0022-538X/92/010469-11$02.00/0
CopyrightC 1992, American Society for Microbiology
The
Six
Conserved
Helicase Motifs of
the UL5 Gene Product,
a
Component
of the Herpes Simplex Virus
Type
1
Helicase-Primase,
Are
Essential for Its Function
LIANGZHUt AND SANDRA K. WELLER*
DepartmentofMicrobiology, The UniversityofConnecticut Health Center, Farmington, Connecticut 06030
Received 19 June1991/Accepted 26 September 1991
The UL5 protein of herpes simplex virus type 1, one component of the viral helicase-primase complex,
contains sixsequencemotifs found inall members ofasuperfamily of DNA and RNA helicases. Although this
superfamily containsmorethan20members ranging from bacteriatomammalian cells and their viruses,the
importanceof these motifs hasnotbeenaddressed experimentally forany oneof them. In this study,wehave
examined the functional significance of these six motifs for the UL5 protein through the introduction of
site-specific mutationsresultinginsingle amino acid substitutions of themosthighly conserved residueswithin
eachmotif. A transient replication complementationassay wasusedtotesttheeffectof each mutationonthe
function of the UL5 protein in viralDNAreplication. In thisassay, amutantUL5 protein expressed froman
expression clone is usedtocomplementareplication-deficient nullmutantwithamutation intheUL5genefor
theamplification of herpes simplex virusorigin-containing plasmids. Eight mutations in conserved regions and
threesimilar mutations in nonconserved regionsof the UL5gene wereanalyzed, and the results indicate that
all six conservedmotifsareessentialtothefunctionof UL5 protein in viral DNA replication;ontheotherhand,
mutations in nonconserved regions are tolerated. These data provide the first direct evidence for the
importance of these conserved regions in any member of the superfamily of DNA and RNA helicases. In
addition, three motif mutationswere introduced into the viral genome, and thephenotypic analyses of these
mutantsareconsistent withresultsfromthe transient replication complementationassay. Theabilityof these
threemutantUL5 proteinstoform specificinteractions with other members of the helicase-primase complex,
UL8 and UL52, indicates that the functional domains required for replication activity of UL5are separable
from domains responsible for protein-proteininteractions. It is anticipated that thistypeof structure-function
analysis will leadtotheidentification of protein domains that contributenotonlytotheenzymaticactivitiesof
helicaseorprimase but alsotoprotein-protein interactions within members of the complex.
Unwinding of duplexDNAand RNAis essential formany biological processes, including replication, recombination, repair, transcription, and translation. This activity is per-formed incellsbyhelicaseswhich contain anintrinsic DNA-orRNA-dependentATPaseactivity.Thehydrolysisof ATP supplies energy for unidirectional translocation along one strand ofduplexDNA orRNA, resultinginunwinding ofthe duplex. The essential role ofDNAhelicases in DNA repli-cation has been studied invarious prokaryotic systemsand in the eukaryotic virus simian virus 40 (9, 25, 31, 39). A helicase
activity
in herpes simplex virus type 1 (HSV-1)-infected cells wasfirstreported in 1988 (7).Protein sequence analysis has revealed the existence of well-conserved motifs ina superfamily ofknown and puta-tive helicases, including Escherichia coli proteins UvrD, Rep, RecB, and RecD, the yeast helicase PIF,
proteins
involved inpositive-strandRNAvirusRNAreplication,
and fourproteins
oftheherpesvirus family (Fig. 1) (18, 19, 23). Consistent with theirability
tohydrolyze
ATP, these pro-teins all contain two highly conserved motifs known todefineanucleotide
binding
domain(motifs
IandIIinFig. 1)
(43).Inaddition, four other conserved motifs (IIItoVI)have beenobserved. The strong conservation of these sixmotifs in a large number ofRNA and DNA helicasesimplies
thatthese sequence elements may be important for helicase
*Correspondingauthor.
tPresentaddress: MGH CancerCenter,Charlestown,MA02129.
function;however, nodirect evidenceexists to demonstrate
theirfunctional significance.
The UL5 geneof HSV-1has beenidentifiedas anessential
gene for viralDNA replication through the studies of
tem-perature-sensitive (ts) (48) and host range mutants (50).
Temperature shift experiments with ts mutants in theUL5
gene indicatethat thisgene isrequired continuously
during
viral DNA synthesis, suggesting a direct role for the UL5
geneproduct(48).Theexistence ofaconsensusATP
binding
site within the gene raised thepossibility
that thisprotein
maybeanATPase or ahelicase(32, 48). Itisnowclearthat
theUL5geneproduct ispartofacomplex made upofthree
viral proteins (the products of the UL5, UL8, and UL52
genes) which exhibits helicase and
primase activity
(6,8).
These activities can be assembled in vivo in insect cells
triply
infected with baculovirusesexpressing
UL5,UL8,
andUL52 gene products (3, 10). The
coexpression
of UL5 andUL52 gene products has been shown to be sufficient for DNA-dependent ATPase and helicase (3) and for helicase andprimase (11) activitiesthatare
indistinguishable
from the three-subunitcomplex.Sequence analysis of the UL5 gene indicates that it is a
member of the
superfamily
of helicases described above (17-19, 23). As with other members ofthissuperfamily
of helicases, the functionalsignificance
ofthe six conserved motifs in UL5 has not beenaddresseddirectly.
We report hereinourinitialattemptstoprobe
theimportance
of these regionsin the UL5 geneby
introducing
directed aminoacid substitutions into each ofthe conservedregions.
Because469
on November 10, 2019 by guest
http://jvi.asm.org/
470 ZHU AND WELLER
0 100 200 300 400 500 600 700 800 900 1000 1100
I 11 III IV V V1 882
721
-.:.f.,S',S...,a..-..iS... ''Z .' :
1181
638 683
S~~~~~~~~~~~.fi.".
... ...f'''i:s:::....eFIG. 1. Conserved motifs inasuperfamily of helicases.The top line represents the number of amino acid residues. Sixproteins(UL5;E. coli proteinsRep,UvrD, RecB, and RecD andthe yeastproteinPIF)arerepresentedbyboxes,withthelengthof the boxcorresponding to the size of the protein(18,19, 23). The six conserved motifsarerepresentedbydarkbars. The motifsarenumberedasindicated,and theorder ofthe motifs in each protein is identical.
the HSV genome is large (152 kb), it is not convenient or
straightforward to introduce a large number of mutations
directly into it. Therefore, we establisheda transient assay
systemtoanalyze thefunctionofmutantUL5geneproducts
expressed from a cloned copy of the gene. Our results
indicate thatall of the six conserved motifsareessential to thefunctionof the UL5geneproduct in HSV DNA
replica-tion, thusproviding directevidence for the functional
signif-icance ofthese motifs. Three motif mutations were
intro-duced into the viral genome, and phenotypic analysis of
these viralmutantsconfirms the importanceof these motifs in UL5 function. Furthermore, the ability ofmutant UL5
proteins to form specific interactions with UL8 and UL52
indicates that the functional domains required for helicase
activityareseparable fromdomains responsible for
protein-proteininteraction. It is anticipated that this type of
struc-ture-function analysis will lead to the identification of not
only protein domains that contributetotheenzymatic
activ-ities ofhelicasebut alsodomainsinvolvedinprotein-protein
interactions.
MATERIALSANDMETHODS
Cells and viruses. African green monkey kidney cells
(Vero; American Type Culture Collection, Rockville, Md.)
were propagated and maintained as described previously
(45). The KOS strain of HSV-1 was usedas the wild-type
virus. ts mutants of strain KOS, tsK13, and tsM19 were
provided by P. A. Schaffer (Dana-FarberCancer Institute,
Boston, Mass.). ThelacZ insertionmutanthr99, which does
notsynthesize the UL5geneproduct, and the cell line L2-5,
whichsupportsthe growth of UL5mutants, aredescribedin
theaccompanyingreport (50).
Plasmids. plOO-1 containing
oris
on a 100-bp MspIfrag-ment was kindly provided by M. D. Challberg, National Institutes of Health, Bethesda, Md. (36). Plasmid pDG2
contains theupstream regulatory element of the ICP6gene
on a549-bp fragment in the vectorBluescribe (Stratagene, San Diego, Calif.) (see Fig. 1 in the accompanying report
[50]). ThevectorBluescribe containsanoriginofreplication
from the phage M13 and thus can be packaged as
single-stranded DNA in the presence of helper phage, R408 (38).
