Copyright ©1984, American Society for Microbiology
Polyomavirus and Simian Virus 40 Large
T
Antigens
Bind
to
Common DNA
Sequences
BETSY J. POMERANTZAND JOHNA. HASSELL*
Department ofMicrobiology andImmunology, McGill University, Montreal, Quebec, Canada H3A 2B4
Received 2 October1983/Accepted 5December1983
The large T antigens of polyomavirus and simian virus 40 (SV40) recognize and bind to specific, noncoding DNAsequenceswhicharelocated between thebeginning of the early and late transcription units intheir respectivegenomes.Eachlarge T antigen bindstomultiple sites within this intergenicDNAstretch. Polyomavirus large T antigen bindsto atleasttwosites within its DNA,andSV40largeTantigen bindsto three sites within SV40 DNA. Comparison of the DNA sequences which comprise the binding sites in polyomavirus DNA orthose which make up the binding sites in SV40 DNA has led to recognition ofa
common sequence, -GAGGC-, which is repeated within each large-T-antigen-binding site. We tested the hypothesis thatrepeatsof this pentanucleotide form therecognition-bindingsiteforpolyomavirusandSV40 large T antigen. This was accomplished by measuring the binding of each large T antigen to both polyomavirus andSV40DNAandto synthetic DNA substrateswhich did ordidnotcontainrepeats ofthe -GAGGC- sequence. Polyomavirus large Tantigen bound to specific fragments ofSV40 DNA, and SV40 large T antigen bound with specificity topolyomavirus DNA. In each case, the DNAfragments bound by the heterologous large T antigen were the same as those bound by the homologous large T antigen. Moreover, polyomavirus and SV40 large T antigen only bound to synthetic DNA substrates which contained repeats of the pentameric sequence. This synthetic DNA also competed effectively withnative polyomavirusorSV40 DNAas asubstrateinbinding reactions withone orthe otherlargeTantigen. These results ledustoconclude that repeatsof the-GAGGC- sequenceform therecognition-binding site for both polyomavirus and SV40large T antigen.
Polyomavirusandsimian virus40(SV40),twomembersof
the papovaviruses, share a common virion architecture,
genome structure and organization, and replication cycle.
With some exceptions, this synonomy extends tothe
num-ber,molecularweights, amino acidsequences, and functions of the viral-encoded proteins (13, 20, 42). Nonetheless, important differences distinguish these viruses. They
repli-cate in radically different hosts; polyomavirus multiplies in murine cells, and SV40 multiplies in simian cells. The
products of their transcription units are not all identical;
polyomavirus encodes one moreearlygeneproduct(middle Tantigen) than does SV40. There are sequencedifferences within the intergenic, regulatory regions of their genomes
(20).The latterincludenot only alterationsinprimaryDNA structure,but alsodifferencesin theorganizationof the viral controlelementssuch astheorigin forDNAreplication (31),
the early and late promoters (15, 24), and the number and
locationofbinding sites for regulatory proteins (15, 34). We havebegun todefinethe borders and toidentify the
impor-tant DNA sequences within these intergenic regulatory
regions in polyomavirus andhavecompared their sequence andorganizationtosimilarcontrolelements inother papova-viruses to derive general models for the initiation and
regulation of DNAreplicationandtranscription (24, 31, 34). Because both viruses make extensive use ofhost cell
ma-chinery to implement these processes, we anticipate that what we learn about replication and transcription from studies of papovaviruses can be generalized to mammalian cells.
Thereplicationofcyclesofpolyomavirus and SV40 occur in twotemporal stages. Thefirst, defined as the early phase,
begins soonafterinfection and extendsto the onsetof viral DNA replication. During this time the viral early
transcrip-*Corresponding author.
tion units are expressed to yield theearly proteins. Polyoma-virus encodes three early proteins, large, middle, and small Tantigen, whereasSV40 codes for two early proteins, large
and small T antigen (20). Thesynthesis of viral DNAsignals the beginning of the late phase of infection. During this phase, the late transcription units areexpressed to yield the three structural proteins of polyomavirus and SV40 VP-1,
VP-2, and VP-3 (20). Now the relative synthesis of tran-scripts from the early transcription unit is drastically re-duced. Both the initiation of viral DNA replication and the repression of early mRNAsynthesis at late times is mediated by large T antigen (7, 8, 44, 48). The mechanism by which large T antigen performs these functions has not been elucidated in equal detail for both viruses. However, it is verylikelythatin both caseslargeTantigenmustphysically
interact with specific viral sequences to effect both replica-tion and early transcription (41).
The large Tantigens ofpolyomavirus andSV40are both nuclear phosphoproteins with calculated molecularweights of87,991 (785 amino acids) and 81,632 (708 amino acids),
respectively, althoughdirect measurements of their molecu-lar weights have led to higher estimates ofbetween 88,000 and 100,000 (20). Both proteinsexist as multiple molecular species (2, 11, 12, 19, 22, 27, 36). A fraction of the oligomeric
species ofSV40 large T antigen, and perhaps of
polyoma-virus large T antigen, contain a cellular-encoded
phospho-protein, P53, also referred to as nonviral tumorantigen or Tau antigen (11, 19, 22, 26). The biological roles of these variousforms oflarge T antigen are not known with certain-ty, but they differ in their extent of phosphorylation and possess different activities in vitro (2, 11, 17, 36). One
activityof the largeTantigenencodedbypolyomavirusand
SV40 is the capacity to hydrolyze ATPto ADP and
Pi
(14, 16, 21, 49). Anotheractivity is the capacity to bind to DNA (4, 14). Polyomavirus large T antigen binds to at least two 925on November 10, 2019 by guest
http://jvi.asm.org/
926 POMERANTZ AND HASSELL
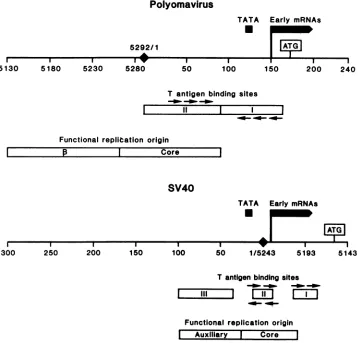
sites within polyomavirus DNA (34) (Fig. 1). One of these
sites overlaps with DNA sequences which form part of the
replication origin, whereas the other straddles sequences
importantfor earlymRNA synthesis (15, 24, 31). SV40large T antigen binds to three, closely spaced sites within SV40
DNA which occupy a region 120 to 180 base pairs (bp) in length (9, 40, 43, 45-47) (Fig. 1). These large-T-antigen-binding sites overlap sequences which comprise the early promoter and origin for DNA replication (20) (Fig. 1). Experiments with SV40 DNA and a large-T-antigen-related
protein, the D2 protein, have shown that the guanine resi-dues in the binding sites which are part of the sequence
-GAGGC-areprotected by the D2 protein from methylation withdimethyl sulfate (47). This pentanucleotide is repeated in two of the SV40 large-T-antigen-binding sites (Fig. 1).
Repetitionof this experimentunder avariety of experimen-talconditionswithauthenticSV40 large Tantigen has led to
nearlyidentical conclusions (9).
Inspectionofthe DNAsequences comprising the
large-T-antigen-binding sites in
polyomavirus
DNA reveals that therelated sequence -A/TGAGGC- is repeated in both of its binding sites (Fig. 1). The conservation of
homologous
hexameric and pentameric repeats within the large-T-anti-gen-binding sites in polyomavirus andSV40 DNA suggests
that these sequences are functionally important and might
serve as the recognition-binding sites for both large T antigens (34). To test this hypothesis, we measured the
bindingofpolyomavirusorSV40 largeTantigen to the DNA of the other andtosyntheticDNApolymerswhich contained repetitions of the -GAGGC- sequence. The results ofthese experiments substantiate the contention that repeats ofthe
pentanucleotideform the coreofthepolyomavirus and SV40
large-T-antigen-binding sites.
MATERIALS AND METHODS
Cells and viruses. Cells were grown onplastic disheswith Dulbecco modified Eagle medium supplemented with 10%
(vol/vol) fetal bovine serum and antibiotics and maintained
Polyomavirus
TATA Early mRNAs
*
AT 5292/1
I I I I _
5130 5180 5230 5280
T antigen binding sites
--D --Om --O
II
I I I --I
___- -
-Functional replitation origin
I
pI
CoreSV40
I 2
250 200
TATA Early mRNAs
I I I5
rw
150 100 S0 1/5243 5193 51
Tantigen binding sites
~ -00
Functional replication origin Auxiliary I Core
FIG. 1. Polyomavirus andSV40large-T-antigen-bindingsitesrelative to other controlsequences.Theregulatory regionsofpolyomavirus andSV40areshown. The nucleotidenumberingscheme of Soedaetal.(42)wasused forpolyomavirusandthat of Buchman et al.(3)wasused
forSV40. The solid diamond represents thejunctionbetween thefirst and last nucleotide in the DNA sequence.The -TATA- consensus sequenceand the5'termini of the abundantearlymRNAsareshownbythe solid boxes(15, 20).Thedirection ofearlytranscriptionisfrom
lefttorightasshownby thearrow onthe solid boxrepresentingtheearlymRNAs. Theputativeinitiationcodons for translation of theearly mRNAsareboxed. Thelarge-T-antigen-binding sitesareillustrated byopenboxes. The boundaries of thepolyomavirus large-T-antigen-bindingsitesweretaken from Pomerantzetal.(34)and those ofSV40weretaken from DeLucia etal.(9).Thearrowsabove and belowthe
large-T-antigen-bindingsites refertothe -GAGGC-sequence,and theirpositionaboveorbelowtheopenbox refer to their locationononeor theother DNA strand. The minimal DNAsequenceswhich functionasreplication originsarealso shownasopen boxes. For eachvirusthere
are a minimumoftwo sequence domains comprising the functional replication origin. For polyomavirus these are referred to as ,3 and
core,whereasforSV40 theyarereferredtoasauxiliarysequencesandcore.The borders of these sequencesaretaken from Muller et al.(31) forpolyomavirus and fromMyersandTjian(33), DiMaio and Nathans(10), andBergsmaetal.(1)forSV40.
