Vol.41, No.2 JOURNALOFVIROLOGY,Feb. 1982, p. 542-556
0022-538X/82/020542-15$02.0O/0
Long
Terminal Repeat of Murine Retroviral DNAs: Sequence
Analysis,
Host-Proviral Junctions, and Preintegration Site
CHARLESVAN BEVEREN,1 ELAINE RANDS,2 SISIR K.CHATTOPADHYAY,2 DOUGLASR. LOWY,2ANDINDER M.VERMAl*
TumorVirologyLaboratory, TheSalk Institute, San Diego, California92138,1and DermatologyBranch, Laboratory of Tumor Virus Genetics, National Cancer Institute, Bethesda, Maryland202052
Received 27 July1981/Accepted25 September1981
The nucleotide sequence of the long terminal repeat (LTR) of three murine
retroviral DNAs has been determined. The data indicate that the
U5
region(sequences originating from the 5' end of the genome) of various LTRs is more
conserved than the U3 region (sequences from the 3' end of the genome). The
locationand sequence of the control elements such as the 5' cap, "TATA-like"
sequences, "CCAAT-box," and presumptive polyadenylic acid addition signal AATAAA inthe various LTRs are nearly identical. Some murine retroviral DNAs
containaduplication ofsequences within the LTR ranging in size from 58 to 100
basepairs. Avariant ofmolecularly cloned Moloney murine sarcoma virus DNA
inwhichoneofthe two LTRs integrated into the viral DNA was also analyzed. A
4-base-pair duplicationwas generated at the site of integration of LTR in the viral
DNA. The host-viraljunctionof two molecularly cloned AKR-murine leukemia
virusDNAs (clones 623 and 614) was determined. In the case of AKR-623 DNA, a
3- or4-base-pair direct repeatof cellularsequences flanking the viral DNA was
observed. However, AKR-614 DNA contained a 5-base-pair repeat of cellular
sequences. Thenucleotide sequence of the preintegration site of AKR-623 DNA
revealedthatthe cellular sequences duplicated during integration are present only
once.Finally,astrikinghomology between the sequences flanking the
preintegra-tion site and viralLTRs wasobserved.
During
thelifecycle ofretroviruses, the viralgenomic RNA is transcribed toDNA, some of
which is integrated into the host chromosome
(4). This crucial function carried out by viral
encoded reverse transcriptase is obligatory for
establishment of infection (42). Although the
mechanismofreversetranscriptionisquite
com-plex (16)and notfully understood, several forms
of double-stranded viralDNA
including linear,
circular, and supercoiled DNAs have been
re-ported in infected cells (41). Both the in
vivo-andin vitro-synthesized viral DNAs have two
typesofgenome-length molecules (2, 6,
15,
20, 32): (i) those that contain 5'-end genomic se-quences (U5) repeated at their 3' end (U3),formingastructure5' . . .U3U53' and
(ii)
thosethat, in addition to having 5' genomic RNA
sequencesrepeated attheir 3'
end,
alsocontain3'genomicRNA sequences repeated attheir5'
end, forming a structure
5'-U3U5
... U3U5-3'.However, analysis of the
integrated
viralDNA shows only structures
containing
U3U5... U3U5 sequences (21, 31). The U3U5
unit is referred to as the long terminal repeat
(LTR).
Thebiological function of theLTRisnotclearly
understood, but its structure warrants several
speculations. TheLTRcontains control elements
for boththepromotion (14a, 44) and termination
of viral RNA transcripts and may mediate or
facilitatetheintegration ofviralDNA into thehost
chromosomal DNA. Nucleotidesequence
analy-sisofLTRsfromseveralproviralDNAs(11, 19, 25, 34, 38) has shown: (i) direct duplication of
sequences at theterminiof viralDNA,i.e., LTRs;
(ii) inverted repeats at the termini of eachLTR;
and (iii) direct repeat of adjacent cellular
se-quences atboth termini ofproviralDNA. These structural attributes ofLTRs are reminiscent of
structures associated with bacterial transposons
(7, 8)TY] elements of yeasts(14),and thecopia
element inDrosophila (13).Inthis
manuscript,
wehave analyzed and compared the nucleotide
se-quences of severalLTRsfrommurine retroviral
DNA.Thehost-viraljunctionsoftwo
molecularly
cloned, integrated, infectious AKR-murine
leuke-mia virus (MLV) DNAs (24) have also been determined. We have also identifiedthe
preinte-gration site ofone oftheintegrated AKR-MLV
DNAs.
542
on November 10, 2019 by guest
http://jvi.asm.org/
MATERIALSANDMETHODS
Recombinant DNA. The construction of recombi-nant DNA clones X-AKR-MLV DNA(clones623and 614) (24), plasmid pMLV-lA containing the uninte-grated form ofMoloney MLV (Mo-MLV) DNA (3, 39), plasmid pMLV-101 containing the 5' halfof an integratedform of Mo-MLV (38), and cloneXAMSV-1 (43)containing unintegrated form ofMoloneymouse sarcoma virus (Mo-MSV) with inverted LTRs have been described. The construction of the Mo-MLV cDNA clone pMLV-201 has also been previously described (38).
To molecularlyclone from uninfected cells a DNA fragmentinwhichthe AKR-MLV DNA had integrat-ed, we used thefollowing approach. About 1 mg of cellular DNA from uninfected NIH/3T3 cells was digested with EcoRI andelectrophoresed on a0.7% agarose gel for20 h at 70 V/cm. The DNA from a portion ofthe agarosegelwastransferredonto nitro-cellulosefilters andhybridizedto aprobe containing the LTR andcellularflankingsequencesof the AKR-MLV DNA clone 623 (pAKR-LC-623), preparedas follows. The X-AKR-MLV DNA clone 623 (X-AKR-623)contains asingle HindIllsite about 2.5kilobase pairs (kbp) upstream from the 5' LTR-cellular junc-tion. TheHindIII-EcoRIfragmentofX-AKR-623 con-tainingthe entire AKR-MLV DNA and someflanking cellular sequences was subcloned in pBR322. Itwas then cleaved with KpnI, and the largest fragment, containing pBR322,2.0kbp of5'cellularsequences, 0.5kbp of5'LTR,0.1kbp of3'LTR, and1.0kbpof3' cellular sequences, was religated and used to trans-form Escherichia coli C600 (seeFig. 4A). Two other plasmidswereconstructedtocharacterize the recom-binant DNA containing the preintegration site. The plasmid pAKR-101 contained a KpnI-digested DNA fragment encompassing about 500 nucleotides of 5' LTRand 2.0kbp of adjacent cellularsequences (see Fig. 4A). Theplasmid pAKR-102 contained a KpnI-digestedDNAfragmentcontaining130nucleotides of 3' LTR andadjacent cellularsequences(seeFig. 4).In both cases, the DNA fragments were tailed with oligodeoxycytidineand annealedto oligodeoxyguano-sine-tailed, PstI-cleaved pBR322 DNA, followed by transformationashas beendescribed(9, 22).The 32P-labeled probes were made by nick translation, and specificactivitiesof3 x10'to5x107 cpm/,Lgof DNA wereusually obtained.
Afterhybridizationof the blot of NIH/3T3 DNAto nick-translatedpAKR-LC-623DNA, asingleband in the size range of 7 to 8kbpwasdetected. The 7- to 8-kbp region ofthe remainderofthe agarosegelwascut, eluted, andretested forhybridization to pAKR-LC-623 DNAand to afragment containing only cellular sequences. The size of thefragment was that predict-ed,since theX-AKR-623DNAis 16.4kbplong and the AKR-MLV DNAis about 8.8kbp. About I to 2 ,ug of thecellularDNAwas ligatedto 5 ,ug of separated arms of modified A phage Charon 4a as describedpreviously (5). Theligated material was packaged in vitro and assayed for viable phage. One positive clone was isolated after screening10,000 to20,000 plaques. The primary plaques were purified by two rounds of screeningtoobtain a pureplaque population.
Sequence determination procedures. The chemical modificationmethodofDNAsequencing as described
byMaxam and Gilbert (26) wasused.Sequence lad-ders weredisplayed on 6, 12, or 18% polyacrylamide-8 M urea gels 0.04 cm thick.
