0022-538X/81/110501-07$02.00/0
Epstein-Barr
Virus DNA
X.
Direct
Repeat Within the
Internal Direct
Repeat
of
Epstein-Barr
Virus
DNA
ANDREW CHEUNG ANDELLIOTT KIEFF*
SectionofInfectious Disease,Department ofMedicine,Microbiology, VirologyandImmunology,Kovler
Viral
Oncology
Laboratories,University
of
Chicago, Chicago,
Illinois 60637Received22April1981/Accepted1July1981
The 3,360-base-pair intemal direct repeat (IR) in Epstein-Barr virus DNA separatesthe shortand long unique DNAdomains. IR hasasingle BamHI site. The juncture between the short unique domain and IR has been mapped by
restriction endonucleases and is less than 2,600nucleotides before the BamHI site in IR. The junction between IR and the long unique domain has been sequenced and is approximately 650 nucleotides after the BamHI site in IR. Thus, relativetothestartof IRatthejuncture withthe short uniquedomain,the lastrepeat isatleast90basepairs short of beingcomplete.There is homology
between the 250-nucleotide fragments to the left and the right of the unique
BamHIsite in IR. A35-base-pairsequenceof the leftfragment is directly repeated within theright fragment,oncefullyandoncepartially. The implications of these findingsarediscussed.
Completerestrictionendonucleasemapshave
been derived for atleast seven isolates of Ep-stein-Barr virus (EBV)(2-4, 7-9,11,20)and for twoisolates of the relatedherpesviruspapio(12, 13, 16). The commonfeatures of this subgroup of herpesviruses include a variable number of directtandemrepeatsofa500-to600-base-pair
sequence (TR)atboth ends ofthelinearvirion
DNA molecule; a shortunique DNAsequence
of15 x 103basepairs (US),avariable number
of tandem direct repeats of a 3,360-base-pair
sequence (IR), andalong
largely
uniqueDNA sequence of 150 x 103 base pairs (UL). The tandem direct repeats of IR separate US and UL (3,4,7, 8, 10,22).Aside from its
organi4tional
role in EBV DNA, IR is animportantfunctionalelement in nonpermissively infected,growth-transformned
cells. Thenucleotidesequencecomplexity equiv-alent ofonestrand of IR is transcribed innon-permissivelyinfected,transformed cells(14,26).
Most of the RNA encoded by IR is processed and accumulates in polyribosomes (14, 26). In nonpermissively infected, growth-transformed cells IR encodes cytoplasmic polyadenylated,
andpresumably messenger, RNAs of3.0 x 103 and 1.5x 103bases (26).
Partial denaturation mapping and buoyant densitymeasurementsindicate that IRisricher inguanineplus cytosinethan mostof EBV DNA (5, 10).IR has asinglecleavage site forBamHI
andBglII restriction endonucleases (8, 10, 22).
Digestionof EBV DNA witheitherenzyme
gen-erates fragments which are the junction frag-mentsbetween IR and UL or US and a much moreabundantfragmentwhich is the fullrepeat unit(3, 10, 22; Fig. 1).Ithas been assumed that thejunctionfragmentstogether haveone
com-pleterepeatunit,althoughneither thebeginning
nor the end of the repeat has been mapped
relativetorestrictionendonuclease sites within
the repeat. The BamHI and BglII sites are asymmetrically placed relativetoeachother and divide IR into small and largecomponents (8).
Labeledsmallcomponent wasfoundtohybridize
weaklyto thelarge component and vice versa,
suggesting that there might be a direct or
in-vertedrepeatwithin IR (8).
The purpose of theexperiments described in this communication is(i)tolocate theregionof
homologywithinIR, (ii)tosequencethe
homol-ogousregionstodeterminetheextentof homol-ogyandwhether thehomologoussequences are onthesame oroppositestrands, and (iii)tomap
thebeginningand end of IR relative to
restric-tionendonucleasesites within IR.
