JOURNAL OF VIROLOGY, May 1975, p. 1262-1266 Copyright 01975 American Society for Microbiology
Vol.15,No. 5 Printed inU.S.A.
NOTES
Morphology
of
BHK-21
Cells Infected with Sindbis Virus
Temperature-Sensitive
Mutants
in
Complementation Groups
DandE
DENNIS T. BROWN* AND JONATHAN F. SMITH
Institutfur Genetik der Universitdt zuKoln,5Koln41, Weyertal121, West Germany
Receivedforpublication 18December1974
BHK-21 cells infected with temperature-sensitive mutantsofSindbis virus in
complementationgroupsDand E differed in theirappearance under
nonpermis-sive conditions. Cells infectedatnonpermissivetemperaturewith virus defective incomplementationgroupE hadnucleocapsids attached in large numberstothe
inside surface of the host plasma membrane. Infection with a group D mutant
produced nucleocapsids that didnot attachtothe plasma membrane but rather
remained free in the cell cytoplasm.
The morphogenesis of the alpha-togaviruses invertebrate cellscanbe dividedintotwomajor
processes: (i) the assembly of the viral
nu-cleocapsid from the capsid protein and single-stranded RNA, and (ii)the envelopmentof the
completed nucleocapsid by the process of
bud-ding through the virus-modified
plasma
mem-brane. Little morphological information iF
available
regarding
theassembly
of the viralnucleocapsid sincenoobvious intermediates in
assemblycanbe detectedinthe electron micro-scope. On the otherhand, the buddingprocess attheplasma membrane hasbeendescribed
by
using various electron
microscopic
techniques
(1, 2).
The modified plasma membranesofSindbis
virus-infected cellscontainthreevirusproteins,
PE2, El, and E2 (7, 16). PE2 is cleaved toE2 during virus
morphogenesis,
and thiscleavage
seems tobe essentialforthe
budding
process(7,
12-14, 16). Enzymatic radio-iodination studies
reveal that El and E2 are exposed on the cell surface, whereas PE2 cannot be iodinated and ispresumablyburied in the cellmembrane (16).
To characterize further the final stages of
Sindbis virus maturation, we studied cells
in-fected with temperature-sensitive mutants (3,
4) by electron microscopy. Those mutants
which produce complete nucleocapsids but no
mature virus were of particular interest, since these mutants are likely to be defective in the
budding process. The two complementation
groups that meet these conditions are D and E
(4), and we have chosen as representative of
these groups temperature-sensitive mutants ts-23 and ts-20 (obtained from E. R.
Pfeffer-korn, Dartmouth College). The known
charac-teristics ofthese mutants may be summarized
asfollows. (i)The intact ts-23 virion has
reversi-ble temperature-sensitive hemagglutinating
ca-pability. However, this virion can adsorb to
cellsatnonpermissive temperatures (4, 18). (ii)
The surfaces of cells infected with ts-20 at
nonpermissive temperatures hemadsorb (4) and
bind ferritin-conjugated antibodyagainst intact virus (Brown, unpublished data), whereas
ts-23-infected cells neither hemadsorb (4) nor
bind antivirus ferritin-antibody conjugates
(Brown, unpublished data). (iii) Cells infected
with either mutant produce complete
nu-cleocapsids (4). (iv) Cells infected with either mutant contain uncleaved PE2 but not E2 at nonpermissive temperatures (7; K. Jones,
per-sonal communication; Brown and Smith,
un-published data), and in ts-20-infected cells
some of this protein can be located in the
plasma membrane (7). It appears that the phenotypes of the two mutants under nonper-missiveconditions are similar in most respects,
although one might conclude from the above data that themembranes ofts-20-infectedcells contain inserted viral glycoproteins, whereas ts-23-infected cells may not.
For electron microscopy, BHK-21 cells
(ob-tained from P. Faulkner, Queens University,
Canada) infected withapproximately50PFU of
1262
on November 10, 2019 by guest
http://jvi.asm.org/
%!s.4",
4 ~
OF4>~7~,
5 p b
'i
I
-'Ri~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~2
rP
',a3.
