p. 0022-538X/93/052672-09$02.00/0
Copyright ©1993, American SocietyforMicrobiology
Relating
Structure
to
Function in the
Hepatitis
Delta
Virus
Antigen
DAVID W. LAZINSKIANDJOHN M. TAYLOR*
FoxChase Cancer Center, 7701 Burholme Avenue, Philadelphia, Pennsylvania 19111
Received 29 October1992/Accepted20January1993
Hepatitisdeltavirusexpressestwoformsofasingle protein,the small(BAg-S) andlarge (bAg-L) antigens,
whichare identicalexceptforan additional19residues presentatthe C terminus ofbAg-L.WhilebAg-Sis
requiredto promotegenomereplication,bAg-L potentlyinhibits thisprocessandalso facilitatespackagingof the viral genome by envelope proteins of the helper virus (hepatitis B virus). Regions within the antigens responsiblefornuclearlocalization,RNAbinding,and dimerization have beenidentified,yetit isnotclear how theseparticularactivities contributetothe ultimatereplicationandpackagingphenotypes.Herewereport the following findings. (i) Although theremovalof the nuclear localizationsignalfrom eitherantigenresulted in significant cytoplasmic accumulation, both proteins still had access to the nucleus. As a consequence, no
functionaldefectwasobservedwith eithermutant.(ii)TheRNA-binding domain, althoughnecessaryforbAg-S function,could be deleted frombAg-Lwithoutcompromisingitsabilitytoeither inhibitreplicationorpromote
packaging. (iii) In contrast, the coiled-coil dimerization domain was required for both the activation of
replicationbybAg-Sand theinhibition of replication by bAg-L. This region,withanadditional 20 amino acids
C-terminal to it, was necessary and sufficient to potently inhibit replication by interacting with the small antigen. (iv) The packaging propertyofbAg-L required aC-terminal Pro/Gly-rich regionwhich is
hypothe-sizedtointeractwith thehepatitis Bvirus envelope proteinsduringthe assemblyprocess.
Hepatitis delta virus (HDV) is a novel human subviral
pathogen which is able to infect only persons who were
previously or are simultaneously infected with hepatitis B virus (HBV) (27). The HDV genome is a single-stranded
circular RNA of approximately 1.7kbwhich isableto fold into an unbranched rod structure, in which 70% of its nucleotidesareinvolvedinintramolecular pairing (31).HDV
is fully capable ofgenome replication in the absence of all
helpervirusfunctions; however, the hepadnaviral envelope proteins (s antigens) arenecessarytopromotethe assembly and release of infectious virions (17, 26, 27). Replication
occursinthenucleus,where ahost-encoded, RNA-directed
polymerase isthought to synthesize the complement of the
genome, termed the antigenome, which then serves as the template forgenome synthesis (30).
HDV encodes a single protein, the 195-amino-acid (a.a.)
delta antigen (MAg-S), which functions in the nucleus to promotegenomereplication(17). Currently, the mechanism
underlying thistrans activation is notunderstood, noris it
knownatwhatstageinthereplication cycletheantigenacts. From both invitro and in vivostudies, however, it is clear thatbAg-S bindstothe HDV rod-structuredRNAtoforma
ribonucleoprotein (RNP) which is found both within the nucleus and in virions (5, 8, 29). Although only bAg-S is synthesized early in infection, a second antigen variant,
bAg-L, isobservedlater ininfection (1, 31, 32). This protein is identical to bAg-S except that it has an additional 19
carboxy-terminal residues which result from the mutationof thebAg-Sambercodonas aconsequenceofRNAediting(3,
22, 35). This editingevent is essential for viralpropagation because, unlike bAg-S, bAg-L is able to function with the hepadnaviral envelopeproteinstodirect the packaging of the HDV RNP structure into virions (4, 28). bAg-L differs phenotypically from bAg-S in a second way; not only is it
* Correspondingauthor.
unable to support genome replication, but it acts as a
trans-dominant inhibitor of this process. When bAg-L is expressed to only 1/10 the level of bAg-S, the number of
genomesobserved is reduced almost 10-fold(7).Thisdegree of potency can be explained if a single bAg-L molecule interacts withamultimericcomplex,suchasthe HDVRNP, somehowrendering it incompetentforreplication yet
com-petentforpackaging.
How does the addition of 19 a.a. to the C terminus of bAg-Sresult insuch drasticphenotypic changes?Part of the
answer has recently been discovered by Glenn et al. (13), whofound that thesolecysteine residue ofbAg-L (position 211; not present in bAg-S) is isoprenylated. Furthermore, when this residue is mutated to serine to prevent the modification, packaging is abolished. The role of the
car-boxy-terminal 19a.a. in inducingtrans-dominant inhibition is less clear, however. For instance, an 11-a.a. C-terminal
truncation mutant that lacks the isoprenylation site and should not therefore be packaged is still able to inhibit replication, indicatingthat thetwophenotypes of bAg-Lmay
be separable (14). This possibility has been confirmed by Chenetal.,whodescribedamutantwhich iscompetentfor packagingbutunableto inhibitreplication (10).
There is agrowing bodyofgenetic evidence concerning
thelocations ofanumber offunctional domainswithin the
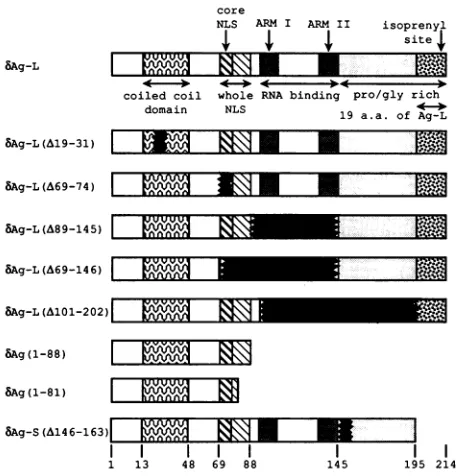
deltaantigenprimarysequence(Fig.1).Alocale
encompass-ing residues 89 to 163 has been shown, in vitro, to direct RNA binding (21). Within this region are two Arg-rich motifs, sequences that promote RNAbinding in a class of
proteins which includes bacteriophage antiterminators as
wellasthe humanimmunodeficiencyvirus Tat and Revtrans
activators (19). Although RNAbindingis thoughtto be an
essentialactivity requiredfor thefunction ofbAg-S,this has
notyetbeenestablishedexperimentally,andtheimportance of this region tothephenotypesofbAg-Lisunknown.
An adjacent region, a.a. 67to 88,constitutes the nuclear localization signal (NLS) (33). Residues 69 to 75 can be 2672
on November 9, 2019 by guest
http://jvi.asm.org/
8Ag-L
MAg-L(A19-31)
core
NLS ARM I ARM II isoprenyl
M| X X site4,
coiled coil whole RNA binding pro/gly rich
Alom>I CT
SAg-L(A69-74)
I EP.