Plasmid pCW8 containing an XbaI-to-KpnI fragment
(se-quencecoordinates 10636 to 16269) encompassing the UL5
gene (12487C to 15133) was kindly provided by M. D.
Challberg. p6UL5, in which the UL5 openreading frame is
placed under the control oftheupstreamregulatory region of
theICP6gene,isdescribed-inthe accompanyingreport(50).
Recombinant
plasmids
werepropagated
in E. coliDH5aF'
by
standardprocedures
(30).
Site-directed
mutagenesis. Single-stranded
DNA fromp6UL5
wasgenerated
following
infection with thehelper
phage
R408(1).
Site-directed mutagenesiswasperformedonsingle-stranded
DNAwith theT7-GENinvitromutagenesis
kit as instructed
by
the manufacturer(United
StatesBio-chemical
Corp.,
Cleveland, Ohio). Mutagenic
oligonucleo-tideswere
synthesized
by using
aCyclone
DNAsynthesizer
(Biosearch
Inc., Burlington, Mass.) (Table 1).
Inadditiontoincorporating
the desired basesubstitution,
manyoligonu-cleotides also contain silent mutations which generate
re-striction enzyme site polymorphisms; thus, the restriction
maps ofthe mutant genes can be
distinguished
from wildtype. All of the mutationswere verified
by
DNA sequenceanalysis using Sequenase (United
States Biochemical) and thesupplier's
instructions.Transient
replication complementation (TRC)
assay. Veroor L2-5 cells were transfected with the
oris-containing
plasmid plOO-1 by
itselfor cotransfected with wild-type or mutantp6UL5 by
using a modification of the standardcalcium
phosphate coprecipitation
procedure(21).
A mix-ture ofDNAsincluding
0.2 jigofplOO-1,
1.0 jigofp6UL5
(where indicated),
and 5,ug
ofsonicated salmonspermDNA inN-2-hydroxyethylpiperazine-N'-2-ethanesulfonic
acid(HEPES)-buffered
saline (pH 7.05) was precipitated in thepresenceof 125 mM
CaCl2
for 20minatroomtemperature.Rapidly
growingVeroorL2-5 cells(1.5 x 106)werepelleted
by low-speed centrifugationand resuspended in 600,ul of the
precipitated DNA mixture, and the cell suspension was
incubated in a shaking incubator at37°C for 30 min.
Com-plete
mediumwasadded,and thecellsweretransferredtoa60-mm tissue culture dish at 37°C. Four hours later, the medium was removed and the dish waswashed oncewith
prewarmed phosphate-buffered saline(PBS), incubated in 2
ml ofHEPES-buffered saline with 15%glycerolat37°C for 2
min,
and washed twice with PBS before the dish wasreplenished with complete medium. At 24 h
posttransfec-tion,
the mediumwaschanged. At 30 hposttransfection, thecellswere superinfected with KOS orhr99ata
multiplicity
ofinfectionof 10 PFUpercellandincubatedat34°C. At 48 hposttransfection, cellswereharvested and total DNAwas
isolated as described previously (46). Two micrograms of DNA was analyzed by digestion with EcoRI alone or in
combination with DpnI and then subjected to agarose
gel
electrophoresis and Southern blot hybridization. The blot
was probed with Bluescribe DNA labeled with 32P as
de-scribedpreviously
(13).
MOTIF
UL5
rep
uvrD
recB
recD
PIF
1,:.. :.-.:
", '.-, i'm""M... ... --. :::m
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.135.482.72.190.2]TABLE 1. Sequences of mutagenic oligonucleotides
Mutation Oligonucleotides(5'-3')' Newrestrictionsite
G-102 to V GCACGTGCTCTTTACGGATCCAGCGTTGCC BamHI
K-103 to A TGTCTGCACGCACGTGCTCGCTCCGGATCCAGCGTTGCC BamHI
ReversionA-103 to K CGCACGTGCTCTTTCCGGAGCCAGCGTTGC Loss of BamHIsite
D-249E-250 to AA CCCAAGGAGGCCGGCAGCAGCGATGACGATb NaeI
G-290toS TCGGCGAGCTCACACACAC None
R-345to K GTTACCGAATTCGTGCTCGACGCACTTTTTGTTGTT EcoRI
R-416 to K CTCCCCCTCTTTCGTCACCTTC None
E-579to A GCCGTTAGTGCTCCCATGC None
E-757E-758to AA AGGGCAGGGGCGCCGCTGCCAGTAACT Narl
T-809toI CGCGTGATGATCATGGCAA None
G-815toA GTCCAGGCTGAGGGCCTGGGAGC AlwNI
Y-836 to A TGGCCACGGCCGCGCTGT None
a Desired mutationsarerepresentedbydouble underlining,andsilent changestocreatenewrestrictionenzymesites areindicatedby singleunderlining.
bThreeofthreeisolatescontainingthedesired DE-to-AAmutation asdeterminedby DNAsequencingdo not contain thecleavagesite forNaeIasexpected;
instead,they contain thewild-typesequence at this postion.
Immunoprecipitation and immunoblot analysis. To detect
the UL5 protein synthesized from the expression clone
p6UL5, immunoprecipitation of[35S]methionine-labeled
ex-tracts was performed as described in the accompanying
report (50). To detect UL5 protein and protein-protein
interactions in cells infected with KOS or viral mutants, a combination ofimmunoprecipitation followed by immuno-blotanalysiswasperformedasdescribedin the
accompany-ingreport (50).
Isolation of UL5 motif viral mutants. Infectious hr99 ge-nomicDNA waspreparedasdescribedpreviously(48). HSV
mutants containing motif mutations were recovered by
cotransfecting L2-5 cells with infectious hr99 DNA and
mutant versions of p6UL5. Marker transfer experiments
were performed as described previously (16) except that
whiteplaques were picked and purified from abackgroundof
blue plaques.
Southern blot analysis of viral DNA. Viral DNA was
recovered and analyzed as described in the accompanying
report (50).
Analysis of viral DNA synthesis.Theability ofmutants to
induceviralDNAsynthesiswasanalyzedasdescribed in the
accompanying report (50).
RESULTS
Developmentofafunctionalassayfor UL5 gene mutations.
The functional
significance
of the six conserved putative helicase motifs wasinvestigated
byintroducing
mutationsintothe most conserved amino acid residues in themotifs.
Because ofthelarge size(152
kb)
oftheHSVgenome, it isnot convenient or straightforward to introduce a series of site-directed mutations intothe viral genome. To facilitate theanalysis of the effects ofthese
engineered mutations,
wedeveloped
anassayto testthefunction of various UL5gene mutationsexpressed fromplasmids.
Theassaywasbasedonthe demonstration thata
plasmid bearing
anHSVorigin
of replication(oris
or oriL) can beamplified
in a transient transfectionexperiment
ifall ofthe necessarytrans-acting
functions areprovided.
trans-acting
replication
functions canbe supplied bysuperinfection
with HSV(40,
41,
46)
orby cotransfectionwitha setofsevenclonedHSVgenes
(4,
22, 47). Weshowedthatinfectionwithts mutantscontaining
mutations intheUL5 gene(tsK13
ortsM19)
failstosupport thereplication
oforigin-containing plasmids
at the nonper-missive temperature and that the defect can becomple-mented by the presence ofa
plasmid
expressing
thewild-typeversionoftheUL5geneproduct (data not shown). This
assaywillbe referredto asthe TRC assay.
The TRC assay was optimized with respect to the virus
providing trans-actingreplication factors and the clone
pro-viding functional UL5protein.Tocircumventpossible
inter-ference or complications due to the presence of a ts gene product in the TRC assay, a UL5 insertion mutant, hr99,
which fails to synthesize UL5 protein (50) was used. To
provide a functional UL5 gene product, p6UL5, in which UL5isexpressed fromtheICP6promoter, was used. In the accompanyingreport(50), we show that the levelof expres-sion fromp6UL5 isatleast 10-foldhigherthanfrompCW8, inwhichUL5is expressed fromits ownpromoter. Thus, the use of p6UL5 avoids potential problems associated with inefficient expression of functional UL5 in the TRC assay. Furthermore, the ICP6 promoter is inducible by
superinfec-tion with hr99 (49, 50). Thus, in the TRC assay, all the
trans-acting factors neededforDNAsynthesisexcept UL5 areprovided by hr99, andfunctional UL5 isprovided inan
inducible mannerby p6UL5.
