codling moth, Cydia pomonella granulovirus, CpGV, resistance
Introduction
The codling moth (CM, Cydia pomonella L) is one of the most destructive insect pests of apples, pears and walnuts. CM is distributed all over the world, where pome fruits are grown. In unmanaged orchards CM infestation can reach to > 95% and result in total loss of marketable harvest.
Cydia pomonella granulovirus (CpGV) (virus family Baculoviridae) is highly pathogenic for CM and its infection results in high mortality of early instar larvae. It can efficiently control the codling moth and reduce damage to the apples. CpGV has been registered for the control of CM in many Euro-pean countries and the US. In recent years, CpGV products are one of the most successful com-mercial baculovirus insecticides. At present, a number of CpGV products are commercialised in Europe, e.g. MADEX® (Andermatt Biocontrol, Switzerland), GRANUPOM® (Probis GmbH, Ger-many), and CARPOVIRUSINE® (Arysta, formerly Calliope, France). CpGV insecticides meet all the demands of a sustainable and environmentally benign CM control agent. Although their efficacy is comparable or even superior to chemical insecticides, they are totally innoxious to any other form of life in the apple plantation and to users and consumers. Nowadays, CpGV application and mating disruption are the two corner stones of CM control in organic apple production.
Any change of one of the crucial steps during the replication cycle might have an effect on the host virus interaction and might confer host resistance to the infecting virus.
Three major mechanisms of resistance development are conceivable:
1. Changes in the midgut cell structure (impairing entrance to midgut cells)
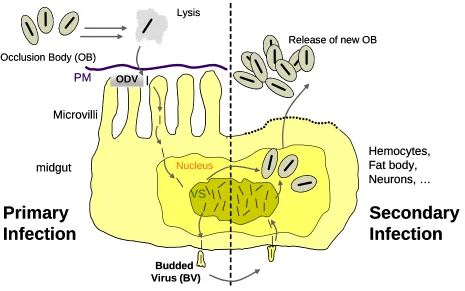
[image:2.595.72.537.227.518.2]2. Changes in the midgut cell physiology (impairing primary infections of midgut cells) 3. Changes in immune status (impairing secondary infection of other tissues)
Fig. 1: Schematic replication cycle of CpGV. Infection starts with solubilization of occlusion body in
the midgut. Occlusion derived virions initiate infection of midgut epithelium cells. Primary infection of midgut cells result in production of budded viruses. Budded viruses are transported to other tissues and initiate secondary infection. Late in infection cycle occluded form of CpGV is produced. Larva die from infection and cadavers release billions of viral occlusion bodies.
Occlusion Body (OB)
Lysis
ODV
PM
VS
Nucleus
Budded Virus (BV)
Hemocytes, Fat body, Neurons, … midgut
Release of new OB
Microvilli
Primary
Infection
Secondary
Infection
Occlusion Body (OB)
Lysis
ODV
PM
VS
Nucleus
Budded Virus (BV)
Hemocytes, Fat body, Neurons, … midgut
Release of new OB
Microvilli
Primary
Infection
Resistance of hosts against baculoviruses
There are a number of reports where differences among geographically distinct insect host popula-tions to baculoviruses were observed (Briese, 1986). In some cases it could be shown that resis-tance of host populations had a genetic factor. However, in most of theses studies resisresis-tance was provoked in the laboratory by selection pressure. In general, the levels of decreased host suscepti-bility to baculoviruses following selection have been low. Field observations of resistance are very rare and where never systematically investigated (Cory & Myers, 2003). In contrast, Fritsch et al. (2005) found field populations of CM with a extremely reduced susceptibility to CpGV.
What do we know about codling moth resistance to CpGV?
There are CM populations, which show 100-1000 times lower susceptibility to CpGV than other populations or susceptible lab strains (see contribution of Huber, this issue). In the laboratory, we have reared one CM population revealing low susceptibility to CpGV in the laboratory without sig-nificant loss of resistance for more than one year. Hence, the mode of resistance is inherited and not lost. This indicates also that the stability of resistance might be high, whereas the costs of resis-tance are low (Milks et al., 2002).
In order to test the susceptibility of different instars of CM to CpGV we have performed the following experiment. Both neonate L1 larvae of the susceptible Cp-NW and the low-susceptible Cp-BÜR were grown to different instars L1, L2, L3, L4 and L5. When they reached the respective instar, 20- 30 larvae were singly held on artificial diet contaminated with 2x105 OB/ml. The larvae were kept at 26°C at 16 h light: 8 h dark. The mortality of the larvae was scored daily for 20 days.