U ISAg-L(A89-145)
I
|1IJArtJf
8Ag-L (A69-146)
8Ag-L(AlOl-202) |
-8Ag(1-88)
I
m
8Ag(1-81) 1
8Ag-S(&4-6) "I" I I
1 13 48 69 88
FIG. 1. Diagram of the structural domains
and mutantsconstructed for thisstudy.Deleted
outoromittedineachdiagram. ARM, arginint
considered the core NLS, as they disp
targeting activity in the absence of adc whilea.a.85to88arereportedtoenhance Although even this core sequence is noi
HDV bAg isolates, we note here that it
consensus, K(K,R)X(K,R), and all isolate consensus(9). Although both bAg-S and #
tofunction in thenucleus, there hasbeen this work, to investigate the significance relationto the function of the deltaantige Indirect evidenceindicatesthattheresii nal tothe NLS (1 to57) facilitatedimeriz this region, an LX6LX6LX6L motifwas
the authorstosuggestthat theantigenuse
of coiled-coil structure known as the dimerize. More recently, both sequence
additionalHDVisolatesand mutationalan
the deltaantigenhasno strictrequirement dposition of the helix, as istypical forle
Nevertheless, this region very likely dc
coiled coil. Acomputeralgorithm predicts 99%confidencethat,ataminimum,residu
thisstructureand that the coiled coilprob atposition13(23).The role of dimerizatioi phenotypes of bAg-S and bAg-L is unkr least part of the coiled-coilregionof bAg-tobedispensable forpackaging (10).
To date, no function has been ascrib
structural domain which spans a.a. 145 t
remainder of the protein, the region is n
predictedtolackanyhelical structure,an4
andglycine.Wesetouttoboth elucidateI
of thisregionand determine how the othei
ute to the ultimate phenotypes of the
antigens.Discreteregionswithin theconte
were individually deleted, and the result
tested for their ability to dimerize in viv
nucleus,
trans activate or inhibitreplication,
and becomepackaged bythe HBV smalls antigen (sAg). MATERIALSANDMETHODS
19 a a. of Ag-L Recombinant plasmids. The wild-type small and large delta
antigen expression
vectors,pDL444
andpDL445,
respec-tively, were constructed by inserting the antigen openread-ing
frame(ORF)-containing
ScaI-BamHIfragments
from pSVL(Ag-S) and pSVL(Ag-L) into theSmaI
andBamHI
sites of a plasmid composed of the NdeI-simian virus 40 E..Jfs! -0XtJ
;'.(SV40)
latepromoter-SmaI region
ofpSVL
and the NdeI-vectorbackbone-SmaI region of pGem4Z (7). The8Ag(1-88)-producing
plasmid pDL439
wasgenerated by
insertion of the deltaORF-containing ScaI-StuI fragment into the SmaI site ofpSVL. Thisconstruct wasthenrestricted with BamHI and blunted with Klenowfragment, and the synthetic NheI linker 5'-TAGCTAGCTAGCTA-3'wasinserted into the site soastointroduce stop codons in all three reading frames.In addition to the first 88 a.a. of the delta antigen, bAg(1-88) contains six aminoacidsat itscarboxy terminus(GELGSS) EIZ777 |whichare derived from thepolylinker region.
Thisplasmid
l l l wasthen usedtogeneratethe
SAg(1-81)-expressing
plasmid145 195 214 pDL476. The antigen ORF-containing
SacII-AccI
fragment of the delta antigen was blunted at the AccI site with Klenow fragment and portionsareblacked inserted into theSacII-blunted
NheI backbone of pDL439. e-rich motif.bAg(1-81)
is extended by an additional 2 a.a. (A andS)
at its C terminus. In order to delete 13 a.a. from the coiled-coil region, apGem4Z derivative that contains the HDV BglII-ilay some nuclear EcoRI fragment inserted between the BamHI and EcoRI litional sequence, sites was restricted with XhoI and religated to create a this function(33). 39-nucleotide deletion. Thedeletion-containingSacII-EcoRI t conserved in all fragment from this vector was then inserted into the same includes an NLS sites withinpDL444 and pDL445togeneratethebAg-S(A19-esconform to this 31)- and bAg-L(A19-31)-producing plasmids pDL448 and 6Ag-Larethought pDL449, respectively.
noeffort, priorto The 8Ag-L(A1l1-202)-expressing plasmid pDL497 was of this region in constructed by excising and purifying the antigen ORF-ns. containing EcoRI-BstXI fragment from pDL445, cleaving duesamino-termi- this with Sau3A, and thenreligating the products with the ation (33). Within pDL445EcoRI-BstXIvectorbackbone. Thesameapproach noted, prompting wasusedto constructpDL499,the
bAg-L(A89-145)-produc-s aparticulartype ingvector, aswell as the
8Ag-L(A69-146)-expressing
plas-leucine zipper to midpDL503 except that thepurifiedEcoRI-BstXI fragment information from wascleavedwith StuI and NaeI forpDL499and withHaeIII alysissuggestthat andNlaIV for pDL503. pDL499was used to constructthe tfor leucine in the bAg-S(A89-145) producer pDL501 by inserting its antigen -ucinezippers (6). ORF-containingEcoRI-SmaI fragment into the same sites Des function as a within pDL444. pDL503 was used in the same manner toswith greater than generate the bAg-S(A69-146) expressor pDL505. In the tes21to48exist in processoffusingtheNlaIV sitetoHaeIIIinpDL503,a new iablystartsearlier, ApaI sitewascreated 18 nucleotides upstream and in frame
nin thereplication with theApaI site that naturallyresides within the antigen
iown,
although at ORF.Thus,theBAg-S(A69-74)-
andbAg-L(A69-74)-produc-L has been shown ing plasmids pDL507andpDL508were
generated by
intro-ducing theApaI-BstXIfragments
frompDL444andpDL445,
'ed
to a potential respectively, into the same sites withinpDL503. pDL500,
to 214. Unlike the whichproduces
8Ag-S(A146-163),
wasgenerated
by
purify-lot hydrophilic, is ing the ORF-containing EcoRI-NaeI fragment of
pDL444
d is rich inproline andligatingit into thepDL444EcoRI andSmaIsites. thefunctional role The plasmid which initiates HDV
replication, pDL481,
rdomains contrib- comprises the NdeI-SV40 late
promoter-Sacl fragment
of small and large pSVL and the SacI-vector backbone-NdeIfragment
of -xtof eachantigen pGem4Z. Within thepGem
polylinker,
the1.1-genome-ting mutants were length SalI-SalI-XbaI fragment of
pSVL(D2M)
has been ro, localize to the inserted in theantigenomic
orientation with respect to theE FJLl* X|l w-
-...
l-aomain Nilb
I aM
L
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.60.288.74.312.2]SV40promoter
(17).