AtypicalTRC assayis showninFig.2. VerocellsorL2-5 cells, containingthewild-type version ofthe UL5 gene,were
transfectedwith
oris-containing
pl00-1
aloneorin combina-tion with the UL5 expression clone p6UL5. At 30 h post-transfection, cellsweresuperinfected
with KOS orhr99. At 40 hposttransfection,
total DNAwasharvestedanddigested
with EcoRI alone (Fig. 2, lanes a) or in combination with
DpnI (lanes b). EcoRI cleaves
pl00-1
once,generating
afragment of2.8 kb(labeled
pl00-1
inFig.
2).DpnI
recognizes
only cleavage sites whichhavebeenmethylated by
thedam methylation system ofE. coli. Thus,plasmid
DNA which hasbeenpropagatedin E.coli is sensitivetoDpnI (lane
lb),
whereas DNA which has replicated in mammalian
cells
isDpnI
resistant (lanes2b, 4b,
and5b).
The presence ofDpnI-resistant
bands indicates that theplasmid
DNA hasreplicated
in Vero cells. When Vero cells weretransfected withpl00-1
andsuperinfected
withKOS,
aDpnI-resistant
band of 2.8 kb,
representing
linearizedpl00-1,
wasob-served,
indicating
that KOS can support thereplication
of thisorigin-containing plasmid (lane
2b).
The smallerDpnI
digestion
fragments
(lanes
b)
representinput
plasmid
andserve as internal controls for the
efficiency
oftransfectionand recovery. The absenceofthe2.8-kb
DpnI-resistant
bandin cellstransfected with
pl00-1
andsuperinfected
with hr99indicates that hr99cannot support the
replication
ofpl00-1
(lane 3b). However,ifafunctionalUL5
product
issupplied
on November 10, 2019 by guest
http://jvi.asm.org/
472 ZHU AND WELLER
1 2 3 4 5
a b a b a b a b a b
I
_ o p6UL5a NoonNow .4 b m-._---
pl00-i
.;
_m
em
DpnI sensitive bands
FIG. 2. TRC assay. The TRC assay was performed in either Vero orL2-5cellsasdescribed inMaterials and Methods. At48 h after transfection, total cellular DNA was isolated. DNA was digested with eitherEcoRI alone (lanes a)orEcoRI plus DpnI (lanes b); thus, every sample is represented byapairof lanes (a and b). Lane1contains5ngofpl00-1containing HSV-1orispropagatedin E.coli, andlanes 2 to 5 contain2,ugof total cellular DNA harvested from theTRC assay as follows: lane 2, transfection withpl00-1and superinfection with KOS; lane 3, transfection with pl00-1 and superinfection with hr99; lane 4, transfection with pl00-1 and p6UL5 and superinfection with hr99; lane 5, transfection of L2-5 cells with pl00-1 and superinfection with hr99. After restriction enzymedigestion, the samples were subjected to 0.8% agarose gel electrophoresis and blottedonto aGeneScreen Plus membrane.The blot was hybridized with 32P-labeled Bluescribe plasmid DNA. Positions of the linearized pl00-1 and the 6.3-kb fragment from p6UL5aremarked. Smallfragments resulting from Dpnl digestion arealso marked.
either by cotransfection with the UL5 expression clone
p6UL5 or by transfection of L2-5 cells which contain a
wild-type version of the UL5 gene, the defect in hr99 is complemented (lanes 4b and Sb, respectively). The 6.8-kb
band seen with EcoRI digestion (lane 4a) represents input
p6UL5 plasmid. This plasmid lacks an HSV origin and therefore cannot replicate in Vero cells and is sensitive to
DpnI digestion (lane 4b). In L2-5 cells, the p6UL5 band is
DpnI resistant because it is integrated and replicates with
cellularDNA(lane Sb). By usingthisassay,the ability of the
UL5protein expressedfrom the expressionplasmidp6UL5
to complementthe replication defect in hr99 canbetested.
This assay provides a convenient method for testing the
function of ULS protein expressed from plasmids bearing
wild-type and mutantversions of the ULS gene.
ULS contains six conserved helicase motifs. Sequence
anal-ysis of the ULS gene reveals considerable homology with
members ofa superfamilyof helicases of E. coli,
Saccharo-myces cerevisiae, and RNA and DNA viruses (Fig. 1) (17-19, 23). The presence ofsix well-conserved motifs and
the similarity of their positions relative to one anotherare
noteworthy (Fig. 1). In this report, the functional
signifi-canceof these six conserved motifs in UL5wasexaminedby
the introduction of engineered mutations into the most
conserved residue(s) in each motif (Fig. 3).
Motifs I and II. Oligonucleotide mutagenesis was
per-formedonsingle-stranded DNA from p6UL5asdescribedin
Materials and Methods; oligonucleotides used to generate
each mutation are listed in Table 1. The first target for
mutagenesis was motif I (also called the A site)containinga
sequence (G/AX4GKS/T) which is highly conserved among
most enzymes utilizing ATP and GTP (43). This motif
containsaflexibleloop believedtobeinvolved in thebinding
of the pyrophosphate moiety of nucleoside triphosphates
(NTPs). Two invariant residues weretargeted for
mutagen-esis; the first mutation resulted in an alteration of G-102 toa
0 100 200 300 400 500 600 700 800 900
a . .1 a . I
11 III IV
4 4 R44
R to
II III
V VI
4
E to A
IV
4 44
EE to AA
v VI
UL5 94 ITGNAGSGKSTCVQ 136 VIVIDEAGLLG 26 LVCVGSPTQTAS 44 NNKRCVEHE 442 AMTITR SQGL SLDKVAICF 8 SAYVAMSRT uv?D 26 VLAGAGSGKTRVLV 174 NILVDEFQNTN 16 VMIVGDDDQSIY 26 QNYRSTSNI 267 LMTLHSAKGL EFPQVFIVG 23 LAYVGVYRA
cap 19 VLAGAGSGKTRVIT 175 YLLVDEYQDTN 16 FTVVGDDDQSIY 26 QNYRSSGRI 271 LMTLHA SKGLEFPYVYMVG 22 LAYVGITRA
zBcS 20 IEASAGTGKTFTIA 345 VAMIDEFQDTD 18 LLLIGDPKQAIY 24 TNWRSAPGM286 IVTIHK SKGL EYPLVWLPF 44 LLYVALTRS rSaD 164 ISGGPGTGKTTTVA 82VLVVDEASMID 16 VIFLGDRDOLAS 24 QLSRLTGTH 198AMTVHK SOGS EFDHAALIT 11 LVYTAVTRA
PIF 255 YTGSAGTGKSILLR 46 ALVVDEIS4LD 25 LIFCGDFFQLPP 29 KVFRQRGDV 219MQTIHQNSAGKRRLPLVRFKA 33 QAYVALSRA
IDENTICAL RESIDUES
TARGETED
ISXIDUES
GK DE
GK MUTATED TO VA
DE AA
G 0 T G
G T G
s I A
Y R
y
A
FIG. 3. Site-directedmutagenesisof theUL5gene.The UL5 proteinis represented byashadowedbox,andthesix conserved motifsare
representedby dark bars.Arrows underthe UL5 protein indicate the positionsof site-directed mutations.Thesubstitutedaminoacids in three mutations in nonconserved regionsaremarked witharrows.Theaminoacidsequencesof the sixconservedmotifsfrom six proteins(UL5,
Rep, UvrD,RecB, RecD, and PIF)areshown (18, 19, 23); the numbersbetween the amino acidsequencesrefertothe number of the residues
separatingeachmotif.Themostconservedresidues withineachmotifarelistedbelow the amino acidsequences.Conservedresidueswere
subjected tomutagenesis, and the amino acid substitutionsareshown.
POSITIONS Or MOTWS
POSITIONS OF MUTATIONS
MOTIF SEQUE= I
4
J.VIROL.