MATERIALS AND METHODS
Viral DNAs. Plasmid pDK14, pDK10, and
pDK322, which contain the BamHI-C, -V, and -X
fragments of EBV (B95-8) DNA cloned into the
BamHI site of pBR322 were grown in X1776 underP1
EK2 conditions(3).BamHI-Visthefragmentformed
by cleavageofadjacent tandem repeats of IR at the
single BamHI site in IR. BamHI-C is the fragment
extending from the last BamHI site in US to the 501
on November 10, 2019 by guest
http://jvi.asm.org/
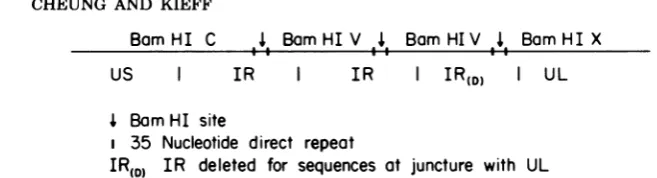
BamHI C ,4, BamHIV ,4, BamHIV 4 BamHIX
US I IR I IR IR(D) I UL
4 BamHI site
i 35 Nucleotide direct repeat
IR(D)
IR deleted for sequences at juncture with ULFIG. 1. Summary of organizational features and BamHI map ofEBVDNA around the internal reiteration.
Letters above the line indicate the BamHI fragments (3). BamHI-C is the rightward end of US and the
beginning of IR uptotheunique BamHI site in IR (3). BamHI- V is a full repeat unit from the unique BamHI
site in one repeat to the unique BamHI site in the second repeat(3).BamHI-X is the end of the last repeat of
IR and the beginning of UL(3).
BamHI site in the first repeat of IR. BamHI-X is the fragment extending from the BamHI site in the last
repeatof IRtothe firstBamHI site in UL (Fig. 1).
Separationand size determination of
restric-tionendonucleasefragments. Restriction
endonu-cleases were purchased from Bethesda Research
Lab-oratories (Bethesda, Md.) and used under conditions
specified by the manufacturer. Restriction digests of cloned EBV DNAs were electrophoresed at 40 V and
4°Cin 1% agarosegels in a buffer consisting of 40 mM
Tris-hydrochloride (pH 7.8),1mM EDTA, and 5 mM
potassium acetate. The size of fragments was deter-mined relative to intact and restriction endonuclease
fragments ofOX174andpBR322 DNAs (25).
Preparation for blot hybridization. After elec-trophoresis, the DNA fragments in the gel were
de-natured in situ with1MKOH, neutralized with 1 M
Tris-hydrochloride (pH7)-iMHCl,and rinsed with
6x SSC (lx SSC is 0.15 M NaCl-0.015 M sodium
citrate). The DNAwastransferred ontonitrocellulose
filters(MilliporeCorp.) as described bySouthern(24).
The filters were then rinsed with 2x SSC (pH 7),
blotted, dried, and baked for2hat80°C in a vacuum
oven.
Filter hybridization andautoradiography.All
stepsin filterhybridization werecarriedout at37°C.
Bakednitrocellulose filters were presoaked in
hybrid-ization buffer which consists of 50% formamide, 5x
SSC,0.5%sodiumdodecyl sulfate,20mM
Tris-hydro-chloride (pH 7.4), and 0.08% each of Ficoll, bovine
serumalbumin, andpolyvinylpyrollidonefor 30 to 60
min(6, 27). Thefilterswerethen allowed to hybridize
withalkaline-denatured, nick-translated (21)
radioac-tiveprobe in hybridization buffer for24h.After
hy-bridization thefilterswerewashed twice in
hybridi-zationbufferfor 1 h andthenin 2xSSC (pH 7) for 1
h. Thefilterswerethenair dried and exposed to
X-rayfilm.
DNA sequence analysis. DNA fragments were
treated with bacterial alkalinephosphatase (Bethesda
Research Laboratories), labeled attheir 5' end with
polynucleotidekinase (Bethesda Research
Laborato-ries) and [y-32P]dATP (Amersham Corp.), cut with
restriction enzyme,separatedonagarosegel, and
iden-tifiedby autoradiography. TheDNAfragmentswere
eluted andpurified byDEAEchromatography (18). In
otherexperiments, end-labeledDNAstrandswere
sep-arated onacrylamidegels. The chemical degradation
methodwas usedfornucleotide sequence
determina-tion (17). The sequenceswereanalyzedforregions of
homologyandreading frameson anAmdahlcomputer
by usingpreviouslyderivedprograms(15, 19).
