0022-538X/00/$04.00⫹0
Copyright © 2000, American Society for Microbiology. All Rights Reserved.
Multiple Blocks to Human Immunodeficiency Virus Type 1
Replication in Rodent Cells
PAUL D. BIENIASZ1*ANDBRYAN R. CULLEN2
Aaron Diamond AIDS Research Center, The Rockefeller University, New York, New York 10016,1and Howard Hughes Medical Institute and Department of Genetics, Duke University Medical Center,
Durham, North Carolina 277102 Received 12 June 2000/Accepted 6 August 2000
The recent identification of human gene products that are required for early steps in the human immuno-deficiency virus type 1 (HIV-1) life cycle has raised the possibility that rodents might be engineered to support HIV-1 infection. Therefore, we have examined the ability of modified mouse, rat, and hamster cell lines to support productive HIV-1 replication. Rodent cells, engineered to support Tat function by stable expression of a permissive cyclin T1 protein, proved to be able to support reverse transcription, integration, and early gene expression at levels comparable to those observed in human cell lines. Surprisingly, however, levels of CD4-and coreceptor-dependent virus entry were reduced to a variable but significant extent in both mouse CD4-and rat fibroblast cell lines. Additional posttranscriptional defects were observed, including a reduced level of un-spliced HIV-1 genomic RNA and reduced structural gene expression. Furthermore, the HIV-1 Gag precursor is generally inefficiently processed and is poorly secreted from mouse and rat cells in a largely noninfectious form. These posttranscriptional defects, together, resulted in a dramatically reduced yield of infectious virus (up to 10,000-fold) over a single cycle of HIV-1 replication, as compared to human cells. Interestingly, these defects were less pronounced in one hamster cell line, CHO, which not only was able to produce infectious HIV-1 particles at a level close to that observed in human cells, but also could support transient, low-level HIV-1 replication. Importantly, the blocks to infectious virus production in mouse and rat cells are recessive, since they can be substantially suppressed by fusion with uninfected human cells. These studies imply the existence of one or more human gene products, either lacking or nonfunctional in most rodent cells that are critical for infectious HIV-1 virion morphogenesis.
Historically, the identification and characterization of host cell gene products that are essential for retroviral replication has been considerably facilitated by genetic complementation in cell lines derived from species that are unable to support the replication of a given retrovirus. The inability of CD4-express-ing nonhuman cells to support human immunodeficiency virus (HIV) and/or simian immunodeficiency virus entry is a partic-ularly noteworthy example (11, 30) and was crucial for the initial identification and, in part, subsequent characterization of chemokine receptors as entry cofactors (2, 3, 10, 14–18, 26, 37, 39). The rodent cell-specific defect in HIV-1 Tat-mediated transactivation provides a second example. In this case, the defect was alleviated by gene products encoded on the human chromosome 12 (1, 21, 35). It later became clear that the mechanistic explanation for the defect was the inability of the murine form of the essential Tat cofactor, CycT1, to support efficient interaction with the TAR element when bound to Tat (5, 19, 20, 27, 46). Therefore, the poor activity of a variety of primate lentiviral Tat proteins in murine cells can be rescued by expression of the human CycT1 protein, which is encoded on human chromosome 12 (46). Alternatively, substitution of a single amino acid in the murine CycT1 protein, tyrosine 261, with its human CycT1 counterpart, cysteine, results in a CycT1 protein that can be efficiently recruited to TAR by Tat proteins and, hence, support transactivation (4, 5, 19, 20, 27).
