JOURNALOFVIROLOGY, Dec.1977,p.826-835
Copyright©1977 American Society forMicrobiology Printed in U.S.A.
Transcriptase
Activity
Associated with Rabies
Virion
AKIHIKO KAWAI
Institute for VirusResearch, Kyoto University, Kyoto 606, Japan
Received forpublication8June1977
Rabies
virion-associated transcriptase activity wasinvestigated
in vitroand
compared
with that of the New Jersey serotype ofvesicular
stomatitis
virus.The concentration of detergent that affected
[3H]GMP
incorporation
intoacid-insoluble material
wassignificantly
different for both viruses.Vesicularstomatitis
virus New Jersey
required
0.05 to0.1% nonionic detergent, whereas rabiesvisioncould
notbe
fully
activated unless 4 to 5% detergent was used. Otheroptimal
conditions
were asfollows: 40 mMNaCl,
5 mMMg2",
40mMTris-hydrochloride
(pH 7.4),
5mM dithiothreitol,
and30'C.
Thereactionrequired
fournucleoside
triphosphates.
The initial rate ofRNAsynthesis
by rabies vision enzyme was 140pmol of GMP incorporated/mg
of viralprotein
per h andlinearlyincreased
until
about 8h,
withaslight
initiallag phase. The enzyme activity thatcorrelated
with
the content of Lprotein
was highest when rabies virions were grown at330C. The product
wassingle-stranded
RNA, which wascomplementary
in basesequences
torabies viral RNA. Most of the RNAsynthesized sedimented
at6-16S.Rabies virus is a
representative
animal virusbelonging
to the rhabdovirus group(11).
Thesingle- and
negative-stranded genome
RNA with amolecular weight
of 4.4 x106
to 4.6 x106,
synthesized
inthe
cytoplasm,
forms ahelical
structure
of
nucleocapsid
in association with aprotein and
iscovered
withthe
envelope by
theprocess of
budding
atthe
cellularmembrane
(19,
21).
Ithas been
proposed
that
the
synthesis
of mRNA
atthe
early
stepof
infection(primary
transcription) of
negative-strand
RNAviruses,
such
asrhabdo-, orthomyxo-,
andparamyxovi-ruses, is
carried
outby
thevirion-associated
transcriptase (2).
This concept hasbeen
con-firmed
for many viruseshaving
anegative-stranded genome
RNA(5).
Attempts to demonstrate this
enzyme
activity
in
the rabies
virion in vitrohave
apparently
been
unsuccessful
(5, 22,
24).
Bishop
reported
that the
highest activity observed
incorporated
only
5 to 10pmol
of
UMP into
theproduct
perhper mg
of
protein (Abstr.
Symp. Adv. Rabies
Res.,Atlanta, p. 9, 1976). Thislowactivity,
cal-culated
tobe about 1/2,000 that of the Indianaserotype of
vesicular stomatitis
virus(VSV)
(6),seemed
tobeparadoxical,
because apolypeptide
molecular
weight
thatcorresponds
tothatof an Lpolypeptide
of VSVrequired
fortranscription
has been observed in rabies virus
preparations
analyzed by
sodiumdodecyl
sulfate(SDS)-poly-acrylamide gel electrophoresis (PAGE).
Thepresent paper
is adescription
of theoptimal
conditions
required
to demonstrate rabiesvir-ion-associated transcriptase activity
invitro.MATERIALS AND METHODS
Viruses and cells. TheHEP-Flury strain of rabies virus,obtained fromA. Kondo (NationalInstitute of
Health, Tokyo, Japan), was passaged more than40
timesthroughBHK-21 cells and cloned five timesby the isolation ofa large plaque formed on BHK cell plates containing lessthan twoplaques per plate (10), and clone 2026/20was used throughout this experi-ment.Anegg-adaptedstrain of VSV New Jersey was obtained from the National Institute of Animal Health (Tokyo, Japan) and cloned twice in our laboratory, and clone NJ3013wasused inthisexperiment. Both virusesweregrowninBHK-21 cells. Toprepare the rabies virusinoculum free of defective interfering par-ticles,50 to 100wholeplaqueswere isolated, pooled in10ml,sonically treated for15 switha40-W Sonifier, and thenusedastheinoculum. The suspensions usu-ally contained about
105
PFU/ml.Both viruses werepurifiedfrominfected culture fluidsbyaslight
modi-fication ofaproceduredescribed elsewhere (10). After culturefluidswereclarifiedbycentrifugationat4,000 rpm for30min at4°C,viruseswereprecipitated by
the polyethylene glycol method (14) and suspended
in 24 ml of NTE buffer. This virus suspension was layered on 6 ml of a 20% (wt/vol) sucrose cushion andspunat64,000xg for120 minat4°C.Thepellet wasresuspendedin 2ml of NTEbuffer and sonically treated for2 minin ice bya40-WSonifier (20 kc). The virussuspensionwaslayeredon28ml ofa 10 to
45% (wt/vol) linear sucrosegradient inNTE buffer
andthencentrifugedat33,000 x gfor 90 min at4°C.
Thegradientwasfractionated into30tubes and mon-itored for optical density at 260 nm. Two or three tubes, which corresponded to the band ofinfective bullet-shaped (B) particles, werepooled and concen-trated bycentrifugationat64,000x g for 120minat 4°C.Thepellet, resuspendedinNT bufferatanoptical
826
on November 10, 2019 by guest
http://jvi.asm.org/
RABIES VIRION 827
density of 2.0 at 260 nm, was estimated to contain 250 to 255
jtg
of protein per ml and was stored at -70'Cuntil use (13). Under these conditions of virus purification, B and defective interfering particles form individual bands at a distance in the sucrose gradient, andboth bands could be separately isolated. Electron microscopic studies revealed that more than 90% of the particles in B-particle preparations were bullet shaped (10).Infectivity assay. Infective rabies virus was ti-trated byplaque formation in suspended BHK-21 cells (20).VSV New Jersey was titrated by plaque forma-tion onmonolayers of BHK cells.
Estimation of protein content. Concentration of protein inpurifiedvirus stocks was determined by the method ofLowry et al. (13), using bovine serum al-bumin as thestandard.