The HBV serotype AYWsAg
expres-sion clonepMSVgptSMwaskindly provided by
C.Mangold
and contains the
XhoI-sAg ORF-BglII fragment
cloned into theBamHI site ofpNI2
(16).
DNA transfections and sample harvest. For all
experi-ments, the human
hepatoma
cell line Huh7 was grown in Dulbecco's modifiedEagle's
mediumsupplemented
with 10% fetal bovine serum and transfected atapproximately
80% confluence with calcium
phosphate-precipitated
DNA(15).
Forpackaging, immunofluorescence,
andcross-linking
experiments, samples
were harvested at 3days
posttrans-fection andat6
days
fortransactivation andtransinhibitionstudies, respectively.
Forprotein analysis, samples
werelysed
andresuspended
withLaemmlisamplebuffer(18),
andRNA was isolated by a modification of the acid
phenol-guanidinium-thiocyanate procedure (12).
SecretedHBVsAgparticles
were recovered from the medium as describedpreviously (28). Briefly,
10 ml of medium was clarified bylow-speed centrifugation,
and theparticles
werepelleted by
high-speed centrifugation (32,000
rpm, 16h, 4°C) through
2ml of20% isotonic sucrose. In vivo
cross-linking
was doneby
a modification of the method ofZapp
et al.(34).
The medium wassupplemented
with 0.3%methanol,
10 mMNaCl,
0.1 mMEDTA,
and0.01%glutaraldehyde
and addedtothe cells for 20 minat
37°C,
followedby
a3-hincubation at4°C.
Cross-linked cells were washed and then directlylysed
andresuspended
insample
buffer.Immunoblot
analysis.
Proteinsamples
were subjected toelectrophoresis
on 14%polyacrylamide gels by
the methodof Laemmli
(18). They
were then transferred tonitrocellu-lose
electrophoretically,
and deltaantigen
was detected byusing
rabbitpolyclonal
antiserum raisedagainst bacterially
expressed antigen
andby
incubation with'25I-labeled
pro-tein A
(Du
Pont).
RNA analysis. Total cell RNA was
separated
electro-phoretically
on1.5%agarosegels
in the presense ofethidiumbromidesothat both the
quality
andquantity
of RNA in each lane couldbemonitored. RNAwasthenelectrophoretically
transferred to a
nylon membrane,
and successful transferwasconfirmed
by
visualization with UVlight.
RNAwasUV cross-linked to the membrane and thenhybridized
with a radiolabeled RNAprobe
specific
for the genomic strand as describedpreviously
(11).
Immunofluorescence assays. Cells were transfected and thenfixedon
coverslips by
treatmentwith-20°C
methanol for 3 min and then with-20°C
acetonefor 30s. Fixed cellswere incubated for 2 h at room temperaturewith a rabbit
polyclonal antibody
raisedagainst
recombinant deltaantigen
expressed
inEscherichia
coli,
washed,
and then incubatedfor 2 h at room temperature with
rhodamine-conjugated
protein
A(Boehringer
Mannheim).
RESULTS
Rationale for design of delta antigen mutants and their associated activities. Our
goal
was todetermine the contribu-tions that thecoiled-coil,
NLS, RNA-binding, and Pro/Gly-richdomains make in the function of the deltaantigens.Thisquestion
wasaddressedby individually deletingeachregionwithin both
bAg-S
andbAg-L
and thentestingthedeletion's effectontheability
of theprotein
tolocalizetothenucleus,oligomerize,
support or inhibit replication, and associate with HBVsAg
in secreted particles. Figure 1 depicts the series ofmutants constructed for this study. The mutantswere
designed
to delete thefollowingsequences: coiled-coildomain,
bAg(A19-31);
nuclear localizationsignal,bAg(A69-74); RNA-binding domain, bAg(A89-145); RNA-binding do-main plus NLS, bAg(A69-146); Pro/Gly-rich region, bAg(A146-163); RNA-binding domain plus Pro/Gly-rich re-gion, bAg(A101-202); RNA-binding domain plus Pro/Gly-rich region plus isoprenylation site, bAg(1-88) and bAg(1-81). In most cases, a given deletion was created in both bAg-L and bAg-S; however, only the bAg-L mutants are diagrammed. The deletion mutation spanning residues 145 to 163 was introduced into both bAg-S and bAg-L, but its effects on the stability of the two proteins were very differ-ent.bAg-S(A145-163)wasreadily detectable, whereas bAg-L(A145-163)was either undetectable orbarely detectable, dependingonthe timeposttransfection that itwasanalyzed. Forthis reason,noconclusions about thephenotypes of this mutantcould be drawn.
All proteins were expressed from SV40 late promoter-derived vectors via transient transfection of the human hepatoma cell line Huh7. The mutantswerethenassayed for relevant phenotypes.
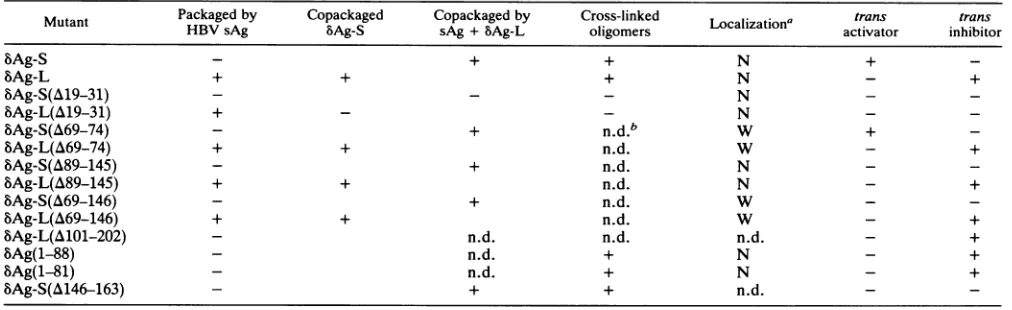
The ability of a protein to localize to the nucleus was assessed by immunofluorescence. Glutaraldehyde cross-linkingofintactlivingcellswasusedtodetermine whethera mutantcouldmultimerizeinvivo. Theabilityof eachmutant tosupport genomereplicationwasassayed by cotransfecting aplasmid thattranscribesanantigenomicHDVRNAwhich contains a frameshift mutation in the antigen ORF and requiresfunctionalbAg-Sintransin ordertoreplicate.Total cellular RNA was then subjected to Northern (RNA blot) analysiswithaprobespecificfor theproductofreplication, i.e., HDV genomic RNA. trans-Dominant inhibition of rep-licationwasdetermined in the same manner except that in addition to the HDV RNA and mutant protein-producing constructs,cellsweretransfected withanequivalentamount of a vector which expresses wild-type bAg-S. A protein's abilitytobepackaged bythehepadnaviral envelopeproteins was determined by cotransfecting a plasmid that produces the HBVsAg. Secreted sAg particleswerethen isolated and subjectedto Western immunoblot analysis with delta anti-gen-specificantiserum.Asummaryof thephenotypes of the variousmutantsisgiven inTable 1.