RESULTS
Restriction endonuclease map of IR. BglII, PvuII,XhoI,SstI,orSstIIcutthe BamHI-V (IR) insert only once, whereasHinfl, HhaI, AluI, and MboII each cutBamHI-Vatseveral sites. The locationof thesingle-cutenzymesites wasdetermined from the size of the fragments
generated by cleavage of pDK 14 with these
enzymes individually and in combination with BamHI. The location of multicut restriction
en-donuclease siteswasalso determined inpartby
sequential cleavage and determination of the size offragments. The mapwas completed by
determining the size ofpartial digestion
prod-ucts. For these experiments BamHI-V was la-beledatthe5'ends. Thelabeled DNAwasthen
cutwith BglJI to separate the 760- and
2,600-base-pair fragments, each labeledonlyatthe 5'
BamHIend. The sizes of theproductsof
partial
digestion of the 760- and 2,600-base-pair
frag-mentswithasecondenzymewereusedto deter-mine the distancefrom the labeled BamHI end totheinternal restriction endonuclease sites for the secondenzyme (Fig.2). The restriction
en-donuclease mapsaresummarized in Fig. 3. Al-thoughonlypartofIRhas beensequenced,for conciseness fragments ofIR are subsequently
denotedbytheir mapcoordinates inbasepairs,
asshowninFig.3.
Map ofhomology within IR. The labeled
fragmentbetween nucleotides0 and760 (0-760
fragment) of IR (BamHI-V) hybridizes much lesstothe760-3,360fragment ofBamHI-V than tothe 0-760 fragment, indicating that there is
only partial homology between the two
frag-ments (Fig. 4a).Thepart of the0-760fragment
homologoustothe760-3,360fragmentis the 0-245component(Fig.4blane3,4dlane2,and4e
lane 2). The part of the 760-3,360 fragment
homologoustothe 0-760fragmentis the 3,120-3,360 component (Fig.4b lane 2,4c lane2,and 4flane2).Thus,theregionofhomologyis within 250nucleotidesoneachside of theBamHI site in IR.
Sequence of homologous
fragments
within IR. The 0-245 BamHI-PstI fragmentand3,120-3,360Hinfl-BamHIfragment(Fig.3)
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.497.74.403.61.150.2].2 22 2 2
[image:3.497.96.391.77.302.2]I.~~~~~~~~a
FIG. 2. Mapping ofmulticut restriction endonuclease sites by analysis of partial digestproducts (23).
BamHI-V(5'endlabeled)wasdigestedwithBgiI (a)togive760-base-pair (lane1)and2,6(X)base-pair (lane
2)fr-agments.Bothfr-agmentsweresubjectedtopartial digestion byenzymesindicatedatthetopofb,.c, d,
ande. The sizeofthepartial digestionproducts (giveninnucleotides in thefigure)wasdeterminedfr-omtheir
electrophoretic mobilityasdescribed inthetext.
Ban
HI-V
0 0 20 30 40 0 60 70 80 90 IC
I
I
II
qI
I I II~~~~~~~~~~~~~~~~~~~~~
I
0 10 20 30 40 50 60 70 80 90 IOC
0 840 1680 2520 3360
FIG. 3. Restrictionenzymemapofthe internalreiteration. Restriction endonucleasesitesofone- ortwo-cut
enzymes werederivedfromdouble digestion. The sitesof multiple-cut enzymeswere derivedfromparital
digestion of5' end-labeled DNAfragments. The scaleatthe bottom is inpercentageofoveralllength and in
nucleotidepairs (basedonsizeestimates).
were sequenced by the chemical degradation
method (17). The nucleotide sequences are
shown inFig.5asacontinuoussequence
begin-ningattheHinfl site and extending through the BamHI site to the PstI site. An identical
35-nucleotide sequence,
CCAGGCCAGCCGGA-GGGACCCCGGCAGCCCGGGCG, occurs as a
direct repeat202 to 168nucleotidesbefore and
91 to125nucleotides after theBamHI site. Also,
anidentical17-nucleotide subsequenceof the35,
GCCGGAGGGACCCCGGC, occurs as a direct
repeat212 to228 nucleotides after the BamHI
site.Theseresultsareindicatedonthesummary
map (Fig. 1). Several other shorter direct and
invertedrepeatsarealsopresent.