PAGE. Slabgel electrophoresis with SDS was per-formed by using Laemmli's discontinuous buffer sys-tem (12). Separating gels were 10% acrylamide con-taining 0.267% N,N'-methylene-bisacrylamide and 0.12% SDS, and stacking gels were 4.5% acrylamide containing 0.12% NN'-methylene-bisacrylamide and 0.1%SDS. Purified virussuspensionwasdissolved in the same volume ofsample buffer, whichwas com-posed of0.125 M Tris-hydrochloride (pH 6.8), 4.6% SDS, 20% glycerol, 10% 2-mercaptoethanol, and 0.005% bromophenol blue, andwasheatedat 1000C
for2minbefore electrophoresis. Electrophoresis was carriedout at a constantcurrentof4to 5 mAuntil thedye front had reached the bottom of the gel; then gelswerestained with 0.25% Coomassie brilliant blue R-250 in50% methanol and 7% acetic acid for2h at
370C
and destained in 50% methanol and 7% acetic acid. Thegelswere then either dried under vacuum onto Whatman 3MM filter paper or scanned by a microdensitometer(model MKIII, Joyce,Loebl& Co. Ltd.,Gateshead,England)beforedrying.RNA polymerase assay. The standard 0.25-ml
reaction mixture for rabies virus contained, unless otherwisementioned, 10
.tmol
ofNaCl, 1.25tmolofMg2e,
10,molofTris-hydrochloride(pH7.4at300C),
0.25jumol
ofATP,CTP,andUTP,0.05,umol of GTPand 10,Ciof[8-3H]GTP (10.5Ci/mmol), 1.25Mmol
ofdithiothreitol (DTT), 12.5mgof Triton X-100 or Nonidet P-40(NP-40), and variousamountsof rabies virionsasindicated in thetext.The standard reaction mixture forVSV-associatedtranscriptase followed the formula of Naito and Ishihama (16). Reaction mix-tures wereincubated at 30°Cfor varyingperiodsas described in thetext.
Radioactivity incorporated into the acid-insoluble
fraction was determined by a modification of the method of Baltimore et al. (3). Reaction mixtures werecooled inanicebox and made upto5%(wt/vol)
with trichloroacetic acid and 100 mM with sodium
pyrophosphate,afterwhich 200,ugof yeast RNAwas
added. After 30 min at 4°C, samples were filtered throughamembranefilter(typeHA;MilliporeCorp.),
andeach filterwaswashedin5%trichloroacetic acid forat least6h (the5% trichloroacetic acid solution waschanged every hour) andfinally in99%ethanol for 30min, after whichtheyweredried.Sampleswere
thenassayedforradioactivityinaliquidscintillation
spectrometer(Nuclear-ChicagoMarkII)at15to20%
efficiency, using a toluene scintillator. Background counts weredetermined, unless otherwise mentioned, for the zero-time samples and usually ranged from 50 to70cpm.
Extraction of RNA. The extraction of rabies virion RNAwascarried out by the SDS-hot-phenol method (18). Fifteenmilliliters of purified virus sample, con-tainingabout 3.6 mg ofprotein in NTE buffer, was mixed with anequal volume of liquified hot-phenol solution (preheatedat65°C) immediately after addi-tionof SDSto afinal concentration of 0.5% and was vigorously shaken for 3 min at 65°C. The solution wasthen chilled, and the aqueous phase wasobtained after high-speed centrifugation. The hot-phenol ex-traction wasrepeatedonce. RNA was obtained from the aqueousphasebytwocycles of ethanol precipita-tionandfinally dissolved in2xSSC (0.3 M NaCl plus 0.03M sodium citrate) to afinal concentration of 15
tg/ml.
Annealing experiment of virion polymerase product. 3H-labeled products were prepared as fol-lows. After incubation for 15 h at 30°C, 0.5 ml of reaction mixturewasmade 0.2% with SDS and0.1M
with NaCl, and 200,g ofpurified yeast RNA was
addedas acarrier; the fraction containing RNAwas thenextractedby cold ethanolprecipitation. The pre-cipitatewasdissolved in0.3mlof2x SSC containing
5
jg
ofpolyvinylsulfate per ml.Annealingwasdoneasfollows. A 0.11-ml amount of3H-labeled product, 0.25ml of virion RNA (15jg/mlin2xSSC),and0.04
ml of 1% SDS in 2x SSC were mixed, boiled for 8 min, and then incubated for2 h at 70°C. The first
sampleswereneitherboilednorannealed,the second
samples were boiled but quickly chilled, the third
samples were annealed without addition of excess
virionRNA, and the fourthsamplesweredone in the
completesystem,thatis,annealed withexcessrabies
RNA(Table4). After beingchilled,thesampleswere
divided into two equal portions, one of which was treated with 25U of RNasesAandT,per ml for30
minat 25°C. Sampleswerethenchilled and
precipi-tated with coldtrichloroaceticacid,andacid-insoluble radioactivitywasdeterminedasdescribed above.
Velocity sedimentation analysis of product
RNA.3H-labeledproduct wasextracted from0.75ml
of reaction mixture, after the addition of 123,ug of rRNA from BHK cellsas a carrier, by cold ethanol
precipitation and were dissolved in 0.5 ml of
SDS-NAE buffer. The solutionwasboiled for4min,layered
on4.5ml ofa5 to20% (wt/vol) sucrose gradientin SDS-NAEbuffer,andcentrifugedat130,000 x g for4
hat 22°C, usinganRPS-40rotor (Hitachi Co.Ltd.,
Tokyo). Extractions of the gradientwere monitored
foroptical densityat260nm,and then each fraction
wasdivided intotwoequal portions,oneof whichwas treated with RNase asdescribed above at 30°C.All
samples were then precipitated with cold
trichloro-acetic acid, and acid-insoluble radioactivity was
as-sayed.
Chemicals and buffers.
Phosphate-buffered
sa-line contained: NaCl, 0.137 M; KCl, 0.0027 M;NaH2PO4, 0.008 M; KH2PO4, 0.0015M; CaCl2, 0.009 M;
MgCl2,
0.005 M;pH 7.4at 25°C. NT buffercon-tained:NaCl,0.13M;
Tris-hydrochloride,
0.05M;
pH
7.4 at 250C. NTE buffer contained: NT bufferplus
VOL. 24,1977on November 10, 2019 by guest
http://jvi.asm.org/
828 KAWAI
0.001 M EDTA. NAE buffer contained:NaCl,0.05M; sodiumacetate,0.01M; EDTA,0.001M;pH5.1. SDS-NAEbuffer contained: NAE bufferplus0.2% SDS.
[8-3H]GTP (10to11 Ci/mmol)and[8-3H]ATP (29 Ci/mmol)wereproductsof theRadiochemicalCentre, Ltd., Amersham, England. ATP, CTP, GTP, UTP, rifampin, a-amanitin,and DNaseI (200 U/mg) were
purchased from Boehringer-Mannheim-Yamanouchi Co. Ltd., Tokyo, Japan. RNase T. (5 x 105 U/mg)
wasfromSankyo Co. Ltd., Tokyo, Japan. RNase A
(3,233 U/mg) was from Worthington Biochemicals
Corp.,Freehold,N.J. Triton X-100wasfromNakarai Chemical Co.,Kyoto, Japan. NP-40wasfrom BDH
Chemicals Ltd., Poole, England. These commercial productswereallused without furtherpurification.