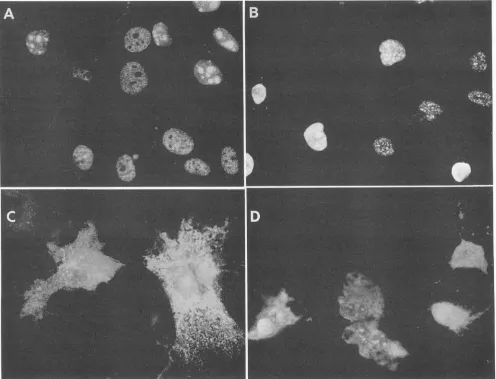
Corenuclear localization signalisdispensableforthe func-tionofbAg-SandBAg-L. Both forms of the deltaantigenare
reportedtoshow three distinct nuclear immunofluorescence stainingpatterns(33).Withmostcells, apunctatenucleolar patternof localization isobserved,in whichboth the nucleoli andadditional subnuclearstructuresshowpreferential stain-ing. However, in some cells, both the nucleoplasm and nucleoli arestained,whereas inothers, antigenis localized tothenucleoplasmand excludedfrom the nucleoli. We have also observed these three patterns, and the first and third typescanbe seeninadjacentcellsin Fig. 2A. Currently, we have no explanation for these differences in subnuclear localization, nordoweunderstand theirrelevance,if any,to the HDV lifecycle.Consistent with earlierfindings,we were unabletodetectdifferences in the localization of bAg-S and bAg-Leither in thecontextof thewild-type proteinsorwith thedeletionmutantsmentioned below.
Itwaspreviously shown that residues 1to 88 ofthe delta antigenweresufficienttopromote nuclearlocalization when fusedto acytoplasmic protein (33). Notsurprisingly, there-fore,ourbAg(1-88)mutantlocalizedtothe nucleus with the same staining pattern as the wild-type protein. However, aminoacids 85to88arereportedtoenhance the function of thecoreNLS,yetwhen theseresiduesweredeleted [bAg(1-81)],weagainobservedwild-typelocalization(Fig. 2B) (33).
Aswould beexpected, when the core NLS was removed
on November 9, 2019 by guest
http://jvi.asm.org/
TABLE 1. Antigen mutants and their characteristics
Mutant Packaged by Copackaged Copackaged by Cross-linked Localization' trans trans
HBV sAg bAg-S sAg +bAg-L oligomers activator inhibitor
bAg-S + + N +
-bAg-L + + + N - +
bAg-S(A19-31) - N - _
SAg-L(A&19-31) + - - N _ _
bAg-S(A69-74) + n.d.b W +
bAg-L(A69-74) + + n.d. W - +
bAg-S(A&89-145) + n.d. N
-bAg-L(A89-145) + + n.d. N - +
bAg-S(A&69-146) + n.d. W
-bAg-L(A69-146) + + n.d. W - +
bAg-L(A101-202) - n.d. n.d. n.d. - +
bAg(1-88) - n.d. + N - +
bAg(1-81) - n.d. + N - +
bAg-S(A146-163) - + + n.d.
-aN, nuclear;W, whole cell.
bn.d.,notdone.
in bAg-S(A69-74) and bAg-L(A69-74), a significant amount ofprotein wasseeninthe cytoplasm(Fig. 2C and D). These mutants were not excludedfrom the nucleus, however, and in fact, some cells showed preferential staining of the nucle-oli (Fig. 2C and D). The presence of a portion of the NLS (a.a. 75 to 88) does not account for this residual nuclear staining,asbAg(A69-146)localized in the same manner (data notshown).
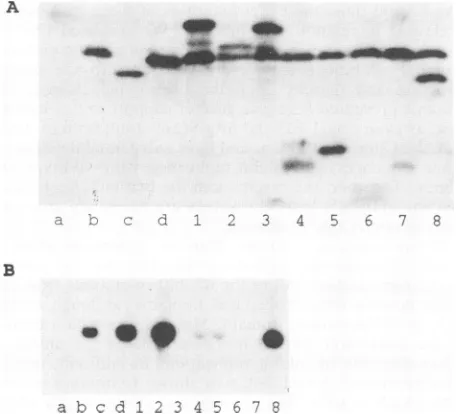
Theremaining nuclear access of the NLS deletion mutants isnot atrivial issue, given their surprising phenotypes. In the trans activation assay, anHDVcDNA clone was unable to initiatereplication, asassessedby the detection of genomic RNA in a Northern blot, unless functional bAg-S was supplied in trans (Fig. 3B, lanes a andb). The presence or absence of bAg-S was confirmed by Western blot analysis (Fig. 3A, lanes a and b). As shown in Fig. 3B, lane d, bAg-S(A69-74) was fully capable of supporting genome replication withwild-type efficiency. Inthetrans inhibition assay, the cotransfection of a bAg-L-expressing plasmid with aplasmid expressing bAg-S and HDV RNA abolished replication (compare lanes b and 1 of Fig. 3B). bAg-L(A69-74) displayed wild-type activity in its ability to inhibit replication (Fig. 3B, lane 3). Furthermore, in a packaging assay,bAg-Lcanbe secreted into the tissue culture medium if HBV sAg is also expressed, and bAg-L(A69-74) was
similarlyfoundtobeefficientlypackaged into sAg particles (Fig. 4, compare lanes b and B with lanes d andD).
Coiled-coil regionneeded forbothtransactivation of repli-cation by bAg-S and trans inhibition by bAg-L. A program designedtorecognizecoiled-coil domains inprotein primary sequences predicts that the deletion ofa.a. 19 to 31 in the delta antigen will abolish the protein's capacity to assume this structure (23). This deletion was introduced into both bAg-S andbAg-L, and its effecton dimerizationwas evalu-ated bytwo independent methods. When cells whichwere
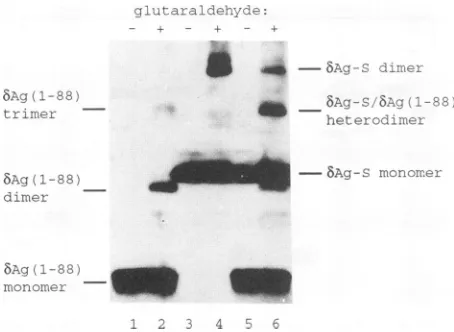
transiently expressing wild-type bAg-Sweresubjectedto in vivocross-linking, a significantamount ofaproductthat is consistent in size with a dimer and whose production is dependent on the addition ofglutaraldehyde was observed (Fig. 5, lane4).The assignment of this novelspeciesas an
bAg-S
dimerwas further substantiated by results obtainedwith the coiled-coil-containing truncation construct bAg(1-88). Glutaraldehyde-treated cellswhich expressed this pro-tein did not produce a product of the same size as the bAg-S-expressing cells but rather two smaller
products
whose sizes are what would be expected for bAg(1-88) dimers and trimers (Fig. 5, lane 2). Furthermore, in addi-tional experiments with gels containing a lower percentage ofacrylamide, dimers and trimers of both wild-type bAg-S andbAg-L could be resolved (datanotshown). These results clearly established that both wild-type proteins as well as bAg(1-88) oligomerized in vivo. However, when cells pro-ducing bAg-L(A19-31) were treated in the same manner, only monomeric proteinwasvisualized (Table 1).