Junction between IR and USor UL.The
internal reiteration begins at the right end of
BamHI-C (3, 7, 8, 10). EcoRI, whichmakestwo cutsatthe left end ofBamHI-C, separatesUS components ofBamHI-C from theright end of
BamHI-C which contains the beginning of IR
(Fig 6A). The distance fromthe beginningof the
firstcopyof IRtothe BamHI sitein IR (which separates BamHI-C from BamHI-V [Fig. 1])
wasdetermined bycleavingBamHI-C with
re-strictionendonucleaseswhichcutIR beforethe Hinf I
HhoI Alu I Mbol SstI
XhoI
Pst I PvuI Bgl E Sst I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.497.104.395.363.489.2]BamHl site. Enzymeswhich cutBamHI-C be-tween thebeginningofIRand thefirstBamHI site should yield the same size fragment from BamHI-C as from BamHI-V. PvuII and SstI, whichcut IR at 840 and 2,500nucleotides before theBamHI site, generate IRhomologous frag-mentsof840and2,500nucleotidesfrom BamHI-C (Fig. 6A). BglII, which cuts IR at 2,600 nu-cleotides before the BamHI site, does not cut BamHI-C (recombinant orvirion DNA). Thus, the beginning of IR in BamHI-C is less than 2,600 andprobably more than 2,500nucleotides beforetheBamHl site in IR.
The internal reiteration ends in BamHI-X (Fig. 1). HindIIImakes a single cut in
BamHl-X and separates UL components ofBamHI-X from theleftend ofBamHI-X, whichcontains the end ofIR (Fig. 6B).PstI, whichcuts IRat 245nucleotides aftertheBamHIsite, andXhoI, which cuts IR at 550 nucleotides after the BamHI site, both generate homologous
frag-mentsofidentical size from BamHI-X (Fig.6B).
However, BamHI-X (recombinant or virion
DNA) lacks the BglII site at 760 nucleotides. Labeled IR hybridizes almost halfas much to
thepart ofBamHI-X tothe rightofthe XhoI site at 550 nucleotides as it does to the 0-550
fragmentwhichisincommon toBamHI-V and
-x(Fig. 6B). Thesedatasuggestthat IR extends for 100 to 200nucleotides past the XhoI site into BamHI-Xandstopsbefore theBglII siteat760 nucleotidesinIR.
Sequenceatthe end ofthelast repeat of
IR.Todefinemoreprecisely the end of the last repeat of IR in BamHI-X, the nucleotide se-quences oftheregionstotheright oftheXhol
cutsites inBamHI-Vand-Xweresequencedby
chemicaldegradation. The nucleotide sequences ofBamHI-V and-X(Fig. 7)areidentical for117 nucleotides pasttheXhoIsite. Thereafter, the nucleotide sequences diverge. Thus, US begins
118nucleotides after theXhoIsiteinBamHI-X.
_.
_:-'.-_IW.
o _
FIG. 4. Mappingofthehomologousregionsin IR. The coordinatesofthe DNAfragmentsareindicated in
basepairs between BamHI sites in the orientation shown inFig.1and 3. The coordinatesarethose shown in
Fig. 3.BlotsofBamHI-VdigestedwithBglII (a) orPstI,XhoI,BglII,and PvuII(b) werehybridizedwith
labeled BamHI-V(lane 1), with labeled (0-760)probe (lane 2), orwith labeled(760-3,360)probe(lane 3).
Homologousregions werefurtherdefined byhybridization oflabeled(0-760) probeto a blotofanMboII
digestofthepartofBamHI-Vfromcoordinates2,520to3,360(c), by hybridization oflabeled (2,520-3,360)
probe(d)oroflabeled(0-245)probe (e)toblotsofPstI-andPvuII-digestedBamHI-V, andbybybridization
of labeled(0-245)probetoblotsofaHinfl digest ofthe2,520-3,360fragment(f).