While these studies illustrate how the complementation of defects in nonpermissive rodent cells has aided the
identifica-tion and characterizaidentifica-tion of molecules that are critical in the HIV-1 life cycle, this work has also raised the possibility that rodents could be engineered to support HIV infection (7, 40). This would provide an inexpensive in vivo model for the eval-uation of therapeutic agents and could aid molecular studies of host-virus interactions at an “organism” level, since both virus and host could be genetically manipulated. However, while expression of CD4, a coreceptor, and a functional CycT1 pro-tein would be predicted to render murine cells permissive for HIV replication, previous work has demonstrated that this is not the case. Murine NIH 3T3 fibroblasts expressing CD4, CCR5, and hCycT1 were found not to be permissive for HIV-1 replication, although these cells were able to support at least some level of gene expression from an integrated provirus (20, 32). Conversely, CHO cells transiently expressing these mole-cules have been reported to support a single cycle of replica-tion following cocultivareplica-tion with HIV-1-infected human cells (47), although in this case, it is likely that infected human cells would fuse with the hamster cells to form heterokaryons.
Previously, few examples of species-specific defects in retro-viral assembly and egress have been described. Simple mam-malian retroviruses, such as murine leukemia virus, can be readily assembled and released by a variety of nonmurine cells (13). Similarly, the foamy retroviruses possess an extremely broad tropism and can be cultivated in a wide range of mam-malian avian and even reptilian cells (22). However, an insect cell-specific defect in simian D-type retroviral particle assem-bly has recently been reported, as has a defect in HIV-1 as-sembly in mouse NIH 3T3 cells (32, 36). In this study, we extend this work and describe defects in HIV-1 replication in rodent cell lines derived from multiple lineages and species. Cumulatively, rodent cell-specific properties can result in an up * Corresponding author. Mailing address: Aaron Diamond AIDS
Research Center, 455 First Ave., New York, NY 10016. Phone: (212) 448-5070. Fax: (212) 448-5159. E-mail: [email protected].
9868
on November 9, 2019 by guest
http://jvi.asm.org/
CD4, CXCR4 and CCR5 (obtained by digestion of pBC12/CMV-based plas-mids) (3) into the retroviral expression vectors LXSH (33) and pBABE Puro (34), respectively.
The pIIIB full-length HIV-1 molecular clone has been described previously (24). Similar pNL/IIIB and pNL/JRFL infectious clones were constructed by replacing the envelope-encoding sequences (SalI-BamHI fragment) of pNL4-3 with pIIIB- and JRFL-derived sequences, respectively. The green fluorescent protein (GFP) reporter viruses R7/3/GFP and R7/YU2/GFP, which contain the enhanced GFP (EGFP) cDNA (Clontech) in place of the Nef open reading frame, were gifts from Mark Muesing. R7/E⫺/GFP was generated by replacing
the envelope sequences (SalI-BamHI fragment) with those of pNL4-3E⫺luc (12),
which contains a defective, frameshifted envelope gene.
Cell lines, viruses, and transfections.Retroviral stocks, generated by transient transfection of Phoenix Gagpol cells (25) with 10g of a retroviral expression vector and 1g of pHIT/G (42), were used to generate stable cell lines express-ing mCycT1(Y261C), CD4, CXCR4, CCR5, or combinations thereof. The cell lines used in this study were derived from mice (NIH 3T3,Mus dunni tail fibroblasts [MDTF], BW5147), rats (Rat2, XC), hamsters (CHO, BHK-21), or humans (HeLa, HOS, CEMx174). Specifically, all rodent cells were engineered to express mCycT1(Y261C), and NIH 3T3, Rat2, and CHO cells were, in addi-tion, transduced with CD4 and either CXCR4- or CCR5-expressing retroviruses. HOS-derived cells expressing CD4 and coreceptors were obtained from the NIH AIDS Reagent Program (GHOST cell lines). Expression of CD4, CXCR4, and CCR5 was verified by fluorescence-activated cell sorter (FACS) analysis using phycoerythrin (PE)-conjugated antibodies (Pharmingen), and where necessary, pure populations of receptor-expressing cells were obtained by sorting. Func-tional expression of mCycT1(Y261C) was verified by enhancement of proviral gene expression (see Results and Fig. 1).