The inability of bAg-L(A19-31) to oligomerize was also confirmed by packaging experiments. Previously, it was noted thatalthough bAg-S isnotpackaged into hepadnaviral particles per se, it iscopackaged if bAg-L is also present in the cell(28). This observationwasinterpreted as indicatinga directprotein-proteininteraction betweenbAg-S and bAg-L. Consistent with thisnotion,when the coiled-coil domainwas
disruptedinbAg-S(A19-31),theproteinwas notcopackaged by bAg-L(Table 1). Likewise, although bAg-L(A19-31) did package itself, itwas unableto copackagebAg-S (Table 1). Inadditiontoprovidingfurther evidence that deletion ofa.a. 19 to 31 inactivates the coiled coil, this result confirms a previous finding, that the packaging (although not the co-packaging) phenotype of bAg-L is independent of dimeriza-tion (10). Disruption ofthe coiled-coil domain completely inactivated both the trans inhibition and trans activation phenotypes, as bAg-L(A19-31) did not interfere with repli-cation andbAg-S(A19-31)wasunabletopromote it(Fig. 3B, lanes 2 and c).
Inhibition of replication by bAg-L does not require an RNA-binding domain but functions via interaction with
BAg-S. Toexplain the potency ofbAg-Lin inhibiting repli-cation, we proposed that the protein interacts with and "poisons" the replicating delta virus RNP structure (7). bAg-Lmight use its RNA-bindingdomaintogain access to this RNP. However,anumber ofourmutantsindicated that the regionwhich binds RNAisnot neededfor inhibition of replication. bAg(1-88), bAg(1-81), S(A89-145), bAg-L(A89-145), bAg-S(A69-146),
bAg-L(A69-146),
and bAg-L(A101-202)allpotentlyinhibitedreplication, although
none contain this domain (Table 1 andFig. 3B, lanes4to 7).How then do these proteins interact with their target? Since all of the mutants contain the coiled-coil
domain,
aregion whichwehavejustshown isrequiredfor
inhibition,
it seems likelythattheycould form heterodimers withbAg-S,
which in turn would bind HDVRNA. This
possibility
wason November 9, 2019 by guest
http://jvi.asm.org/
FIG. 2. Immunofluorescence analysis of the localization of wild-type and mutant delta antigens. Huh7 cells were fixed 3 days posttransfectionand incubated witharabbitpolyclonal antibodyraisedagainstrecombinant deltaantigen expressedin E.coli.(A) BAg-S;(B) 8Ag(1-81); (C)bAg-S(A69-74); (D) MAg-L(A69-74).
verifiedby in vivocross-linking. When cellsexpressingboth
5Ag-S
and bAg(1-88) were cross-linked, a uniquehet-erodimericspecieswasobserved whichwas not seenincells that expressed either protein individually (Fig. 5, compare lanes2, 4, and6). Although this heterodimeric species(289 a.a.) is similar in predicted size to and nearly comigrated withthe8Ag(1-88) trimer (282 a.a.),it could be discernedby its relative abundance. While the bAg(1-88) trimer was far less abundant than the dimer, the 8Ag(1-88)/8Ag-S het-erodimerwas inexcessof either homodimericspecies (Fig. 5, compare lanes 2 and 6). This result is in excellent agreement with what would be predicted theoretically. If bAg-S andbAg(1-88) arepresentinequimolar amounts and theenergetics of homodimer and heterodimer formation are equivalent, then a 1:2:1 ratio of bAg-S dimer to bAg-S/ bAg(1-88) heterodimer to bAg(1-88) dimer would be
ex-pected.
Packaging of bAg-L into HBV sAg particles requires a C-terminalPro/Gly-rich domain. Of the aforementioned mu-tants which potently inhibited replication, several were unabletobepackaged byHBVsAg. For instance,bAg(1-88)
was notfound in sAgparticles; however, when the Pro/Gly-richregion spanningresidues 146to214wasappended toit [bAg-L(A89-145)], packagingwasrestored(Table1andFig. 4, laneF). Similarly, a mutantwithanevenlargerdeletion, bAg-L(A69-146), was both packaged and competent in its abilitytocopackage bAg-S(Fig.4, laneG). Since thebAg-S version of the a.a. 89 to 146 deletion both lacked the isoprenylation site andwas notpackaged, the restoration of packaging seen with the addition of a.a. 146 to 214 to
bAg(1-88) mightsolely reflect the requirement forthis site. Another mutant,bAg-L(A101-202), however, indicated that the Pro/Gly-rich region was also needed, as this mutant contained the isoprenylation site butwasstillnotpackaged (Fig. 4, laneE).
DISCUSSION
Wehave systematically deleted domains within the delta antigens in order to determine their contributions to the various functions of these proteins. This approach has the general limitation that it is possible, in deleting a given
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.66.562.75.454.2]A
- 4b
: .40
'Rm momo
--- L 1 2 4 5 6 7 8
a
B
[image:6.612.66.294.75.282.2]a b c d l 2 3 4 5 6 7 8
FIG. 3. transactivation and transinhibition of replication. (A)
Western blot analysis of delta antigen and muntant proteins
ex-pressed in Huh7 cells at 6 days posttransfection and probed with rabbit polyclonal anti-delta antigen antiserum. Thepredicted sizes
of allproteinswereingoodagreementwith actual sizes, as
deter-mined by comparison with nonradiolabeled, prestained protein
standards. (B) Northern blot analysis of genomic HDV RNA
syn-thesized inthepresenceofwild-type andmutantantigens in Huh7
cells at 6 days posttransfection. RNAwasprobed with uniformly
labeled, in vitro-generated antigenomic RNA transcribed by using T7 RNApolymerase fromapGEM4Z derivative containing trimeric HDVcDNA. Lanesatod, samples tested fortransactivation; lanes 1to8, samples tested fortransinhibition. All sampleswereisolated
from cells whichwere transfected withequal amounts ofavector
which produces antigen ORF frameshift-containing antigenomic
RNA(pDL481); the samplesinlanes1to8wereisolated from cells whichwereadditionally transfected withabAg-S expression
plas-mid. Lanes: a, no-protein control; b, bAg-S; c, bAg-S(A19-31); d, bAg-S(A69-74); 1, bAg-L; 2, bAg-L(A19-31); 3, bAg-L(A69-74); 4, bAg-L(A101-202);5, Ag-S(A89-145); 6, bAg(1-81); 7, Ag-L(A69-145); 8, Ag-S(A146-163). As assessed by theintensityof ethidium
bromide-stained RNAon the membrane after transfer, less RNA
was loaded in lane b than in the other lanes, while lane 2 was somewhat overloaded.