RESULTS
Comparison of sequences of LTRs. Since the LTR sequences may beinvolved in the integra-tion and transcripintegra-tion ofviral DNA, we were interested to determine whether differentmurine retroviral LTRs share some common features. Sizeand sequence heterogeneity of the LTR of murine C-type viruses have been predicted by Rands et al. (28) on the basis of restriction
endonuclease mapping of ecotropic MLV
DNAs. We haverefinedthisanalysis furtherby
determining the nucleotide sequence ofseveral
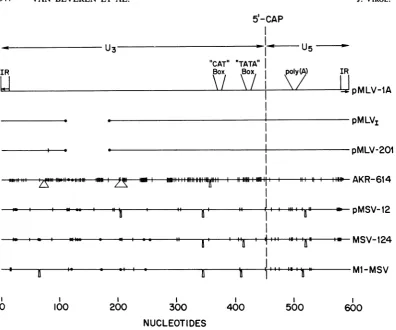
MLV LTRs. Figure 1 shows a diagrammatic
sketch of the comparison of nucleotide
se-quencesof LTRs from various murine retroviral DNAs (11, 29, 38). The complete nucleotide sequence of several of these LTRs is given below (see Fig. 6). Several generalizations can be made. (i) The size of each LTR is different,
rangingfrom 519 nucleotides (Mo-MLV ofBalb/
Mov-1 locus)to 626 for AKR-614 or AKR-623
DNA. (ii) In comparing Mo-MLV and AKR
LTRs, the U5 region isconserved, whereas the
U3region shows considerable variation.A
com-parison ofthe LTRsof various Mo-MSV isolates
andthatofMLV-1A suggeststhat cloneml
Mo-MSVismoreclosely relatedtoMo-MLVclone1
than is Mo-MSV clone 124. (iii) The 5'-cap
nucleotide islocated atapproximately thesame
position inall cases.(iv)In all cases,a
transcrip-tional control signal like theRNA PolII
initia-tion site (TATA-likebox)ispresent-25 to-31
nucleotides fromthe RNA 5'-capnucleotide. A
similarsequence atpositions+46 to +52 maybe
involved in polyadenylation ofRNA (27).
An-other control
signal
CCAAT, located aroundposition-80andimplicated intranscription (10,
17), is presentinallcases at a similar position.
(v)SomeLTRs, forinstance those of AKR-614,
pMLV-lA,andpMSV-12reportedhere andthat
ofml-MSV(11), contain an internalduplication
of sequences ranging from 58 to 100 base pairs
(bp).However, other LTRs likepMLV1-101(38)
or pMLV-201 (36), containing 3' LTR, do not
containduplication ofsequences.(vi) All
provi-ralLTRshavelost two A residues at the 5' and
3'termini,whereas the LTRsfrom unintegrated
viralDNAs retain the terminal two A residues. Itshould be noted that the loss of two A residues in the 3' LTR sequence derived from cDNA clone pMLV-201 probably occurred during S1
nuclease treatment (36). (vii) The inverted re-peatsatthetermini ofall MLV- orMSV-related
LTRs have the same sequence, namely
TGAAAGACCCC . . . GGGGTCT'TCA.
To determine whether the LTRs encode a
41,
on November 10, 2019 by guest
http://jvi.asm.org/
544 VAN BEVEREN ET AL.
5'-CAP
U3-"CAT"
Box
IR
Li
'TATA" Box
--
a s d - iSsll lz lS,.Il|- *-l.. ffi|fis ulmllSii~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~rr|nZY-nHOM /Z , 1..7, ..I -11. lm
9, , " , , | , , a
'MI
"' '"-. . _ ,,.a.I..
.
_._._...
,0
0 100 200 300
I.
*iiII I
400
poly(A)
.a
D
500
IR
I
I
!pMLV-iA
-pMLV1
-pMLV-201
AKR-614
pMSV-12
MSV-124
KA iieA4%1
600
NUCLEOTIDES
FIG. 1. Schematiccomparison of various murine retroviral LTRs. The nucleotide sequence of the LTR of pMLV-IA (unintegrated Mo-MLV [39]) is compared to those of the integrated 5' LTR from theMov-1 locus (pMLVr-101[38]),the Mo-MLV cDNA clone of the 3' LTR (pMLV-201; see text), AKR-614 (24), pMSV-12 (Fig. 2,structureII),anindependentrecombinant cloneof MSV clone 124 (29), and a recombinant clone of Mo-MSVclone ml (11).Comparisonswerecarried out using the ALIGN program (M. 0.Dayhoff, W. C. Barker, and B.C.Orcutt,personalcommunication) with a unitary matrix and a gap penalty of 3. Symbols: 0,deletions versus pMLV-lA; A, insertions; and 1, single base changes. Control signals: I.R., inverted repeat 5'-AATGAAAGACCCC-3'; CAT-box (10, 17), 5'-CCAAT-3'; TATA-box (10, 17), 5'-CAATAAA-3' for all except AKR-614(5'-CTATAAA-3');poly(A), possible polyadenylation signal (27) 5'-AATAAA-3'.
proteinthe three possibleframes of translation
ofMo-MLV,Mo-MSV, and AKR in the 5'--* 3' directionareshown in theappendix. Thelargest possible open reading frame is foraprotein of about10kilodaltons encoded by AKR-614LTR. In the direction opposite tothe genomic RNA
transcription, the largest possible protein
syn-thesized is less than 7 kilodaltons. Since no mRNA encoded by a murine type C retroviral LTR has been reported, the role ofsplicing in generation ofan RNA molecule which can en-codeaprotein cannotbe ruledout.
Inversion of LTR in Mo-MSV. Although the LTR appears to be structurally analogous to movable genetic elements, no transposition of LTRs has been observed in the infected cell
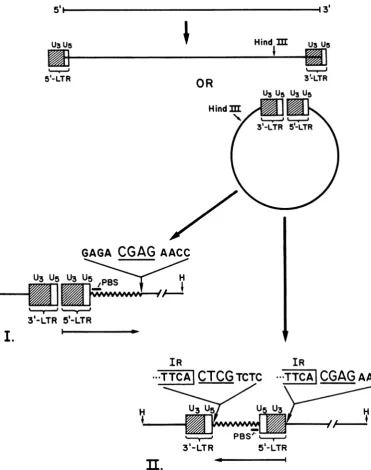
DNA. Shoemaker et al. (35) have, however, reported variants of molecularly cloned Mo-MLV DNAin whichoneofthe LTRsintegrated into the viral DNA, thuscreatingamolecule in which the two LTRs are not adjacent to each other.Asimilar situation hasnowbeen encoun-tered in amolecularly cloned Mo-MSVDNA.
We havepreviously described the molecular
cloningofthecircular form ofunintegrated
Mo-MSV DNA containing two LTRs in
bacterio-phage ) and its subsequent subcloning in pBR322 (43). Since the circular DNA was cleavedattheHindlIl site,themolecular clone was permuted with respect to the linear viral DNA.However, the two LTRsshouldbe pres-ent adjacent to each other as shown in Fig. 2 J. VIROL.
_ _
,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.490.55.452.63.393.2](structure I). Out of a total of three clones
analyzed,twoclones appearedtohavestructure
I, whereas in thethird Mo-MSVDNAclone,
A-MSV-1 (43; subcloned in pBR322, pMSV-12),
the LTRs are not adjacent but separated by a
stretch of 321 nucleotides (structure II). Further-more,the 5' LTR is inverted from the expected
orientation in Fig. 2 (compare structures I and
II). Several conclusions can be made from the
sequenceof inverted LTR in pMSV-12. (i) The
5'. i .,l
'I
U3 U5
5'-LTR
OR
Hind
m
Us
U5
I
3-LTR
GAGA
CGAG
AACC
U5
PB~~~
3-LTR/M I5MMLTA
t It 0
3'-LTR
5'-LTRI.
IR
*
TT
CTCGTCTC
ss s7 s~~~1 H
I
IR
*jTTCA
ICGAG
AACC
U3 l H
3'-LTR 5'-L-TR
D:.