RESULTS
Incorporation
ofribonucleoside
mono-phosphate by rabies virion. The activity ofthe rabies virion forincorporation of
[3H]GMP
was measured by the same procedure as that
appliedtoVSV NewJersey (16). Nosignificant
activity could be observed (Table 1, experiment
A). Todetermine whether such negative results
under 1/1,000 of VSV New Jerseymight be due
simplytoinappropriate conditions for the rabies
virion, the amount ofvirions was increased to
more than 50 ,g of protein and the specific
activity of 3H-labeled nucleotidewas increased
to2,000to2,500mCi/mol. Incubation timewas
also
prolonged
to 12to 15 h. Evenwith suchasimple device, the count ofradioactivity
incor-porated into the acid-insoluble materials was
500to1,000cpm.Theseweresignificantlyover
the background counts, which usually ranged
from 50 to 70 cpm, though far below that of
VSV New Jersey(Table 1, experiment B).
Optimal conditions for incorporation of
[3H]GMP
by the rabies virion. Theoptimal
concentration of NaClwas40mM, whereas that
of VSV New Jerseywas 70 mM (16). Optimal
pH was tested by using
Tris-hydrochloride-buffered solution in therange ofpH 6.8 to8.2,
whichwasdeterminedattheworking
tempera-tureand the concentration utilized in the
exper-iment, andoptimal activitywasobtained atpH
7.4(datanotshown). At various concentrations
of
Tris-hydrochloride
(pH 7.4), the maximalac-tivitywasobtainedataround 40mM, and higher
concentrations of
Tris-hydrochloride
wereinhib-itory. Testing of other Good buffer systems
[BES:
NN'-bis(2-hydroxyethyl)-2-aminoeth-anesulfonic
acid; HEPES:N-2-hydroxyethyl
piperazine-N'-2-ethanesulfonic
acid; TAPS:N-tris-methyl-3-aminopropanesulfonic
acid; TES:N-tris-methyl-2-aminoethanesulfonic
acid; Tricine:
tris-methyl-glycine]
at aconcen-tration of 40 mM (pH 7.4) revealed the
effec-tivenesstobe in theorderof
Tris-hydrochloride,
BES, Tricine, TES, TAPS; however, HEPES
wasmostinhibitory. Therefore,
Tris-hydrochlo-ridewasusedthroughout this experiment.
To determine the
optimal
concentration ofribonucleoside triphosphates, the following
ex-periments were
performed.
(i) Reactionmix-tureswerepreparedtoinclude various
concen-tations butthesamemolar ratiosof four
nucleo-tides. Maximalactivity requiredto incorporate
GMPintoacid-insoluble materialwasobtained
when the nucleotide concentrationwasabout 1
mM. (ii) Reaction mixtures were
prepared
toTABLE 1. Comparisons of
[H]GMP-incorporating
activities of rabies virus and VSV New Jersey under various conditionsAmt of Assay
conditions"
[:H]GMP incorporatedExpt Virus viral Spactof Incuba- pmol/mg
protein Detergent [3H]GMP tion time cpm of protein
(pg)
(mCi/mmol) (h) per hA Rabies 13.2 TritonX-100(0.1%) 133 5 W
VSV 6.6 TritonX-100
(0.1%)
133 512,010
8,290
B Rabies 62 TritonX-100(1%) 2,000 12 543 1.1
VSV 18 TritonX-100
(1%)
2,000
12277,500
1,947
C Rabies 20 NP-40(4%) 200 5 947 140
D VSV 16.5 TritonX-100 (0.2%) 200 3 101,900 23,400
a Assayconditionsfor[3H]GMP-incorporatingactivity ofexperiments A, B, and D were the same as those described for virion transcriptase of VSV,NewJerseyserotype (16) except for themodifications indicated in thistable.Namely,the reaction mixture contained 70mMNaCl, 100 mMTris-hydrochloride (pH8.0),5 mM
Mg2e,
5mM DTT,0.4mM (A andB) or1.0mM (D)ATP, CTP, and UTP, 0.04 mM (A and B) or 0.2 mM (D)GTP, and othersasindicated. Assay conditions for experimentCwith rabies virus were as described in thetext.Reactionswerecarriedoutat300C,
and theacid-insoluble radioactivity was determined.bBackgroundcountsdetermined for the zero-time samples were 94 cpm for experiment A, 70 cpm forB,65 cpm forC,and40cpm forD, andweresubtracted.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.491.56.453.445.575.2]RABIES VIRION TRANSCRIPTASE 829
include various concentrations of cold GTP but
a constant concentration
of
ATP, CTP, andUTP (1
mM)
and3H-labeled GTP (10
uCi pereach reaction mixture).
Astheconcentration ofcold GTP increased,
the radioactivityincorpo-rated
into the acid-insoluble material firstin-creased
and attained amaximal level when thecold GTP was 0.1 to 0.2 mM. When the cold
GTP
wasadded
athigher
concentrations, theincorporation
ofradioactivity decreased
consid-erably, although the total GMP
incorporatedgradually increased. Thus,
when the standardreaction mixture contained
0.2mM GTP and 1mM
ATP,
CTP, and UTP, the maximal
incor-poration of
radioactivity could be obtained,
andthe
total GMP
incorporation
wasabout
two-thirds that obtained when cold GTP
was addedat 1mM.
Requirement
of themetal ion
was investigated incomparison
with that of VSV New Jersey. Theactivity
at various concentrations ofMg2'
reached the
plateau level
ataround 5 mM.Sim-ilar
results
werealso obtained for
VSV New
Jersey.
The
activity of rabies virion
wasstrictly
de-pendent
onthe
presenceof
DTT inthe reaction
mixture.
Less than 10% of maximal
activity
wasobtained
inthe absence of DTT.
The optimal
concentration
was 5mM, but concentrations
higher
than 10 mM wereinhibitory (data
notshown).
Asimilar
dependence
onthesulfhydryl
agentwas
observed with VSV
NewJersey,
andoptimal
activity
was obtained at 5 mM DTT(S. Naito,
personal communication).
The most
conspicuous difference
was foundin the
optimal concentration of
nonionicdeter-gent. In
the
caseofVSV New
Jersey, the
tran-scriptase
activity
wasfully
activated
at0.05%
Triton
X-100, whereas the rabies
virionrequired
4 to
5% for maximum
activity
of
[3H]GMP
in-corporation
(Fig. 1).
Asimilar
high
concentra-tion
wasrequired
when
adifferent
detergent,
NP-40,
wasused.