domain, to alter the structure of the protein so as to affect other regions. The loss of a function associated with the
deletion ofadomain could indicate that domain's direct role in the process or may only reflect the inactivation of a
second critical domain. For this reason, deletion mutants which maintain function arefarmore informative, asit can
beconcluded,without ambiguity, that thestructure
respon-sible for that activity has remained intact. As a
multifunc-tionalprotein,the deltaantigenhasprovedtobeparticularly amenable to this approach, since it has been possible to
abolish one function while preserving others. All of the major conclusions that arise from this workwere obtained by usingmutantswhich retain the function of interest. These conclusionscanbe summarizedasfollows.(i)ThecoreNLS isdispensablefor the replicationandpackagingactivities of bAg-SandbAg-L. (ii)Neither thepackaging norreplication inhibition activities ofbAg-L require the RNA-binding do-main. (iii) Replication inhibition by bAg-L occurs via a
protein-protein interaction with bAg-S. (iv) bAg-L uses a
FIG. 4. Packaging and copackaging assay; Western blot analysis of proteins expressed within cells and secreted from cells in the presenceof HBV sAg. Lanes a to g show 1/100 of the cell protein from a 100-mm petri dish, and lanes A to G show 1/10 of the secreted materialisolated from tissue culture medium from the same sam-ples. Lanes: a, bAg-L(A19-31); b, bAg-L plus bAg-S(A69-74); c, bAg-L(A69-74)plus bAg-S; d, bAg-L(A69-74) plusbAg-S(A69-74); e,5Ag-L(A101-202);f,bAg-L(A89-145)plus bAg-S; g, bAg-L(A69-146)plusBAg-S. The predicted sizes of each protein were in good agreementwith actualsizesdeterminedby comparison with nonra-diolabeled, prestained protein standards.
C-terminal Pro/Gly-rich region in its association with the hepadnaviral envelope proteins.
We deleted six of the seven amino acids which were previously identified asthe core of the nuclear localization signal(33), andasexpected,thesemutantsdisplayedaltered compartmentalization and were present in both the cyto-plasm and the nucleus. Additionalmutantsdemonstrate that theremainingamino acids of the NLS (a.a. 76to88)are not responsible for the residualaccess tothe nucleusdisplayed by thea.a. 69to74deletions. Surprisingly, and presumably as a consequence of this ability to reach the nucleus, bAg-S(A69-74) is fullycompetent insupporting replication, whereas BAg-L(A69-74) inhibits replication with wild-type efficiencyand ispackaged byHBVsAg. The small andlarge versions of BAg(A69-74) may be entering the nucleus by passive diffusion, as the nuclear pore is thought to be permeable to molecules smaller than 50 kDa (25). Alterna-tively, since both proteins were present in the nucleoli of somecells,it remainspossiblethat there isasecondaryNLS withinthefirst 68 a.a. Consistent with thispossibility,Chang etal.recently reported that sequences within the first 50a.a. of the deltaantigencan, undersomecircumstances, directa fusionprotein tothe nucleus(6).
So far, we have observed no defect in function after deletion of thecoreNLS.However, since several aspects of the viral life cycle have not been assayed in this study, it would be prematuretoconclude that these residuesaretruly dispensable. During the onset of infection, theviral RNP, having just entered the cell and shed its envelope, is ex-pectedtobefartoolargetofreelydiffusethroughthe nuclear pore. Itseemspossible,therefore,that thecoreNLS maybe required to facilitate this process.
A number of mutants have enabled us to delineate the domain which governs the
ability
ofbAg-L
tobepackaged
by the hepadnaviral envelope
proteins.
Since both bAg-L(A19-31)andbAg-L(A69-146)
areefficiently packaged,
wecanconclude that the residues which have been deletedare
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.324.555.75.225.2]8Ag 1-83) tri-
--glut raldehr1 de:
t - + - +
5
het
_rroi-ner
, het er^olami
-iAgrl-8&)mnomeer8Ag-S
dir-mer
Ag(1-88)
monomer
1 2 3 4 58
FIG. 5. Western blotanalysis of invivo-cross-linked bAg-S and bAg(1-88). Samplesineven-numbered laneswereisolated fromcells
for which 0.01% glutaraldehyde was added to the tissue culture medium,andsamplesinodd-numbered laneswerenotcross-linked.
Lanes: 1and2, bAg(1-88);3 and4, bAg-S;5 and6,bAg(1-88) plus
bAg-S. Predicted sizes: bAg(1-88) monomer, 94 a.a.; bAg(1-88) dimer, 188 a.a.; bAg(1-88) trimer, 282 a.a.; bAg-S monomer, 195
a.a.; bAg-S/Ag(1-88) heterodimer, 289 a.a.; bAg-S dimer, 390 a.a. The predicted sizeswere in good agreement with actual sizes as
determinedbycomparisonwithnonradiolabeled,prestained protein
standards.
not involved in this process. Recently, Chen et al. (10) similarly showed that bAg-L(A33-73) is proficient in this
process.Fromtheseresults,we canconcludethata.a. 19to
146 are dispensable for packaging. Of the remaining resi-dues,wehave shownadirectrolefora.a.146to214.When thisdomain isappendedtoapackaging-deficientmutant,the wild-typepackaging phenotypeisrestored.
Boththeaminoacid contentand thepredicted secondary structure ofthisdomainareverydifferentfromthe
remain-derof theantigen. Morethan39% oftheaminoacids inthis region are either proline or glycine, yet these residues
constitute less than 12% of the amino-terminal portion. While residues 1 to 145 are extremely hydrophilic and
predicted to include several a-helices, a.a. 146 to 214 are
relatively nonpolar and are thought, because of the high
content ofproline and glycine, to be unableto assumethis
structure. These properties are consistent with a domain
which issequestered fromthe solvent, possibly by interac-tion with another protein. Although there is, as yet, no
evidence that demonstrates the existence of an bAg-L/sAg
protein interface, it is difficult to imagine how the large antigen could be so efficiently and selectively packaged without such an interaction taking place. Wepropose that thePro/Gly-rich regioninteractsdirectlywiththe HBVsAg
tofacilitatepackaging.