FIG. 2. Structure of recombinant clones of Mo-MSV clone 124. Thegenomic RNAgivesrise to
double-stranded DNAhavingone(not shown)ortwoLTRs in either linearorcircularconfiguration.Thearrangementof
recombinant clones ofsupercoils,isolated fromproductivelyinfected cellsand clonedattheunique HindIII site, isgenerally that shown instructureI(e.g., clonepMSV-1L [39a]). Inonerecombinantclone, pMSV-12, the
regionindicatedbythearrow,encompassingthe 5' LTR and the 321nucleotides downstream from it(including the tRNAprimer binding site [PBS]),has been invertedtogivethe arrangement shownin structureII.Thetarget
tetranucleotideCGAG,foundonceinstructure I,is foundrepeatedatthe end of the inversion in structureII.
H
I
VOL.41, 1982
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.490.57.429.138.609.2]546 VAN BEVEREN ET AL.
5' LTR and 321 nucleotides downstream from it areinverted with respect to their orientation in pMSV-1L (structureI)(39a).(ii)The 321 nucleo-tides between the two LTRscorrespond to se-quencesrepresentingthetRNAbindingsite and downstream viralsequences. Theinversion pre-sumably occurred during viral DNA synthesis
andis not an artifact ofcloning (35). (iii) Two terminal A residues from the U3 region of 5' LTR and two terminalT residues from the
U5
region of 3' LTR are lost. (iv) A 4-bpinverted repeat(Fig. 2)is observedattheU3 terminusoftheinverted 5' LTR, andatthejunction ofthe
additional 321-bp sequences and
U5
terminus of the 3' LTR. It may benotedthat in contrast to theproviral DNAdescribedbelow(Fig.3and5)inwhich there isdirectrepeatofadjacent
cellu-lar sequences, the 5' LTR and the additional 321-bp unit are inverted; thus, the flanking se-quences display an inverted rather than direct
orientationbecause the molecule has
integrated
into itself. (v) The 4-bpinvertedrepeatispresent once in the parental DNA atthe site of
inver-sion.
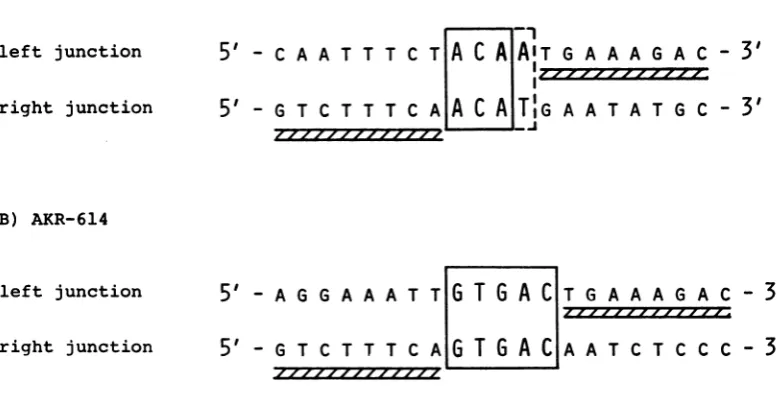
Host-viraljunctions of AKR-MLV 623and614.
A hallmark of structures resembling bacterial
transposons is the direct repeat ofcellular se-quences atbothtermini oftheintegrated mole-cule. We have recentlyreported the molecular
cloning of two infectiousintegrated moleculesof ecotropic AKR-MLV DNA (clones 623 and614) from a chronically infected NIH/3T3 cell line (24). We wanted to determine the nucleotide sequence of host-viral junctions of these two proviral DNAs. In particular, we wanted to determine whether the viral DNA has any pref-erencefor host sites. Figure 3 shows the nucleo-tide sequence of the junctions of AKR-MLV DNAs and mouse cellular DNAs. The inverted repeats at the termini of an LTR define the boundary of the viral sequences. In the case of AKR-623 DNA, the 5' LTR and host junction is ACAA, whereas the 3' LTR and host junction is ACAT. In all the retroviral DNAs analyzed so
far,the terminal two nucleotides are lost during the integration process. Assuming that similar rules apply to AKR-623 DNA, there appear to be either 3- or 4-nucleotide direct repeats of cellular sequences. If one out of the two terminal Aresidues is retained at the 5' junction, then the repeatis3nucleotides.Alternatively,the repeat is 4 nucleotides, and 1 nucleotide at either the 5'
or3'junctionhas undergone transversion.
In the case of AKR-614, the direct cellular repeatis5nucleotides (Fig. 3). Again, the termi-nal two Aresiduesare lost during integration. It
is interesting to note that the two AKR MLV
DNAs, depending on their site of integration,
A) AKR-623
left junction
right junction
5'
- C A A T T T C5'
- G T C T T T CB) AKR-614
left junction
right junction
5'
- A G G A A A T5'
- G T C T T T C--I
T
A
C
A AIT
G A A A G A C-I,4 ,7 7-.,7 ,ro,~,- /r~
A
A C A
T|G
A A T A T G C-T
G
T G
ACT
_--
G A A A G A CAIG
T
G
A C
A3
'
3'
-
3'
A T C T C C C -
3'
FIG. 3. Nucleotide sequenceatjunctions of viral AKR and cellular DNA. Thefragments sequencedfor the junctions of AKR-623 are given in the legend to Fig. 5.Todetermine thejunctions of AKR-614, the 12.9-kbp EcoRI insert ofX-AKR-614wassubclonedinto theEcoRIsite ofpBR322. The leftjunctionwasdeterminedby labelingthePstIsite in the 5' LTR, cleaving the 2.5-kbp PstIfragmentwithEcoRI,andsequencingthe1.75-kbp fragment. The right junction was sequenced by labeling the SmaI site in the 3' LTR, cleaving the 9.1-kbp fragmentwithBamHI,andsequencing the 2.9-kbp fragment. Duplicate nucleotidesareenclosed in openboxes, andthe11-nucleotide inverted repeatsatthetermini of the LTRsareindicatedbyhatched boxes.A,Junctions in cloneX-AKR-623;B,junctions in clone X-AKR-614.
J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.490.54.443.407.606.2]generatedirect repeats of cellular sequences of
either3 or 4bp asinthe caseofAKR-623 DNA
or a5-bprepeatinAKR-614DNA.
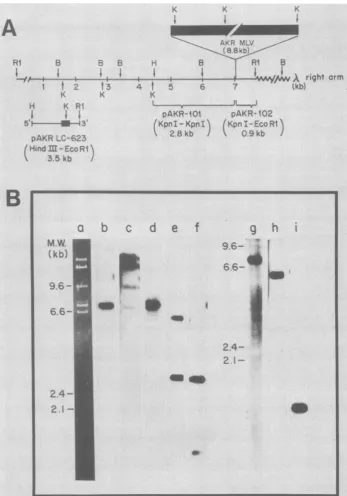
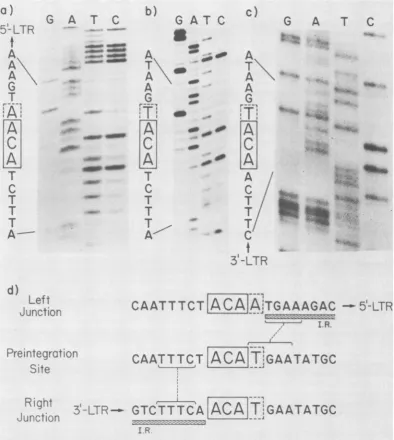
Nucleotide sequence ofpreintegration site. In
theproposed models for integration of movable genetic elements, it is imperative that the direct
cellular repeat sequences be present only once
at thesite of integration (33, 35). Since we were not sure whether the directcellularrepeat is 3 or 4nucleotides in the case of AKR-623, we deter-mined thenucleotidesequence of the
preintegra-tion site forAKR-623. Figure 4 shows the
char-acterization of the recombinant DNA clone, X-NIH-623, containing the preintegration site. The ethidium bromide stainingpattern(Fig.4B, lane a) shows that after cleavage with EcoRI, twoinserts of an average size of 7.0 and 7.8 kbp, in addition to the A Charon 4a arms, can be identified. However, when the DNA from the same gel is transferred tocellulose nitrate and hybridized to a nick-translated probe made from pAKR-LC-623 DNA, only the 7.8-kbp insert can
beidentified(Fig. 4B, lane b). The same 7.8-kbp
band hybridizeswhen thefilter is hybridized to
nick-translated X-AKR-623 DNA (Fig. 4B, lane
c). Inaddition, however, the A arms and some uncutDNA can also be identified. The A-AKR-614 DNA does not hybridize to the X-NIH-623
insertDNA (data not shown). Lanes d through i
of Fig. 4B display the analysis of X-NIH-623 DNAbyhybridization to 101 and pAKR-102 DNA. TheBamHI (Quint and Berns, per-sonalcommunication), KpnI, and EcoRI
restric-tion endonuclease map of AKR-623 DNA is shown in Fig. 4A, which also shows the se-quencesin plasmids pAKR-101 and pAKR-102. The restriction map of X-NIH-623 should be
identical to that of AKR-623 DNA minus the
AKR-MLV DNA sequences. To test this
predic-tion,we cleavedX-NIH-623 DNA with BamHI,
EcoRI, andBamHI plus
EcoRI
and hybridizedto labeled pAKR-101 and pAKR-102 DNA as
probes. In both cases, digestion with
EcoRI
shouldyieldaninsert of 7.8 kbp(Fig. 4B, lanes d
andg).DigestionwithBamHIand hybridization
topAKR-101 DNA should show a major band at about 2.8 kbp and a 6.0-kbp band containing NIH-623 and X right arm sequences (Fig. 4B, lanee). On the other hand, hybridization of the
BamHI digest topAKR-102 DNA should show
only the 6.0-kbp
fragment
(Fig. 4B, lane h).Digestion withBamHIandEcoRIand hybridiza-tion to pAKR-101 DNA should yield the 2.8-kbp band and a smaller 1.4-kbp
fragment
because EcoRIcleaves at the junction ofNIH-623 andXrightarm(Fig. 4B, lane f). However,
hybridiza-tion of pAKR-102 DNA of the
BamHI-EcoRI-digestedDNAshould yield onlyonefragmentof
1.4 kbp (Fig. 4B, lane i). Thus, it appears that the 7.8-kbp insert of X-NIH-623 DNA has a
restriction endonucleasepattern similar to t-hat
of the flankingcellular sequences of X-AKR-623
DNA.