I I I I I
50 100 150 200 240
300 43
---s
J.VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.131.489.281.624.2]at 37°C in a humidified CO2 atmosphere. Polyomavirus stocks were prepared by infection of primary baby mouse
kidney cells at low multiplicity (0.01 to0.1 PFU per
cell).
Infected cells were harvested 10 to 14 days postinfection. Stocks ofSV40were
prepared similarly by
infection ofCV-1 cells. Viruswas releasedfrom infected cellsbythreecycles
offreezingand thawing.
Construction of recombinant plasmids. pPSVE1 is
com-prised of pBR322 sequences, the SV40HindlIl C fragment
(SV40 nucleotide 1047 to5171), and the
polyomavirus HphI
(nucleotide 154) to EcoRI (nucleotide 1560) fragment (see Fig. 2). The numberingsystemsusedarethoseproposed by
Soedaet al. (42)and Buchmanet al. (3)for the A2 strain of
polyomavirus and the 776 strain ofSV40, respectively. We
cite nucleotides from the clockwise direction on the
poly-omavirus and SV40
physical
maps. The SV40sequences inpPSVE1werejoinedtothe
polyomavirus
sequences insuchawayas tojuxtapose the SV40earlypromoterimmediately adjacenttothe polyomavirusDNA which encodes the
early
proteins. The details concerning the construction of this
recombinant plasmid will be described elsewhere.
Impor-tantly, pPSVE1 contains the natural sites of
binding
forSV40largeTantigen
(within
thebordersof theSV40 HindlIlC fragment)butdoes notcontain the natural sites of
binding
for the polyomavirus large T
antigen.
The methods usedto construct pPH1-8 (25) andpSVOl (33)
have been described (see Fig. 2).Isolation of DNA and its modification. Recombinant
plas-midDNAswere isolated fromEscherichia coli strainHB101
orDH1 and
purified by
CsCldensity centrifugation.
Restric-tion enzyme
digestions
wereperformed
in accordance withthe
specifications
of the manufacturer. To end label theDNAfragments, 100 ngof restriction
enzyme-cleaved
DNA was incubated in a volume of 50 ,ul with 10 mMTris-hydrochloride (pH 7.6), 5 mM MgCl2, 1 mM
dithiothreitol,
25 p.Ci of the appropriate a-32P-labeled
deoxynucleotide
triphosphate (3,000
Ci/mmol),
and 5 U of the Klenowfragment ofE. coli DNA polymerase I for 1 h at
15°C.
Thereaction was terminated by phenol
extraction,
and the aqueous phase waschromatographed
on aSephadex
G-50column to separate the
unincorporated
deoxynucleotide
triphosphates from the labeled DNA
fragments. Typically,
the specific activity of the labeled DNA
fragments averaged
108 cpm/,ug.
Synthetic
oligodeoxynucleotides
werephosphorylated
and ligated foruse asDNAsubstrates in the
binding
assays. From 1 to 4jig
of DNA was labeled with 50p.Ci
of[y-32P]ATP
and 5 U ofT4kinase in abuffercontaining
70 mMTris-hydrochloride (pH 7.6), 10 mM MgCl,, and 5 mM
dithiothreitol. After 1/2 h at 37°C, unlabeled ATP
(final
concentration, 0.5 mM) and 5 U ofT4 kinase were
added,
and thereactionwascontinuedfor another 1/2 hat37°C.
Thephosphorylatedlinkerswere
ligated
for12to16 h at 15°Cin 50 mMTris-hydrochloride (pH
8.0)-10 mMMgCl2-13
mMdithiothreitol-1mMATPwith 10 U ofT4
ligase
in avolumeof100
[L.
Preparation ofnuclear extractsfrominfected cells. Nuclear
extracts were prepared from 3T6 cells infected with
poly-omavirus or CV-1 cells infected with SV40
essentially
as describedby McKay (28). From 0.5 x109
to 1.0 x109
cellsgrowingon the surface of plastic petri dishes were infected with virus at a multiplicity of infection of about 10. The cells
wereharvested by scraping (40 to 46 h postinfection at
37°C)
and then washed twice with ice-cold phosphate-buffered salinesolution. The cellswereresuspended in
lysis
buffer (10mM Tris-hydrochloride, pH 7.4, 10 mM NaCl, 1.5 mM
MgC92,
0.5 mM dithiothreitol, 100p.g
of phenylmethylsul-fonyl fluoride per ml) ata density of5 x107
cells per ml.Theywere allowedtoswell at4°Cfor10 min and thenlysed
by 10to 15 strokes with atype B pestle in a glassDounce
homogenizer. The nuclei were pelleted by centrifugation
(2,000 rpmfor10 min at4°C)andwashed inlysisbuffer. The
nucleiwerethenresuspended in nuclearextraction buffer(10 mM Tris-hydrochloride, pH 8.0, 150 mM NaCl [for
SV40-infected CV-1 cells] or 300 mM NaCl [for polyomavirus-infected 3T6 cells] plus 0.5 mM dithiothreitol, 100 ,ug of phenylmethylsulfonyl fluoride per ml) and incubated at4°C
for5 to 15 min. The nucleiwere pelleted bycentrifugation,
and the supernatantwasusedas a sourceoflargeTantigen. These preparations were storedat -70°C until use.
Immunoassay. The conditions of the
immunoassay
wereessentially those described
by
McKay(28).
Briefly,
reac-tions were carried out in a 1-ml volume
containing
20 mMNaPO4
(pH 7.0),2 mMdithiothreitol,
0.01%(wt/vol)
bovineserum
albumin,
0.1 mM EDTA, 0.05%(vol/vol)
NonidetP-40,40 ,ugof
phenylmethylsulfonyl
fluoride perml,
and3.0%(vol/vol) dimethyl sulfoxide.From 1to10 ngof labeled DNA
fragmentsandvariousamountsofthe
large-T-antigen
prepa-rationswerealso included. Aftera 1-hincubationat
25°C,
2 ,ugofunlabeledratcellular DNAwasadded,
andincubationcontinuedfor 20 min at
25°C.
The inclusion ofnonradioac-tive DNA reduces the
nonspecific binding
oflarge
Tantigen
to the labeled DNA. The
large
Tantigen-DNA complexes
wereimmunoprecipitated by
sequential
incubation with anti-bodies directedagainst
the virallarge
Tantigens
and then with Formalin-fixedStaphylococcus
aiureius
cells. Thesereactions were
performed
at25°C
for 20 min in eachin-stance.
Polyclonal
antisera directedagainst polyomavirus
large T antigen were obtained from ascites fluid of brown
Norwegian rats
bearing
tumors inducedby
polyomavirus-transformed rat cells. Hamster antiserum directed
against
SV40largeTantigenwas obtainedfrom the National
Insti-tutesofHealth(serum-ID = 81 x0000001). Neither
antiser-um was purified before use. The
immunocomplexes
were then washed twice with 1 ml of 10 mMTris-hydrochloride
(pH 8.0-150 mM NaCI-0.5% Nonidet P-40.
Finally,
the DNA was released from theimmunocomplexes
with 1% (wt/vol) sodium dodecyl sulfate-10 mM EDTA and depro-teinized withphenol
andchloroform/isoamyl
alcohol(24:1
[vol/vol]), and a portion was electrophoresed through an
agarose or
polyacrylamide gel;
1.5%(wt/vol)
agarosegels
and 12%
(wt/vol) polyacrylamide gels
were used toanalyze
the DNA.
Electrophoresis
wasperformed
in 89 mMTris-hydrochloride
(pH 8.0)-89 mM boric acid-2 mM EDTA.Before
autoradiography,
agarosegels
were immersed in ethanol for 30 min and dried under vacuum withheating.
Polyacrylamide
gelswere notdried beforeautoradiography.
Generally,
gels wereexposed to Kodak XAR-5film for6to 36 h in casettes with DupontLightning-Plus
intensifying
screens.
RESULTS
Weemployedtheimmunoassay described by McKay (28)
to measurethe bindingoflargeTantigen presentin nuclear
extracts from
lytically
infected cells to end-labeledfrag-mentsofpolyomavirus orSV40 DNA. This assay makesuse
of the fact that DNA bound to a
particular
protein
can beseparated from free DNA by immunoprecipitation with
antibody
specific
to that protein. Inbrief,
defined radiola-beled DNAfragments
were reacted with nuclear prepara-tions oflargeT antigen,and thefragments boundby large
Tantigen
wereimmunoprecipitated
withanantiserumdirectedon November 10, 2019 by guest
http://jvi.asm.org/
928 POMERANTZ AND HASSELL
V
O3
75
e4~
VP=;:
;
~~~~~53
as EcoRN 10
9so
/2
oso0)
EcoR11
\6/(5092) 3 \
225 7
a8
PsVO1
6 24948713bp_
\~~~~~~~~~~~~~~~
11
X /270
1743
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
against large T antigen and Formalin-fixed S. aureus cells. The immunocomplexes were washed, and the eluted DNA
was analyzed by gel electrophoresis and autoradiography. Only DNA fragments which were boundspecifically by the large T antigen were displayed on the autoradiogram. For convenience, and to simplify the interpretation of the
re-sults, we used recombinant plasmid DNAs which were
known to harbor the large-T-antigen-binding sites as
sub-strates inthe binding reactions. Their structures are shown
in Fig. 2togetherwith appropriate restriction endonuclease cleavage maps. In addition, the location of the
large-T-antigen-binding sites, themajorcap site, and the direction of transcription of the early mRNAs are shown. pPH1-8
con-tains the two known polyomavirus large-T-antigen-binding
sites (34) within the 604-base pair (bp) Hinfl 4 fragment, which is composed of only polyomavirus DNA (Fig. 2).
pPSVOlandpSVE1 containthe threeSV40 large-T-antigen-binding sites (33) within the borders of the Avall-HindlIl 2 and 4fragments, respectively (Fig. 2).