An anionicdetergent
such assodium
deoxycholate
wasinhibitory
when added
to
the
reaction mixture
at0.125%
(Table 2).
Utilizing
these
optimal conditions,
the
specific
activity of GMP
incorporation by
rabies virionincreased
to 140pmol (Table
1,
experiment
C).
Identification
of the active
entity
that
catalyzes
the
incorporation
of
[3H]GMP.
Ta-ble3shows that the
activity
of[3H]GMP
incor-poration
by
rabies virionrequires
fournucleo-side
triphosphates.
Theactivity
observed in thereaction
mixturelacking
UTP was abouthalf
that in the
complete
system andless
than 1%when
lacking
ATP. In the case ofVSV
NewJersey,
the omission of UTPand
ATPresulted inactivity
of 12 and 7.8% that of thecontrol,
respectively.
Similar results
wereobtained
when-C
0
4Ut
FC
0~~~~~~~~~~
0 >0
VSVewersy. he eacionmixureforrabies
viio -a vrprdaecirus2ntetx n
0
a. ~~~~~~~~~~~~~~~~~~~a.
cotane th niae ocnrabiens ofnnoi
zgofrabies virus 22
0g,A.25of DTT, 0 CTn
A
E
0 0.01 0.1 1 10
Detergent (%)
FIG. 1. Effect of detergent concentration on
I'H]GMP-incorporating
activity byrabies virus and VSV New Jersey. The reaction mixture for rabies virion wasprepared as described in the text and contained theindicated concentrations of nonionicdetergent Triton X-100 (0) or NP-40 (A) and 16.5
a-gofrabies virusprotein. Thereaction mixturefor
VSVNew Jersey contained
.amol
17.5 ofNaCi,
25pmnol
ofTris-hydrochioride
(pH 8.0), 1.25/amol
ofMg2',
1.25/fmol
of DTT,0.1/.onol
ofATP,CTP,andUTP,
[
0.015Polof GTP,2.0it
Ciof[8-3H]GTP
(10.5Ci/mmol),
25 pgofVSV NewJerseyprotein,
and
theindicated
concentrations ofTriton X-100(0).
Afterincubation for10
(rabies
virus)or3(VSV)
hat 30'C,acid-insoluble
radioactivity was
determinedandcal-culated
as shown.[8-(H]ATP
wasused
instead
of[8-iH]GTP;
that
is,
theresidual
activity
of
[3H]AMP
incorpora-tion was
high
in the case ofUTP
omission.Varieties
of
residual
RNApolymerase
activity
when each
nucleotide
wasomitted
werealso
reported
in
manyparamyxovirus preparations
(4, 17, 23).
Aplausible explanation
isthat each
nucleotide
maybe contaminated with others.
However,
supportive data have
notbeen
ob-tained.
The dose response curveinFig.
2shows
that
GMP
incorporation
is
inlinear
proportion
to
the
amountof virions
in the reactionmixture,
as
tested
from 5 to 100 ,ug ofprotein.
Thus,
there
appeared
tobe
noinhibitororcooperative
factor that could
act athigh
concentrations.Sequentially,
theincorporation
commenced with
no or
only
a very shortlag phase
attheinitialstep of incubation and
linearly
increased upto7 to 8 h
(Fig. 3).
The maximumactivity
wasobserved
when the reactionwas carried out ataround
30°C.
The reaction was sensitive toRNase,
but resistanttoDNase,
actinomycin D,
rifampin,
a-amanitin,
andgentamicin
(Table
2).
Theresults indicated that the
activity
was notbrought
aboutby
DNA-dependent
RNA polym-VOL. 24,1977on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.491.270.417.67.246.2]TABLE2. Properties of
[fH]GMP-incorporating
activityof rabies virionpmolof GMP Expt Reactionmixturea incorpo- Ratio
rated/mgof (% protein
1 Complete 34.1 100
+Actinomycin D,2Ag/mI 29.7 87.1 +Actinomycin D,20jyg/ml 28.8 84.5 +DNase I,20,ug/ml 37.0 108 +DNaseI,100ytg/ml 33.8 99.1 +RNaseA, 50tug/ml 0.92 2.7
-Virion <0.44 <1.3
2 Complete 23.4 100
+Rifampin, 20jyg/ml 21.3 90.9 +a-Amanitin,2Iug/ml 23.1 98.8 +Gentamicin,100Jg/ml 23.1 98.8
-Triton X-100 3.5 14.8
-Triton X-100 + DOC 0.59 2.5 (0.125%)
+DOC (0.125%) 3.2 13.6
aThereactionmixture wascomposed of 40 (experiment 1) or 70(experiment 2) mMNaCl,100mMTris-hydrochloride (pH 7.4), 5mMMg2+, 0.4 mMATP, CTP,and UTP,0.04 mMGTP,25 JACi of[8-3H]GTP (10.5 Ci/mmol), 5 mM DTT, 1%TritonX-100, and21,gof viralprotein. The reactionwas carriedout at30°C for12(experiment 1) and10(experiment 2)h.Acid-insolubleradioactivitywasdeterminedasdescribed in the text and calculated in terms ofpicomoles ofGMP incorporated permilligram of viral protein. DOC, Deoxycho-late.
erase derived from host cells or
other
contami-nated
microorganisms,
whichcould
multiply
inthe presence of a
high concentration of
deter-gent.
To
determine
whether or not the enzymethat
catalyzes
[3H]GMP incorporation
wasassoci-ated
withthe rabies
virion, purified virions
werelayered
on a 10 to45%
(wt/vol)
sucrosegradient
and
centrifuged
asdescribed
inMaterials
andMethods. Each fraction
obtained from
thegra-dient was
assayed
forinfectivity,
[3H]GMP-in-corporating
activity,
andprotein
content. Boththese activities
corresponded exactly
tothe
virusband
(Fig.
4). The results andthe
requirement ofdetergent
for the reaction suggest that theenzyme
concerned
with[3H]GMP-incorporating
activity is located in
the core ofthe
rabies
virion.Analysis of the
product.
To examine thenature of the acid-insoluble material, the
3H-labeled product was extracted by cold ethanol
precipitation
after 15 h ofincubation. The
ex-tracts were
examined
forsensitivity
to RNaseand
complementarity
totheviral genomes. Ta-ble4shows
that alarge
portion of product wassensitive toRNase but
completely resistant
toRNasewhen annealed with the addition of
ex-cessviral
RNA. Annealing
experiments revealedthat a
large
portion ofthe product self-annealed,andthis wasinterpreted as follows: the amount
of
3H-labeled
complementary
RNA product wasrelatively small, whereas the
amountof
template
viral RNA in the reaction mixture was so
large
thatthe
product could
hybridize
withoutaddi-tionof viral
RNA.