S$1Q
O.r4
.~~..'tk
S
~~
,~~~~*
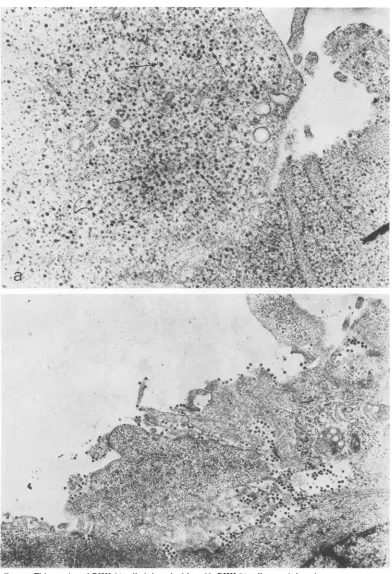
-FIG. 1. Thinsection of BHK-21 cells infected with ts-23. BHK-21 cells were infected at room temperature
with approximately 50 PFU of ts-23 per cell for 1 h. The inoculum was removed and replaced with Eagle
medium (5) which had been prewarmed to 40 C. At the end of the incubation period, cells were fixed in situ with
1.5%glutaraldehyde in the phosphate buffer of Millonig (9). The cells were scraped from the monolayer and,
after postfixation with osmium tetroxide, dehydrated and embedded in epon (8). After sectioning, the
preparations were stained with uranyl acetate and lead hydroxide. (a) Incubated for 8 h after infection at
39.5C.Nucleocapsids (arrows)areseenrandomlydistributed in the cell cytoplasm. Magnification x40,000. (b)
Infected and incubated as described in (a) and then shifted to 28 C and incubated for an additional hour.
Magnification x20,000.
1263
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.505.48.440.34.608.2]1264
NOTES*'.
v ,S ^
, 1% W
;,. 4rS..
* t 4 f
* ; ' a
J. VIROL.
8 .
'.t
41
%4 *4
::
)
0<vPwsX~~~
tsof:~
~~
ls
~~
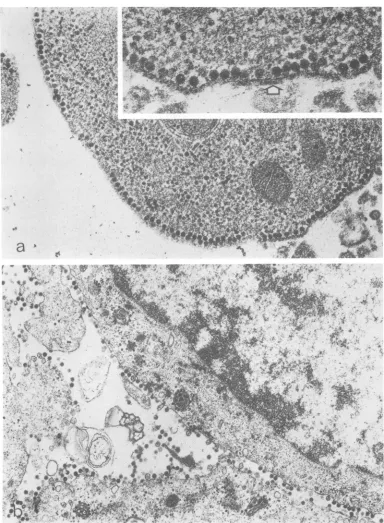
FIG. 2. Thinsection of BHK-21 cellsinfected with ts-20. Cells were infected, incubated, and prepared for
microscopyasdescribed for Fig. 1. (a)Incubation atnonpermissive temperature for 8 h. Nucleocapsids are seen
attached to the internal surface of the plasma membrane. The insert shows an area of this cell at higher
magnification. The bilayer nature of the modified membrane is evident, the nucleocapsids are intimately associated with its inner surface, and the outer surface appears thicker than normal in some areas (arrow).
Magnification x48,500 andx97,000 for insert. (b) Infected andincubated as described for (a) and then shifted
to 28C and incubatedforanadditional hour.Magnification x25,000.
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.505.76.460.67.590.2]ts-20 or ts-23 percellwereincubated at
nonper-missive temperature (39.5
C)
for 8 h andpre-pared for electron microscopy as previously described (2). Identical cultureswere shiftedto permissive temperatures (28C)for 1hafterthe initial incubation beforefixation.