HIV-1 virus stocks were generated by transfection of 293T cells with 10g of pIIIB, pNL/IIIB, pNL/JRFL, pR7/3/GFP, or pR7/YU2/GFP by using Lipo-fectamine Plus (Life Technologies). In some experiments, virions pseudotyped with the vesicular stomatitis virus envelope glycoprotein (VSV-G) were gener-ated by cotransfection of 293T cells with pHIT/G and either pIIIB, pNL4-3E⫺luc, or R7/E⫺/GFP.
Viral replication and entry assays.HIV-1 replication assays were performed with NL/IIIB for CD4⫹CXCR4⫹cells and NL/JRFL for CD4⫹CCR5⫹cells.
Cells were inoculated with approximately 1,000 infectious units (IU) of virus (determined by using CD4⫹CXCR4⫹or CD4⫹CCR5⫹CHO cells and the focal
immunoassay described below). The following day, cells were washed extensively, and p24 levels in the culture supernatant were monitored for the ensuing 10 days. To determine the relative efficiency with which VSV-G and HIV-1 enveloped viruses could enter rodent cell lines, NIH 3T3, Rat2 CHO, and HOS cells expressing CD4 and either CXCR4 or CCR5 were infected with R7/3/GFP, R7/YU2/GFP, or R7/E⫺/GFP(VSV-G). For each envelope, the quantity of virus
used was selected so that 5 to 50% of the most susceptible cell line became infected. The number of infected cells was determined 36 to 40 h postinfection by FACS.
Comparative analysis of HIV-1 production in rodent and human cells during a single cycle of replication.For analysis of HIV-1 production during a single cycle of replication, cell lines lacking HIV-1 receptors were used. Human and rodent cell lines expressing mCycT1(Y261C) (approximately 2.5⫻105
cells/35-mm-diameter dish) were infected with approximately 106 infectious units of
HIV-1IIIB pseudotyped with the VSV-G envelope protein. The following day, cells were washed extensively, and the growth medium was replaced. After a further 24 h, cells and supernatants were harvested, and viral RNA, proteins, and infectivity were analyzed by RNase protection assays, enzyme-linked immunosor-bent assay (ELISA), Western blotting, and focal immunoassays as described below.
Viral RNA analysis.A PCR-generated fragment of pIIIB (nucleotides 78 to 340, relative to the start site of transcription) was inserted into theHindIII and
SmaI sites of pBluescript KS⫹to provide a template for the synthesis of an antisense RNA probe spanning the HIV-1 major 5⬘splice donor. This plasmid was linearized withHindIII, and the probe was generated by transcription with
an antimouse immunoglobulin G-peroxidase conjugate and were developed by using chemiluminescent detection reagents (Roche).
Infectivity assays.Infectious HIV-1 in supernatants was quantitated by using a focal immunoassay similar to those previously described (9, 26). Briefly, CHO cells expressing mCycT1(Y261C), CD4, and either CXCR4 or CCR5 were seeded in 24-well plates and inoculated with serial dilutions of HIV-1-containing supernatant. Forty-eight hours later, the cells were fixed, and foci of infection were enumerated microscopically after sequential incubations with the HIV-1 p24Gag-specific 183 clone H12-C monoclonal antibody and an antimouse
immu-noglobulin G-fluorescein conjugate.
Viral production by heterokaryons.MDTF or Rat2 cells (2.5⫻105
cells/35-mm-diameter dish) were infected with approximately 106 IU of HIV-1IIIB
pseudotyped with the VSV-G envelope protein. Twenty hours later, the cells were washed extensively, trypsinized, and plated with an equal number of MDTF, Rat2, or HOS cells. After 6 h, cells were washed once with serum-free medium and overlayed with a 50% solution of polyethylene glycol (PEG) for 2 min at 37°C. After washing, fresh growth medium was added for 16 to 20 h, and infectious virus production was measured with the focal immunoassay.