Itshouldalso benotedthatall ofthepackaging-competent
mutantspresentedcontain the19C-terminalresidues unique
to bAg-L, underscoring the importance of this region. The relevance of these 19 a.a. to replication inhibition is less obvious, however. Previously, we hadobserved that when 84heterologousamino acidswereappended tothe C
termi-nusofbAg-S,orwhenanE. coliprotein (MalE)wasfusedto its amino terminus, the inhibitory phenotype was elicited
(20). Neitherprotein containedanyofthe 19a.a. uniqueto
bAg-L, and thus, the exact identities of these residues are irrelevant in relation to inhibition. We surmised that the C-terminal 19 a.a. aswell as the other amino- and
carboxy-terminal alterations act sterically to perturb the protein
structure and thereby elicit the phenotypic change. The mutantspresented here give further support to this
hypoth-esis, as even bAg(1-81) and 8Ag-S(A69-146), both ofwhich lack the C-terminal 19a.a. and have substantial deletions of other sequences, still inhibit replication with wild-type effi-ciency. Together, the results with the two mutants indicate that the antigen's first 68 residues are necessary and suffi-cientto conferthis phenotype.
Within residues 1 to 68, there is a domain which is predicted to assume a coiled-coil structure that can potenti-ate multimerization. All of the inhibitory mutants from this study contain intact coiled-coil domains, although several lack the RNA-binding domain. Mutants in which a portion of the coiled-coil domain has been deleted are unable to either dimerize or inhibit replication. In addition, one in-hibitory mutant, bAg(1-88), was shown to interact directly with bAg-S in vivo. The potency and stoichiometry ofthe inhibition exhibited by bAg-L can be explained if its target is a complex containing multiple bAg-S molecules. From a theoretical analysis of inhibition data, it was calculated that this complex should include 23 bAg-S monomers (7). Recently, Ryu et al. found that, based on equilibrium density centrifugation analysis, the HDV nuclear RNP contains approximately 25 bAg-S molecules, consistent with the notion that this structure is the target for bAg-L inhibition (29). The results presented here demonstrate that bAg-L (and the other trans-dominant inhibitor mu-tants) do not require the RNA-binding domain to access the RNP but can act via a coiled-coil interaction with bAg-S.
Ag-L could act through either a passive or active mecha-nism to poison the HDV RNP complex. In the passive model, any and allproteins which are themselves unable to support replication but can still interact with bAg-S would have access to the RNP and, by their mere presence, inhibit replication. This possibility is negated by the results ob-tained withbAg-S(A146-163), which does not support repli-cation and does not inhibit replirepli-cation but does have a functional coiled coil (Table 1). The active model asserts that the inhibitor should not only interact with bAg-S but also mustcontain an activated function which interferes with the replication process.
What might this inhibitory function be? In addressing this question, we should make three points. First, there is no evidence to suggest that bAg-L directly prevents either the initiation, elongation, or maturation steps of HDV replica-tion. All that is known is that, in the presence of bAg-L, fewerreplicative products are observed. Thus, bAg-L could work through an indirect mechanism. Second, there is currently a gapin our understanding of how the HDV RNP, which is observed in the nucleus, can encounter and become packagedby the exclusively cytoplasmic hepadnaviral enve-lope proteins. Finally, there are a number of shuttle proteins, suchas No38, nucleolin, and Noppl40, whichshow exclu-sively punctate nucleolar localization by immunofluores-cence(2, 24). Despite this staining pattern, these proteins are notrestricted to the nucleus. More sensitive techniques have been used to demonstrate that the proteins can "piggyback" larger molecules associated with them in and out of the nucleus (2, 24). All three of these proteins are phosphory-lated and contain potentially complex nuclear localization signalswith multiple NLS consensus sites. Similarly, bAg-L
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.67.294.72.238.2]is a nucleolar phosphoprotein which has abipartiteNLS (5,
33).
With these points in mind, we speculate that bAg-L can escort theHDVRNPfrom the nucleustothecytoplasm. A nuclear export signal might reside within the first 68 a.a. of the antigen, which would be silenced in bAg-S but conformationally activated upon addition of the C-terminal 19 a.a. in bAg-L. bAg-L, through a coiled-coil interaction with the small antigen, would bind to the HDV RNP and move it to the cytoplasm,wherethe large antigen's isopre-nylated Pro/Gly-rich region could interactwith HBV sAgto induce packaging. This model also accounts for the trans-dominant inhibition simultaneously caused by bAg-L. As HDV RNP structures are removed from the nucleus, the nuclear pool of these species would be depleted, and as a consequence, replication would be reduced. Thus, a single-molecule mechanism, nuclear export, can account for the superficially different phenotypes of packaging and replica-tioninhibition. A direct investigation of the ability of bAg-L toexit the nucleus will be required to testthe validity ofthis model.
ACKNOWLEDGMENTS
D.W.L. wassupportedbypostdoctoral training grant CA-09035 and individualfellowship1F32 AI08637-01 from the National Insti-tutes for Health. J.M.T. was supported by grants CA-06927, RR-05539, and AI-26522 from the National Institutes for Health, by grantMV-7Q fromtheAmericanCancerSociety, and by an appro-priation fromtheCommonwealthof Pennsylvania.
Wethank C.Mangold (Universityof Mainz, Mainz,Germany) for kindlysupplying the HBV sAg vector pMSVgptSM and A. Lupas andJ.Stock (Princeton University, Princeton,N.J.)forgenerously providingthecoiled-coil predictingprogram. Wethank C. Seeger, R. Katz, W. Mason, W.-S. Ryu, and H. Netter for their valuable criticalreadingof themanuscript.
REFERENCES
1. Bergmann,K.F., and J. L.Gerin. 1986. Antigens of hepatitis delta virus in the liver andserum of humans and animals. J. Infect. Dis.154:702-706.
2. Borer, R.A.,C.F.Lehner, H. M. Eppenberger, and E. A. Nigg. 1989. Major nucleolar proteins shuttle between nucleus and cytoplasm.Cell 56:379-390.
3. Casey, J. L., K. F. Bergmann, T. L. Brown, and J. L. Gerin. 1992.Structural requirements forRNAeditinginhepatitisdelta virus: evidence for a uridine-to-cytidine editing mechanism. Proc.Natl. Acad. Sci. USA 89:7149-7153.
4. Chang,F.L., P.J. Chen, S. J. Tu, M. N. Chiu, C. J. Wang, and D. S. Chen.1991. Thelargeformofhepatitis8antigeniscrucial for the assembly ofhepatitis 8 virus. Proc. Natl. Acad. Sci. USA88:8490-8494.