Thenucleotide sequence of the preintegration site and flanking sequences is shown in Fig. 5. A sequence ACAT which constitutes the direct cellular repeat of X-AKR-623 can be identified to occuronce. This sequence is flanked at its 5' and 3' ends by sequences whichoverlap with flank-ing cellular sequences of X-AKR-623. It is
diffi-culttodecidewhether thedirect cellular repeat
inAKR-623is 3 or 4 nucleotides.
A striking feature of the sequences flanking thepreintegration site of AKR-623 DNA is the apparenthomologywith viral LTR sequences. A sequenceTTCC at the 5' end and TGAA at the 3' end of the preintegration site have homolo-gous sequencesin the inverted repeatsof 3' and 5' LTR sequences. No such homology can be
observedin the caseofAKR-614DNA.
DISCUSSION
Establishment of infectionbyretroviruses
re-quires the integration of viral DNA into host
chromosomes. Usually more than one copy of
the vital DNA is
integrated, although
not allproviralDNAsaretranscribed. The mechanism
ofintegration remains
totally
obscure.Howev-er, all proviral DNAs have aunique structure,
namely, U3U5. . .
U3U5.
Detailed nucleotidesequence analyses of the structure of several
retroviral DNAs
provide
a stronganalogy
withthe structure of
transposable
genetic
elementslikebacterialtransposons, the Tyl elements of
yeasts, and copia in
Drosophila
(reviewed
in reference 19). The retroviral DNA structure ischaracterizedbythe presenceof LTRs. The size
ofthe LTRs is
variable,
ranging
from 273bp
foravian endogenous
provirus (ev-1) (19)
to1,327
nucleotides for mouse mammary tumor virus
(12). The murineretroviruses vary insize from 500to650
nucleotides,
whereas the avian retro-viruses have a widersizerange,from 273nucle-otides forev-1virusto330 nucleotides for
RSV-SR-A(37), 350nucleotides forRSV-SR-D
(23),
and 569bp forspleen necrosis virus
(34).
Each direct repeat has inverted repeatsatits termini. The number of nucleotides in the inverted re-peats rangesfrom 3 forspleen
necrosis virusto 11 forintegrated Mo-MLV(38). Aspointed
out by Hishinumaetal.(19),the 5' and 3'termini ofallproviraland manytransposableelementsare
always TG ... CA. The LTRs described here are no exception to this rule. In the case of
unintegratedviralDNAs,however,the 5'
termi-nusisAATG,andthe 3' terminus is CATT. The
last 2 nucleotides at each terminus are lost
during integration.
Presumably,
thestaggered
cuts are madeatAA l TG and CA
t
TT(33,
35).
on November 10, 2019 by guest
http://jvi.asm.org/
w-AK~AR MLV
(8 8kb)
H B Ri B
f4w
uright
armt 5 6 7 (kb)
K
L _.f
pAKR-1I0 pAKR-102 KpnI-KpnI (Kpn I-Eco Rl
2.8kb / 09kb
A
RI B B B
I..
Si
IS
1 t 2 t3 4
K K
H K RI
5'I
I--.S..,4
3' pAKR LC-623 (Hind I -EcoRI3.5 kb J
M.W.
(kb)
9.6
-6.6
-a
b
c
d
ef
e0
g
h
9.6- t
6.6
2.4-
2.1-
2.4-
[image:7.490.73.421.55.552.2]2.1-aw
FIG. 4. Characterization of recombinant clone X-NIH-623.A,Predicted restrictionmapofthe7.8-kbp EcoRl fragment containing the AKR-623 preintegration site. Lengths are shown in kbp. Restriction enzymes: RI, EcoRI; B, BamHI; K, KpnI; H, HindIll. (Datafor BamHIarefromQuintandBerns, personal communication.) Regions ofhomology with AKR-623 recombinant subclones pAKR-101 and pAKR-102 are indicated with
brackets. Thelocation of the AKR-623integrationis shownbythe closed box. Thestructureof theinsertinthe AKR-623 subclonepAKR-LC-623,from theHindIIIsitetothe 5' LTRKpnI site, ligatedtothe 3'LTRKpnI site, andthroughtotheEcoRI site is alsoshown.B,Restrictiondigestpatterns ofX-NIH-623 DNA.X-NIH-623 DNA
wascleaved with EcoRI (lanesa,b,c,d, and g), BamHI (laneseandh),orEcoRI plus BamHI (lanesf andi).In
lanea,theDNAwasstained withethidium bromide. Inall othercases,theDNAwastransferredontocellulose nitrate sheetsand annealed tovarious nick-translated[32P]DNAs,and theresultant bandswere visualizedby autoradiography (3). The [32P]DNA probes usedwere:pAKR-LC-623 (lane b),X-AKR-623(lane c),pAKR-101 (lanes d,e,andf)andpAKR-102 (lanesg,h,andi). Wild-type phage DNAdigestedwithHindIIIwasused for size markers.
B
K K K
I on November 10, 2019 by guest
http://jvi.asm.org/
a
)
5'-
LTR
t
A
A
A
G
T
I
T
T
T
A
G
A
T
C
b)G AT C
- -a _
t_
A
-
A\-A
-0
r --^"
:T,
_
[A
, .-C
- T
T
-~~
A/
d)
Left
Junction
Preintegrotion
Site
c)
A
T
A
\2s
i
A
A
,A
T
C
G
A
T
C
*., :.Bs *4,
4
4%:~..,
*4
% 1
4
W
..
Q
* -s _*
.UW
..
t
3 -LTR
CAATTTOT
A
CAThjTGAAAGAC
r
I.R.