RESULTS
Mouse and rat cell lines that express functional CycT1, CD4, and coreceptor proteins do not support HIV-1
replica-tion. Rodent cell lines that are capable of supporting Tat
function were generated by transduction with retroviral vectors that express mCycT1(Y261C). To verify that expression of a functional CycT1 protein was sufficient to increase HIV-1 pro-viral gene expression to a level comparable to that observed in human cell lines, NIH 3T3, Rat2, and CHO cells expressing mCycT1(Y261C) were infected with VSV-G-pseudotyped NL4-3E⫺luc. For comparison, the human cell lines HeLa,
HOS, Jurkat, and CEMx174 were identically infected. As can be seen in Fig. 1A, this analysis revealed that introduction of the mCycT1(Y261C) expression vector into rodent cells was sufficient to increase gene expression from an integrated pro-virus to a level that fell within the range observed in human cell lines. Surprisingly, NL4-3E⫺luc-infected, but otherwise
un-modified, Rat-2 cells expressed high levels of luciferase, com-parable to that seen in human cells. Luciferase expression was only modestly (approximately threefold) enhanced by mCycT1(Y261C). We speculate that Rat2 cells express a form of CycT1 that is able to support at least some level of Tat function. In contrast, mCycT1(Y261C) expression in NIH 3T3 and CHO resulted in 21- and 76-fold increases in proviral gene expression, respectively (Fig. 1A). All subsequent experiments were done with rodent cells that expressed mCycT1(Y261C).
Surprisingly, a large degree of variation in luciferase expres-sion was observed among the various cell lines. However, this phenomenon was found to be a property of this particular reporter virus, and does not appear to be due to genuine differences in HIV-1 long terminal repeat promoter activity. The observed variation was not species or cell type specific and was not observed in other experiments in which similar VSV-G-pseudotyped GFP-expressing reporter viruses were used
on November 9, 2019 by guest
(see Fig. 4A and data not shown) or when wild-type virus was used and viral RNA was analyzed (see Fig. 5).
Since mCycT1(Y261C)-expressing rodent cells are capable of supporting high levels of HIV-1 gene expression, compara-ble to that of permissive human cells, we additionally trans-duced these rodent cells with retroviral vectors that expressed CD4 and one of the two major HIV-1 coreceptors, namely CXCR4 or CCR5. The expression of these molecules was confirmed by FACS, and the widely used HIV-1 permissive HOS-derived CXCR4⫹ and CCR5⫹GHOST cell lines were
included for comparison. As can be seen in Fig. 1B, the levels of expression of CD4, CXCR4, and CCR5 on NIH 3T3, Rat2, and CHO cells were equal to or greater than that measured on GHOST cells.
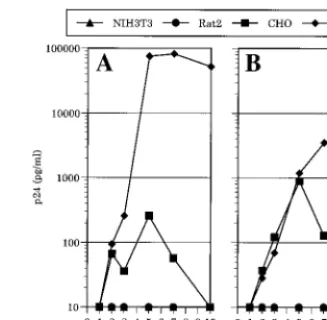
The rodent cells expressing mCycT1(Y261C), CD4, and a coreceptor, as well as GHOST cells were infected with X4- or R5-tropic NL4-3-derived viruses (NL/HXB and NL/JRFL, re-spectively). Virus replication was monitored during the subse-quent 10 days by measuring p24 in culture supernatants. As shown in Fig. 2, GHOST cells that expressed CXCR4 and CCR5 supported rapid and robust replication of HIV-1 NL/ IIIB and NL/JRFL, respectively, but viral replication was not
detected in mCycT1(Y261C)-, CD4-, and coreceptor-express-ing NIH 3T3 or Rat2 cells. Interestcoreceptor-express-ingly, the hamster cell line CHO could support at least some level of HIV-1 replication, although this was transient and clearly at a lower level than that observed in the human GHOST cells (Fig. 2).