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.91.264.72.267.2] [image:4.612.112.504.450.670.2]Motif I controlp6UL G102V K103A
a ba b a b a b
Motif 11 A103KDE250AA
a b a b
_p6UL5 --
-_ pl00-1 **_
,S
[image:5.612.100.285.78.254.2]£ :
FIG. 4. TRCassayofUL5 proteinswith site-directed mutations inmotifs I and II. TheTRC assaywasperformed asdescribedin Materials and Methods and the legendto Fig. 2. In each sample, plOO-i was used in transfection either alone (lane control) or in
combinationwith p6UL5containingwild-typeUL5(lanep6UL5)or
variousmutant constructsofp6UL5asindicated.Transfected cells weresuperinfected with hr99.
valine residue (G102V), and the second resulted in an
alteration of K-103 toalanine(K103A) (Fig. 3). Both
muta-genicoligonucleotides alsogenerateanovelBamHIsite, and
theintroductions of both the targeted motif mutation and the
novel restriction site into p6UL5 were confirmed by DNA
sequencing (data not shown). The motif I mutations were
testedfor theirabilitytocomplement hr99 in the TRCassay
as described above. As shown in Fig. 4, wild-type UL5
(p6UL5) can complement hr99 in its ability to support the
replication of plOO-1 in Vero cells (lane p6UL5); however,
mutants G102V and K103A failtodo so (lanes G102V and
K103A, respectively). The transfection efficiencies of both
plOO-1 and p6UL5 in each assay are roughly similar, as
indicated by the presenceof the small DpnI-sensitive
frag-ments (lanes b); as described above, these bands represent
input DNA. The loss of UL5 function inG102VandK103A
demonstrates theimportance of conserved motifI.
Further-more, areversion of the K103A mutation backtowildtype
(A103K) results in the recovery of UL5 function (lane A103K), confirming that the observed loss offunction of
K103A is duetothe targeted mutation atposition 103.
Thesecondregion targeted for mutagenesiswasmotifII,
aregion containing one ortwo negatively charged residues
(D or DE) at the COOH terminus of a p strand; the
negatively charged residue(s) is believedtointeract with the
Mg2+
associated with the0
or -y phosphate of a purinenucleotide substrate(motif II) (14, 26). Toassessthe
signif-icance of this motif, we replaced both the aspartic acid at
residue 249 and the glutamic acid at residue 250 with two
alanine residues (Fig. 3). The plasmid carrying mutant
DE250AAwasunabletocomplement hr99inthereplication
assay (Fig. 4), indicating that the presence of negatively
charged aminoacids within this motif is indeed essential.
MotifsIll to VI. In additiontothe well-known motifsIand
II, four other motifs are shared among over 20 proteins
which have known or putative helicase activity (19, 23).
However,unlike motifs I and II,the functional significance
ofmotifs IIItoVI has notbeenpreviously addressed either
genetically or biochemically. To determine whether these
MotifIII controlp6UL5 G290S a b a b a b
0 -p6UL5
-Motif IV MotifV
R345K T8091 G815A
a b a b a b
MotifVI Y836A
a b
am to p6UL5p6UL-...
a.
4*@
-p100-i -_ 0. - p100-i1-1_._ a a
la 4
FIG. 5. TRC assay ofUL5 proteins with site-directed mutations inmotif II, IV, V, and VI. For details, see the legend to Fig. 4. Pairs ofsamplesaremarkedwith the names of thep6UL5 constructs used intransient transfections.
four motifs are essential for UL5 function, we introduced
alterations into each conserved region (Fig. 3). Motif III
contains an invariant glycine at position 290 which was
replaced with a serine (G290S). A conservative alteration replacing the invariantarginine residue at position345with lysine(R345K) wasintroduced intomotif IV. An EcoRI site wasintroduced into p6UL5 together with the R345K muta-tion, and as expected, the band representing p6UL5 is
smaller insize(Fig. 5). Twomutationswereintroducedinto
motifV,onereplacingthreonine 809 with isoleucine (T8091)
and the other replacing glycine 815 with alanine (G815A).
Eachof the mutations in motifs III, IV, and V resulted in
failure to complement hr99 inthe replicationassay (Fig.5).
Motif VI can be divided into two components, one well
conservedamong RNA and DNA helicases of this
superfam-ily and the other a single tyrosine found in known DNA
helicases butabsentinRNAhelicases(23).Atyrosinein this positionisfoundin E. colienzymesUvrD, Rep,RecB, and RecD,in theyeastenzymePIF,and in theUL5 counterparts
of the herpesviruses Epstein-Barr virus, human
cytomega-lovirus, varicella-zoster virus, and HSV. To assess its
im-portance, wechangedthe
tyrosine
atposition
836 ofUL5toan alanine (T836A). Mutation T836A completely abolishes
the ability of the plasmid to complement hr99 for UL5
function(Fig. 5, laneY836A).
Nonconservedregionsof UL5. Mutations in nonconserved
regions of UL5 were introduced todetermine whether the UL5gene product can tolerate similarchanges outside the putative functional motifs (Fig.
3). Thus,
an arginine-to-lysine mutationatposition416(R416K),aglutamic
acid-to-alanine substitution at
position
579(E579A),
and a doublereplacementof twoglutamicacid residuesat757 and 758to
twoalanine residues (EE758AA)
(Fig.
3 and Table2)
weretested as described above. All three mutant
plasmids
wereabletocomplementhr99aswellasdidthe wildtype
(Fig. 6).
This result indicates that the UL5
protein
can toleratechangesoutside the conserved motifs.
Detection ofmutantUL5 geneproducts.The loss of
func-tion of mutations in the six conserved motifs of UL5 describedabovecould in
principle
be due eithertochanges
inglobal
protein
conformation or tochanges
in localized. 4mmm
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.328.565.78.257.2]474 ZHU AND WELLER
TABLE 2. Plaquingefficiencies ofKOS,hr99,hr99G201V, hr99K103A,andhr99R345Kon Vero and L2-5cellsa
Virus ~~~Cell Titer
Virus
Cline
(PFU/ml)KOS Vero 5 x 109
L2-5 5 x 109
hr99 Vero <1 x 103
L2-5 5 x 108
hr99G102V Vero <1 x 103
L2-5 5 x 108
hr99K103A Vero <1 x 103
L2-5 3 x 108
hr99R345K Vero <1 x 103
L2-5 6 x 108
aPlaquing efficiencieswere measuredby determining the titers of virus stocks on monolayers of Vero or L2-5 cellsasindicated.
residues essential for function. Changes in global protein conformation often affectprotein stability, resultingin deg-radation (37). Todemonstrate that theloss-of-function
mu-tations still encode a stable UL5 protein, UL5 expression
frommutantconstructs wasexamined in transient
transfec-tion experiments. Cells were transfected with wild-type p6UL5 or mutantversions of theplasmidandsuperinfected withhr99. Cells were labeled with
[35S]methionine,
and cellextracts were immunoprecipitated with anti-UL5 antiserum
(a-UL5)
asdescribed in Materials andMethods. Cells trans-fected with salmonsperm DNAaloneandsuperinfectedwith hr99 fail to synthesize a protein ofthe expected molecularsize,
99 kDa(Fig. 7, leftmost lane). When cellswere trans-fected withp6UL5 andsuperinfected withhr99,aprotein of 99 kDawasspecifically precipitated by a-UL5(Fig. 7, lane Wild type). In the absence of superinfection, cells trans-fected withp6UL5 failtosynthesize detectable amounts of the UL5 gene product (data not shown). Thus, consistentwithresults presented in the accompanying report(50), hr99
is able to provide trans-inducing factors which stimulate expression of p6UL5. Cells transfectedwith UL5constructs containing mutations in motifs I through VI produce UL5 protein recognized by a-UL5 in amounts similar to those produced by the wild type after superinfection with hr99
control p6UL5 R41 6K
a b a b a b
E579A EE758AA
a b a b
-p6UL5 -d Uw
__ "" - p100-i-S
FIG. 6. TRCassayofUL5 proteins with site-directedmutations inthe nonconserved regions. For details, seethe legendto Fig.4.
Pairsofsamplesaremarked with thenamesoftheconstructsused intransienttransfections.
0 c
200-kDa
-97.4-kDa
-69-kDa
30-kDa
21.5-kDa
FIG. 7. Detectionof UL5 proteins in transient transfection by
immunoprecipitation with a-UL5. Equalamounts(12 ,ug) ofp6UL5
with mutations thatabolished the UL5 function(asmarked) were usedtotransfect Verocells, and the cellswerethensuperinfected
withhr99. Inthe leftmostlane, 12 ,ugof salmon spermDNA was
used.Transfected cellswerelabeled andprocessed for immunopre-cipitationasdescribed in theaccompanyingreport(50). Positionsof
'4C-labeled
rainbowproteinmolecularweightmarkers(Amersham,Arlington Heights,Ill.)electrophoresed in parallelareshownonthe left.
(Fig. 7). All mutantproteins showed electrophoretic mobil-ities identicaltothat of thewild-type protein. Thus, although these mutations abolish the ability ofUL5 to complement hr99 in the replication assay, the stabilities ofthe mutant UL5geneproductsarecomparable tothatofthe wildtype.