Analysis of virion
protein
by SDS-PAGE.
The
specific
activity of rabies virion-associated
transcriptase varied
greatly
depending
onvirus
stocks of the
samestrain. The
activity
wasrel-atively high
instocks in which viruses had been
replicated
atthe lower
temperaturesof
33 to33.5°C, but
nosignificant difference
wasfound
in
thermal inactivation of
[3H]GMP-incorporat-ing
activity by the
purified virion
atbetween
33 and37°C (data
notshown). Furthermore,
vi-rusesthat were
propagated
asthefirst
passageof
plaque isolates showed
higher activity than
those
that
replicated after
passagesthrough
BHK
cells.
Although defective
interfering
par-ticles
wereproduced after several
passages,pu-rified
B-particle preparations,
which wereprac-tically free from defective
interfering
particles,
could be obtained
by
velocity
sedimentation
cen-trifugation in the
sucrosedensity gradient,
asdescribed
inMaterials and
Methods. Inaddition,
noinhibitory effect
was observed on thetran-scriptase
activity
of rabies
virionwhen
defective
interfering
particles
wereadded
tothe reactionmixture
(unpublished
observations). Thus,
itwasassumed that a
quantitative
orqualitative
TABLE3. Propertiesof
[fH]GMP-incorporating
activityof rabies and VS virions[3H]GMPincorporated'
Rabies virus VSV NewJersey
Reaction
mixture pmol/ pmol/
mg of Ratio mg of Ratio
pro- M pro- (%
tein tein
perh per h
Complete 80.8 100 23,400 100
-ATP <0.8 <1 1,830 7.8
-CTP 0.68 0.85 170 0.72
-UTP 37.8 46.7 2,800 12
-ATP, CTP, <0.8 <1 190 0.83 UTP
aThecompletereaction mixture(0.25ml)forrabiesvirion contained40 mMNaCl,40mMTris-hydrochloride (pH 7.4), 5mMMg2+,5mMDTT,1.0mMATP, CTP, and UTP, 0.2 mMGTP, 10JACiof[8-3H]GTP (10.5Ci/mmol),4%Triton X-100,and 16.5jigof viral protein. The complete reaction mixture for VSVNewJerseywas asfollows:70mMNaCl, 100 mMTris-hydrochloride (pH 8.0), 5mM Mg2+, 5 mM DTT,1.0mMATP, CTP, and UTP,0.2mMGTP and 10 ,uCi of[3H]GTP,0.2% TritonX-100,and 16.5 ,g ofVSV New Jerseyprotein. The samenucleosidetriphosphate solutions wereusedtoprepare the reactionmixture for both viruses.
bThereaction wascarriedoutfor 5 (rabies virus) and 3 (VSV)h at30°C,and[3H]GMPincorporated into acid-insol-uble material was determined and calculated in terms of picomolesofGMPincorporated per milligram of viral protein perhour.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.491.55.245.66.252.2]RABIES VIRION TRANSCRIPTASE 831
V 0
0
O3
0 C a.2
o I El
0-0 20 40 60 80 100
[image:6.491.87.208.67.245.2]Hg Protein
FIG. 2. Incorporation of
[3H]GMP
by different amountsofrabies virions. Reaction conditionswere asdescribed in Table2(experiment 1), exceptfor the amount of virion. The incorporation ofPHIGMP
wascalculated andexpressed intermsofpicomoles of GMPincorporatedper10h.C._
C-20 a E,o a
0
E0
0 0.
m
0.
(0a, E
0.
4 6 8
Time (hr)
sey. In
addition
tofour
major proteins(G,
N,
Ml, and M2), rabies virus contains several minor
components, such
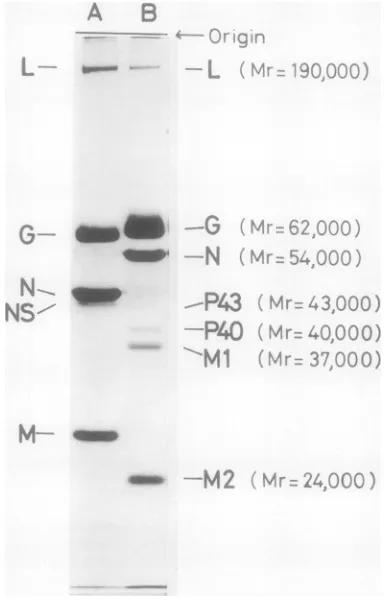
as P40(molecular
weight,40,000), P43
(molecular
weight, 43,000), and Lprotein, a
large-molecular-weight polypeptide
that
correspondsexactly
to that of VSV NewJersey (25). It was noted that the ratio of four major proteins was constant but amounts of
minor components varied
greatly.
Therelative
ratio
of these
polypeptides
was examined forseveral virus
stocks,
which werepropagatedun-der different
conditions by
amicrodensitometer
and
compared withthe
RNApolymerase
activ-ity
(Table
5). The resultsindicate
that the po-lymerase activity was in a fairly linearpropor-tion to the amount of L protein.
On the
otherhand,
nosignificant difference
wasfound
inthecontents
of
four major
proteins.Concerning
other minor components,
noclear
correlation
was
detectable. Thus, it
wasassumed that
there
was a
parallel
relationship between the activity
of
virionpolymerase and
the contentof
Lpro-15
12
i
I-I
E
9
T7
-0
._
'. 3
FIG. 3.
[3H]GMP-incorporating
activityof rabies virionsfordifferenttimes andatdifferent tempera-tures of incubation. Standard reaction mixtures, which contained191ug
ofrabiesvirionprotein, were incubated at 25 (A), 28(5),
30 (0), and 350C (x), andat indicated times samples wereremoved and assayed for acid-insoluble radioactivity. The incorporationofPH]GMPwascalculatedasshown.change might
takeplace
inthe RNApolymerase
of the virion grown under different
conditions,
such as differenttemperatures and viral passage
levels.
Figure 5 shows the
comparison
ofpolypeptide
composition
of rabies virus and VSV NewJer-I.
lk
Ir
x
- E
6-U
D-a
%-
5
- w *Ah
24
0
a-C.)
. 2
1
Fraction
No.
FIG. 4. Locationof3H]GMP-incorporating
activ-ity ofrabies virion by velocity sedimentation on a sucrosedensity gradient.Purifiedrabiesvirionswere
loadedon a10to45%(wt/vol)sucrosegradientand centrifuged at 65,000x gfor90min in an RPS-40 rotor (Hitachi Co. Ltd., Tokyo). Nineteenfractions
obtained were submitted to the determinations of
proteincontent, infectivity, andactivity of
[3H]GMP
incorporation. Proteincontent(0)
wasdetermined by the method ofLowry et al. (13), infectivity (A)wasdeterminedbyplaque formationasdescribed in the text, and
f3H]GMP-incorporating
activity(0)
wasassayedunder the reactionconditions described
in thetextfor12hat30'C.