5. Chang, M.-F.,S.C.Baker, L. H.Soe,T.Kamahora,J. G. Keck, S.Makino, S.Govindarajan,and M. M. C. Lai. 1988. Human hepatitisdelta antigen isa nuclear phosphoproteinwith RNA binding activity. J.Virol. 62:2403-2410.
6. Chang, M.-F., S. C. Chang, C.-I. Chang, K. Wu, and H.-Y. Kang. 1992. Nuclear localization signals, but not putative leucine zipper motifs, are essential for nuclear transport of hepatitisdeltaantigen. J. Virol. 66:6019-6027.
7. Chao, M., S.-Y. Hsieh, and J. Taylor. 1990. Role oftwoformsof thehepatitis delta virusantigen: evidence for amechanism of self-limitinggenomereplication. J.Virol. 64:5066-5069. 8. Chao, M., S.-Y. Hsieh, and J. Taylor. 1991. The antigen of
hepatitis deltavirus: an examination of in vitro RNA-binding activity.J. Virol. 65:4045-4062.
9. Cheisky,D., R. Ralph, and G.JonaL 1989. Sequence
require-mentsfor synthetic-peptide-mediated translocation to the
nu-cleus. Mol.Cell. Biol. 9:2847-2492.
10. Chen, P.-J., F. L. Chang, C. J.Wang, C. J. Lin, S. Y. Sung, and D.-S. Chen. 1992.Functional study of hepatitis deltavirus large antigen in packaging and replication inhibition: role of the amino-terminal leucine zipper. J. Virol. 66:2853-2859. 11. Chen, P.-J., G. Kalpana, J.Goldberg, W.Mason, B. Werner,J.
Gerin, and J. Taylor. 1986. Structure and replication of the genome of hepatitis 8 virus. Proc. Natl. Acad. Sci. USA 83:8774-8778.
12. Chomczynski, P., and N. Sacchi. 1987. Single-step method of RNAisolation by acid guanidinium-thiocyanate-phenol-chloro-formextraction. Anal. Biochem.162:156-159.
13. Glenn, J. S., J. A. Watson, C. M. Havel, and J.0.White. 1992. Identification of a prenylation site in the delta virus large antigen. Science256:1331-1333.
14. Glenn, J. S., and J. M. White. 1991.trans-Dominant inhibition of human hepatitis delta virus genome replication. J. Virol. 65:2357-2361.
15. Graham, F. L., and A. J. Van der Eb. 1973. A new technique for theassay ofinfectivity of human adenovirus 5 DNA. Virology 52:456-467.
16. Israel, N., N.Chenciner, C.Houlmann,and R. E.Streeck 1989. Expression vectors for high-level protein synthesis in Vero cells. Gene 81:369-372.
17. Kuo, M. Y. P., M. Chao, and J. Taylor. 1989. Initiation of replication of the human hepatitis delta virus genome from cloned DNA: role of delta antigen. J. Virol.63:1945-1950. 18. Laemmli,U. K. 1970. Cleavage ofstructural proteins during the
assembly of the head ofbacteriophage T4. Nature (London) 227:680-685.
19. Lazinski, D., E. Grzadzielska, and A. Das. 1989. Sequence-specificrecognition of RNA hairpins by bacteriophage antiter-minators requires a conserved arginine-rich motif. Cell 59:207-218.
20. Lazinski, D., and J. Taylor. Unpublished observations. 21. Lin, J.-H., M.-F. Chang, S. C. Baker, S. Govindarajan, and
M. M. C. Lai. 1990.Characterization of hepatitis delta antigen: specific binding to hepatitis delta virus RNA. J. Virol. 64:4051-4058.
22. Luo, G., M. Chao, S. Hsieh, C. Sureau, K. Nishikura, and J. Taylor. 1990. A specific base transition occurs on replicating hepatitis delta virus RNA. J. Virol.64:1021-1027.
23. Lupas, L., M. VanDyke, and J. Stock. 1991. Predicting coiled coils from protein sequences. Science252:1162-1164.
24. Meier, U. T., and G. Blobel. 1992.Noppl40shuttles ontracks between nucleolus and cytoplasm. Cell70:127-138.
25. Paine, P. L., L. C. Moore, and S. Horowitz. 1975. Nuclear envelope permeability. Nature (London) 254:109-114. 26. Ponzetto, A., P. J. Cote, H. Popper, B. H. Hoyer, W. T. London,
E. C. Ford, F. Bonino, R. H. Purcell, and J. L. Gerin. 1984. Transmission of the hepatitis B virus-associated 8 agent to theeasternwoodchuck. Proc. Natl. Acad. Sci. USA 81:2208-2212.
27. Rizzetto, M., B. Hoyer, M. G. Canese, J. W. K. Shih, R. H. Purcell, and J. L.Gerin. 1980.8 agent:association of 8 antigen with hepatitis Bsurface antigen and RNA in serum ofb-infected chimpanzees. Proc. Natl. Acad. Sci. USA 77:6124-6128. 28. Ryu, W.-S., M. Bayer, and J. Taylor. 1992. Assembly of
hepatitis delta virus particles. J. Virol. 66:2310-2315.
29. Ryu, W.-S., H. J. Netter, M. Bayer, and J. Taylor. Ribonucleo-proteincomplexes of hepatitis delta virus. J. Virol., in press. 30. Taylor, J. 1990. Hepatitis delta virus: cis and trans functions
needed forreplication. Cell 61:371-373.
31. Wang, K.-S., Q.-L. Choo, A. J. Weiner, J.-H.Ou,C.Najarian, R. M.Thayer, G. T.Mullenbach, K. J. Denniston, J. L.Gerin, and M. Houghton. 1986. Structure, sequence and expression of thehepatitis delta viral genome. Nature (London) 323:508-513.
32. Weiner, A. J., Q.-L. Choo, K.-S. Wang, S.Govindarajan,A. G. Redeker, J. L.Gerin, and M. Houghton. 1988. A single antige-nomic open reading frame of the hepatitis delta virus encodes the epitope(s) of both hepatitis delta antigenpolypeptides p24 and p27. J. Virol.62:594-599.
on November 9, 2019 by guest
http://jvi.asm.org/
33. Xia, Y.-P., C.-T. Yeh, J.-S. Ou, and M. M. C. Lai. 1992. Characterization of nuclear targeting signal of hepatitis delta antigen: nuclear transport as a protein complex. J. Virol.
66:914-921.
34. Zapp, M. L., T. J. Hope, T. G. Parslow, and M.R.Green. 1991. Oligomerization and RNA binding domains of thetype1 human
immunodeficiency virus Rev protein: a dual function for an
arginine-rich binding motif. Proc. Natl. Acad. Sci. USA 88: 7734-7738.
35. Zheng, H., T. B. Fu, D. Lazinski,and J. Taylor.1992. Posttran-scriptional modification of genomic RNA of human hepatitis delta virus. J. Virol. 66:4693-4697.