This result suggests that the loss of function seen in the
mutant constructs ismost likelydue to changes in residues
involved in UL5 function rather than disruptions which
affect the three-dimensional structure and destabilize the
protein.
Isolation of viral mutants with UL5 motif mutations. The
transient replication complementation test has provided a convenientway ofassessing the effects of motif mutations onthefunction ofUL5proteinin viral DNA replication.To confirmthatthis transientassayindeed serves as a reason-able facsimilefor viralinfection,weintroducedseveralmotif mutations intotheviralgenome and tested their effects inthe
contextofviral infection.Theisolationof mutants withUL5
motifmutations was facilitated by the
availability
of theUL5null mutanthr99 (50).hr99contains an ICP6::IacZinsertion
nearthe Nterminus ofthe UL5 gene and forms blueplaques
on the UL5 permissive cell line L2-5. Viruses containing motifmutations wererecoveredfollowing cotransfection of L2-5 cells withinfectious hr99DNA and plasmidscarrying motif mutations(see Materials and Methods). Homologous J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
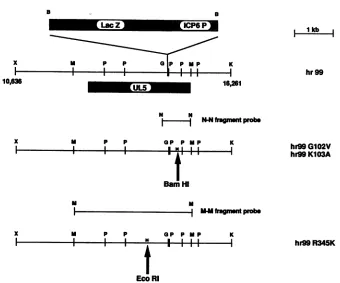
[image:6.612.362.502.75.357.2] [image:6.612.54.294.99.215.2] [image:6.612.81.265.514.681.2]B B
LacZ icp6P
M P p a P PMP
I I I II I
UmL5
K 16,261
N
NN-N
fagnmntprobe
x
F-p p GP p MP
I I I B H1l
Barn Hi
x
K
hr99GI02V hr99K103A
p p OP P NP K
I l * l l l l |hr99R345K
[image:7.612.148.488.67.350.2]EcoRI
FIG. 8. Genome structures ofmutants hr99, hr99GlO2V, hr99K1O3A, andhr99R345K. Diagrams showing the mutations in each viral mutantareshown inrelationtoanXbaI-to-KpnI fragment(sequencepositions10636to16261)whichspansthe UL5openreading frame. The
UL5openreading frame is shownas ahatched box withan arrowindicating the direction of translation within the HSVgenome.The positions
ofsite-directed mutations and thenewrestrictionsites generated concurrently in hr99GlO2V, hr99K103A,andhr99R345Karemarked by
arrows.Thepositions of DNA probes for Southern blot hybridizationare shown; the N-N probe and the M-M probeareDNAfragments
generated by NaeI and MluI digestion, respectively. Restrictionenzymerecognition sites: B, BamHI; G, BglII; K, KpnI; M, MluI; P, PstI;
X,XbaI.
recombination between hr99 DNA and the UL5 sequences
results inviralgenomes which have lost the lacZgene and
thereforeform white plaques in the presenceof
5-bromo-4-chloro-3-indolyl-,-D-galactopyranoside (X-Gal) (15). Since
new restriction sites were introduced by silent nucleotide
changes at positions next to many of the motif mutations
(Table 1), thepresenceof the mutations could be determined
by restriction analysis of viral DNA from the white plaque
isolates. We have introduced two mutations in motif I
(G102Vand K103A)and onemutation in motif IV (R345K)
into viral genomes, resulting in the isolation of mutants
hr99G102V, hr99K103A, andhr99R345K, respectively (Fig.
8). In each case, over 50% of the white plaque isolates
contained the desiredmutation,asdeterminedby restriction
enzyme analysis. KOS, hr99, and the three motifmutant
DNAs were analyzed by restriction enzyme digestion and
Southern blothybridization (Fig. 9). DNAsfromhr99G102V
and hr99K103A would be expected to contain a novel
BamHIsite,and hr99R345K would beexpectedtocontaina
novel EcoRIsite,asdiagrammedinFig.8. InFig. 9A, KOS,
hr99,hr99G102V, and hr99K103A DNAsweredigestedwith
BamHI andprobedwith the N-Nprobe (anNaeIfragment;
Fig. 8). Wild-type KOS contains theexpected 9.8-kb band
(Fig. 9, lane 1). In hr99, the size of the corresponding
fragment increasestoapproximately 14 kbas aresult of the
ICP6::lacZ insertion (lane 2). In both hr99G102V and
hr99K103A, the new BamHI sites are clearly present, as
indicated by the disappearance of the 14-kb band and the
appearanceof3.0-and 6.8-kb bands (lanes3 and4,
respec-A
1 2 3 4
14kb\ 9.8 kb
B
5 6 7
16.7kb
- w w -6.8 kb
_*
m_-
2.2kb- 1.5 kb
FIG. 9. Southern blotanalysisof viral DNA from KOSandthree
UL5mutants.(A)Viral DNAsweredigestedwithBamHI,andthe
blotwashybridizedwith the N-Nprobe (see Fig. 8).Lanes:1,KOS; 2, hr99; 3,hr99G102V; 4, hr99K103A. (B)ViralDNAsweredigested with EcoRI andprobedwith the M-Mprobe (see Fig. 8).Lanes:5, KOS; 6, hr99; 7,hr99R345K.Sizes of thefragmentsareasmarked.
x 10,631B
i I i m I i - - hr 99
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.327.565.485.666.2]476 ZHU AND WELLER
L2-5* * hr99R345K
Vero hr99GlO2V L2-5 * 0
VerO L2-5 hr99Kl 03A
Vero L2-5 hr99
Vero L2-5 KOS
Vero L2-5 Mock
Vero
e
q
0
e
*
0
0
FIG. 10. Analysis of viral DNA synthesis by KOS and four viral mutants. Vero or L2-5cells wereinfected with the indicated virusat amultiplicity of infection of 5 PFU per cell for 18 h at 37°C.Aseries of fivefold dilutions of each cell suspension was spotted onto a GeneScreen Plus membrane. Cells were lysed on themembrane, and the membrane washybridized with 32P-labeledEcoRI F frag-ment asdescribed in the accompanying report (50).
tively). The intensities of the 3.0- and 6.8-kb bands differ
depending on the position of each fragment with respect to
the probe. In Fig. 9B, KOS, hr99, and hr99R345K DNAs
were digested with EcoRI and probed withthe M-M probe
(an MIuI fragment; Fig. 8). As predicted, KOS contains a 16.7-kb band (lane 5). The 2.2- and 15.9-kb bands seen in hr99 DNA (lane 6) are expected and are due to internal EcoRIsites within the ICP6::lacZinsert itself. The presence
ofanew EcoRI site in hr99R345K (lane7) is confirmed by
the appearance of1.5- and 15.2-kb bands. The large
frag-ments in lanes 6 and 7 are very faint because of the small
regionof homology with the M-M probe.
Phenotypic analysis of motif mutants hr99G102V, hr99K
103A, and hr99R345K. Mutants hr99G102V, hr99K103A,
and hr99R345K were propagated on L2-5 cells, and the resulting stocks were titered on L2-5 and Vero cells. As
shown in Table 2, these mutants could form plaques
effi-ciently in L2-5cells; however, all three mutants are unable
toformplaques on Vero cells. Thus, these motif mutations
severelyimpair the normal growth on Vero cells, suggesting
that the mutations affect UL5 functions essential for virus
growth.The abilityof mutantshr99G102V,hr99K103A,and
hr99R345K to synthesize viral DNA was analyzed by dot blot hybridization to a representative HSV DNA probe (32P-labeled HSV-1 strain KOS EcoRI F fragment) as de-scribed in Materials and Methods. Figure 10 shows that all
three mutants were unable tosynthesizeviral DNA in Vero
cells,whereas theinabilitytosynthesizeviral DNAwasfully complementedtowild-typelevelsby growingin the permis-sive cell line L2-5.
Motif mutantssynthesize stable UL5 proteinwhichretains
its ability to interact with UL8 and UL52 specifically. As
mentioned above, loss-of-function mutations could be due either to changes in global protein conformation or to changesin localized residues essential for function. There-fore, stability of the mutant UL5 proteins in thecontextof viral infection was examined. Extracts from mutant and
wild-typevirus-infected cellswere immunoprecipitatedwith
a-UL5, and the immunocomplexes were subjected to
so-dium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and immunoblotted with a-UL5 asdescribed inMaterialsandMethods.Figure11Ashowsthatwhile hr99
did not synthesize detectable UL5 protein, all three motif
mutants were capable of synthesizing full-length UL5
pro-teins at levels similar to that of the wild-type virus KOS. Thus,consistent with results of transient transfection exper-iments described above, the mutations do not appear to affectthestability oftheUL5peptide.