Specific bindingofpolyomavirusandSV40largeTantigen
to each other's DNAs. To test thehypothesis thatrepeats of the sequence -GAGGC- arerequired for the specific binding of polyomavirus and SV40 large T antigen to DNA, we
measuredthe binding of each large T antigen to its own DNA and to theheterologous DNA. We employed this approach because each of the large-T-antigen-binding sites in the genomes of these viruses contains repeats ofthe aforemen-tioned sequence(Fig. 1). Wereasoned that ifrepeats of this sequence are important forbinding oflarge T antigen then eachspecies of largeTantigenought to be capable of binding toboth DNA substrates. Initially, wemeasured the binding
ofpolyomavirus largeTantigen present in a nuclear extract from infected 3T6 cells to end-labeled DNA fragments
obtained by Hinfl hydrolysis ofpPH1-8 DNA (a recombi-nantplasmidDNAwhich containspolyomavirus sequences) (Fig. 2) and to the end-labeled fragments obtained after cleavage of pSVOl DNA (a recombinant plasmid DNA whichcontains theSV40large-T-antigen-binding sites) (Fig. 2) withAvall and HindIII (Fig. 3A and B). We previously
showed thatpolyomavirus largeTantigen binds specifically
totheHinfl4fragment of pPH1-8DNA(34), a resultwhich is confirmed here (Fig. 3A). When polyomavirus large T
antigen was incubated with the end-labeled AvaIl-HindIll fragments of pSVO1 DNA, the second largest fragment in themixture was selectively bound(Fig. 3B). Thisfragment contains the three SV40large-T-antigen-binding sites (Fig. 2).
One of the problems encountered with this assay is the
immunoprecipitation of DNA fragments other than those whichcontainthelarge-T-antigen-binding sites(Fig. 3A and
A B
DNA Hinf L,'pPH1- 8 Ava I1+Hina 111ir)SVO|
(ng Iml) 1 1 5 5 1 1 5
T-antigen Polyomavirus PolyornaviIr.s l
(ul1) 25 50 25 50 25 50 f5 50
1W
-1 - 1 _ _
2 0 -* 5
3 40
8
9 ~~~~~~~4
10.11 5
12,13 A* 14.15S
16-20 8-11 i
M 1 2 3 4 M 1 2 3 4
FIG. 3. Immunoprecipitation ofpolyomavirus and SV40 DNA fragments after reaction with nuclear extract from polyomavirus-infected cells. (A) pPH1-8 DNA was cleaved withHinfl (see Fig. 2A), and thefragments were endlabeled and usedas substrate in binding reactionsattheconcentrations shown in thefigure. Several concentrations of nuclearextractfrom polyomavirus-infected 3T6 cellswereassayed. The marker lane (M)containsaportion ofthe original DNA substrate (pPH1-8 cleaved with Hinfl). Fragment 4 contains the previouslymappedpolyomavirus large-T-antigen-bind-ing sites(seeFig.1and 2).The driedagarosegelwasexposedtofilm for18 h. (B) Nuclearextractfrompolyomavirus-infected 3T6cells was reacted with end-labeled fragments generated by AvaIl and
HindIll cleavage of pSVO1 DNA (see Fig. 2). M represents a portion of the
AvaIIIHindIII
digestion products of pSVO1 DNA usedassubstrate. Fragment 2contains the previouslymapped SV40 large-T-antigen-binding sites (Fig. 1 and 2). The driedagarosegel wasexposed tofilm for18 h.B). These fragments are immunoprecipitated to a lesser extent than thosewhich containthebinding sites forlarge T
antigen (Fig. 3A and B). The largest fragments are usually
preferentially precipitatedunder these conditions (see
espe-cially Fig. 3A lanes 3 and 4and Fig. 3B lanes3 and 4). We suspect that thisoccurseither because large T antigen binds
nonspecifically to DNA fragments or because the protein
bindsspecificallytoweakrecognition-binding sitesin DNA. In addition, we have found that many preparations of
antiserum contain DNA-binding proteins. Their interaction
with the labeled DNA fragments also contributes to the
background bindingweobserved.
To determine whether SV40 large T antigen would bind
FIG. 2. Structures and restriction endonucleasecleavage mapsof recombinantplasmidDNAs. TherecombinantplasmidpPH1-8contains polyomavirussequencesbetweentheHindIll cleavagesitesatnucleotides3918 and 1656(clockwiseon thepolyomavirus DNAmap)inserted into theuniqueHindIll site of pBR322.The large openbox representspolyomavirusDNA; thethin line represents pBR322 sequences. Small arrowsdesignate sites ofHinflcleavage, and the two larger arrows denote those Hinfl sites which flank the previously mapped large-T-antigen-binding region. Sizes of restriction fragments (inbp) are notedaround the periphery of the plasmid. Therelativesize of each fragment isindicated bythe number within the circle. The symbols above the plasmid represent features of polyomavirus DNA which reside between the early andlate transcription units. The open triangle denotes the area in which DNA replication initiates, the closed box represents the sequencesboundby the polyomaviruslarge Tantigen,and the open arrow depicts the site ofinitiationandthedirectionof transcription of the early viral mRNAs. pSVOlcontainsthe EcoRIIG fragmentofSV40 DNA(nucleotide5092 to 160)inserted into the uniqueEcoRI site of pBR322DNA(nucleotide4362/1). The SV40large-T-antigen-bindingsites arecontained within the EcoRII G fragment. The hatched area represents SV40 sequences. The other symbols are the same as those used above. pPSVE1 contains theHindIlI C fragment ofSV40 DNA (nucleotide 1047 to 5171; thehatched box) and theHphItoEcoRIfragment of polyomavirus DNA (nucleotide 154 to 1560; the open box) cloned between theHindIll (nucleotide29)andEcoRI(nucleotide4362/1)sitesof pRB322 DNA. AHindIll recognitionsite is present at the junction oftheSV40(nucleotide5171) andpolyomavirus (nucleotide 154) DNAsequences. The hatched box represents SV40 DNA, the open
boxrepresentspolyomavirusDNA, and thethinline represents pBR322 DNA. The othersymbols are the same as those used above.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.332.547.75.265.2]930 POMERANTZ AND HASSELL
with specificity to DNA substrates which contain the binding sites for polyomavirus large T antigen, we measured its
capacity tobindto defined fragments of pPSVE1 and pPH1-8 DNA (Fig. 2 and 4A and B). As expected, the nuclear extractcontaining SV40 large T antigen bound with specific-ity to the AvaII-HindIII 4 fragment of pPSVE1 DNA (Fig. 4A).The latter fragment carries the sites of binding forSV40 large Tantigen. In addition, the Hinfl 4 fragment of pPH1-8 DNA, which contains the sites of binding for polyomavirus large T antigen, was specifically bound by SV40 large T antigen (Fig. 4B). These results show that SV40 large T antigen binds with specificity to a DNA fragment which contains the sites of binding for polyomavirus large T antigen.
To better gauge the affinities of polyomavirus and SV40 large T antigens for each other's natural binding sites, we compared the binding of the two large T antigens to the other's DNA under conditions where the concentration of
nuclearextract was limiting and all the other components in the bindrigassay were in excess. To make the comparison
asmeaningfulaspossible, we simultaneously prepared fresh nuclear extracts from 3T6 cells infected with p;lyomavirus and CV-1 cells infected with SV40. In each case, the cells were infected at a multiplicity of 10, and extracts were prepared 48 h postinfection from the same number of cells and were handled identically before they were assayed. A
A B
DNA Ava IX+Hind IIf/pPSVE1 HinfIf/pPH1-8
(ng/mI) 1 1 5 5 1 1 5 5
T-antigen SV40 SV40
(ul) 50100 50 100 50 100 50 100
2,3 ES
44 4
--_*
~~5-7
67,889
96
i10,19!10
11,12 12.13
13
14,15
14 16
15--17 * 17-20
M 1 2 3 4 M 1 2 3 4
FIG. 4. Immunoprecipitation of polyomavirus and SV40 DNA fragments after reaction with nuclear extract from SV40-infected CV-1 cells.(A)pPSVE1DNAwasdigestedwithAvaIl andHindlll (Fig. 2), and the fragments were end labeled and reacted with nuclearextractfromSV40-infectedCV-1 cells. Theconcentration of DNAand extract used in each reaction is shown in thefigure. M represents asampleoftheAvaIl andHindIll fragmentsofpPSVE1 DNA used as substrate. Fragment 4 contains the SV40
large-T-antigen-binding sites. The dried agarosegel was autoradiographed for 24 h. (B) pPH1-8 DNA was cleaved with Hinfl, and the fragments were end labeled and used as substrate in binding
reactions with nuclear extractfromSV40-infectedCV-1 cells. The concentrations of DNA and extract are shown in the figure. M shows the Hinfl digestion products of pPH1-8 DNA used as
substrate. The fourth largest fragment contains the polyomavirus large-T-antigen-binding sites.Thedriedagarose gelwas autoradio-graphed for24 h.