X - i.. .0:
;'..,. -- I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.497.110.401.311.592.2]30 60 90 GAG TCC AGA GGT CAG GGG CAC CTC AGG GTG CCC CCG GGT CC C GCC CGGGCG GCC CCA GAG GCC GGT GLU SER ARG GLYGLN GLY HIS LEU ARG VAL PRO PRO GLY PRO ARG PRO ALA GLY GLY THR PRO ALA ALA ARGALA ALAPRO GLU ALA GLY
120 150 10
TCC TCG CCC CTTCCC CGG GCT TCA GAG CCCAGG ATG TCCCCC AGA AGG GAC CCTAGG CGTCCC CTC TCC TCCCCT CCA GGC CCGAGC CTC SER SER PRO LEU PRO ARG ALASER GLU PROARG MET SER PRO ARG ARG ASP PRO ARGARGPRO LEU SER SER PRO PRO GLY PRO SER LEU
210 240 , 270
TCC CTCGCGGAG AGG CCT CTT TGG GCC CTC AAGTCC AGC CCC ACC GAG ACC CGA GTG GCCCGG ATC CCC CCACCG GCC CTTCTC TCTGTC SER LEU ALAGLU ARG PRO LEU TRP ALALEU LYS SER SER PRO THRGLU THRARG VAL ALA ARG ILE PRO PRO PRO ALA LEU LEU SER VAL
300 330 360
CCC CTGCTC CTC TCC AAC CTT CGC TCC ACC CTA GAC CCC AGC TTC TGG CCT CCC CGG GTC CA CAg4
PRO LEU LEU LEU SER ASN LEU ARG SER THRLEU ASP PRO SER PHE TrP PRO PRO ARG VAL HIS GLN ALA SER ARG ARG ASP PRO GLY SER _390 420 . 450
Q&GGCQAGTCG CCT TCC CTC TCC C(A)TGGC CTC TCC TTC CCG CCTCCC ACC CGA GCC CCCTCA GCT TGC CTCCCC AOCGGG TCC ATC AGG
PRO GLY GLU SER PRO SER LEU SER HIS GLY LEUSER PHE PRO PRO PRO THR ARG ALA PRO SER ALA CYS LEU PRO THRGLY SER ILE ARG
460
[image:5.497.62.452.63.217.2]CCG GCC GGA GGG ACC CCG -G GCC CGG TGT CAG TCC CCC CTG CAG PRO ALAGLY GLY THR PRO ALA ALA ARGCYS GLN SER PRO LEU GLN
FIG. 5. Nucleotide sequenceofthehomologousDNAs around the BamHI site inIR.The 242nucleotidesin
theHinfl-BamHIfragmenttotheleft ofthe BamHI siteweresequencedbydeterminingthe 200nucleotides
fromeach end. The253-nucleotideBamHI-PstIfragmenttotheright ofthe BamHI sitewasalsosequenced
bydeterminingthe200nucleotidesfromeachendand 120to150nuckotidesfromtheHinfIsiteat367to371
nucleotides in thefigure.The continuousnuckotide sequence is shownfromtheHinfIsitethroughthe BamHI
site(indicatedby an arrow between nucleotides242and243)tothe PstI site.Regionsofidentityareindicated
by boxes;regions ofsimilarityareindicatedbyunderlining,with asterisksindicatingunmatched bases. The
amino acidsequence is shown in theonly reading framewhich doesnothaveatermination codon.
DISCUSSION
The data which are summarized in Fig. 1 confirm that there ishomologywithin the inter-nal reiteration in EBV DNA and indicate that thehomology is duetothedirectrepeatofa 35-nucleotidesequencetotheleft and therightof
theuniqueBamHI site in IR. A directrepeatof
the 17-nucleotide subsequenceof the 35 begins
87 nucleotides after the second 35-nucleotide sequence. This 17-nucleotide subsequence is partofalargersequenceduplication,since there is extensive homology to the 35-nucleotide
se-quence onbothsides of the17-nucleotiderepeat.
Thus,therearethree directrepeatsofahighly
conserved sequencein thisregionof IR. These repeatsarelikelytohaveimportantcisor trans
fimction.