0
Lon November 10, 2019 by guest
http://jvi.asm.org/
[image:6.491.253.444.301.521.2] [image:6.491.87.209.318.499.2]TABLE 4. Annealing experiment with3H-labeled products of rabies viriona
Treatment Annealed ~th
Acid-insoluble
cpm
RNase resistance
RNase(-) RNase(+) (%)
None None 703b 102b 14.5
Boiled,quickly chilled None 683 121 17.7
Boiled,self-annealed Endogenous RNA 690 530 76.8
Boiled,annealed Excessrabies virion RNA 680 688 100
a
3H-labeled
productswereprepared by the procedures described in the text, using140[Lg
of rabiesvirusprotein.Annealing procedures were also as described in the text. bBackground counts (20 cpm) were subtracted.
tein,
which had the same functionas anintrinsicsubunit
of VSVpolymerase.
Analysis
of the low specific activity
ofrabies vision
polymerase.
The highest
activ-ity
of rabies virion RNA
polymerase
remained atapproximately 1/100
to1/200 of
that of VSV
FIG. 5.
Ekctrophoretic
patternofvision polypep-tidesofrabiesvirus and VSV. TheSDS-solubilized 16.5lig
ofviralproteinsin 504u
wasseparated
byPAGE. (A) VSV New
Jersey;
(B) rabies virus. Themolecular
weight
of eachvision
polypeptide
wases-timated by comparison with several
co-electropho-resedmarkerproteins, such as
fi-galactosidase
(13,-500), bovine serum albumin(67,000), ovalbumin (43,-000), andchymotrypsinogen
(25,000).New Jersey (Table 1). Three possibilities were
proposed
toexplain this:
(i) rapid digestion of
product RNA into acid-soluble material by
RNase, which could not be eliminated during
virus
purification; (ii)
verylow
speed of chain
elongation of RNAsynthesis; and (iii) very low
frequency of RNAsynthesisinitiation.
Detection of
RNaseactivity
inpurified
virions
was carried out with VSV NewJersey, because
a large amount of 3H-labeled complementary
RNA
could
beeasily obtained by VSV
transcrip-tase in
vitro.
This3H-labeled
complementary
RNAwas mixed with variousamounts of
puri-fied
rabiesvirions and incubated for
30 min at300C
inthe presence ofTriton X-100.Activityto convert
3H-labeled
complementary RNAintoacid-solublematerial was not observed.Support
was
also obtained
inanother
experiment
inwhich
the
velocity sedimentation
patternof
product RNA
wasanalyzed.
The'H-labeled
product
from a 14.5-henzyme reactionwaschar-acterized
by
5 to 20%(wt/vol)
sucrose gradientcentrifugation.
Eachfraction
ofthe
gradient
wasdivided into
twoequal
portions,
oneof
which wastreated with
RNase,
asdescribed
inMate-rials and Methods. Results
shown inFig.
6in-dicate that
large-sized
RNA
wasclearly
evident
and
only
asmall
amountof
small-sized
RNAhad accumulated.
Mainpeaks
werepresent from 6 to16S, and
afaint
componentsedimented
faster than 20S.
Asmall
amountof
anRNase-resistant fraction
wasfound
inthe
3-4Speak.
The
sedimentation
patternof
product
RNAfrom a 5-h
incubation
wasquite similar
tothatseenin
Fig.
6. These resultsindicate that
it isunlikelythat product RNA was rapidly digested
by
RNaseactivity
present inthe rabies
virionpreparation.
The other twopossibilities
are nowbeing
quantitatively examined.
DISCUSSION
Virion-bound transcriptase
activity wasdem-onstrated herein in rabies virions
and
was100-to
200-fold lower
than that ofVSV
NewJersey.
The
activity
was ranked atthe
samelevel
asthat seen in Newcastle
disease
virus, Sendai
832
on November 10, 2019 by guest
http://jvi.asm.org/
RABIES VIRION TRANSCRIPTASE 833
TABLE5. Comparison ofpolymerase activity and polypeptide composition of rabies virions produced under variousconditions
Conditions for virus
prondutions Polmear
a Relativecontentof virionpolypeptide'
productions Polymerase activity (pmol of GMP/mg of
Passage Temp proteinperh)b L G N P40
Ml
M2no. (OC)
1 33 140 3.5 41.2 33.1 3.1 6.8 11.8
1 33 125 2.8 37.4 36.2 2.4 5.6 13.5
1 37 79 2.0 48.7 30.6 2.2 5.7 10.4
2 37 46 1.3 37.3 34.8 2.9 6.7 13.2
5 37 16 0.8 49.0 31.4 1.4 5.2 11.3
aBHK cellswereinfectedwithplaque isolates prepared as described in the text (passage 1) or undiluted culture fluids of infected BHKmonolayers (passages2and5).
bPolymerase activitywasassayedwith the standard reaction
mixture,
asdescribed in the text.cRelative contentsof eachpolypeptideweredetermined from theelectrophoretic pattern of SDS-PAGE by
scanningwithamicrodensitometer (model MKIII, Joyce, Loebl & Co.Ltd.)andweighingeachbandcutout
fromtracingpapers.Percentagesof fourmajorandtwominor componentsarelisted;whencontentsof several
minor componentswereless than1%, theywereomitted from thetable.
virus, and
mumps virus (4, 9, 17). At the presenttime,
we arenotofficially permitted
touse anystrain of VSV Indiana
inlaboratories for
biolog-ical research
inJapan. For this
reason, VSVIndiana, the
prototypeof
VSV, could
not beused
asthe reference virus. The
specific
enzymeactivity of VSV
NewJersey, which
wasused inthis
experiment,
was23,400
pmol
of
GMP
incor-porated/mg of viral protein
perh, and
thisvalue
was
considerably higher than that reported
forthe
same serotype ofVSV, but it does
corre-spond
tothat of VSV Indiana (6). Thespecific
activity of virion transcriptase
ofthe
HEP-Flury
strain of rabies virus
wasthus estimated
tobe
around 1/150 that of VSV Indiana.
Based
ondata that(i)
[3H]GMP-incorporating
activity
wasassociated
with theviral
core,(ii)
the
product
was sensitive toRNase,
and(iii)
the
product RNA contained
sequencescomple-mentarytogenome
RNA of the rabies
virus, it
was
concluded
that therabies virus contained
transcriptase.