(.ti) 25 5 f 0
24 3 _ 4 5-7*
8
12,13
-14,15 *
16 20 9
L
M 1 2 3 4FIG. 5. Immunoprecipitation of polyomavirus DNA fragments after reaction with nuclear extract from polyomavirus- or SV40-infected cells. pPH1-8 DNA was cleaved with Hinfl, and the fragmentswereend labeled and reacted with nuclear extract from either polyomavirus-infected3T6 cells orSV40-infectedCV-1 cells. AfixedconcentrationofDNA but variousamounts of extract were assayed. M represents a sampleoftheHinfI-cleaved pPH1-8DNA used as substrate. The dried agarose gel was exposed tofilm for 18h.
comparison of the capacities of polyomavirus and SV40 large Tantigenstobindto aDNAsubstrate which contained the polyomavirus large-T-antigen-binding sites is shown in
Fig.5. The results showed that bothlargeTantigensbound
totheHinfl 4 fragment of pPH1-8toapproximatelythe same extent.
Wealso compared thebinding of polyomavirus and SV40 large T antigens to DNA fragments which contained the SV40 large-T-antigen-binding sites (viz., pPSVE1 DNA) (Fig. 6). These results showed that both large T antigens bound specifically to the Avall-HindIl 4
fragment
ofpPSVE1 DNA. However,unlike the results obtained previ-ously, the affinity of polyomavirus large T antigen for the DNAfragmentcontainingthe SV40large-T-antigen-binding
sites appeared to be less than that ofSV40largeT antigen
(Fig. 6).
Insummary,the resultsweobtainedstronglysuggestthat
thelarge Tantigens ofpolyomavirus and SV40are capable
of
binding
to each other's genomes with specificity.More-over, our data are consistent with the hypothesis that the two largeT antigens recognize andbind to the same DNA
sequences.
PolyomavirusandSV40 largeT antigenspecific bindingto synthetic DNA substrates which contain repeats of the se-quence -GAGGC-. To
directly
test thehypothesis
that re-peatsof the sequence -GAGGC-arerequiredfor thebindingof polyomavirus and SV40 large T
antigens
toDNA,
we searched for this sequence amongcommercially
availablesyntheticDNAlinkers andadaptors. Wenotedthatthe
self-ligation of XhoI linkers
(-CCTCGAGG-)
resulted in thecreation of the -GAGGC- sequence and that trimers of XhoI linkers caused the duplication of this sequence within the molecule separated by3 bp.This arrangement of the penta-nucleotides mimics their topography in the large-T-anti-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.369.508.74.292.2] [image:6.612.75.291.357.575.2]gen-binding sites of polyomavirus and SV40 DNA. There-fore, we tested the capacity of self-ligated XhoI linkers to serve as a substrate for the binding ofpolyomavirus and SV40 large T antigen. To ensure that the conditions we employedfortheseassayspermitted only specific binding to bescored, we also measured thebindingoflarge T antigen to three other synthetic DNA substrates. These included
self-ligated BamHI (-CGGGATCCCG-), BclI (-CTGATCAG-), andClaI (-CATCGATG-)linkers. Polymersof these linkers
do notform the -GAGGC- repeat. The results of measuring
the binding of polyomavirus large Tantigen to these DNA substrates are shown in Fig. 7. Only the ligated XhoI linkers were specificallybound after reaction with nuclear
prepara-tionsof polyomavirus large T antigen. Moreover, the
larger-molecular-weight polymers were preferentially bound by
polyomaviruslarge T antigen (Fig. 7). The smallest multimer
of the ligated XhoI linkers to be bound was a pentamer. Pentamers contain four repeats of the -GAGGC- sequence per DNA strand, with each repeat separated from the
adjacentrepeat by 3 bp. Inspection of Fig. 7 also reveals that there was virtually no binding of polyomavirus large T
antigen to the ligated BamHI, BclI, or ClaI linkers. This suggests that the background binding which we observed
previously (see Fig. 3A and B) was caused bythe specific binding of polyomavirus large T antigen to weak binding sites in theDNA.
Werepeated this experiment butemployed SV40 large T
antigenin thebindingassay(Fig. 8). Again,the
polymerized
XhoI linkerswereefficiently bound by SV40largeTantigen, but under identical conditions, polymers ofBamHI, BclI,
andClaI linkerswere not(Fig.8).AlsoshowninFig.8Aisa
comparison between the capacities of polyomavirus and SV40large Tantigenstobindtotheself-ligatedXhoIlinkers.
DNA Ava I1+Hind lII/pPSVE1
(naQ/nil) 5 5 5 5
T-antigen SV40 Py
(yI) 25 50 25 50
2,3 *
4 -4 Q
5
7,8-
9- I10-1
1,12k-13 " 14
15-17
M 1 2 3 4
FIG. 6. Immunoprecipitation of SV40DNAfragments after reac-tion withnuclear extractfrompolyomavirus-orSV40-infected cells. pPSVE1 DNA was cleaved with AvaII and HindIII, and the fragments wereend labeledand reacted withnuclearextractfrom eitherpolyomavirus-infected3T6cellsorSV40-infected CV-1 cells. Afixed concentration ofDNA wasreactedwithincreasingamounts of nuclear extract. Mrepresents asample ofthe DNA fragments used as substrate. The dried agarose gelwas exposed to film for 24 h.
FIG. 7. Immunoprecipitation of ligated synthetic linkers after reaction withnuclearextractfrompolyomavirus-infected cells. The linkers were phosphorylated andligatedand thenusedassubstrates in binding reactions with nuclearextractfrom polyomavirus-infect-ed 3T6 cells (30 ,ul perreaction).Theconcentration of linkerDNA was 100ng/ml (lanes 1, 6, and 9), 50 ng/ml (lanes 2,4, 7and 10) and 25ng/ml (lanes 3, 5, 8, and 11); 100 pg of eachDNAsubstratewas run as amarker(M). Thepolyacrylamide gelwasexposedtofilm for 18 h.
SV40 large T antigen clearly binds to this substrate to a greater extent than does polyomavirus large T antigen.
These data and those shown in Fig. 7 demonstrate that repeats of the pentanucleotide sequence -GAGGC- are
rec-ognizedand bound by both
polyomavirus
andSV40largeTantigens under conditions which only allow for
specific
binding.
Binding of large T antigens of polyomavirusand SV40 to common DNA sequences. The data we obtained heretofore
showed that both large T antigenswere capable of binding with specificity to DNA fragments encompassingeach
oth-er's binding sites, and that they werecapable ofbindingto
synthetic DNA substrates containing repeats ofthe
penta-meric sequence -GAGGC-. To prove that both large T
antigens bind to the same sequences in
polyomavirus
andSV40DNAand to prove that theimportant characteristic of this sequence is the repeat of the
pentanucleotide,
weconducted the
experiments
outlined below. First, wemea-suredthecapacity of variousunlabeledpolymers created by self-ligation of linkerstocompetewithlabeledpolyomavirus
DNA as substrate in binding reactions with polyomavirus large T antigen. We repeated this
experiment
with SV40DNA and SV40 large T antigen. Second, we determined
whether thesynthetic DNAs could competeeffectivelywith
SV40 DNA as a substrate when polyomavirus large T
antigen was used as a source ofprotein and whetherthese
synthetic DNAs could act as competitors in binding
reac-tions with polyomavirus DNA and SV40 large T antigen. To conduct these experiments, we end labeled the DNA
fragments obtained by restriction endonuclease cleavage of various recombinantplasmids and reacted these with a fixed
quantity of large T antigen and various amounts of
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.378.497.74.315.2] [image:7.612.108.251.419.645.2]932 POMERANTZ AND HASSELL
A
Xho
U
U1
isaa
li'.
_lip
to-__
__
M1234
B
BamHIBclI Cla I
6
U0
M 1M34M5
M 12M34 M56
FIG. 8. Immunoprecipitation of ligated synthetic linkers after
reaction with nuclearextract from polyomavirus- orSV40-infected
cells. (A) Synthetic XhoI linkers were phosphorylated and ligated
and then reactedataconcentration of25ng/ml (lanes1and3)or50
ng/ml (lanes 2 and 4) with 30 ,u1 of nuclear extract from
SV40-infected CV-1 cells (lanes 1 and 2) or polyomavirus-infected 3T6
cells(lanes 3 and 4). The marker lane (M) contained the phosphory-lated ligated XhoI linkers usedassubstrate. The polyacrylamide gel was exposed to filmn for 6h. (B) Synthetic BamHI, BclI, and ClaI
linkers were phosphorylated and ligated and then reacted at a
concentration of 25ng/ml(lanes1, 3, and 5)or50ng/ml(lanes 2, 4,
and 6) with 30 p.l ofextract from SV40-infected cells. The marker lanes (M) contain a sample of each DNA substrate used in the
binding reactions. The polyacrylamide gelwas exposed to film for 18 h.
beled, self-ligated linkers. The latter included those linkers
used previously, namely BamHI, BclI, ClaI, and XhoI. First,we measured thecapacity of self-ligated XhoI linkers,
which contain repeats of the pentanucleotide, to compete
with the end-labeled Hinfl fragments ofpPH1-8 DNA (the
polyomavirus DNA-containingplasmid; Fig. 2A) in binding
reactions withanuclearpreparation of polyomavirus large T antigen (Fig. 9A). They reveal that the self-ligated XhoI linkers servedas very effectivecompetitorswith the
natural-ly occurring polyomavirus sequences in binding reactions
with polyomavirus large T antigen. By contrast, the
self-ligated BamHI, BclI, and ClaI linkers did not compete with polyomavirus DNA for thebinding ofpolyomavirus large T antigen, evenatconcentrationsatleast 10timesgreaterthan that required for effective competition with the self-ligated XhoI linkers (Fig. 9B).
We repeated this experiment with the same competitors
but withSV40 DNA as substrate andSV40 largeT antigen
(Fig. 10A and B). The results were the same as those
reported above. The self-ligated XhoI linkers competed effectively withSV40 DNA for the binding ofSV40 large T antigen, but none of the other self-ligated linkers did. In
addition, we also observed thatXhoI linkers which had not
beenligated didnotcompetewithSV40DNAforthebinding of theSV40largeTantigen (Fig.10A). Thissameresult was
obtained when monomers of the XhoI linkers were used as a competitor inbinding reactions with polyomavirus DNA and polyomavirus large T antigen (data not shown). This result was not unexpectedbecause the -GAGGC- sequence is only formed when the XhoI linkers are ligated to each other.