CAATTTOT
t_TIGAATATGC
_-__-
5-LTR
Right
Junction
3'-
LTR-
GTCTTTCA ACA
I--JTGAATATGC
[image:8.490.51.446.90.531.2]I.R
FIG. 5. Preintegrationsite and cellular-viraljunctions of recombinant clone X-AKR-623.All sequencesare
read from bottomtotopinthe 5'to3' direction. The nucleotides enclosed in boxesarethose sharedbyallthree sequences,with thepossiblefourth nucleotide enclosed inadotted box. a,Leftjunction.DNA sequenceladder ofthe fastermigratingstrand ofa 5' terminally labeled410-bpSau3A fragment ofthe 920-bpBamHI-KpnI fragmentof subclonepAKR-101. Reactions(26): G,Galone;A,A>C; T,T+C;C,C alone. Partialdigestion
productswereseparatedon a12%polyacrylamide-8Mureagel.b,Preintegrationsite. DNA sequence ladder of the slower migrating strand ofa 5' terminally labeled 255-bp Sau3A partial digestion fragnent ofa925-bp
BamHI-EcoRIfragmentof cloneX-NIH-623. Chemicaldegradationreactions and sequencegelwere asdescribed above. c,Rightjunction. DNAsequenceladder of the800-bpKpnI-EcoRIfragmentof subclonepAKR-1023' terminallylabeledattheKpnIsite. The lanes have been labeledtoallowdirectreadingof thecomplementary
strand. Partial chemical degradation products were separated on a 6% polyacrylamide-8 M urea gel. d,
Alignment of 5' and 3' junction sequences with the preintegration site. Sequences shared between the preintegration site and the inverted repeats (I.R.)of theLTRsareshownbybrackets.
4.0
on November 10, 2019 by guest
http://jvi.asm.org/
550 VAN BEVEREN ET AL.
A comparison of the nucleotide sequence of several integrated andunintegrated murine
re-troviral DNA LTRs allows certain
generaliza-tions. (i) TheU5 regionof the LTR is relatively conserved when compared with the U3 region,
which undergoes rather extensive changes,
in-cluding deletionsand substitutions (Fig. 1).The
heterogeneity in the U3 region is extensive
enough in that we were able to prepare a
Mo-MLVspecificprobefromtheU3 sequences(40).
Similar conclusionswere reached when the
nu-cleotide sequences of several avian retroviral
DNAs were compared(23, 37). (ii) The
impor-tant regulatory signals like the 5' cap, RNA
polymerase II initiation sites (TATA-like
se-quences), and CCAAT box sequences are
pres-entessentiallyatthe samerelativepositionwith
aminimum of change. Wehaverecently shown
thattheTATA-likesequences presentfrom -25
to -31from the5' capof bothunintegrated and
integrated Mo-MLV DNA are involved in the
initiation ofRNApolymeraseII(14a).Asimilar
region for RNA polymerase II initiation has
previously been identified in the avian sarcoma
viral LTR(44). (iii) Theinverted repeats atthe
termini of theLTRareidentical inallcases.(iv)
TheU3 regionof severalLTRs(e.g.,AKR-623,
unintegratedMo-MLV,Mo-MSV-ml, etc.)
con-tains a 58- to 100-bp repeat. However, other
LTRs(e.g.,
pMLVI-101
andpMLV-201)containonly one copy of sequences involved in the repeats. The nucleotide sequence of SV40
shows a 72-bp repeat about 300 nucleotides
upstream from the 5' capforthe late mRNA's (18, 30). It hasbeen shown that whenonlyone
copy of the 72 bp is present, no effect on the
transcription of simian virus 40is observed(1),
whereas when bothcopies of the 72-bp repeat
are removed, no transcription is detected. We
have previously shown that
pMLVI-101,
con-taining onlyonecopyofrepeat sequencesinthe
5'LTR, isinfectiouswhenligatedto aMo-MLV
DNAcontainingthe 3' half ofthe molecule (3).
One of the salient features of transposable
elementsisduplication of hostsequencesatthe
site ofintegration. The proviralDNAs havealso been shown to generate duplication of the host DNA as witnessed by the presence of direct repeatofcellularsequencesatitstermini(11,19, 25, 34). The number of bases involved in the
duplicationrangesfrom 6bpin MMTV andev-1
proviral DNA to S bp in the case of spleen
necrosis virus (SNV) and 4 bp in the case of severalmurineretroviral DNAs. Twointegrated
AKR-MLV DNAs have been analyzed in this
report.One of them, AKR-623, generates a 3- or
4-bp repeat, whereas the other, AKR-614, cre-ates a5-bpdirect repeat. Because the sequence at the 5' junction is ACAA and that at the 3'
junctionisACAT, there isanuncertaintyin the
precise assignment of the size of the direct
cellular repeat in the case of AKR-623. The
preintegration site in the case of AKR-623 is
ACAT,similar to that of the 3' junction. Asingle
base mutation at the 5' junction couldaccount for thedifference. Alternatively, it may be that thedirect repeat is only three nucleotides and thatonlyoneinsteadoftwo Aresiduesislostat the 5' terminus. Another explanation may be suggested as a result ofsequence analysis of a two-LTR clone ofMo-MSV (39a). This clone hasthe sequence TTAAA at the junction of the
LTRs, so that a loss of the terminal two A
residues from such a clone would still leave a 5'-terminal A on the 5' LTR. Since all murine retroviral DNAs analyzed so far, including the inverted LTR variant reported here, show a4-bp
repeat, itis tempting to suggest that AKR-623
also has a4-bprepeat.However, in the case of AKR-614, the direct repeat appears to be 5bp.
Thus, AKR viral DNA seems to be anexception
among murine retroviral DNAs, being able to integrate either by generating 3- or 4-bp repeats orby creating a 5-bp repeat of host sequencesat its termini. It appears from various retroviral
DNA data that the virus and not the host deter-mines the number ofnucleotides duplicated at the site ofintegration. For instance, Mo-MSV
growninheterologousmink cellsalso shows a 4-bp repeat like that of other murine retroviruses. Among avian viruses, SNV and ev-1 generate
different-size direct cellularrepeats, namely, 5
and 6 bp, respectively. The observation that
AKR-MLV DNA integrated at two different
sites in the same host generates a different number ofduplicatedsequencesis in agreement withthisnotion.
Animportantconstraint in themechanism of
integration oftransposable elements is that
du-plicatedcellular sequences can be present only
once atthesiteofintegration(33).Thishas been
demonstrated in the case of MMTV and ev-1
virus (19, 25). Inthis manuscript, we have also shown that the cellular sequences constituting
the directrepeat in the case ofAKR-623 DNA
arepresent only once in the preintegration site. Inthe case of MMTV and ev-1, there seems to be no apparenthomology between theflanking
cellular sequences and viral LTRs. However, in the caseofAKR-623, onenucleotide to the left of the preintegration site is a sequence TTTC
homologous to TTTC at the 3' terminus of3'
LTR(Fig. 5). Similarly, a sequence TGAA
im-mediatelytotheright ofthepreintegrationsite is
homologoustoTGAA presentatthe 5' terminus
of 5' LTR(Fig.5). Thus,hadacircular form of AKR-MLV been the precursor to theAKR-623
clone, there would have been considerable
se-quence homology between cellular DNA and viral LTRs atthe siteofintegration.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
4) UO C) C 44 CD Id 00C
go U .-. U
Id 0 01~~~~~ C)~ go 0 4.45
440'~~~P4 4 54 r $4 0 50 04
ra ggoo. 010
go14 5 co UI 0
4.' 0>d '0g 0> 5
goU 4)U~~~~~cn MUL 01 4 C
t rI~ > -1C 01 d e4 U
$4 ty050'
.454 >~~~~~~'0 0 r-o go % *Vco 0c
~~go~ 54r*'4 0d v 0nu40
,a E-4~1'0>5>
I4 0 4o ra U>E54go )'
0i UN 44' U4 .- o u)
> $020 > 4 4J U C
..454 V IdO~~~~~~~~~~0 >1
4)U 010 ~~~~~~~~~01'0 ~ CDU go4 U104
§(4 IUM go la "- <
54 :1001.0
)Ue
$440 go'0*-E-4>1IdOIC 44tp4gJ0 5 01 0
04 UO UO r4 54 0 0- go UO
0'Id~ 0 U0 go UON go44 n 4
-4i ~~~~om ~
-r4E-4 CU g UD go D
laa0% LgoC" 40n
AtU olU 4U0 U)
U Q.' UAd0U a
01 39~~~~0' 4
>54 go U go~~~~~~~~$ UEV '0
~~ 0102 l~~O o~~ god . 4
tyi co L)~~~~~~0 '44 4
Ad tp 4UOU 4 54 Y
01- V) C) goE-Cm 4$ 54 C
44 t~o U ' 4 OUM .--4~U)~
waCD'0 go U > U- ) o
010O010 - -01'0 -4~ .IJU n 1.