Thefragments which have beensequencedon
bothsides of the BamHI site in IRare242and 253 nucleotides (Fig. 5). These sequences are
continuous in IR andare495 ofthe total3,360 nucleotides of IR.Through the continuous 495-nucleotide sequence, there is no polypeptide
chain termination codon in the reading frame beginning GAG. There isonetermination codon (TAG) in the secondreading frameand one in the third. Translation from theoppositestrand isblocked byatleasttwoterminator codonsin each reading frame. The amino acid sequence whichcould make maximumuse ofthe coding capacity of this sequence by translating it in framefrom thefirst nucleotide is shown below thenucleotidesequenceinFig.5.
The BamHI sitein IR is lessthan2,600 nu-cleotides afterthebeginningof IR inBamHI-C. The nucleotide sequence which begins IR has
not been determined. Relative to the start, which isless than2,600 nucleotides before the BamHI site, the last repeat of IR would be expectedtoterminatemorethan760nucleotides after the last BamHI siteso astoterminate in acompleterepeat.However, the lastrepeat ter-minates 117nucleotidesafter the XhoI site and before the Bglll site at 760. The XhoI site is
approximately550nucleotides after the BamHI
site. Thus, the last repeat stopsapproximately 670nucleotides after the last BamHIsiteorat
least90nucleotidesshortof completingthelast
copyof IR(relativetothestartofIRatthe
US-IRjunction).
Presumably, the repetitions of IR are struc-turallyorfunctionally usefultoEBV. The num-ber ofrepeatsofIR varieswithin populations of molecules produced bya singleisolate of EBV (8). Theaverage number ofrepeatsalso varies among EBV isolates (11). The B95-8 isolate whichhasadeletion of approximately 15 x 103
nucleotides in the UL tendstohave10 repeats of IR, whereas other isolates of EBV and of
herpesviruspapio tendtohavesix repeats of IR
(11-13). This suggests that the number of re-peats ispartially dependentonoverall DNA size. Increasesinoverallsizehave also been reported with passage and may be due to an average increase in thenumber of repeats of IR (1). One consequence of the finding in this study of a deletioninthe last(orfirst)repeatof IR is that it raises the possibility that multiple repeats could be generated from a parental molecule which consists oflessthan two complete repeats.
Althoughdeletionsasa result ofhomologous
recombination between the repeats within IR
on November 10, 2019 by guest
http://jvi.asm.org/
|~~~~~~~~
*hid-FIG. 6. MappingofthejunctureofIR andUS(BamHI-C;panel A)orUL(BamHI-X,panel B). BamHI-C (panel A) orBamHI-X(panel B)wasdigestedwithrestriction endonucleases whichcutwithin IRorwithin the adjacent USor UL to determine the extent ofIR in BamHI-C and BamHI-X. Blots ofrestriction
endonucleasefragmentsofBamHI-C,-X,or-VwerehybridizedtolabeledBamHI-Vto indicate thefragments
which contain sequences ofIR. The recombinant DNAs were digested with EcoRIor HindIIIor with a
combinationofBamHI andSst,PvuII, XhoI,orPstL.The numbersindicate the coordinatesofthefragment
within IR in basepairsrelativetothemapshown inFig.3orthe sizeofthefragmentinbasepairs. Fragments
consisting ofbothpBR322and EBVDNAareindicatedby"P" and theletterofthe EBV DNAfragment.
A. Bam HIV
10 20 30 40 50 60 70 80 90 TCGAGTAGGT GCCTCCAGAG CCCCTTITTGC CCCC(C)TGGCG GCCCAGCCCG ACCCCCGGGC GCCCCCAAAC TTTGTCCAGA TGTCTAGGGG
100 110 120 130 140 150 160 17O
TCCCCGAGGG TGAGGCCCAG CCCCCTCCCG CCCCTGTCCA CTGCCCCGGT CCCCCCAGAA GCCCCCAAAA GTAGAGGCTC AGGCC
B BamHI X
10 20 30 40 50 60 70 80 90 TCGAGTAGGT GCCTCCAGAG CCCCT TT TGC CCCC(C)TGGCG GCCCAGCCCG ACCCCCGGGC GCCCCCAAAC TTTG TCCAGA TGTC TAGGGG
'00 110 120 130 140 ISO
TCCCCGAGGG TGAGGCCCAG CCCCCTCTCG CCCAAGCTGC TTTGATTCTT GGGATATTTT T
FIG. 7. Nucleotidesequencesofthe DNAstotheright ofthe XhoIsites in BamHI-V and -X. TheDNAs
were5'end labeledatthe XhoIsite, and thenucleotidesequencesweredeterminedbychemicaldegradation (17).