We also measured the capacity of the various self-ligated linkers to compete with SV40 DNA for the binding of polyomavirus large T antigen and vice versa. The results (Fig. 11) revealed that only the self-ligated XhoI linkers competed with SV40 DNA for the binding of polyomavirus large T antigen. This is indicated by the decrease in the intensity of theAvaII-HindIll2fragment that accompanied the inclusion of increasing quantities of self-ligated XhoI linkers in the binding reaction (Fig. 11A). Similarly, only the self-ligatedXhoI linkers competed effectively for the binding ofSV40 large T antigen to the Hinfl 4 fragment ofpPH1-8 DNA (Fig. 12). The latter DNAfragment contains bothsites of binding for polyomavirus large T antigen. These results strongly suggest that both large T antigens recognize and bind to the same sequences in eitherpolyomavirus orSV40 DNA and that a crucial featureof thesesequences isarepeat of the pentanucleotide -GAGGC-.
DISCUSSION
We tested the hypothesis that repeats of the sequence -GAGGC- form the recognition-binding site for polyoma-virus andSV40large T antigens. First, we showed that both large Tantigens bind with specificity to each other'sbinding sites. A common feature of the sequences comprising the large-T-antigen-binding sites in polyomavirus and SV40 DNA is repeats of the pentanucleotide -GAGGC-. Second,
wemeasured the capacity of both large Tantigens tobind to synthetic DNA substrates containing repetitionsofthe pen-tanucleotide and to controlDNAs thatdo not andfound that only those DNAs with the pentanucleotide repeatare
specif-icallybound by the large T antigens. Finally, weshowed that among the synthetic DNAs tested, only substrates
contain-ing repeats of the pentanucleotide effectively compete with
the natural DNA substrates (polyomavirus or SV40 DNA) for the binding of the two large Tantigens to either theirown
genome or theheterologous DNA. The extentofcompetition
iscomplete at high levels ofcompetitor DNA,indicatingthat the competitor can block the specific binding ofthe large T
antigens to all of the bindingsites in both polyomavirus and SV40 DNA. These data substantiate the hypothesis that repetitions of the -GAGGC- pentanucleotide form the recog-nition-binding site for the large T antigens ofpolyomavirus
andSV40. The sequences of the various DNAstowhichthe large T antigen bind are shown in Fig. 13.
Tjian (47) first proposed that the pentanucleotide was
important for the binding of the D2 hybrid large T antigen
(23) toSV40 DNA because the protein shielded the guanine
residues in this sequence from methylation by dimethyl
sulfate. Subsequently, measurements of the capacity of authenticSV40 large T antigen to bind tomutantSV40DNA
substrates confirmed theimportanceof the pentanucleotide.
For example, a deletion of 21 bp within binding site I (between nucleotides 5207 and 5186) in the cs1085 SV40 mutant genome drastically reduces the
capacity
of SV40large T antigen to bind to thatsite (10, 29, 45). This deletion removes a pair of -GAGGC- residues in binding site I
(Fig.
13B). Similarly, DNA of the mutantcs1088, which contains
multiple mutations in bindingsite Iwhichchangetheguanine
residues in both pentanucleotides to adenines, is also not
bound by large T antigen (29, 45). Finally, SV40 large T
antigen fails to bind tosite Iof the DNAoftheSV40mutant J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.612.122.244.71.321.2]d11086 (10). The genome of this mutant is deleted by 4 bp
(between nucleotides 5192 and 5197) and this results in the removal ofone ofthe -GAGGC- repeats in binding site I (Fig. 13B).
Although theoccurrenceofrepeatsofthe pentanucleotide is sufficient to account for the binding of SV40 large T antigen to sites I and II in SV40 DNA, this sequence does notoccurin site III. The latterisalow-affinitybinding site, and SV40 large T antigen is incapable of bindingtothis site when itis separated from sites I and11(9, 10, 29, 33, 43, 46,
47). Recently, DeLucia et al. (9) have identified 13 DNA pentanucleotide sites in SV40 DNA that areprotected from methylation by SV40 large T antigen and deriveda
consen-sus sequence for them (-G/T A/G GGC-).
Large-T-antigen-binding site III contains six ofthese pentanucleotides ar-ranged as direct repeats withina 59-bp span of DNA (Fig. 13B). It is likely that repetition of these weak
recognition-binding sequences (-TAGGC-, -GGGGC-,and-TGGGG-)in
binding site IIIallows for the stablebindingofSV40large T antigen only after a fraction of the protein has already covered sites I andII.
The -GAGGC-pentanucleotide alsooccurswithin
large-T-antigen-binding sites I and II in polyomavirus DNA (Fig.
13A). Here, the sequence -A/TGAGGC- is found repeated three times inbindingsite I andtwoorthreetimesinbinding siteII,dependinguponthe strain ofpolyomavirusexamined.
Interestingly, the firstnucleotide in each repeatisseparated from thesame nucleotide of theadjacentrepeat by 10or 11 bp. This arrangement ofrepeats would place them on the sameface of the DNA helix. Theimportance of hexanucleo-tiderepeatfor thebinding of largeTantigeninpolyomavirus DNA is supported by the results we have presented here.
However, there is nodirect evidence that largeTantigen of
polyomavirus contactsthese sequences inthebindingsites.
We examined the sequences comprising the
large-T-anti-gen-binding
sites in polyomavirus DNA for sequencesho-mologoustotheconsensusfamily derivedby DeLuciaetal. (9). This led tothe identification of three related sequences
which are located between the -A/TGAGGC- repeats in
binding
sites I and II (Fig. 13A). One such sequence,-TGGGG-(between nucleotides 128 and 134), forms a con-tinuation of therepeats presentinsite I (Fig. 13A). The other two sequences are separated from each otherby 10bp, but
theyoccuratadistancegreaterthan 10bpfrom the repeats insites I andII(betweennucleotides 92 and 107) (Fig. 13A).
This set ofrepeats may constitute a third
large-T-antigen-binding site in polyomavirus DNA that is located between those identified previously.
Interestingly, there are also four other pentanucleotides
homologoustotheconsensussequencederived byDeLucia et al. (9), arranged as tandem pairs on each DNA strand
(betweennucleotides 5286 and 14) inourstrain of polyoma-virus. These sequences are located within the core of the
polyomavirus origin for DNA replication (31). Moreover,
theirarrangement is remarkably similarto the arrangement
oftheconsensuspentanucleotides in binding siteIIofSV40
(Fig. 13B).InSV40, binding site II comprises thecoreofthe SV40 origin for DNA replication (1). Whether, the pentanu-cleotides in the polyomavirus origin for DNA replication
compriseyetanotherlarge-T-antigen-bindingsite remainsto
bedetermined. In this regard itmaybe noteworthy that the A2 strainofpolyomavirus does notcontain three of thefour repeatsalluded to previously.
Theaffinity of SV40 large T antigen for the bindingsitesin SV40 DNA isnearly thesameasthatforthebindingsitesin polyomavirus DNA.However,polyomaviruslarge T antigen apparently binds tightertothebinding sites in polyomavirus DNA thanto those in SV40 DNA. This interpretation rests
A B
M C Xho! C BamHI L Clal M
2 _ 2
3 _,
4 _ __ 3
5-7 4m4I. I 4
8 4' * t , g
5-10.11 8
12,13 10-11
14,15
16 14-15
17-20 41
1 2 3 4 1 2 3 4 5 6 7 8 9 10
FIG. 9. Competition between end-labeled polyomavirus DNA fragments and ligated, unlabeled synthetic linkers for reaction with polyomavirus largeTantigen.(A)Theend-labeled DNAfragmentsobtained afterHinfldigestion ofpPH1-8DNA (1ng/ml) were mixed with various concentrations of ligated, unlabeled XhoI linkers (lane 2, 100ng/ml;lane 3, 500
ng/ml;
lane 4, 1,000ng/ml),and were reacted with 30R1
of nuclearextractfrom polyomavirus-infected 3T6 cellsas described in the text. The marker lane (M) contains a sample of the end-labeled HinflfragmentsofpPH1-8DNA used as substrate. The control lane (C or lane 1) shows thefragmentsimmunoprecipitatedafter reaction of theend-labeled,HinflfragmentsofpPH1-8DNA with 30p.1 ofnuclear extract frompolyomavirus-infected 3T6cells in the absence of added linkers. The driedagarose gel was exposed to film for 6 h. (B) The end-labeled HinflfragmentsofpPH1-8DNA were mixed with unlabeledli-gatedBamHIlinkers(lane 1, 100 ng/ml; lane 2, 500ng/ml;lane 3, 1,000ng/ml),BclI linkers (lane 4, 100ng/ml;lane 5, 500 ng/ml; lane 6, 1,000 ng/ml), or Clal linkers (lane 7, 100 ng/ml; lane 8, 500 ng/ml; lane 9, 1,000 ng/ml) and were reacted with 30
,u1
ofnuclear extract from polyomavirus-infected 3T6 cells. The control lane (C) shows the fragments immunoprecipitated after reaction of the end-labeled Hinfl fragmentsofpPH1-8DNA with 30p.1of nuclear extract frompolyomavirus-infected3T6 cells in the absence of added linkers. The lane labeled Mcontainsasample of end-labeled Hinfl-cleavedpPH1-8DNA used as a substrate in the binding assays. The dried agarose gel was exposed tofilm for10 h.on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.612.155.470.432.609.2]934 POMERANTZ AND HASSEiL
A B
XhoI Xho
ligatod minomers -
-3J-~_
2|w_*3____
6j44
6
ii * g-t 1
lM1t 3 4 M n32 456789g
FIG. 10. Competition between labeledSV40 DNA fragments and unlabeled self-ligated synthetic linkers as substrates for binding to SV4O large T antigen. (A) The fragments obtained after
AvaII-HindIIIcleavage ofpSVOl DNA were endlabeled and mixed with various concentrations of unlabeled ligated XhoI linkers before reaction with 30 ,ul of nuclear extract from SV40-infected CV-1 cells.ThepSVOl DNA fragments were present at a final concentra-tion of 1 ng/ml. In lane 1, no linkers were added, whereas lane 2 contained ligated XhoI linkers at a concentration of 100 ng/ml. Lanes 3 and 4 contained unlabeled XhoI linkers that had not been self-ligatedaFt concentrations of 100 and 500nglml,respectively. The dried agarose gel was exposed to film for 10 h. (B) The AvaIl-HindIII digestion products of pSVO1 DNA were end labeled and mixed with various concentrations of self-ligated BamHI, BclI, or
ClaI linkers before incubation with 30 p.1 of nuclear extract from SV40-infected CV-1 cells. The labeled DNA fragments were present at aconcentration of 1 ng/ml in each reaction. The concentrations of unlabeled ligated BamHI linkers (lanes 1 to 3), unlabeled self-ligated BcII linkers (lanes 4 to 6), and unlabeled self-ligated ClaI linkers (lanes 7 to 9)were 100, 500, and 1,000ng/mlwithrespect to the three lanes of each. The lane labeled M contained a portion of the AvaII-HindIII fragments of pSVO1 used as substrate in the bindingreactions. The dried agarose gel was exposed to film for 16 h (lanes 1 to 9).However, the marker lane (M) was exposed to film for 3 h. The final photograph was prepared by splicing lane M to the others (lanes 1 to 9).