Host cell-dependent variation in HIV-1 entry mediated by
CD4 and coreceptors. It has been shown that expression of
CD4 and an appropriate coreceptor on nonhuman cells is sufficient to permit HIV-1 envelope-induced fusion and entry (10, 14–17). Nevertheless, the low level of Tat function in rodent cells has precluded any quantitative comparison of the relative efficiency of the early stages of HIV-1 infection in this context. To examine whether the mouse, rat, and hamster cells could indeed support efficient HIV-1 entry, the cell lines de-scribed in Fig. 1 were infected with HIV-1 reporter viruses that express GFP in place of Nef. Virus entry mediated by the HIV-1IIIB (X4-tropic), YU2 (R5-tropic), and VSV-G enve-lopes was compared by counting the number of infected cells 36 to 40 h later by FACS. An example of these experiments is presented in Fig. 3A; the accumulated data for all of the cell lines examined are shown in Fig. 3B.
Remarkably, large differences (⬎400-fold) in infection effi-ciency were observed among cell lines that expressed similar levels of CD4 and coreceptors (Fig. 3). Specifically, HIV-1 infection of Rat2 cells mediated by the HIV-1IIIB envelope, CD4, and CXCR4 was reduced by more than 400-fold com-pared to that in CHO or HOS cells (Fig. 3). HIV-1IIIB enve-lope-mediated infection of CD4⫹CXCR4⫹NIH 3T3 cells was
also less efficient (approximately 30- to 50-fold) than that of CHO or HOS cells (Fig. 3B). To determine whether this was a coreceptor-specific phenomenon, similar experiments were performed with target cells that expressed CD4 and CCR5 and GFP reporter viruses containing the R5 tropic YU2 envelope. Substantial variation was also observed in the efficiency of YU2 envelope and CD4 CCR5-mediated infection (Fig. 3B). In this case, CD4⫹ CCR5⫹ Rat2 and NIH 3T3 cells were infected
approximately 20- and 6-fold less efficiently than GHOST cells, respectively. Interestingly, CD4⫹CCR5⫹CHO cells were
[image:3.612.349.513.73.233.2]in-fected approximately 10-fold more efficiently than GHOST cells, although the latter difference might, in part, be explained by higher levels of CD4 and CCR5 expression on CHO cells (Fig. 1B). Importantly, these differences in entry efficiency
FIG. 1. Rodent cell lines that express human gene products required for HIV-1 replication. (A) NIH 3T3, Rat2, and CHO cells either unmodified (open bars) or transduced with retroviral vectors that express mCycT1(Y261C) (solid bars). These cell lines, as well as various human cell lines (hatched bars) were infected with NL4-3E⫺luc, pseudotyped with the VSV-G envelope glycoprotein.
Forty-eight hours after infection, luciferase activity in cell lysates was deter-mined. RLU, relative light units. (B) The cell lines in panel A were sequentially transduced first with an LXSH-derived retrovirus vector that expresses CD4 and subsequently with pBABE Puro-derived vectors that express either CXCR4 or CCR5. The expression level of each of these proteins was determined by using PE-conjugated antibodies. CD4⫹CXCR4⫹cell lines are displayed with dark
grey lines, and CD4⫹CCR5⫹cell lines are displayed with pale grey lines. The
parental, mCycT1(Y261C)-expressing cell lines were used as negative controls (black lines). For comparison, HOS cells and the CD4- and coreceptor-positive derivatives (GHOST cell lines) were simultaneously analyzed.
FIG. 2. HIV-1 replication in mCycT1(Y261C)- and CD4-, and coreceptor-expressing rodent cell lines. (A) The CD4⫹CXCR4⫹NIH 3T3, Rat2, CHO, and
HOS cell lines described in the legend to Fig. 1 were infected with NL/IIIB. Alternatively (B) CD4⫹CCR5⫹counterparts were infected with NL/JRFL.