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 40, 1981
might be expected, the deletion ofpart of IR outside of these regionsand at the juncture of IR withUL orUS isasurprising finding which could be of significance. IR and the adjacent partof ULencode atleastone andpossibly two abundant cytoplasmic polyadenylated RNA(s) in nonpermissively infected, growth-trans-formed cells (14, 26). Translationof an mRNA from IR would be blocked once in all three reading frames between theXhoI site and the switch into UL (Fig. 7). In the reading frame beginning CGA (Fig. 7) a second terminator sequenceTAG occurs 162nucleotides after the XhoI site in IR (Fig. 7). This terminator is de-leted by the switchinto UL at 118nucleotides after the XhoIsite in the lastcopyofIR at the IR-UL juncture in BamHI-X (Fig. 7). If the terminator codonTAG, which is five nucleotides totheright of the XhoI site,wereeliminatedby RNAprocessing, aconsequence of the deletion of the lastpartof IR in BamHI-X could be that amessageextending from IR into UL would be translated by using codons from both IR and UL.
ACKNOWLEDGMENTS
Thecontribution of recombinant EBV DNAs by T. Dam-baugh and of computer assistance by Sophia Kholodenko from Bernard Roizman's laboratory are gratefully acknowl-edged.
This research was supported by Public Health Service grants CA 19264 and CA 17281 from the National Cancer Institute andby grant MV 32F from the American Cancer Society.E.K. isaFaculty Research Awardee of the American CancerSociety.
ADDENDUM IN PROOF
TheSstIsite inBamHI-C (Fig. 6) is within the US
component. Thus, the extent of the deletion of
se-quences intheincomplete copy of IR, relative to the
beginning ofIR, is larger than anticipated from the
datareportedhere.
LITERATURE CITED
1.Adams, A.,G.Bjurse1l,C.Kaschka-Dierich,and T. Lindahl.1977.CircularEpstein-Barrvirusgenomesof reduced size inahumanlymphoidcellline ofinfectious mononucleosisorigin.J.Virol. 22:373-380.
2. Bornkamm, C. G.,H.Delius,U.Zimber,J. Hudew-entz,and M. A.Epstein.1980.Comparisonof Epstein-Barr virus strainsof differentorigin byanalysisof the viral DNAs. J.Virol. 35:603-618.
3. Dambaugh, T.,C.Beisel,M. Hummel, W.King, S. Fennewald,A.Cheung, M.Heller,N.Raab-Traub, andE.Kieff.1980.Epstein-Barrvirus DNA. VII. Mo-lecularcloningand detailed mappingof EBV(B95-8) DNA. Proc.Natl. Acad.Sci.U.S.A.77:2999-3003. 4. Dambaugh,T.,N.Raab-Traub,M.Heller,C.Beisel,
M.Hummel,A.Cheung,S.Fennewald,W.King, and E. Kieff.1980.Variations among isolates of Ep-stein-Barr virus.Ann.N.Y.Acad. Sci.354:309-325. 5. Delius,H.,andG. W. Bornkamm.1978.Heterogeneity
ofEpstein-Barrvirus.II.Comparisonofatransforming andanontransformiing virusby partial denaturation mapping of their DNAs. J. Virol.27:81-89.
6. Denhardt,D. T.1966.Amembrane-filtertechniquefor the detection ofcomplementaryDNA.Biochem.
Bio-507
phys.Res.Comm. 23:641-646.
7. Given, D., and E. Kieff. 1978. DNA ofEpstein-Barr
virus.IV.Linkage mapfor restriction enzymefragments of the B95-8and W91 strains of EBV. J. Virol. 28:524-542.