Cells
infected with eithermutant atnonper-missive temperature had large and
approxi-mately equal numbers ofnucleocapsids at 8 h
afterinfection;however, the intracellular distri-bution of the
nucleocapsids
wasconsiderably
different. ts-23 nucleocapsids were uniformly distributed in the cell
cytoplasm (Fig.
la), whereas ts-20 nucleocapsids were attached totheinner surface oftheplasma membrane(Fig. 2a). Examination ofcells infected withts-20 at
earlier times (5 to 6h after infection) revealed
the presence of some nucleocapsids, but few
could be found in association with the
plasma
membrane (data not shown). The
observation
that cells infectedwithts-20readily hemadsorb
at this early time (4) suggests that membrane modification isquite extensivebeforethe
bind-ing of nucleocapsids. External to bound
nu-cleocapsids, the plasma membrane of ts-20-infected cells was often observed to be coated
with anamorphous materialapproximately12.0
nm thick (Fig. 2a, insert). This layer was not
found on uninfected cells or on cells infected with ts-23. Thesimilarity of the cross-sectional
thicknessofthis layertotheknownlength ofthe
spikes in the mature virion (1, 6, 17) suggests
that this layer contains the virus-specific
pro-tein(s) responsible for thehemadsorbing
capa-bility of the ts-20-infected cells (3, 18). The
tightassociation ofthe viral nucleocapsids with the modified plasma membrane in
ts-20-infected cells was also demonstrated by an
examination of membrane fragments obtained
fromgradientsofdisruptedinfected cells. Large
numbersof structuresresemblingnucleocapsids were found attached to these membrane
frag-FIG. 3. Negatively stained membrane fragment with associated nucleocapsids (arrows) obtained from
disruptedcellsinfected with ts-20 at nonpermissive temperatures. Cells were disrupted after 8 h of infection by
Douncehomogenization. An aliquot was layered onto a 10 to 45% (wt/wt)potassium tartrate gradient and
centrifuged at 30,000 rpm for 12 h in the SW-41 (Spinco) rotor. Membrane fragments of the type shown were
found at a density of 1.27gm/cm'. Thepreparation was stained with neutral 1.5%phosphotungstic acid.
Magnification x110,000.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.505.92.403.302.601.2]NOTES
ments (Fig. 3). No mature or budding virions
could be found in cells infected with either
mutant at nonpermissive temperatures. Both
intact and budding virions could be seen in
large numberswhen either mutant-infectedcell was returned to permissivetemperature(Fig.lb
and 2b). The processing oftheviral
nucleocap-sids produced at nonpermissive temperatures
into mature virion after the change to
permis-sive temperature demonstrates the reversible
nature of the mutant system with respect to virus production. Virus production after
tem-perature shift does not, however, mean that
membrane-associated viral proteins produced at nonpermissive temperature are actually chased into maturevirions.
It may be concluded from this work and
previous studies that ts-20-infected cells have
virus-specific proteins on the outer surface
which can hemadsorb and bind virus-specific
antiserum, and virus-specific alteration on the
inner surface of theplasmamembrane towhich
nucleocapsids attach. ts-23-infected cells have neither of these capabilities at nonpermissive
temperatures. Thus the
binding
of viralnu-cleocapsids to the inner surface of the
plasma
membrane, like the
hemadsorbing
capability, is avirus-specific function thatmaybe attributedto either or both PE2 and El present in the modified cell membrane (7). The inability to
attach
nucleocapsids
totheplasma
membrane and the absence ofhemabsorption
ints-23-infectedcellsmaybe duetothe lackofinsertion ofthe membrane proteinsincells infected with this mutant.
Alternatively,
if the viralmem-brane proteins areinserted inthe ts-23-infected cells, the absence of
hemadsorption
andex-posed viral antigenssuggeststhat the insertion is less complete than ints-20-infected cells. In
eithercase,the defectin ts-23preventsboth the
binding
of thenucleocapsids
and the properinsertion oftheproteins. Either thesetwo func-tions are controlled
by
one protein or, ifsepa-rately, by different proteins, one function is
dependent upon the prior
completion
of theother. Such a sequential relationship could be
invoked if the membrane proteins must be
installed in a particular arrangement before
nucleocapsidscouldbe attached. Suchseems to
bethecase, since ts-20-infected cells hemadsorb
before detectable quantities of membrane-associated nucleocapsids can be found (4; this
study). It is
possible, therefore,
that theinabil-ity of the virus membrane proteins to bind
nucleocapsids in the ts-23-infected cells is a
result ofthe lack of an appropriate
configura-tion oftheprotein within the membrane rather
than a simple defect in some capsid-binding region of the protein. Investigations are in
progress to determine the organization of the
virus proteins inthe mutant systemsdescribed
and to elucidate the nature ofthe capsid-mem-braneinteraction in ts-20-infected cells.