As demonstrated in the accompanying report (50), the
interactionbetween UL5 and othermembers ofthe helicase-primase complex, UL8 and UL52, can bedemonstrated in KOS-infected cells by coimmunoprecipitation. We exam-ined whetherUL5peptideswithmutations in motifs I and IV arecapable of formingspecific immunocomplexeswithUL8
and UL52. Extracts from Vero cells infected with KOSor
UL5motifmutantshr99G102V, hr99K103A,andhr99R345K were immunoprecipitated with cx-UL5 as described above. Immunocomplexes were subjected to SDS-PAGE and
im-A
-LO cm asO n
0) 0)0 o o
.~
-180-kDa
ULU
-t 16-kDa -84-kDa -58-kDa
IgG_ _ _ _
36-kDa
_ Si* ~ ..
B
i,e
~e
> enL:Ocm Y
0
0 0) 0) a) 0) C) o
00 0 )Y
-180-kDa
52 A - -116-kDa
-84-kDa
58-kDa
_-36-kDa
C Y>UZC CM cr)
et 0
CO) _r _r
cc O Y Y
c0) 0) 0) )
CD CY)0)2 0 0
4 J.. -15.3.','80-kDa
-116-kDa
UL8-_ 848kDa
58kDa
,
A~~~~~3-D
FIG. 11. Interactions of UL5 proteins from KOS and four viral mutants with UL8 and UL52 as detected by coimmunoprecipitation. Vero cellsweremockinfected or infected with the indicated virus at a multiplicity of infection of 10 PFU/ml for 18 h at 34°C. Cells were harvested, andextracts were subjected to a combination of immunoprecipitation and immunoblotting as described in the accompanying report (50). Extracts were immunoprecipitated with a-UL5, and immunocomplexes were divided into three aliquots and subjectedto SDS-PAGE in parallel. Each blot was probed with either a-UL5 (A), a-UL8 (B), or a-UL52 (C). Positions of the UL5 and immunoglobulin G (IgG) proteins aremarked in panel A; positions of the UL8 and UL52 proteins are marked in panels B and C, respectively. Positions of molecular weight markersareindicated.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.612.107.246.76.284.2] [image:8.612.110.502.530.656.2]munoblotted with a-UL8 or a-UL52 as described in the
accompanying
report(50).
Figures
liB
and C demonstrate that mutantpeptides specified by
hr99G102V, hr99K103A, andhr99R345K retain theability
tointeract with both UL8 and UL52. InFig.
liB,
cells infected withthe three motif mutantshr99G102V,
hr99K103A,
and hr99R345Kcontain a smaller band in additiontotheexpected UL52gene product. This band is not present in KOS-infected cells and is notreproducibly
seeninmotifmutant-infectedcells. Its origin is notclear.Thus,
the behavior of the motif mutants with respect to viral DNAsynthesis
andstability
of mutant peptides is consistent with the results obtained in transient assays. Theseresults thus confirm thevalidity
of the TRCassayfortesting
thefunction of UL5 in viral DNAsynthesis.DISCUSSION
The functional
dissection
ofgenesby
site-directedmuta-genesis
hasprovided
manyinsights
into theevolutionary
relationship
and domainorganization
ofmanyprotein
mol-ecules. Inthisstudy,
weinitiateda structure-functionanal-ysis
of the UL5protein.
The presence of six conserved motifs in UL5 sharedby
alarge
superfamily
ofhelicases with six conservedmotifsprovided logical
targetsforsite-specific
mutagenesis.
The resultspresented
demonstratethat all six motifsarein fact essential for the function of UL5in DNAreplication.
TRCassay. Structure-function
analysis
involvestheintro-duction ofaseriesof
specific
mutations intoagene.For the HSV system,itisnotalways
convenientandstraightforward
to introduce a
large
number ofspecific
mutationsdirectly
into the viral genome. To facilitate ouranalysis
of UL5protein
mutants, wedeveloped
aconvenientassayforUL5 function intransiently
transfected cells. The TRCassay is based onthe demonstration thataplasmid
bearing
an HSVorigin
ofreplication
can beamplified
ifall necessarytrans-acting
functionsaresupplied (4, 40, 41,
46,
47).
Inourstudy,
all of the
trans-acting
factors except UL5 areprovided by
the null mutant hr99(50),
andwild-type
or mutantversions of UL5 areprovided by
theexpression plasmid p6UL5.
Thus,
theability
ofmutantversions of UL5 tocomplement
hr99 in the
amplification
oforigin-containing plasmids
pro-vides afunctionalassayfor UL5.All six conserved motifs of the UL5 gene are essential.
Protein sequence
analysis
has revealed that the HSV UL5(as
well as the UL5 gene counterparts in otherherpesvi-ruses)
is a member of asuperfamily
ofDNA and RNA helicases which sharea setof six well-conserved motifs(18,
19, 24).
This remarkable conservation of motifs within helicases frombacteria,
yeast, and mammalian cells and viruses suggests that these motifsareimportant
for helicasefunction; however,
theirsignificance
has not beendirectly
demonstrated.Inthisreport,weshowthatmutationsineach of the conserved motifs of
UL5
completely
abolishUL5
function in the TRC assay, whereassimilar
mutations into nonconservedregions
of the UL5 gene have no effect onUL5function. The effect of amino acid substitutionsonUL5
function
could intheory
be due to either achange
in aresidue that is involved in function or a
change
inprotein
conformationpotentially
leading
toinstability
oftheprotein.
We consider the latterpossibility
unlikely
sincewild-type
levels offull-length
UL5protein
havebeen detectedincellstransiently
transfected withplasmids
expressing
loss-of-function mutations inthe UL5 gene(Fig.
7).
Toconfirmthattransient transfection
experiments
reflect
events whichoccur in infected cells, three motif mutations
(G102V, K103A,and R345K) were introduced into the viral genome. Thisallowed us to confirm the validity of the TRC
assay and to analyze single amino acid substitutions inthe
UL5gene in the contextof the viral infection. Introduction
of motif mutations intothe viral genome was facilitated by
the existence of the lacZ insertion mutant hr99, which provides a color screenfor recombinant viruses. The
phe-notypesofmutantshr99G102V,hr99K103A, andhr99R345K
fullysupport the results obtained from the TRC assay inthat
these mutants fail to synthesize viral DNA when grownin
Vero cells, although wild-type levels of mutant proteinsare
present in infected cells. Consistent with their inability to
synthesizeviral DNA, these three mutants also fail to form plaquesonVero cells.
As wehave shown inthe accompanyingreport(50), UL5
proteinexpressed during viral infection can be assayed for its ability to associate with UL8 and UL52 proteins by a combination immunoprecipitation and immunoblot proce-dure. To date we have not been able to observe this association in cells transiently transfected with expression plasmids, presumably because of the sensitivity of our detection methods (51). In this report, we show that UL5 proteins expressed from the threeUL5motifmutantsretain the ability to interact with UL8 and UL52 as
:assayed
bycoimmunoprecipitation.
The ability of these mutant UL5 proteinsto associate specifically with UL8and UL52indi-catesthat the conserved motifsI and IV areunlikely tobe
involved
directly
inthe interaction betweencomponents of thehelicase-primase
complex. In addition, these results indicatethatthetertiarystructureofthemutantproteinshas notbeengrossly altered by themutations. Taken together,ourresults suggestthat the loss ofUL5function exhibited by
the eight mutations in motifs I through VI was due to changes in residues required for UL5 function. These results
provide
thefirstdemonstration ofthefunctional importance of motifsconservedwithin membersof thelarge superfamily of helicases.Possible roles of the six conservedmotifs. The importance
of motifs I and IIin
NTP-binding
proteins has been recog-nized for some time on the basis of (i) nuclear magneticresonanceand X-ray diffraction data(14, 26) and(ii) genetic
analyses
inwhichreplacement
of residues in motifs I and II abolishes the ATPaseactivity
ofNTP-binding
proteins.Interestingly,
mutation of the invariantlysine in motif I ofthe yeast RAD3
protein
toanarginine
residue resulted inaprotein
which could still bind ATP but was defective inhydrolysis
(42).Similarly,
mutations introduced into the invariantglycine
orlysine
of motif I inShe
multidrug resistancegeneofthe mouse(mdrl)resulted in theab'olition ofdrug
resistance; again
these mutants still retained theability
tobindATPbutfailedtohydrolyze
it(2).