Id0 J U>
4 '0 01 '0 go oo 44 00 ri
-~~U ~~~~01~OA o'u 0- 4.
AdI C
4() U cooU
.~~~~,'0 OUm % 0 go U gDo UZN
U r4N 01 5r4L
~~ .I.IUO la0540 01.00 .U0
U4 >
gojglIi~~~ >0*rgo~ U
to r-4 QU co uo
0
U> of U DC
Os go 0 Ln Ur- *i C
od -'0- E0'4gU uc r
on November 10, 2019 by guest
http://jvi.asm.org/
552 VAN BEVEREN ET AL. J. VIR~OL.
01 0
C.r-4.1 U0 aU o a01 40C 04 CD
V4r IV 44 U%O Ul Uco
~~~01 U) U
~~~~~~~~~~0
U > E'UE-4 *.-~~~4E-4 'UU01
wi
01 ~ 4)Olb
01b.401cn , inL ~
0r-01 0'-4 4) f1 '-4 E-4
-~~01~~~~~' 01 -'~~~~4 "--4 E-4
012
. 4> E-4 Uto U 44
0101 '1-~~~44E-4 r-4
E-4>E-U) CD 01, CeJ w4 0 w 010
-"4 E-~~~-4 'Ug U4 Q cUr U) Uw
VeU~ U)0 0)U "-4E-4
U) "-4E&4 '-4 E-4 00
4.4 E4 04 U 'UU
> E-4 04 U>
-9 C0CD t 010 U) Ui E-40C
.9 V 4 4-4 r E-.4un
4)AdU
'UU V J ~'>4E4
01E-4) D~ Va ~oC 44 E-40 c 0 010
4' 01 aCD ~ >E-4c,4 r-4E-i
4i
Ea~~~~~~>
E44U ~~~~~~~ 0U
co)
014'~~~~~~~~U 4.4 E-4 co U
"4>E-4UU go 4 0 0E1
-4E-4 >E-4 CDr'UU D E4c
C: r-. ~ U) U-4 04 Urn~
Ad w ~~01"-4 4.4 VQu
441 "4W E-4 r
4) E4 ~ C >- 01
010 4)4LaU)
'UE-4 .~~~~~~~~~~~~ >"4 E14
012
mu)01E01 01 01
04
01 U)Umu.
E- "-4 E4 01
4-4E-4 'UU ~~~~~~~r- E4 a
4-4 E- 00 ~ E-4 Q 0
C)4UC)i U~~~~~~~~~~~~~-44
'UU QE U 4' 01 0~~~~~~~~101U
-40 'UO "-4E- 00
C "4-4 r-4" E4 ) U
CD4
U)U-
-4D
UU) 0-4 Er -4 "-4 E4 to 01
r-4~ ~ >2co C)4U U)U
to Eu to U UC 24 to
'-4~ -4 01 4~-i 0 QCI U)4 44 E
'UU~~~~~~4 4) 4
E-4
,-o
Eto4 E2-4-4 C>UEV
.bd C~ > E4 4 01CD ."4 E-40 01 01
01~~~~~ 0 01~~~I'( Er-- gc 01 01
0101 U)~~~toU . C). U 0101
01 4 E-' ) U 01
'-4E-4 C).~r- U- U) 0
E-4C). U U U 01 a
0101 0101~~lm .4.1 >2 U)Uto
01 a Va 'mU 01 > 2-.
~~% 4, ~~o 01~ oC 017 01 01 4C0 010O
Y.., >E-4C4 44U1 w0%01UoOOCD.UMU
0 C -4 . 4 'U Ur C). Un
C'Jof v1
4-01
4.1g c to
v1
CD.U
'44UC U.E-).C UmaCr-E-CD0 "-4 E24C
ra >I U C). U 01- 01 44 U
UC010AC) 01ur1- 0Eit
to u U) U o 44UO 'U2~~~-e0 01010 -4E.4 -42F
>1~~~ 01~4 V1f 44 Ur V 4O
a E~~~~~~-44
on November 10, 2019 by guest
http://jvi.asm.org/
MLV LTR 553
N
-IV
$4 C
)
r- E- o
Y 04 OC) 4) C1 < P >riE-'oQ 4 44 04 COU '-4 E XO U to r- E-4 EU4 oc 44 0E-0 r r 44U 4. 4 e) E01 I E-' U~ Eu U COO EuU Eu E-z 04 U) Utv= Y4 #: E-1U UO Es COO to 440 > :N (A' ON CD oo 4: ON CD 44 R JJ 04 U) LI) r-i E-4 Q > E-4 > .,.'EU 04 : 4 04U EuaU c 04 U) EuU mO U
CUn Q *.4 E-' EuUO 04 04oU to CD E-ie' > 440 ': C Ll u~ P)00 CD
4 01Y toUo UJ Ea Es '04: 440 01 :U 4-4 E-40( c < c4 u > E-irv, JJNO cO 0¢ 4 4 m O 4 4 014 04.
> ^E-1 o 'C4 mcJ to i4 EuU cC v4 4d 440 O EJuU U 0m u EuU
co U CO
E42-i 4. 4J 4 U 4U4 4U4 CD .r4 E-> 2-' 010 E-' '-42-go< r- E-4C ---E-4 r-er 4u UL >u to 4 4 U
--4E2-q
44 u o m cJn U -4E U 44 E-4 Uo E2-i
m cr
6
EnuU 4 E00 ON0 "-4 LI) 4: uo U' Eu .Eu o u0> Euu 0U'-4 E4CD 4E%
toa0 t L
U > E-1 4-0 Uo 04 to un to<: CO1 r- E4 u
> 0110
E2-m -H E-4 to .,, E-i 04u C U 44 0E C
.HE-4 > 2-'9 >: Cu uC,4 > C PO U uo P u u 04EU U 4 CD uun
-4 Ln > E4% 04e
---4CD-C
>l
04 EUe
U cr(
U f u
oU
on November 10, 2019 by guest
http://jvi.asm.org/
554 VAN BEVEREN ET AL. J.VIROL.
Eu (O 01 (3o 4) CDO 4 (Do co (.C w (30 k.Co >1 co0
al24~ 04(dcU 0s4 n
4) U 0al (. ON0 (
04 d E- 0 (
Ad~ ~ E(d4-4~ .
'. ( r4 >1 04 (
.NC r04d-4E-4 E(
44
d,4 E-4cD 04 .) fCQ
J 01
>>
-4c E4EUn
(Ed
01 4 co 1 cnr4)
-> ~ > E44) (d W 3 to (
01(3 01 (3 V~~~~~~~~r oc > E-4
W4(0CD3(3o (aCD Q4 0130C
.-4 E-1 to (d coU? (
Va -~4 E- o 44'4) (
01a,.- E-' to (d14 (
U) (d V~~~~r 94 04 > E-' ) (
>4E-'o '~(o 1 V ~
4t(do
EU0>U) W300 EU,- 01 1 04a, CDU 04 (dN
(A a, gc4(d~ '.4(ON 4) v)dE 4)(rU
C,4 toU
-W (3 ~~~~~04u 04 (d EU (d V
EU ( 04 d 01 ( LI(3
-4E-4 01(3EU( W.43r-4E-4
0~~~~O~ 3% (0C > E4 4 E-4a Vo CD W 300 0E-'c 01(3C4 4)(dU t ~
0 ~~~~~ V~r 4c~ 01(3(4) > ~Ln
440E414
'14E-4 44 E-4 4J U~~d '-
E-EU (d > E-4 04 (
4)(d0 U)(3o CD30 01toU CDo >1 04 01 C1 '-4 4) (dr' .-4 E-"n
raoc W0 u ~ 0
01 (3 Vu) ( co (3 01 (
:3(3 w(3 *.~~~~~~~'i -4E to 01
~~EO
01~~~~
Eo
) 4mu.9
gc O 4) (00o 0
01(3 ~~~~~~~~~~~~>E-*4 E-4 04 (
1.4 (3 4.4(3 (A
EU(d04 (d U) (d ~~~~~~~~~~~-4
E-'~~W
C)E-~~4E-' 01 (.9 '4.~~~~~V4 E-' U) (3 EU E 4)(d01~~~~~~u-0 r(44 0 (E- s
U) v EU 4( U) (to U)
EU(d EU (d~ ~~~01 E (A) d Ca(3
.4 a(3 01 ( U)> UQ 4-i E'D W
4i u 0(o U(0u(d*04E-4oL
U) 34 E (d' to(3a 4&4
E-io>E-W a ~ 0) (3- 01&44E
.-a4r- E-4( EU( 4.4>
~~-IE' 04 ~~(dU~~~( W LI 04 (
Ae -4E-4 '.E-1(3 v i
EU (d EU (do-4 ( u t
U) (3r U) 44 V-u 0r-'-4 E-4.