The technical assistance of Doris Renz is gratefully ac-knowledged.
This research wassupportedby the Deutsche Forschungs-gemeinschaft, SFB 74.
LITERATURE CITED
1. Acheson, N. J., and F. Tamm. 1967. Replication of semliki forest virus: an electron microscopic study. Virology32:128-143.
2. Brown, D. T., M. R. F. Waite, and E. R. Pfefferkorn. 1972.Morphology andmorphogenesis of Sindbis virus as seen with freeze-etching techniques. J. Virol. 10:524-536.
3. Burge, B. W., and E. R. Pfefferkorn. 1966. Isolation and characterization ofconditional-lethal mutants of Sind-bis virus.Virology 30:204-213.
4. Burge, B. W., and E. R. Pfefferkorn. 1968. Functional defects of temperature-sensitive mutants of Sindbis virus. J. Mol. Biol.35:193-205.
5. Eagle, H. 1955. Nutritional needs of mammalian cells in tissue culture. Science 122:501-504.
6. Harrison, S.C., A. David, J.Jumblatt,and F. E. Darnell. 1971.Lipid and protein organization in Sindbis virus. J. Mol. Biol. 60:523-528.
7. Jones, K. J., M. R. F. Waite, and H. R. Bose. 1974.
Cleavage of a viral envelope precursor during the morphogenesis of Sindbis virus. J. Virol. 13:809-817. 8. Luft, J. H. 1961.Improvements in epoxy resin embedding
methods. J.Biophys. Biochem. Cytol. 9:409. 9. Millonig, G. 1961.Advantagesof aphosphatebuffer for
0804solutions in fixation. J.Appl. Physiol.32:1637. 10. Pfefferkorn, E. R., and M. K. Boyle. 1972. Selective
inhibition of thesynthesis of Sindbis virion proteins by aninhibitionofchymotrypsin.J. Virol. 9:187-188. 11. Scheele, C. M., and E. R. Pfefferkorn. 1970. Virus
spe-cific-proteins,synthesizedincells infected with RNA+ temperature-sensitive mutants of Sindbis virus. J. Virol. 5:329-337.
12. Schlesinger, M. J., and S. Schlesinger. 1973. Large-molecular-weightprecusorsofSindbis virusproteins.J. Virol. 11:1013-1016.
13. Schlesinger,M.J., S.Schlesinger,and B. W.Burge.1972. Identificationof asecondglycoproteininSindbis virus. Virology47:539-541.
14. Schlesinger, S., and M. J. Schlesinger. 1972. Formation of
Sindbisvirusproteins:identification of a precursor for oneofthe envelope proteins. J. Virol. 10:925-932. 15. Sefton, B. M., and B. W. Burge. 1974. Biosynthesis of
Sindbis virus carbohydrates. J. Virol. 12:1366-1374. 16. Sefton, B. M., G. G. Wickus, and B. W. Burge. 1973.
Enzymatic iodination of Sindbis virus proteins. J. Virol. 11:730-735.
17. Simpson, R. W., and R. E. Hanser. 1968. Basic structure ofgroupAarbovirus strains middleburg, Sindbis and semliki forestexaminedby negative staining. Virology 34:358-361.
18. Yin, F. H. 1969. Temperature-sensitive behavior of the hemagglutinin in atemperature-sensitive mutant vi-rion ofSindbis. J.Virol. 4:547-584.
19. Yin, F. H., and R. Z. Lockart, Jr. 1968. Maturation defects in temperature-sensitive mutants ofSindbis virus. J.Virol. 2:728-737.
1266 J.VIROL.