Some motif Imutations,
on the otherhand,
appeartoaffectbinding
ofNTPsper se;for
instance,
athreonine-to-serinereplacement
inmotifI oftheHSV
thymidine
kinaseexhibitedanaltered Km forthymidine
and ATP(29).
Thus,
motif I has beenstrongly
implicated inbinding
andhydrolysis
of NTPs in several knownNTP-binding proteins.
The invariantnega-tively
charged
residues in motifII have notreceived much attentiontodate. Our-results for UL5 indicated the absoluterequirement
forthenegatively
charged
residues in motifII,
while two similarmutations(EE
toAA and E toA)
intro-duced intotwononconservedregions-of
theprotein
didnotaffect UL5function.
Little is known about the
biological significance
of the other fourmotifs sharedby
thissuperfamily
of known andon November 10, 2019 by guest
http://jvi.asm.org/
478 ZHU AND WELLER
putative
helicase proteins. Our results demonstrate thatthese four conserved motifs are indeed essential for the
function of UL5. In addition to an NTP-binding fold, one
might expect that a helicase would contain several other functional
regions,
includingonewhich contacts the nucleic acid substrate. Itisinterestingto notethatmotif VI containscertain residues found only in putative DNA helicases but
notin RNAhelicases; for example, thetyrosine in motif VI
ofUL5is sharedby E. coliproteins UvrD, Rep, RecB, and
RecD,theyeasthelicasePIF,and three other herpesviruses, varicella-zostervirus, Epstein-Barr virus, and human
cyto-megalovirus
(18, 19, 24). Our results demonstrate that thistyrosine
is indeed important for UL5 function, since the substitution oftyrosine
836with an alanine residueabolished theactivity
of UL5 in the replication assay. Further bio-chemicalanalysis
of this mutant will be necessary todeter-mine whether DNA binding is affected (see below). An
additional
region
ofsequence conservation located between motifs I and II and designated motif Ta has also been associatedwith presumedDNA but not RNA helicases (17,24,
27).
It is of interest to note that the superfamily described
above does not contain all known helicases. A second
helicase
superfamily
(SF2) which contains proteins from E.coli,
yeast,insect,
andmammalian cells, poxviruses,herpes-viruses,
andpositive-strand
RNAviruseshas been identified(20,
28).Adegree of similarity exists betweenSF1 and SF2,although
there are substantial differences. One member of theSF2family,
theRAD3protein, has been studied through spontaneous and artificial mutations (42). All mutationsimpairing
itsactivityin DNArepairoressential functionsfellexactly
within the conserved motifs I to V (33-35). Thisfinding
strongly demonstrates thefunctional importance of these conserved motifsoftheSF2.Recently, the presence oftheseconserved motifs ina newly identified and sequenced
protein
encoded by the human ERCC-3 gene was taken as evidence for apotential
DNA-unwinding function (44).Al-though
it will be necessary to demonstrate experimentally that theproduct
of the human ERCC-3 gene is indeed ahelicase,
the results provided in this report provide experi-mentalsupportforthenotionthatsuchconservedmotifs aregood predictors
ofregions likely to be importantfor func-tion.The results presented in this report clearly demonstrate
that it will be possible to carry out a detailed structure-functionanalysisof the HSV UL5 gene product. UL5 viral
mutants with single amino acid substitutions have been isolated which are incapable of carrying out viral DNA
synthesis
but arestill able toform specific complexes with other viralproteins.
Thenext step will be to assay mutant viralproteins
for otheractivities in order to definedomainsrequired
forvariousfunctionsrequiredof ahelicase,includ-ing
ATP-binding, ATPase, nonspecific DNA-binding, and helicase activities. Direct biochemical assays have beenhampered
bythelackof aconvenientoverexpressionsystem whichiseasilymanipulatedgenetically.The onlyexpression system for UL5, UL8, and UL52 which has consistentlygenerated
active helicase or primase enzyme is thebaculo-virussystem (3, 11), and this system has not beenconvenient
for genetic analysis. Experiments to express functional helicase or primase subunits in a genetically manipulable
systemareunderway. This type ofstructure-function
anal-ysis
willbeappliednot only tothe UL5 gene but also to theUL8 and UL52genes and should facilitate theassignmentof
function to the individual members of the complex; for
instance,
it should be possibleto determine which protein,UL5 or UL52, has intrinsicprimase
activity
andtomap theprotein domains responsible for this
activity.
Furthermore,
the results presentedin this andinthe
accompanying
report(50) indicate that in addition to
determining
theprotein
domainsresponsiblefor helicase andprimase
activity,
itwill
be possible to map the sites required for
interaction
withother members of the helicase or
primase.
The fact thatinteraction
between
members ofthehelicase-primase
com-plex appear to be essential for enzymatic
activity
and thefact that all threesubunitsare
required
in vivoforviral DNAreplication suggest thatdomains of interaction may
provide
useful targetsfornovel antiviral drugs. Aprecedent for this
strategy comes from thefinding thatapeptide derived from
the large subunit ofHSV ribonucleotide reductase inhibits
reductase activity, evidently by interference with subunit
interactions (5, 12).
ACKNOWLEDGMENTS
We are grateful to M. Challberg for providing plasmids and
a-UL52, and we thank members of this laboratory for helpful
commentson themanuscript.
This investigation was supported byPublic Health Servicegrant
A121747. S.K.W. is the recipient of an American Heart
Association-Genentech Established Investigator Award.
REFERENCES
1. Ausubel, F. M., R. Brent, R. E. Kingston,D. D. Moore, J. G. Seidman, J. A. Smith, and K. Struhl (ed.). 1990. Current protocolsinmolecular biology. John Wiley & Sons, Inc.,New York.
2. Aziaria, M., E. Schurr, and P. Gros. 1989. Discrete mutations
introduced in the predicted nucleotide-binding sites of themdrl geneabolishitsabilitytoconfermultidrug resistance.Mol.Cell.
Biol. 9:5289-5297.
3. Calder, J. M., and N. D. Stow. 1990. Herpes simplex virus
helicase-primase: the UL8 protein is not required for DNA-dependent ATPaseand DNA helicase activities. Nucleic Acids Res. 18:3573-3578.
4. Challberg, M. D. 1986. A method for identifying the viral genes required for herpesvirus DNA replication. Proc. Natl. Acad. Sci. USA 83:9094-9098.
5. Cohen, E. A., P.Gaudreau, P. Brazeau, and Y. Langelier. 1986. Specific inhibition of herpesvirus ribonucleotide reductase by a nonapeptide derived from the carboxy terminus of subunit 2. Nature (London)321:441-443.
6. Crute, J. J., and I. R. Lehman. 1991. Herpes simplex virus-1
helicase-primase, physical and catalytic properties. J. Biol.
Chem. 266:4484-4488.
7. Crute, J.J.,E. S.Mocarski, and I. R. Lehman. 1988. A DNA
helicase induced by herpes simplex virus type 1. Nucleic Acids Res. 16:6585-6596.
8. Crute, J. J., T. Tsurumi, L. Zhu, S. K. Weller, P. D. Olivo, M. D. Challberg, E. S. Mocarski, and I. R. Lehman. 1989. Herpes simplex virus 1 helicase-primase: a complex of three herpes-encoded gene products. Proc. Natl. Acad. Sci. USA 86:2186-2189.
9. Dean, F. B., P. Bullock, Y. Murakami, C. R. Wobbe, L. Weissbach, and J. Hurwitz. 1987. Simian virus 40(SV40) DNA replication: SV40large T antigen unwinds DNA containing the SV40 origin of replication. Proc.Natl. Acad. Sci. USA
84:16-20.
10. Dodson, M.S.,J. J. Crute, R. C. Bruckner, andI.R. Lehman. 1989. Overexpression and assembly of the herpes simplex virus type 1 helicase-primase in insect cells. J. Biol. Chem. 264: 20835-20838.
11. Dodson, M. S., andI. R. Lehman. 1991. Association of DNA helicase and primase activities with a subassembly of the herpes simplex virus 1 helicase-primase composed of the UL5 and UL52 gene products. Proc. Natl. Acad. Sci. USA 88:1105-1109. 12. Dutia, B. M., M. C. Frame, S. J. H. Subak, W. N. Clark, and J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
H.S. Marsden. 1986. Specific inhibition ofherpesvirus ribonu-cleotide reductase by synthetic peptides. Nature (London) 321:439-441.