on the fragile assumption that the preparations of the two
largeT antigens whichwecompared contained approximate-ly equal numbers ofactive large-T-antigen molecules. If the
difference in affinities forthetwo DNA substrates displayed
bypolyomaviruslarge T antigen is real,then thismayreflect
itsrequirement for alargerrecognition-binding sequence in
DNA. The hexanucleotide -A/TGAGGC- is repeated in the
binding sites inpolyomavirus DNA. Bycontrast, the
penta-nucleotide -GAGGC-is repeated in thebindingsitesinSV40
DNA (Fig. 13). A plausible explanation for these
observa-tions is that polyomavirus large T antigen preferentially recognizes the hexanucleotide, whereas SV40 large T
anti-gen recognizes thepentanucleotide. Thishypothesis is sup-ported by the observation that SV40 large T antigen binds more readily to the self-ligated XhoI linkers containing repeats of thepentanucleotide
tha~n
does polyomavirus large T antigen.A single pentanucleotide or hexanucleotide is not likely the sole feature of the large-T-antigen-binding sites impor-tant for recognition and binding. These sequences occur
often in isolation
within SV40,
polyomavirus, andpBR322
DNA, but DNAfragments whichbearthem arenot
invari-ably
boundby large
Tantigen.
Forexample, the sequence-GAGGC- occurs a total of 14 times in SV40 DNA and is
representedsix times within theboundariesofbindingsites I and II in SV40 DNA. Similarly, this pentanucleotide is
presentatotal of 20times inpolyomavirusDNA andoccurs seventimes between the borders ofthetwoknown
large-T-antigen-binding
sites.Finally,
-GAGGC- occurs 15 times inpBR322 DNA; however, in noinstancedo these sequences occurclosetoeachother(the shortest spacing betweenany twoofthese pentanucleotidesis 55bp). These observations
suggestthat thespecific bindingof SV40 and
polyomavirus
large T antigen to DNA is facilitated by repeats of the
pentanucleotide or the hexanucleotide within the
binding
sites.The minimum numberof-GAGGC- repeats required for the efficientbindingofpolyomavirusorSV40largeT
antigen
to DNA islikelytwo. This is indicated by theobservations thatonlytworepeatsarerequiredfor the efficientbindingofpolyomavirus large T antigen to either binding site II in
polyomavirus DNA (34) ortobinding site I inSV40 DNA.
A B
M-.,...,--...-MTl
C|
f
Cl
a 1 Am 1 vm_ _ _ __ -__2
-Urn~ ~
2amai3-~~~~~
2 ow4
Dtw
3
_4
1234
_123456789
FIG. 11. Competition between labeledSV4O DNA fragments and unlabeled self-ligated linkers as substrates for binding to polyoma-virus large T antigen. (A) The fragments obtained after AvaII-HindIII cleavage of pSVO1 DNA were labeled and mixed with various concentrations of unlabeled self-ligated XhoI linkers before reaction with 30 p.1 ofnuclear extract from polyomavirus-infected 3T6 cells. In each reaction the concentration of labeled DNA fragments was 2 nglml, whereas the unlabeled self-ligated XhoI linkers werepresent at a concentration of 100 (lane 2), 500 (lane 3), or 1,000nglml (lane 4). Lane 1 (C) contained no added linkers and served as apositive control. The lane marked M contained a portion of the labeled DNA fragments which se'rved as substrate in the binding reactions. The dried agarose gel was exposed to film for 10 h. (B) The AvaII-HindIII digestproducts ofp5 VOl DNA were end labeled and mixed with various concentrations of unlabeled self-ligated BamHI, BcII, or CIaI linkers before they were reacted with 30 p.1 of nuclear extract frompolyomavirus-infected 3T6 cells. The labeled DNA fragments were present in the various reactions at a concentration of 2nglml. The self-ligated, unlabeled BamHI linkers (lanes 1 to 3), the self-ligated unlabeled BclI linkers (lanes 4 to 6), and the self-ligated unlabeledCIaI linkers (lanes 7 to 9) were present atconcentrations of 100, 500, and 1,000 ng/ml with respect to the three lanes of each. The lane marked M contained aportionof the labeled DNA used as substrate in thesereactions. The dried agarose gel wasexposed to film for 6 h.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.612.63.298.71.264.2] [image:10.612.331.547.326.498.2]M
C_
Bjarn H I Bci Cla Xh1o 1I M2 _
3 1 m
4
_Om ~~~~
qp5 -7 5 -a 'a
12.13 F 14,15
16 J
7 19b
20
1 2 3 4 5 6 7 8 9 10 11 12
FIG. 12. Competition between labeledpolyomavirusDNA
frag-ments andunlabeled self-ligated linkersas substrates forbindingto SV40 largeT antigen. The fragments obtained after Hinflcleavage ofpPH1-8DNAwereend-labeled and mixed with various
concen-trations of unlabeled self-ligated linkers before they were reacted with 30,ul of nuclear extractfromSV40-infected CV-1 cells.Ineach reaction the labeledDNAfragmentswerepresentat aconcentration of 2 ng/ml.Lane 1(C) shows the labeledDNAfragments whichwere
immunoprecipitated after reaction of the end-labeled Hinfl
frag-mentsofpPH1-8DNAwith 30p.lof nuclearextractin the absenceof added linkers. Unlabeledself-ligated BamHI linkers wereaddedto those reactions whose outcome are shown in lanes 2 to 4 at
concentrationsof100(lane 2), 500(lane 3), and 1,000ng/ml (lane 4). Unlabeledself-ligatedBcIl linkerswereaddedtoseveralreactionsat
concentrations of 100(lane 5),500(lane 6),and1,000ng/ml (lane 7).
Similarly, unlabeled self-ligatedClaI linkerswereaddedtoseveral reactions atconcentrations of 100 (lane 8), 500 (lane 9), and 1,000 ng/ml (lane 10). Finally, unlabeled self-ligated Xhol linkers were added tothereactions whoseoutcome areillustrated inlanes 11 and 12.Heretheconcentrations oftheself-ligatedXhoIlinkerswere100 (lane 11) and 250 ng/ml (lane 12). The lane markedM contained a portion of the end-labeledHinflfragments ofpPH1-8DNAused as
substrate inthevarious reactions. The driedgelwasexposedtofilm for 16h.
Similarly, SV40 large T antigen binds efficiently to binding site II in polyomavirus DNA and to binding site I in SV40
DNA. Each of these large-T-antigen-binding sites contains
the-GAGGC-pentanucleotide repeatedonly twice (Fig. 13).
Interestingly, the smallest ligated XhoI linker species to be bound by eitherpolyomavirus or SV40 large T antigen is a pentamer. Pentamers of the XhoI linkers contain four re-peatsofthe-GAGGC- sequence per DNAstrand(Fig. 13C). Ifonly a tandem duplication ofthe -GAGGC- sequence is
required forbinding,then weshould have observed binding of the twolargeTantigenstodimersortrimersoftheligated
XhoIlinkers. One explanation ofthisdiscrepancy isthatthe
incapacity ofthe large T antigens to bind to ligated XhoI
linkerssmaller than pentamers mayreflect their requirement foraminimumlength ofDNA(>40bp) for efficientbinding. Alternatively, the weak binding-recognition sequences de-scribed by DeLucia et al. (9) neighboring the -GAGGC-repeats inbinding site II in polyomavirus DNA and binding site I in SV40 DNA may contribute to the facility with which thelarge T antigens recognize and bind to these sites.
There appears to be a correlation between the spacing of the-GAGGC- sequences in the binding sites in SV40 DNA and theaffinity of SV40 largeT antigen for these sites. For example, theaffinity ofSV40 large Tantigen is greatest for
site I where 7
bp
intervene between thepentanucleotides
(Fig. 13B) (32, 48). However,siteII isboundpoorly by large
T antigen, and here the pentanucleotides are separated by
only 1bp (Fig. 13B). Bycontrast,the hexanucleotides which
arerepeatedin thelarge-T-antigen-bindingsites in
polyoma-virus DNA are allseparated by4or 5bp,and
polyomavirus
large T antigen binds with approximately equal affinity to both binding sites in polyomavirus DNA (Fig. 13A) (34).