Viral replication was observed for the subsequent 10 days by monitoring the p24 concentration in the culture supernatants by ELISA.
on November 9, 2019 by guest
http://jvi.asm.org/
were envelope specific. Relatively minor variations in the num-bers of GFP-positive cells were observed when cells were in-fected with R7/E⫺/GFP that was pseudotyped with VSV-G
envelope protein (Fig. 3).
Analysis of a single cycle of HIV-1 replication in rodent
cells.Differences in entry efficiency could partly explain why
HIV-1 failed to establish a spreading infection in mouse and rat cells. However, it remained likely that other blocks in the HIV-1 life cycle contributed to rendering viral replication un-detectable in NIH 3T3 and Rat2 cells. In addition, HIV-1 enveloped viruses could enter CHO cells at least as efficiently as HOS cells, yet spreading infection was significantly attenu-ated in the former. To examine which, if any, of the subsequent steps of the HIV-1 life cycle are blocked in rodent cells, a single cycle of HIV-1 replication was examined in detail. In order to determine whether any potential differences were either species specific or cell line specific, additional cell lines from each species were examined. Fibroblasts from wild mice (Mus dunni) as well as a mouse T-cell line, BW5147, were included, as were additional rat (XC), hamster (BHK-21), and human (HeLa and CEMx174) cell lines. All of the nonhuman cell lines were transduced with recombinant LXSN- or MSCV-derived retroviral vectors expressing mCycT1(Y261C), and,
which were slightly more susceptible (approximately threefold) than was typical of the other cell lines. These data indicate that if viral entry restrictions are bypassed, subsequent steps in the HIV-1 life cycle preceding and including early gene expression occur with similar efficiency in human and in mCycT1(Y261C)-expressing rodent cell lines. Importantly, the large differences in susceptibility to VSV-G-pseudotyped HIV-1 infection that do exist among certain mammalian cells (23) were not ob-served in the cell lines used in this study and therefore cannot explain the large differences in subsequent virus yield de-scribed below.
To compare the abilities of rodent and human cells to pro-duce infectious HIV-1 particles, virions pseudotyped with VSV-G and containing an intact X4-tropic HIV-1 genome (HIV-1IIIB) were generated. This virus stock was used to inoculate the panel of rodent and human cell lines, so that 70 to 100% of the cells became infected, as determined by immu-nofluorescence with a p24 monoclonal antibody (data not shown). Thereafter, the cells were washed extensively, and subsequent infectious virus production was determined by us-ing a focal immunoassay with CD4⫹CXCR4⫹CHO cells as
targets (Fig. 4B). Importantly, no infectious virus could be detected in the supernatant of any cell line when similar CD4⫹
CCR5⫹ CHO cells were used as indicator cells (data not
shown). Thus, the washing procedures completely removed the VSV-G-pseudotyped inoculum, and all subsequently produced virus particles contained only the X4-tropic HIV-1IIIB enve-lope.
In contrast to the relatively uniform susceptibility to HIV-1(VSV-G), Fig. 4B documents the extreme variation observed (up to 10,000-fold) in the ability of the panel of cell lines to produce infectious virus. The human cell lines HeLa, HOS, and CEMx174 each produced similar, high levels of infectious virus (⬎107IU/ml). In contrast, a profound defect in infectious HIV-1 production was evident in mouse and rat cell lines, which yielded only 500 to 3,000 IU/ml. The hamster cell lines exhibited an intermediate phenotype, BHK-21 cells produced approximately 2⫻ 104 IU/ml, and CHO cells produced ap-proximately 2⫻106IU/ml, a value only 10-fold less than that observed in human cells. In marked contrast to the extreme interspecies variation in this phenotype, mouse cells that are genetically divergent (MDTF and NIH 3T3) or of a different type (BW5147 T cells) produced remarkably similar levels of infectious virus, as did human cell lines of different lineages.