EU (d ~~~~~>E-4 04 (d U (d
01( 01 4dCD4.40 E-4
EU(d-> E U)n( >EU (d% ~44 E
01(3 0(3 '.4(3~~c>5-i r- 0
>.( 0( to (dU) (
~0 U) (do : (30 4 E4o 5
(d * : 5- v.1(3 4
>4 0( 4) (
mu( Q1 '4>-EE4
04 (d -UE) -W %O (3 1Om (
VW .4 0 0e' (dO)144 cn t u
(3 04 Ed-4 EU (dc% U) ad
mu tr04 4 5- 4404E(d
U)mu 4i u W E(d> (
on November 10, 2019 by guest
http://jvi.asm.org/
The retroviruses have a strong structural homology with transposable elements. The mechanism of integrationalso suggests thatthey behave likemovablegeneticelements. Further-more,like transposons,theyeastTylelementor the copia element of Drosophila, retroviral DNAs integrate at multiple sites in the host chromosomal DNA. Unlike the bacterial tran-sposons,however,the retroviral DNAs have not been shown to move from theirintegration site in the chromosome. The inverted LTR variant reported here and before (35) offers a direct proof that LTR can integrate like transposable elements by creating a duplication of the se-quences atthe siteofintegration.
APPENDIX
The completenucleotide sequence of the LTRs and the possible open reading frames of the following murine retroviral DNAs have been determined (Fig. 6):
(i)pMLV-lA. Thecircularform ofunintegrated Mo-MLV DNA containing one LTR was molecularly cloned asdescribedpreviously (3).
(ii) pMLV-201. The construction and nucleotide sequence of the viral DNA contained in pMLV-201 haspreviouslybeendescribed (36). There were report-ed tobe afewnucleotide changes in the LTR sequence of pMLV-201 when compared to the LTR sequence of pMLVI-101 (an integrated Mo-MLV DNAclone) (3, 38). Inparticular,atpositions -25 to -31 from the 5'-cap nucleotide, the sequence in pMLV-201 was CAAAAAAascompared withCAATAAA in pMLVI-101. We have independently determined the nucleo-tide sequence of the LTR in pMLV-201 and found only oneof theoriginalthreechanges(atposition88,i.e., -362fromthe5'-cap nucleotide).
(iii)pAKR-614. The13-kbpinsert fromintegrated A-AKR-614 DNA (24) was cleaved withrestriction endo-nuclease EcoRI and subcloned in the unique EcoRI site ofplasmid pBR322. The nucleotide sequence of the LTRof623 DNA is identical to that of AKR-614 DNA.
(lv)pMSV-12. The circular form ofunintegrated Mo-MSVDNAcontaining2LTRs wasmolecularlycloned asdescribedpreviously (43). InpMSV-12,the 3' LTR integratedinto the viral DNA at 463 nucleotides from the 5'-cap nucleotide. Furthermore, the 3' LTR is inverted in its orientation with respect to the 5' LTR as shown in structure II of Fig. 2.
ACKNOWLEDGMENTS
We areindebtedtoJimOstlund for help in the computer analysis of the nucleotide sequences. We thank Janice
Galle-shaw andDoug Murdock forexcellent technicalassistance, andMaureen Brennan for typing the manuscript.
The workreportedherewassupported by grants from the NationalCancerInstitute and the American Cancer Society.
ADDENDUM IN PROOF
Thenucleotide sequence of the LTR ofan indepen-dently isolated clone of MSV-124 (29) was treated separately in Fig. 1 due to the occurrence of 26 differences with respecttothe LTRofpMSV-12 (43). In a recent publication, however, the authors have
made 26 base changes in their 582-nucleotide LTR sequence (E. P. Reddy, M. J. Smith, and S. A. Aaronson, Science 214:445-450, 1981). The revised sequence now has only one difference from that of pMSV-12 (atposition 85). The line in Fig. 1 for MSV-124should, therefore, be almost identical to that for pMSV-12.
LITERATURE CITED
1. Benoist, C., and P. Chambon. 1981. In vivo sequence requirements of theSV40early promoter region. Nature (London) 290:304-310.
2. Benz, E. W., Jr., and D. Dina. 1979. Moloney murine sarcoma virions synthesize full genome length
double-stranded DNA in vitro. Proc. Natl. Acad. Sci. U.S.A. 76:3294-3298.
3. Berns, A. J. M., M. H.-T.La, R. A. Bosselman, M. A. McKennett, L. T. Bacheler, H. Fan, E. C. Robanus
Maan-dag,H.v.d. Putten, and I. M. Verma. 1980. Molecular cloning ofunintegrated and a portion of integrated Mo-loney murine leukemia viral DNA in bacteriophage lamb-da. J.Virol. 36:254-263.
4. Bishop, J. M. 1978. Retroviruses. Annu. Rev. Biochem. 47:35-88.
5. Blattner, F. R., A. E.Blechl, K.Denniston-Thompson,H. E.Faber, J. E. Richards, J. L. Slightom, P. W. Tucker, and0.Smithies. 1978.Cloninghumanfetal-y-globin and mouse a-typeglobin DNA: preparation and screeningof
shotgun collections. Science202:1279-1284.
6. Bosselman, R. A., and I. M. Verma. 1980. Genome
organization ofretroviruses. V. In vitrosynthesized Mo-loney murine leukemia viral DNA has long terminal redundancy. J. Virol. 33:487-493.
7. Bukhari, A. I., J. A. Shapiro, and S. L. Adhya, ed. 1977. DNA insertion elements, plasmids and episomes. Cold Spring HarborLaboratory, ColdSpring Harbor, N.Y. 8. Calos, M. P., and J. H. Miller. 1980. Transposable
ele-ments. Cell20:579-595.
9. Chang,A. C. Y., J. H. Nunberg, R. J. Kaufman, H. A. Erlich, R. T. Schimke, and S. N. Cohen. 1978. Phenotypic
expressionin E. coli of a DNA sequence coding for mouse dihydrofolate reductase. Nature (London) 275:617-624. 10. Corden, J., B. Wasylyk, A. Buchwalder, P.Sassone-Corsd,
C.Kedinger,andP.Chambon. 1980.Promoter sequences ofeukaryotic protein-coding genes. Science 209:1406-1414.
11. Dhar, R., W. L. McClements, L. W. Enquist and G. F. VandeWoude.1980. Nucleotide sequencesofintegrated Moloneysarcomaproviruslongterminalrepeatsand their
hostandviraljunctions. Proc. Natl.Acad. Sci. U.S.A. 77:3937-3941.
12. Donehower, L. A., A. L. Huang, and G. L.Hager.1981. Regulatory and coding potential of the mouse mammary tumorvirus long terminal redundancy. J. Virol. 37:226-238.
13. Dunsmuir, P., W. J. Brorein, Jr.,M. A.Simon,andG.M. Rubin. 1980. Insertion of theDrosophila transposable element copia generatesa 5 basepairduplication. Cell 21:575-579.
14. Farabaugh, P. J., and G. R. Fink. 1980.Insertion of the
eucaryotic transposableelementTylcreates a5 base pair
duplication.Nature(London) 286:352-356.
14a.Fuhrman,S.A., C. VanBeveren,and I. M.Verma.1981. Identification ofanRNApolymerase II initiation site in thelong terminal repeat ofMoloneymurine leukemia viral DNA. Proc. Natl. Acad. Sci. U.S.A. 78:5411-5415. 15. Gilboa, E., S. Goff, A. Shields, F.Yoshimura,S. Mitra,
and D. Baltimore. 1979. In vitro synthesis ofa9 kbp terminally redundant DNA carrying the infectivity of
Moloneymurine leukemia virus.Cell 16:863-874. 16. Gilboa,E.,S. W.Mltra,S.Goff,andD.Baltimore. 1979.