13. Feinberg, A. P., and B. Vogelstein. 1983. A technique for radiolabeling DNA restriction fragments to high specific activ-ity. Anal. Biochem. 132:6-13.
14. Fry, D.C., S. A. Kuby, and A. S. Mildvan. 1986. ATP-binding site of adenylate kinase: mechanistic implications of its homol-ogy with ras-encoded p21, Fl-ATPase, and other nucleotide-binding proteins. Proc. Natl. Acad. Sci. USA 83:907-911. 15. Goldstein, D. J., and S. K. Weller. 1988. AnICP6::lacZ
inser-tional mutagen is used to demonstrate that the UL52 gene of herpes simplex virus type 1 is required for virus growth and DNAsynthesis. J. Virol.62:2970-2977.
16. Goldstein, D. J., and S. K. Weller. 1988. Factor(s) present in herpes simplex virus type 1-infected cells can compensate for the loss of the large subunit of the viral ribonucleotide reduc-tase: characterization of an ICP6 deletion mutant. Virology 166:41-51.
17. Gorbalenya, A. E., and E. V. Koonin. 1988. One more conserved sequencemotif in helicases. Nucleic Acids Res. 16:7734. 18. Gorbalenya, A. E., E. V. Koonin, A. P. Donchenko, and V. M.
Blinov. 1988. A conserved NTP-motif in putative helicases. Nature (London) 333:22-23.
19. Gorbalenya, A. E., E. V. Koonin, A. P. Donchenko, and V. M. Blinov. 1988. A novelsuperfamily of nucleoside triphosphate-binding motif containing proteins whichareprobably involved induplex unwinding inDNAandRNA replication and recom-bination. FEBS Lett. 235:16-24.
20. Gorbalenya,A.E.,E. V.Koonin,A. P. Donchenko,and V. M. Blinov. 1989. Two related superfamilies of putative helicases involved inreplication, recombination, repair and expression of DNAand RNA genomes. Nucleic AcidsRes. 17:4713-4730. 21. Graham,F.L.,and A.J.van der Eb.1973. Anewtechnique for
the assay ofinfectivity of human adenovirus5 DNA. Virology 52:456-467.
22. Heilbronn, R., and H. zur Hausen. 1989. A subset of herpes
simplex virus replication genes induces DNA amplification
within the host cell genome. J. Virol. 63:3683-3692.
23. Hodgman, T. C. 1988. A new superfamily of replicative pro-teins. Nature(London) 333:22-23.
24. Hodgman, T. C. 1988. A new superfamily of replicative pro-teins. Nature(London) 333:578. (Erratum.)
25. Kornberg, A. 1980. DNAreplication. W. H. Freeman &Co., San Francisco.
26. laCour,T. F. M., J. Nyborg, S. Thirup,and B. F. C. Clark.
1985. Structural details of thebindingofguanosinediphosphate to elongation factor Tu from E. coli as studied by X-ray crystallography. EMBO J. 4:2385-2388.
27. Lane, D. 1988. Enlarged familyofputative helicases. Nature (London) 334:478. (Letter.)
28. Linder, P., P. F. Lasko, P. J. Nielsen, K. Nishi, and P. P.
Slominski. 1989. Birth ofthe D-E-A-D box. Nature (London)
337:121-122.
29. Liu, Q.,and W. C.Summers. 1988.Site-directedmutagenesisof
anucleotide-bindingdomain in HSV-1thymidinekinase:effects
oncatalytic activity. Virology 163:638-642.
30. Maniatis, T., E. F.Fritsch, andJ. Sambrook. 1982. Molecular
cloning: alaboratory manual. ColdSpringHarborLaboratory,
ColdSpring Harbor, N.Y.
31. Matson, S., and K. A. Kaiser.Rogers. 1990. DNA helicases. Annu. Rev.Biochem. 59:289-329.
32. McGeoch,D.J.,M. A.Dalrymple,A.Dolan, D.McNab,L.J.
Perry, P. Taylor, and M. D. Challberg. 1988. Structures of herpes simplexvirus type 1 genes required for replication of virusDNA.J. Virol.62:444- 53.
33. Naumovski, L., G. Chu, P. Berg, and E. C. Friedberg. 1985.
RAD3 geneofSaccharomycescerevisiae: nucleotide sequence
ofwild-typeandmutantalleles, transcript mapping, and aspects
of generegulation. Mol. Cell. Biol. 5:17-26.
34. Naumovski, L., and E. C. Friedberg. 1986. Analysis of the essential and excision repair functions of the RAD3 gene of
Saccharomyces cerevisiae by mutagenesis. Mol. Cell. Biol.
6:1218-1227.
35. Naumovski, L.,and E.C. Friedberg. 1987. The RAD3geneof
Saccharomyces cerevisiae: isolation and characterization ofa
temperature-sensitive mutant in the essential function and of
extragenic suppressorsof this mutant. Mol. Gen. Genet. 209:
458-466.
36. Olivo,P.D.,N.J.Nelson, and M. D.Challberg. 1988. Herpes
simplex virus DNA replication: the UL9 gene encodes an
origin-binding protein. Proc. Natl. Acad. Sci. USA
85:5414-5418.
37. Pakula,A.A.,and R. T.Sauer. 1989. Geneticanalysisofprotein
stabilityandfunction.Annu. Rev.Genet. 23:289-310.
38. Russell, M., S. Kidd, and M. R. Kelley. 1986. An improved
filamentoushelper phageforgeneratingsingle-stranded plasmid DNA.Gene 45:333-338.
39. Stahl, H., P. Droege, and R. Knippers. 1986. DNA helicase
activityofSV40largetumorantigen.EMBO J. 5:1939-1944.
40. Stow,N.D. 1982. Localization ofanoriginofDNAreplication
within theTRS/IRSrepeated region of the herpes simplesvirus type1genome. EMBO J. 1:863-867.
41. Stow, N.D., and E. C. McMonagle. 1983. Characterization of theTRS/IRSoriginofDNAreplicationofherpessimplexvirus
type 1.Virology 130:427-438.
42. Sung, P.,D.Higgins, L.Prakash,and S. Prakash. 1988.
Muta-tionoflysine-48toargininein the yeast RAD3proteinabolishes
itsATPase and DNAhelicase activities butnot theability to bindATP. EMBO J. 7:3263-3269.
43. Walker, J. E.,M.Saraste,M.J.Runswick,and N.J. Gay. 1982.
Distantly related sequences in the a and P-subunits of ATP
synthase, myosin, kinases and other ATP-requiring enzymes
andacommonnucleotidebindingfold. EMBOJ. 1:945-951. 44. Weeda, G.,R.C.A.vanHam,W.Vermeulen,D.Bootsma,A.J.
vanderEb,andJ.H.J.Hoeimakers. 1990. ApresumedDNA
helicase encoded byERCC-3 is involved in thehuman repair
disordersxerodermapigmentosum andcockayne'ssyndrome.
Cell62:777-791.
45. Weller, S. K.,D. P. Aschman,W. R. Sacks,D. M. Coen,and
P. A.Schaffer. 1983. Geneticanalysisoftemperature-sensitive
mutants ofherpes simplexvirus type 1: the combineduse of
complementationandphysical mappingforcistronassignment.
J.Virol. 130:290-305.
46. Weller,S.K.,A.Spadaro, J.E.Schaffer,A. W.Murray,A. M.
Maxam, and P. A. Schaffer. 1985. Cloning, sequencing, and functionalanalysisoforiL,aherpessimplexvirus type1origin
of DNAsynthesis. Mol.Cell. Biol.5:930-942.
47. Wu,C. A.,N.J. Nelson,D.J.McGeoch,and M. D. Challberg.
1988. Identification of herpes simplex virus type 1 genes
re-quiredfor
origin-dependent
DNA synthesis. J. Virol.62:435-443.
48. Zhu, L., andS. K. Weller. 1988. UL5, aprotein required for
HSVDNAsynthesis:genetic analysis,overexpressionin
Esch-erichiacoli, andgenerationofpolyclonalantibodies.
Virology
166:366-378.
49. Zhu, L.,R.Martinez, L.Shao,andS. K. Weller.
Unpublished
data.
50. Zhu, L., and S. K. Weller. 1992. The UL5 gene of
herpes
simplex virustype 1: isolation ofalacZinsertionmutantand
association oftheUL5 geneproductwithothermembers of the
helicase-primase complex.J. Virol. 66:458-468.
51. Zhu,L.,andS. K. Weller. Unpublisheddata.