Reduced levels of genomic HIV-1 RNA in infected rodent
cells.Since events up to and including early gene expression
proceeded efficiently when diverse cell lines were infected with HIV-1(VSV-G) (Fig. 4A), we undertook a systematic exami-nation of subsequent steps in the HIV-1 life cycle to determine why rodent cells do not efficiently produce infectious HIV-1 particles. We first analyzed the level at which HIV-1 mRNA is
FIG. 3. A target cell-specific, CD4- and coreceptor-dependent HIV-1 entry defect. (A) FACS analysis of GFP reporter virus infection of CD4⫹CXCR4⫹
CHO and CD4⫹CXCR4⫹Rat2 cells, mediated by the VSV-G envelope (upper
panels) or by the X4 tropic HIV-1IIIB envelope (lower panels). (B) Analyses identical to that shown in panel A were done with all of the CD4- and corecep-tor-positive cell lines, using HIV-1IIIB enveloped GFP reporter virus for CD4⫹
CXCR4⫹cells and a YU2 enveloped virus for CD4⫹CCR5⫹cell lines. The
proportion of GFP-positive cells was determined by setting a gate at a fluorescent intensity of between 10 and 30, depending on the cell line, so that less than 5 positive events were observed in 50,000 mock-infected cells (0.01%).
on November 9, 2019 by guest
present in spliced and unspliced forms, using ribonuclease pro-tection assays and a probe spanning the 5⬘major splice donor. This analysis, shown in Fig. 5, revealed that spliced HIV-1 RNA was present at similar levels in all of the cells analyzed, as expected (less than 3.5-fold variation). In contrast, the full-length, unspliced HIV-1 transcript is present at lower levels (6-to 40-fold) in each of the nonhuman lines, as compared (6-to the human cell lines. Nevertheless, the levels of the unspliced transcript (Fig. 5) clearly did not correlate with the level of infectious virus production by a given cell line (Fig. 4B). For example, the total amount of HIV-1 RNA and the degree to which it is spliced appear almost identical in MDTF, Rat2, and CHO cells (Fig. 5), but the latter produce an approximately 1,000-fold higher level of infectious virus (Fig. 4B). Thus, while the reduced abundance of the unspliced HIV-1 transcript likely contributes to the observed reduction in infectious HIV-1 production by rodent cells, it appears to play only a minor role in determining this phenotype.
Reduced levels of Gag synthesis, processing, and assembly
into infectious viral particles in rodent cells. The reduced
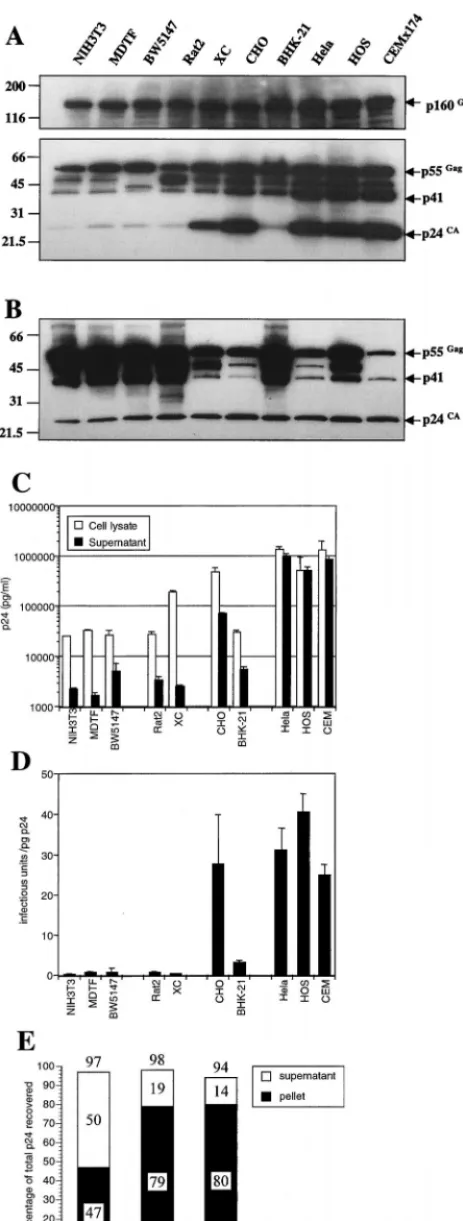
levels of unspliced HIV-1 RNA might be predicted to result in a commensurate reduction in the levels of Gag and Gag-Pol polyprotein expression, since both are translated from un-spliced HIV-1 mRNA. Indeed, as shown in Fig. 6, both West-ern blotting (Fig. 6A) and ELISA (Fig. 6C) analysis of infected cell lysates revealed a significant reduction in the level of Gag proteins that could be detected by using antibodies to the viral capsid protein (p24). When equivalent quantities of cell lysates were examined by Western blotting, however, qualitative as well as quantitative differences in Gag expression among this panel of cell lines were noted (Fig. 6A). While readily detect-able but modest differences in the level of the p55Gag and p160Gag-Pol precursor could be observed, much larger differ-ences in the level of the processed p24CAprotein were evident. Clearly, therefore, the level of intracellular processing of the Gag precursor varied both between and within species.