A detailed model of reverse transcription andtests of crucial aspects.Cell 18:93-100.
17. Grosschedl, H., and M. L.Birnstlel.1980.Identification of
on November 10, 2019 by guest
http://jvi.asm.org/
556 VAN BEVEREN ET AL.
regulatorysequencesintheprelude sequencesofanH2A
histonegeneby the studyof specific deletion mutants in vivo. Proc. Natl.Acad. Sci. U.S.A. 77:1432-1436. 18. Haegeman, G., and W. Flers. 1980. Characterization of
the 5'-terminal cap structures of early Simian virus 40 mRNA. J. Virol.35:955-961.
19. Hishinuma, F., P. J. DeBona, S. Astrin, and A. M. Skalka. 1981.Nucleotide sequenceofacceptorsiteandterminiof integrated avian endogenous provirus ev-1: integration
creates a 6bp repeatof host DNA. Cell 23:155-164. 20. Hsu, T.W., J. L. Sabran, G. E. Mark, R. V. Guntaka, and
J. M. Taylor. 1978.Analysis of unintegratedavianRNA tumorvirus double-stranded DNAintermediates.J. Virol. 28:810-818.
21. Hughes, S. H., P. R. Shank, D. H. Spector, H.-J. Kung, J. M.Bishop, H. E. Varmus, P. K. Vogt, and M. L. Brelt-man.1978.Provirusesof avian sarcoma viruses are termi-nally redundant, coextensive with unintegrated linear DNAandintegrated at many sites. Cell 15:1397-1410. 22. Jones, M., R. A.Bosselman,F. A. V. d. Hoorn, A.Berns,
H.Fan, and I. M. Verma.1980.Identificationand molecu-lar cloning of Moloney mouse sarcoma virus-specific
sequencesfromuninfectedmousecells. Proc.Natl.Acad. Sci. U.S.A. 77:2651-2655.
23. Ju, G., and A. M. Skalka. 1980. Nucleotide sequence
analysis of the long terminalrepeat(LTR)of avian retro-viruses: structuralsimilarities with transposable elements.
Cell22:379-386.
24. Lowy, D. R., E. Rands, S. K. Chattopadhyay, C. F. Garon, andG. L. Hager.1980. Molecular cloningofinfectious integrated murine leukemia virus DNA from infected mousecells. Proc. Natl. Acad.Sci. U.S.A.77:614-618.
25. Majors, J. E., and H. E. Varmus. 1981. Nucleotide
sequences athost-proviral junctions formousemammary tumorvirus.Nature(London)289:253-258.
26. Maxam, A. M., and W.Gilbert.1977. Anewmethodfor
sequencingDNA.Proc. Natl.Acad.Sci. U.S.A.
74:560-564.
27. Proudfoot, N.J.,andG. G. Brownlee.1976.3-non-coding regionsequencesineukaryoticmessengerRNA. Nature
(London)263:211-214.
28. Rands, E., D. R. Lowy, M. R. Lander, and S. K. Chatto-padhyay. 1981.Restriction endonucleasemappingof
eco-tropic murine leukemia viralDNAs: size and sequence
heterogeneity of the long terminal repeat. Virology 108:445-452.
29. Reddy, E.P.,M.J.Smith,E.Canaani, K. C.Robbins, S. R.Tronick, S. Zain, and S. A. Aaronson. 1980.Nucleotide
sequence analysis ofthetransforming regionand large terminal redundancies of Moloneymousesarcomavirus. Proc.Natl.Acad. Sci.U.S.A. 77:5234-5238.
30. Reddy, V.B.,P. K.Ghosh,P.Lebowitz,M.Piatak,andS. M. Weissman. 1979. Simian virus 40 early mRNAs. I.
Genomic localization of3'- and 5'-termini andtwomajor splicesin mRNA fromtransformedandlyticallyinfected
cells.J.Virol.30:279-2%.
31. Sabran, J. L., T. W.Hsu, C. Yeater, A. Kaji, W. S. Mason,andJ. M. Taylor. 1979. Analysis ofintegrated
avianRNA tumor virus DNA in transformed chicken,
duck andquail fibroblasts. J. Virol. 29:170-178.
32. Shank, P. R., S. H. Hughes, H.-J.Kung, J. E. Majors, N. Quintrell, R. V. Guntaka, J. M. Bishop, and H. E. Varmus.1978.Mapping unintegrated aviansarcomavirus DNA:terminiof linear DNA bear 300nucleotidespresent once or twice in two species of circular DNA. Cell 15:1383-1395.
33. Shapiro, J. A. 1979. Molecular modelforthetransposition andreplicationof bacteriophage Mu and other transpos-able elements. Proc. Natl. Acad. Sci. U.S.A. 76:1933-1937.
34. Shlmotohno, K., S. Mizutani, and H. M.Temin. 1980. Sequence of retrovirus provirus resembles that of bacteri-altransposable elements. Nature (London) 285:550-554. 35. Shoemaker, C., S. Goff, E. Gilboa, M. Paskind, S. W.
Mltra, and D. Baltimore. 1980. Structure of a cloned circular Moloney murine virus DNA moleculecontaining aninvertedsegment: Implication for retrovirus
integra-tion.Proc. Natl. Acad.Sci. U.S.A. 77:3932-3936. 36. Sutcliffe,J. G., T. M.Shlnnick,I. M. Verma, and R. A.
Lerner. 1980.Nucleotidesequenceof Moloneyleukemia
virus: 3'-end reveals details of replication, analogy to
bacterialtransposonsand an unexpected gene. Proc. Natl.
Acad. Sci.U.S.A. 77:3302-3306.
37. Swanstrom, R., W. J. DeLorbe, J. M.Bishop, and H. E. Varmus.1981.Nucleotide sequence of cloned unintegrat-edavian sarcomavirus DNA: viral DNA contains direct
and inverted repeats similar to those in transposable elements.Proc. Natl.Acad.Sci. U.S.A.78:124-128. 38. Van Beveren, C., J. G. Goddard, A. Berns, and I. M.
Verma. 1980.Structure of Moloney murineleukemia viral DNA:nucleotidesequenceof the 5' longterminal repeat andadjacent cellular sequences. Proc. Natl. Acad. Sci. U.S.A. 77:3307-3311.
39. Van Beveren, C., J.Galleshaw,V. Jonas, A. J.M. Berns, R. F.Doolitile,D.J. Donoghue, and I. M. Verma. 1981. Nucleotide sequence and formation of the transforming geneofamouse sarcomavirus.Nature(London)
289:258-262.
39a.Van Beveren, C., F. van Straaten, J. A.Gaileshaw,andI.
M. Verma. 1981.Nucleotide sequence of the genome of a murine sarcoma virus. Cell 27:97-108.
40. Van der Putten, H., W. Quint, J. vanRaa1j,E. Robanus-Maandag,I.M. Verma, and A. Berns. 1981.
M-MuLV-induced leukemogenesis: integration and structure of
recombinantproviruses intumors.Cell24:729-739.
41. Varmus, H. E., S.Heasley,H.-J.Kung, H. Oppermann, V. C.Smith,J. M. Bishop and P. R. Shank.1978. Kinetics of synthesis, structure and purification of avian sarcoma
virus-specific DNA made in the cytoplasm of acutely
infectedcells.J. Mol.Biol.120:55-82.
42. Verma, I. M. 1977. The reversetranscriptase. Biochim.
Biophys.Acta473:1-38.
43. Verma, I. M., M. H.-T. Lai, R. A. Bosselman, M. A. McKennett, H. Fan, and A. Berns. 1980. Molecular clon-ing of unintegratedMoloney mouse sarcoma virus DNA in
bacteriophage lambda. Proc. Natl. Acad. Sci. U.S.A.
77:1773-1777.
44. Yamamoto, T., B. de Crombrugghe, and I. Pastan. 1980.
Identificationof afunctional promoter in the long terminal repeat of Rous sarcomavirus.Cell22:787-797.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/