This suggests that thespacingbetweentheputative recogni-tion sequences is important for the binding ofthe large T
antigens. Apparently when the distance between the first
base inadjacent pentanucleotidesis close to 10bp, then the
site is
tightly
boundby large
Tantigen.
Thisgeneralization
also applies to the weak recognition signals in site III of
SV40 DNA. Here the sequence -GGGGC- or -TGGGC- is
repeated five times, and the first base in each repeat is
separated
from thatsamebase in theadjacent
repeatby8 to11bp, dependingupon which pairsofrepeatsarecompared (Fig. 13B).
Therefore,
it seemslikely
that not only is the sequenceoftherecognition
siteimportant
for thebinding
oflarge T antigen but that the spacing between adjacent recognitionsites may also influence thetightnessofbinding.
Theobservationthatthe
large
Tantigens
ofpolyomavirus
and SV40 bind with
specificity
to common DNA sequencespromptedustosearch theamino acidsequence of thesetwo
proteins
forhomology in thevicinity
oftheirorigin-binding
domains. Initial attemptsto map the
origin-binding
domainofSV40
large
Tantigen
have ledtoconflicting
results(5, 6,
18, 35, 37-39, 41). However,
recently
Morrison et al.(30)
have reported that an amino-terminal
17,000-molecular-weight fragment ofSV40 large T
antigen
(amino
acids 1 to130) is capable of
specifically binding
to SV40 DNA.Be-causethe first 82 amino acids oflargeTantigenare shared
with small Tantigen, which does notbind to DNA, these
investigators
havetentatively
concluded that theorigin-binding
domain ofSV40 largeTantigen
is located between amino acids83 and 130.Comparable
studies ofpolyomavirus
largeT antigen have not been
performed.
Nonetheless, wecompared
theamino acid sequenceofSV40large
Tantigen
withthatofpolyomavirus largeTantigenacross the
origin-binding
domain ofSV40large
Tantigen (amino
acids 83 to130), after
alignment
of the two sequences to maximizehomology
between them. Theextentofrelatednessbetweenthe two
proteins
across theorigin-binding
domain is23%,
whichis less than thatdisplayedovertheentirelength ofthe molecules
(36%).
However, much ofthishomology
is clus-tered betweenamino acids124to130 inSV40large
Tantigen
andaminoacids 278to284in
polyomavirus large
Tantigen.
Here, six ofthe sevenamino acidsareidentical between the
two T
antigens.
Moreover, fourofthe amino acids in thisregion
of SV40 large Tantigen
are basic aminoacids,
whereas three in polyomavirus large T antigen are basic amino acids. Becausethetwo
large
Tantigens recognize
thesameorvery
closely
relatedDNAsequences,it islikely
that theprotein-DNA
contacts aremediatedby
thesidechains ofthesame orrelatedamino acidsin thetwo
proteins.
Thesidechains ofthe short stretch ofbasic amino acids described previouslyinpolyomavirusandSV40largeTantigenmay be thosewhichdirectlycontactthe-GAGGC-sequences in the viral genomes. Thissuggestioncanbe testedexperimentally.
Whatever the outcome of such experiments, it will also prove interesting to determine whether the two large T
antigens
canfunctionallysubstitute for each other in any ofthe activities which are unique to large T antigen (i.e., repression ofearly transcriptionandinitiation of viral DNA
replication).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.612.81.275.75.236.2]936 POMERANTZ AND HASSELL J. VIROL.
A
Polyomavirus
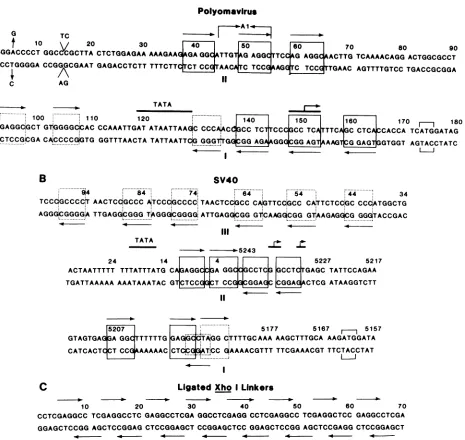
G TC
--f 10 20 30
40]
so~
[so
70 80 90GGGGACCCCTGGCCCGCTTA CTCTGGAGAA
AAAGAAGGA
GGCTTG
GAGGTCCGAGG
ACTTGTCAAAACAGG
ACTGGCGCCT CCCCTGGGGA CCGGGCGAAT GAGACCTCTT TTTCTTCTCTCAACAICTCCAAGGTC
TCCdTTGAAC AGTTTTGTCC TGACCGCGGAA
IC AG
TATA
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~....
1O0 110 120 '140 I5O 160 170 180
TGiGAGGC,GCT GT$GGGGQCAC CCAAATTGAT ATAATTAAGC
CCCAACC,GCC
TCTCCCGCCTCT
TTCAjGC CTCAjCCACCA TCATGGATAG [image:12.612.80.545.71.511.2]ACjCTCCGCGACA;CCCCGiGTG GGTTTAACTA TATTAATTCiG GGG*r1GEqG AGAAGGGIC AGT AGTGTGGT AGTACCTATC
...
T...
T...
AGG_AG
AAAGTAGA_T
G G T GT TB
SV4O
.. ...
* 4 84 74. 64 54 44 34
TCCCGGCCCC'T AACTCdGCCC AiTCCCiGCCCC.TAACTCCiGCC CAJGTTCCiGCC CA.TTCTCCGC CCOCATGGCTG AGGGiCGGGGA TTGAGGCGGG TAGGGiCGGGG.ATTGAGG,CGG GT'CAAG.CGG GGGGTACCGAC
TATA
-. - o-5243
24 14 4 5227 5217
ACTAATTTTT TTTATTTATG CA GAGGCCGA GGC CGCCTCG GCCTCTGAGC TATTCCAGAA TGATTAAAAA AAATAAATAC GT CTCCGGCT CCGaCGGAGC CGGAGACTCG ATAAGGTCTT
5207 5177 5167 5157
GTAGTGAG GA GG TTTTTG GA0GicAGG C'TTTTGCAAA AAGCTTTGCA AAGATGGATA
CATCACTCCT CC AAAAAC CT[ GTOCC GAAAACGTTT TTCGAAACGTTTCTACCTAT
C Ligated Xho ILinkers
10 20 30 40 50 60 70
CCTCGAGGCC TCGAGGCCTC GAGGCCTCGA GGCCTCGAGGCCTCGAGGCC TCGAGGCTCC GAGGCCTCGA
GGAGCTCCGGAGCTCCGGAG CTCCGGAGCT CCGGAGCTCC GGAGCTCCGG AGCTCCGAGG CTCCGGAGCT
FIG. 13. Nucleotide sequence of DNA substrates to which polyomavirus and SV40 large T antigen bind with specificity. (A) The nucleotidesequenceofthe A2strain ofpolyomavirusthatincludesthelarge-T-antigen-bindingsites is illustrated and numbered according to Soedaetal.(42). The strain ofpolyomavirusused in thisresearch, whichwerefer toasAl, differs insequencefrom that of the A2 strain at severalpositions. The A/Tbpatnucleotide5isreplaced byaG/Cbpin theAlstrain, and the Al strain containsaninsertion of 2bpbetween nucleotides14and15.Moreover,theAl strain is deleted of10bp by comparisontothe A2 strain between nucleotides 44 and 54 (shown in the figure by the verticallinesbearingarrows). The dark lines above thesequenceshow the borders of the -TATA- box and theposition of the5' termini of the abundantearly mRNAs (denoted bythe dark line and the vertical lines bearing an arrow) (15). The initiation codon for translationoftheearlymRNAsisbracketed. Boxessurroundtherecognition-bindingsequence,-A/TGAGGC-. The dotted boxes contain the weakrecognition-bindingsequence(-TGGGC-,-TAGGC-,-GGGGC-)identifiedbyDeLuciaetal.(9). Thearrowsaboveorbelow the boxes depicttheorientationoftherecognition-bindingsequenceoneach DNAstrand.(B)The nucleotidesequencescomprisingtheSV40 large-T-antigen-binding sitesareshown. Thesymbolsusedarethesameasthosedescribed above.(C)The nucleotidesequenceofligatedXhoI linkers (-CCTCGAGG-) is shown. The symbols used are the same asthose described above. However, the boxes wereomitted from the figure becausetheywould have cluttered thedrawing. Note thatthe -GAGGC- sequence highlighted byahorizontal arrow ononeDNA strand partially overlaps with anotheron theopposite DNAstrand.
ACKNOWLEDGMENTS LITERATURE CITED
1. Bergsma, D. J., D. M. Olive, S. W. Hartzell, and K. N. We thank Monica Naujokas and Cathy Collins for excellent Subramanian.1982.Territorial limits and functionalanatomyof technical assistance and acknowledge the contributions of Ron the simian virus 40 replication origin. Proc. Natl. Acad. Sci. McKay with whom webegantheseexperimentsin 1981. U.S.A. 79:381-385.
This research wassupportedby theMedicalResearchCouncil of 2. Bradley, M. K., J. D. Griffin, and D. M. Livingston. 1982. Canada and the National Cancer Institute of Canada. J.A.H. is a Relationship of the oligomerization to enzymatic and DNA research associate of the National CancerInstitute of Canada. binding propertiesof theSV40largeT-antigen.Cell 28:125-134.
on November 10, 2019 by guest
http://jvi.asm.org/
3. Buchman, A. R., L. Burnett, and P. Berg. 1980. The SV40
nucleotide sequence, p. 799-823. In J. Tooze(ed.),DNAtumor
viruses, 2nd ed. Cold Spring Harbor Laboratory, Cold Spring
Harbor, N.Y.