8. Given, D., and E. Kieff. 1979. DNA ofEpstein-Barr virus. VI.Mapping of the internal tandem reiteration. J.Virol. 31:315-324.
9. Given,D.,D.Yee,K.Griem,and E.Kieff.1979.DNA ofEpstein-Barrvirus.V. Direct repeatsatthe endsof EBVDNA. J. Virol.30:852-862.
10.Hayward,S., L. Nogee,and G.Hayward.1980. Orga-nization ofrepeated regionswithin theEpstein-Barr virus DNAmolecule. J. Virol. 33:507-521.
11.Heller,M.,T.Dambaugh,and E. Kieff. 1981. Epstein-Barr virus DNA. IX.Variations among viral DNAs. J. Virol.38:632-48.
12. Heller, M., P.Gerber,and E.Kieff.1981.Herpesvirus papio DNA issimilar inorganizationtoEpstein-Barr virusDNA. J.Virol. 37:698-709.
13. Heller,M., and E. Kieff. 1981.Colinearityof Epstein-Barr virus DNAand the DNA ofherpesviruspapio.J. Virol. 37:821-826.
14.King, W., A. Thomas-Powell, N. Raab-Traub, M. Hawke, andE.Kieff.1980.Epstein-Barrvirus RNA. VI.Viral RNA inarestringentlyinfected, growth-trans-formedcellline.J. Virol.36:506-518.
15.Korn,L, C. Queen,and M.Wegman.1977.Computer analysis of nucleic acid regulatory sequences. Proc. Natl.Acad. Sci. U.S.A. 74:4401-4405.
16. Lee, Y.,A.Tanaka,R.Law,M.Nonoyama,and H. Rabin.1981.Linkagemapof thefragmentsofherpes viruspapioDNA.J. Virol. 37:710-720.
17. Masam,A.M.,and W.Gilbert.1977.Anewmethod for
sequencingDNA. Proc.Natl. Acad. Sci. U.S.A. 74:560-564.
18. Muller, W.,H.Weber,F.Meyer,andC. Weissmann. 1978.Site-directedmutagenesisinDNA:generationof point mutation in clonedcomplementaryDNAatthe positionscorrespondingtoamino acids 121to 123.J. Mol. Biol. 124:343-358.
19. Queen, C., and L Korn. 1980. Computer analysis of nucleic acidandproteins methods.MethodsEnzymol. 65:595-609.
20. Raab-Traub, N., T. Dambaugh, andE. Kieff. 1980. DNAofEpstein-Barrvirus.VIII.B95-8,theprevious prototype,isanunusual deletion derivative. Cell22: 257-267.
21. Rigby, P., M. Dieckmann,C.Rhodes, and P. Berg. 1977.Labeling DNA to high specific activity in vitro by nicktranslation with DNApolymerase I. J. Mol. Biol. 113:237-252.
22. Rymo,L., andS. Forsblum. 1978. Cleavage of Epstein-Barr virus DNA by restriction endonuclease EcoRI, HindIIIandBam I.Nucleic Acids Res. 5:1387-1402. 23. Smith, H. O., and M.LBirnsteil.1976.Simple method
for DNA restriction sitemapping. Nucleic Acids Res. 3:2387-2391.
24. Southern, E. M. 1975. Detection of specific sequences amongDNAfragmentsseparated by gel electrophore-sis. J. Mol.Biol.98:503-517.
25. Sutcliffe, J.G. 1978. pBR322 restriction map derived
from the DNA sequence: accurate DNA size markers upto4361nucleotidepairs long. Nucleic Acids Res. 5: 2721-2728.
26. vanSanten, V., A. Cheung, and E. Kieff. 1981. Epstein-BarrVirus (EBV) RNA.VII.ViralcytoplasmicRNA in arestringently infected celllinetransformed in vitro by EBV.Proc.Natl. Acad. Sci. U.S.A.78:1930-1934.
27. Wahl, G. M., M. Stern, and G. R. Stark. 1979. Efficient transfer oflarge DNA fragments from agarose gels to
diazobenzylozymethylpaperand rapidhybridization by using dextran sulfate. Proc. Natl. Acad. Sci. U.S.A. 76: 3683-3687.
on November 10, 2019 by guest
http://jvi.asm.org/