In
asearch for the
optimal detergent
and
divalent
cationconcentrations,
ionicstrength,
pH,
and temperature, certaindifferences
be-tween
the
rabies virus andVSV
NewJersey
were
evident. The concentration of
nonionicde-tergent
required
wasmuch
higher
for rabies
virus than for
VSV.
Athigher concentrations,
the
enzymereaction showed
asharp decrease,
presumably
duetothe viscous condition of thereaction
mixture. This raised thequestion
ofwhether
thedisruption
of virionswasinsufficientor the viral
envelope
inhibited
the enzymere-action,
asreported
with Sendai virus(14).
Anal-ysis by SDS-PAGE
of virions treated withvar-ious
concentrations
of NP-40 inconditions
sim-ilartothose used for the RNA
polymerase
assayshowed
thataconsiderable
amountofenvelope
x
2
On
C
0
5
10
1520
25
Fraction No.
FIG. 6. Velocity sedimentation analysis of
3H-la-beledproducts. The reaction mixture, which con-tained100jug ofrabies virusprotein, wasincubated for14.5hat
30'C,
and3H-labeled product RNAwas extracted. Theprecipitatewasresolved inSDS-NAE buffer, boiledfor4min, loadedon a 5 to20%sucrose densitygradient,andcentrifuged at130,000 x gfor4hat22'C.Eachof26fractionsobtainedfrom the
centrifuged gradientwasdivided intotwoequal
por-tions, oneof whichwastreated with25 UofRNases A andT1 per mlfor30minat30°C,andthen acid-insoluble radioactivity for all fractions was deter-mined. Symbols:
(0)
untreated;(0)
RNase treated.proteins
(G,
Ml,
andM2)
remained bound invirions
treated
evenwithaconcentration
ashigh
as
5%,
andnosignificant
difference
wasobserved
in the
grade
ofremoval
ofenvelope
proteins
VOL. 24,1977
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.491.271.419.258.459.2]834
KAWAIbetween the treatment with 0.2 and 5% NP-40
(Fig.
7C,
D, andE).
In anothercase,eventhough
two-thirds of the
envelope proteins
werere-moved and most of the N and L proteins were
left behind (Fig. 7B), the polymerase
activity
was not
stimulated,
but rather thispreparation
had one-third the
polymerase activity
ofunfrac-tionated
disrupted
virions(data
notshown).
Thus,
the
inhibitory
effect ofenvelope proteins
was not
demonstrated,
and a reconstitutionex-periment
should be done tofully
rule out thispossibility.
Thistype ofexperiment
hashereto-fore not
been
feasible,
since acomplete removal
of the
envelope proteins
from rabies virionsby
treatment with nonionic
detergent
whilepre-serving
theoriginal
level of RNApolymerase
activity has
notbeen successful.
Furthermore,
5%
deoxycholate
treatmentalmost
completely
removed the
envelope proteins,
butneither
kept
back
Lprotein
norpreserved
RNApolymerase
activity
(Fig. 7A).
Therefore,
therequirement
of a
high
concentration of adetergent
is notfully understood.
Concerning
this
point,
anex-periment
concerning
primary
transcription
when
virus-infected cells have
beenpretreated
with
actinomycin
D andcycloheximide
(5, 24)isof interest. These workers showed that
puri-fied
rabies virioncould
carryoutprimary
tran-scription
ininfected BHKcells,
eventhough the
activity
waslow.
Acertain
intracellular
processof
infection in theearly
stage mayprovide
afavorable
condition that islacking
inthe
invitro
transcriptase
assay.The
RNApolymerase
activity
ofrabies virus
and
VSV New
Jersey
wasstrictly dependent
onthe presence of DTT in
the
reactionmixture.
This strict
dependence
on asulfhydryl
com-pound
is in contrast withfindings
withVSV
Indiana but
corresponds
tofindings
with KernCanyon
virus(1).
The enzyme of thelatter
was alsoessentially
inactivewhen
assayed
in are-action mixture
lacking
asulfhydryl
agent, andDTT was most
effective.
Maximal
yield
ofreleased viruses
wasob-tained
at33"C in
BHKcells
infected with
ourHEP-Flury
strain andother fixed viruses
(7). Asimilar temperature
dependence
was alsoevi-dent in the RNA
polymerase activity
of rabiesviruses grown at 33 and
370C.
Although
thereasonfor
such correlation
is unknown, theen-zyme
activity
was in parallel with the amountof Lprotein obtainedby SDS-PAGE. Although
it has notbeen
established
that Lprotein oftherabies virion is thetranscriptase, nevertheless it
is
possible
that Lprotein
may be the mainintrinsic
subunit,
such as is seen inVSV
tran-scriptase
(8,16). Our
findings herein
showthatthe rabies virion contains RNA polymerase,
even
though
the amount is low compared with-4~~~~~~~~~~~~~~~~~~~~~~.:
l'yR-Oi!
.A t__w
A ,
.~
Tb>,_-_
- m am
FIG. 7. SDS-PAGEofpolypeptides of rabies virion treated withdetergentsunder variousconditions. The
samelotof
purified
rabies virussampleswastreated with: (A) 5% deoxycholate (viralprotein, 100,ug/ml)in thebuffer composed of40mM
NaCl,
5mMMg2+,and40mMTris-hydrochloride (pH7.4)for 15min at20'C; (B) 5% NP-40 (viral protein, 40
pg/ml)
in thebuffer composed of16mMNaCl,
5mMDTT,40 mMTris-hydrochloride (pH 7.4), and2mM EDTA for30min at200C;
(C [control], D, and E) 0, 0.2, and 5% NP-40(viral protein,100,ug/ml)
inthebuffer composed of40mMNaCl,
5mMMg2+,1 mMATP, CTP, and UTP, 0.2mMGTP, 5mMDTT, and 40 mMTris-hydrochloride (pH7.4)for30minat30'C. Eachdetergent-treated virus samplewaslayered on acushionof1mlof 15%sucrosesolutioncontaining 5mM DTT and 40 mMTris-hydrochloride (pH7.4)andcentrifugedat 130,000 x gfor 120min at40C.
Thepellet wasresuspended in theoriginal volume
ofNTbuffer containing5mM DTTandthen
ana-lyzedby SDS-PAGEandpolymerase assay. The rel-ative content of each viral protein was estimated from the electrophoretic pattern with a microdensi-tometer,asdescribed in Table5.Polymerase activity
wasassayedunder the standardconditionsdescribed
in the text. Afterincubation for5h at30'C, acid-insolubleradioactivitywasdetermined.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.491.258.446.64.395.2]RABIES VIRION TRANSCRIPTASE
that of another member of the rhabdovirus
group,VSV New Jersey.