4. Carroll,R.B.,L.Hager,and R.Dulbecco. 1974.SV40T-antigen
bindstoDNA. Proc. Natl. Acad. Sci. U.S.A. 71:3754-3757.
5. Chaudry, F.,R. Harvey, and A. E. Smith. 1982. Structure and
biochemical functions of four simian virus 40 truncatedlargeT
antigens. J. Virol. 44:54-66.
6. Clark, R., K. Peden, J. M. Pipas, D. Nathans, and R. Tjian.
1983. Biochemical activities ofT-antigen proteinsencoded by
SV40A genedeletion mutants. Mol. Cell. Biol. 3:220-228.
7. Cogen, B. 1978. Virus-specific earlyRNA in 3T6 cells infected
byatsA mutant ofpolyomavirus. Virology85:222-230. 8. Cowan, K., P. Tegtmeyer, andD. D. Anthony. 1973.
Relation-shipofreplicationandtranscriptionofSV40 DNA. Proc. Natl.
Acad. Sci. U.S.A. 70:1927-1931.
9. DeLucia, A., B. A. Lawton,R. Tjian, and P. Tegtmeyer. 1983.
Topography ofSV40 A protein-DNA complexes: arrangement
ofpentanucleotide interaction sitesatthe originofreplication.
J. Virol. 46:143-150.
10. DiMaio, D.,and D. Nathans.1982. RegulatorymutantsofSV40.
Effect of mutationsataTantigenbindingsiteonDNA
replica-tion andexpression of viralgenes. J. Mol. Biol. 156:531-548.
11. Fanning,E.,C.Burger,and E. G. Gurney.1981. Comparisonof
T-antigen-associationhostphosphoproteinsfromSV40-infected
andtransformed cells of differentspecies.J.Gen. Virol.
55:367-378.
12. Fanning, E., B. Novak, and C. Burger. 1981. Detection and
characterization ofmultiple forms of simian virus 40 large T
antigen.J. Virol. 37:92-102.
13. Friedmann, T.,A.Esty,P.Laporte,and P.Deininger.1979. The
nucleotide sequence and genome organizationof the polyoma
early region: extensive nucleotide homology and amino acid
homologywith SV40. Cell 17:715-724.
14. Gaudray,P.,P.Clertant,and F.Cuzin. 1980. ATP
phosphohy-drolase(ATPase) activityofapolyomavirusT-antigen. Eur. J.
Biochem. 109:553-560.
15. Gaudray, P., D. Tyndall, R. Kamen, and F. Cuzin. 1981. The
high affinity binding site on polyomavirus DNA for the viral
largeTprotein. Nucleic Acids Res. 9:5697-5710.
16. Giacherio, D., and L. P. Hager. 1979. A poly (dT)-stimulated
ATPaseactivity associated with SV40 largeT antigen. J. Biol.
Chem. 254:8113-8116.
17. Gidoni, D., A. Scheller, B. Barnet,P. Hantzapoulos, M. Oren,
andC. Prives. 1982. Different forms of simian virus 40 largeT
antigen varyingin their affinities for DNA. J. Virol. 42:456-466. 18. Gluzman, Y.,and B. Ahrens. 1982. SV40earlymutants thatare
defectivefor viral DNAsynthesisbutcompetentfor
transforma-tion of culturedrat and simian cells. Virology 123:78-92.
19. Greenspan, D. S., and R. B. Carroll. 1981. Complex ofSV40
large T-antigen and 48,000-dalton host tumor antigen. Proc.
Natl. Acad. Sci. U.S.A. 78:105-109.
20. Griffin,B. E. 1980. Structure andgenomic organizationofSV40 andpolyoma virus, p. 61-123. In J. Tooze (ed.), DNA tumor
viruses, 2nd ed. Cold Spring Harbor Laboratory, Cold Spring
Harbor, N. Y.
21. Griffin,J.D., G.Spangler,and D.M.Livingston. 1979. Protein
kinase activity associated with SV40 T-antigen. Proc. Natl.
Acad. Sci. U.S.A. 76:2610-2614.
22. Harlow,E.E.,D. C.Pim,and L. V.Crawford.1980.Complexof
simian virus 40 large-T antigen and host
53,000-molecular-weight protein inmonkeycells. J. Virol. 37:564-573.
23. Hassell, J., A.Lukanidin, G. Fey,andJ. Sambrook. 1978. The
structure and expressionoftwodefective adenovirus 2/simian
virus 40hybrids. J.Mol. Biol. 120:209-247.
24. Hassell, J. A., C. Mueller, A.-M. Mes, M. Featherstone, M.
Naujokas,B.Pomerantz,and W. Muller.1982. The construction ofpolyoma virus vectors: functions required forgene
expres-sion, p.71-77. In Y. Gluzman(ed.), Eukaryotic viral vectors.
ColdSpring HarborLaboratory, ColdSpring Harbor, N.Y. 25. Hassell, J. A., W. C. Topp, D. B. Rifkin, and P. E. Moreau.
1980.Transformation ofratembryofibroblastsbycloned
poly-oma virus DNA fragments containing only part of the early
region. Proc. Natl. Acad. Sci. U.S.A. 77:3978-3982.
26. Lane,D.P., and L. V.Crawford. 1979. T-antigenis boundto a host protein in SV40-transformed cells. Nature (London) 278:261-263.
27. McCormick, F., and E. E. Harlow. 1980. Association of a murine53,000-daltonphosphoproteinwith simian virus 40 large-Tantigenin transformed cells. J. Virol. 34:213-224.
28. McKay, R. D. G. 1981. Binding of a SV40 T-antigen related protein toDNA.J. Mol. Biol. 145:471-488.
29. McKay,R.,and D. DiMaio.1981.BindingofanSV40T antigen-relatedproteintothe DNAofSV40regulatory mutants.Nature (London)289:810-813.
30. Morrison, B., M. Kress, G.Khoury, and G.Jay. 1983. Simian virus 40 tumor antigen: isolation of the origin-specific DNA-bindingdomain. J. Virol. 47:106-119.
31. Muller, W. J., C. R. Mueller, A.-M. Mes, and J. A. Hassell. 1983. Thepolyomavirus originfor DNAreplicationiscomprised ofmultiple geneticelements. J. Virol. 47:586-599.
32. Myers, R. M., D. C. Rio, A. K. Robbins, and R. Tjian. 1981. SV40gene expressionis modulated bythecooperative binding ofT-antigen toDNA. Cell 25:373-384.
33. Myers, R.M.,and R.Tjian. 1980. Construction andanalysisof SV40 originsdefective in tumorbindingand DNAreplication. Proc. Natl. Acad. Sci. U.S.A. 77:6491-6495.
34. Pomerantz, B. J., C. R. Mueller, and J. A. Hassell. 1983. Polyomavirus large T antigen binds independently to multiple unique regionson the viral genome. J. Virol. 47:600-610. 35. Prives, C.,B. Barnet,A.Scheller,G.Khoury,andG.Jay.1982.
Discreteregions of simian virus 40largeTantigenare required fornonspecific and viralorigin-specific DNA binding.J. Virol. 43:73-82.
36. Prives, C., Y. Beck, G.Gidoni, M.Oren, andH. Shure. 1980. DNA-bindingand sedimentationproperties ofSV40T-antigens synthesized in vivho and in vitro. Cold Spring Harbor Symp. Quant.Biol. 44:123-130.
37. Prives, C., Y.Beck, and H. Shure. 1980. DNA-binding
proper-ties of simian virus 40 T antigens synthesized in vivo and in vitro. J. Virol. 33:689-696.
38. Rundell, K.,J. K.Collins,P.Tegtmeyer,H. L.Ozer, C.J. Lai, and D. Nathans. 1977. Identification of simian virus 40 protein A.J. Virol. 21:636-646.
39. Scheller, A., L. Covey, B.Barnet, and C. Prives. 1982. A small subclass ofSV40T antigenbinds tothe viralorigin of replica-tion. Cell 29:375-383.
40. Shalloway, D., T. Kleinberger, and D. M. Livingston. 1980. MappingofSV40DNAreplication origin region bindingsitesfor the SV40T-antigen by protection againstexonuclease III diges-tion. Cell 20:411-422.
41. Shortie, D.,R. F.Margolskee,andD.Nathans. 1979.Mutational analysisof theSV40replicon: pseudorevertantsofmutantswith a defective replication origin. Proc. Natl. Acad. Sci. U.S.A. 76:6128-6131.
42. Soeda, E., J. R. Arrand, N. Smolar, J. E. Walsh, and B. E. Griffin. 1980. Coding potential and regulatory signals of the polyomavirus genome. Nature (London)283:445-453. 43. Tegtmeyer, P., B. Anderson, S. B. Shaw, and V. G. Welson.
1981. Alternative interactions of theSV40Aproteinwith DNA. Virology 115:75-87.
44. Tegtmeyer,P.,M.Schwartz,J. K.Collins,and K.Rundell.1975. Regulationoftumor antigen synthesis by simian virus 40gene
A.J. Virol. 16:168-178.
45. Tenen,D.G.,L. L.Haines,andD. M.Livingston. 1982. Binding ofananalogof theSV40T-antigentowild-typeandmutantviral replication origins.J. Mol. Biol. 157:473-492.
46. Tjian,R. 1978. Thebindingsite onSV40 DNAfora T-antigen-relatedprotein. Cell 13:165-179.
47. Tjian, R. 1978. Protein-DNA interactionsattheorigin ofSV40 DNA replication. Cold Spring Harbor Symp. Quant. Biol. 43:655-662.
48. Tjian, R. 1981. T-antigenbindingand the control ofSV40gene
expression. Cell 26:1-2.
49. Tjian,R.,and A.Robbins. 1979.Enzymaticactivitiesassociated with a purified SV40 T-antigen-related protein. Proc. Natl. Acad. Sci. U.S.A. 76:610-614.