Table 1. Median time to death and end mortality of different instars of susceptible Cp-NW
popula-tion and low susceptible Cp-BÜR strain kept on artificial diet, which was contaminated with 2x105 OB/ml. The duration of the bioassay was 20 days, the mortality was scored daily.
Cp-NW Cp-BÜR
Instars Median time to death (days)
End mortality (%) Median time to death (days)
End mortality (%)
L1 4 100 9 67
L2 5 100 10 56
L3 5 100 -* 48
L4 7 100 -* 39
L5 9 100 -* 36
* end mortality after 20 days below 50%.
As shown in Table 1, obvious differences in the median time to death and the final mortality be-tween the susceptible Cp-NW and the low-susceptible Cp-BÜR strains were observed. The infection concentration of 2x105 OB/ml exceeds the 99% mortality for susceptible larvae of Cp-NW in all in-stars, whereas for Cp-BÜR only mortalities between 67% (L1) and 36 % (L5) were observed. It can be concluded that the resistance is not restricted to a specific instar. Further experiments are going on to identify the mechanisms and the inheritance pattern of resistance.
At the moment only a few cases of low susceptibility are reported. It has to be noted that CpGV products work in more than 95% of applications well and not every failure of control can be attrib-uted to a reduced susceptibility of the CM population. CpGV products have been proven to be highly efficient and environmentally safe. Due to the complex mode of virus action, the development of field resistance was always regarded as rather unlikely. However, the observed phenomenon of low susceptibility of local CM populations against CpGV products might be the first signs of an emerging resistance. If not controlled, the low susceptibility of CM to CpGV products as observed in local populations in Germany will bring about a threat to the successful control of CM in organic ap-ple production.
Therefore, more basic research combined with practical advice to manage this phenomenon is es-sential. The following research is necessary for better understanding this phenomenon and for de-veloping measure to manage it.
1) Analysing the distribution of this phenomenon in different growing areas, where these products are widely used.
Determining the baseline susceptibilities of different CM populations will help to identify local CM populations with low susceptibility, to get a more coherent picture about the plasticity of response of different populations, and to provide evidence about the potential dynamic of spread of this phenomenon.
2) Determining the inheritance pattern of resistance genes
Information of the genetic background of the resistance is important for detection and monitoring resistant populations as well as for development of measure to delay and to manage resistance, as it is for any other pesticide (Tabashnik, 1991).
3) Development of molecular markers of resistance alleles
As soon as molecular markers of resistance alleles are available the prevalence of these alleles can be monitored in CM populations and combined with measurements for resistance manage-ment strategies. Having molecular markers will provide the advantage that genotypic screening can be performed before reduced susceptibility is phenotypically expressed.
4) Determining the mechanisms of resistance
Understanding the CpGV resistance at molecular level will be an key in sustaining the utility of CpGV as a biopesticide. Knowledge on resistance mechanisms will also shed light on virus host interaction.
5) Isolation and development of an improved CpGV strain(s)
All CpGV products that are registered and used for CM control are based on the so-called Mexi-can (M) isolate, which was identified in 1964 from diseased insects. The identification and char-acterization of other CpGV isolates that can serve as alternatives to the currently used CpGV-M will be a promising step forward in sustaining the efficacy of CpGV biopesticides and in gaining a better understanding of CM-CpGV interaction. Such alternative isolates with differing virulence could be critical in a resistance management programme based on different CpGV strains.
6) Development of strategies to manage the resistance against CpGV
Literature Cited
Briese, D. T. (1986). Insect resistance to baculoviruses. The Biology of Baculoviruses, Vol. 2, Prac-tical Application for Insect Control. R. R. Granados and B. A. Federici. Boca Raton, CRC Press: 237-263.
Cory, J. S., Judith H. Myers, J. H. (2003). The Ecology and Evolution of Insect Baculoviruses. Ann. Rev. Ecol. Evol. System. 34: 239-272
Fritsch, E., Undorf-Spahn, K., Kienzle, J., Zebitz, C.P.W., Huber, J., 2005: Codling moth granulovi-rus: First indications of variations in the susceptibility of local codling moth populations. Nachrichtenbl. Deut. Pflanzenschutzd. 57: 29-34
Luque, T., Finch, R, et al. (2001). The complete sequence of the Cydia pomonella granulovirus ge-nome. Journal of General Virology 82: 2531-2547.
Milks, M. L., Myers, H. H.; Leptich, M. K. (2002). Costs and stability of cabbage looper resistance to a nucleopolyhedrovirus. Evolutionary Ecology 16: 369-385.
Steineke, S. (2004). Populationsdynamik des Cydia pomonella Granulovirus. Dissertation. Univer-sität Mainz.