ACKNOWLEDGMENTS
Gratitude is duetoS. Matsumoto for pertinent discussions,
to A.Ishihama and S.Naito for assistancewiththe RNA polymeraseassay,andtoM.Ohara for help with the
manu-script.
This investigation was supported by a grant from the Ministry of Education, Scienceand Culture, Japan.
LITERATURE CITED
1. Aaslestd,H.G.,H. F.Clark,D. H.LBishop,and H.
Koprowski.1971.Comparisonoftheribonucleic acid polymerases oftworhabdoviruses, Kern Canyonvirus andvesicular stomatitis virus. J. Virol. 7:726-735. 2. Baltimore,D. 1971.Expression of animalvirusgenomes.
Bacteriol.Rev. 35:235-241.
3. Baltimore, D., A. S. Huang, andM.Stampfer. 1970. Ribonucleicacidsyntheisofvesicular stomatitis virus. II. AnRNApolymerase in the virion. Proc.Natl.Acad. Sci.U.S.A. 66:572-576.
4. Bernard,J.P., andR.L.Northrop.1974.RNA
polym-eraseinmumpsvirion. J. Virol.14:183-186.
5. Bishop, D.H.L,and A. Flamand. 1975.Transcription
processesof animal RNAviruses,p.95-152. In D. C. Burke and W. C. Russell (ed.),Control processes in virusmultiplication. Cambridge University Press, Cam-bridge.
6. Chang,S. H.,E.Hefti,J. F.Obijeski,and D. H. L. Bishop.1974. RNAtranscription bythe virion
polym-eraseoffive rhabdoviruses. J. Virol.13:652-661. 7. Clark, H. F., and T. J. Wiktor. 1972.
Temperature-sensitivity characteristics distinguishing substrains of fixedrabiesvirus: lack of correlation withplaque-size markersorvirulencefor mice.J.Infect. Dis.125:637-646. 8. Emerson, S.U., and R. R. Wagner. 1973. L protein requirement for in vitro RNAsynthesis by vesicular stomatitisvirus. J. Virol.12:1325-1335.
9. Huang,A. S.,D.Baltimore, and M. A. Bratt. 1971. Ribonucleic acid polymerase in virions of Newcastle disease virus:comparisonwith thevesicular stomatitis viruspolymerase.J. Virol. 7:389-394.
10. Kawai, A.,S.Matsumoto,and K. Tanabe. 1975. Char-acterization ofrabies viruses recovered from
persist-entlyinfected BHK cells. Virology67:520-533. 11. Knudson, D. L. 1973. Rhabdoviruses. J. Gen. Virol.
20:105-130.
12. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly ofthehead of bacteriophage T4. Nature(London)227:680-685.
13. Lowry,0.H., N.J.Rosebrough, A. L. Farr,and R.
J. Randall.1951.Proteinmeasurementwith the Folin phenolreagent.J.Biol. Chem.193:265-275.
14. McSherry,J., and R.Benzinger. 1970. Concentration andpurification of vesicularstomatitis virus by polyeth-yleneglycol "precipitation." Virology40:745-746. 15. Marx, P. A.,A.Porter,and D.W.Kingsbury.1974.
Sendaiviriontranscriptase complex:polypeptide
com-position and inhibition by virion envelope proteins. J. Virol.13:107-112.
16. Naito, S.,and A. Ishihama.1976.Function andstructure ofRNA polymerase from vesicularstomatitis virus. J. Biol.Chem.251:4307-4314.
17. Robinson,W.S.1971.Ribonucleic acid polymerase
ac-tivity in Sendai virions and nucleocapsid. J. Virol. 8:81-86.
18.Scherrer,K. 1969. Isolation andsucrosegradient analysis
ofRNA,p.413-432. In K. Habel and N. P. Salzman (ed.), Fundamental techniques invirology. Academic PressInc.,New York.
19. Schneider,L.G.,and H.Diringer.1976.Structure and molecularbiology ofrabies virus.Curr.Top. Microbiol. Immunol.75:153-180.
20. Sedwick,W.D.,and T. J. Wiktor. 1967.Reproducible plaquingsystem forrabies,lymphocytic choriomenin-gitis,andother RNA viruses inBHK-21/13Sagarose
suspensions. J.Virol. 1:1224-1226.
21. Sokol, F. 1975.Chemical composition andstructure of rabiesvirus,p.79-102.In G. M.Baer(ed.), The natural historyofrabies,vol. 1. AcademicPress Inc.,NewYork. 22. Sokol, F.,and H. F. Clark. 1973.Phosphoproteins, struc-turalcomponentsofrhabdoviruses.Virology52:246-263. 23. Stone,H.O.,A.Portner, and D. W.Kingsbury.1971. Ribonucleic acidtranscriptase in Sendai virions and infected cells. J. Virol. 8:174-180.
24.Villarreal,L.P.,and J. J. Holland. 1974.Transcribing complexesincells infectedbyvesicularstomatitis virus andrabies virus. J. Virol. 14:441-450.
25. Wagner,R.R.,L.Prevec,F.Brown,D. F.Summers, F. Sokol, and R. MacLeod. 1972. Classification of rhabdovirusproteins:aproposal.J.Virol.10:1228-1230.
VOL. 24, 1977
![TABLE 1. Comparisons of[H]GMP-incorporating activities of rabies virus and VSV New Jersey undervarious conditions](https://thumb-us.123doks.com/thumbv2/123dok_us/1536729.106253/3.491.56.453.445.575/table-comparisons-incorporating-activities-rabies-jersey-undervarious-conditions.webp)
![FIG.1.Mg2',I'H]GMP-incorporatingdetergentpmnolculatedvirionacid-insolublecontaineda-gincubationCi/mmol),indicatedVSVVSVUTP,[ Effectofdetergentconcentrationon activity by rabies virus and New Jersey](https://thumb-us.123doks.com/thumbv2/123dok_us/1536729.106253/4.491.270.417.67.246/incorporatingdetergentpmnolculatedvirionacid-insolublecontaineda-gincubationci-indicatedvsvvsvutp-effectofdetergentconcentrationon-activity-rabies-jersey.webp)
![TABLE Properties of ofactivity rabies[fH]GMP-incorporating 2. virion](https://thumb-us.123doks.com/thumbv2/123dok_us/1536729.106253/5.491.55.245.66.252/table-properties-of-ofactivity-rabies-gmp-incorporating-virion.webp)
![FIG. 2.ofamountsasamountwas GMP described Incorporation of [3H]GMP by different of rabies virions](https://thumb-us.123doks.com/thumbv2/123dok_us/1536729.106253/6.491.253.444.301.521/fig-ofamountsasamountwas-gmp-described-incorporation-different-rabies-virions.webp)