Specif-ically, the p55Gagprecursor was inefficiently processed in each of the mouse cell lines. Similarly, Rat2 and hamster BHK-21 cells contained relatively low levels of the processed Gag pro-teins compared to p55Gag. In contrast, p55Gagappeared to be quite efficiently processed in the rat cell line, XC, and the hamster cell line, CHO.
[image:5.612.54.294.72.255.2]Importantly, when equal amounts of viral protein, as mea-sured by p24 ELISA (2.5 ng), were loaded onto acrylamide gels, similar levels of fully processed p24CAwere detected by Western analysis, irrespective of the amount of p55Gag, as shown in Fig. 6B. Therefore, the NEN/DuPont p24 ELISA assay used in these experiments selectively measures the fully processed protein and is apparently unaffected by the presence of a large excess of the unprocessed p55Gag. Of note, it has been shown that p55Gagis efficiently processed in human cells, even when expressed at very high or very low levels (32). Therefore, cell line-dependent differences in the degree to which p55Gagis processed are not determined by the expres-sion level per se. Thus, this p24CA-normalized Western anal-ysis (Fig. 6B) permits a semiquantitative assessment of the degree to which p55Gagis processed within the virus-producing cell, independent of its abundance. Interestingly, there were clear intraspecies differences in the extent to which intracellu-lar Gag protein was processed (Fig. 6A and B). Among rat cell lines, p55Gagwas processed more efficiently in XC than in Rat2 cells, and in hamsters, p55Gagwas processed more efficiently in CHO than in BHK-21 cells. Even among human cell lines, p55Gag was processed more efficiently in the CEMx174 and HeLa cell lines than in the HOS line. Conversely, the mouse cell lines exhibited a uniform and profound defect in the de-gree to which p55Gagwas processed. Given these findings, it is FIG. 4. Posttranscriptional defects in infectious virus production from rodent
cells during a single cycle of HIV-1 replication. (A) Rodent and human cells are similarly susceptible to HIV-1(VSV-G) infection. The indicated cell lines were infected with R7/E⫺/GFP pseudotyped with VSV-G. The number of infected
cells per well was determined by multiplying the percentage of GFP-positive cells by the total number of cells in each well. (B) Cell lines were infected with replication-competent, X4-tropic but VSV-G-pseudotyped HIV-1IIIB. After ex-tensive washing, infectious virus production was analyzed by using CD4⫹
CXCR4⫹CHO cells and a focal immunoassay, as described in Materials and
[image:5.612.314.547.77.308.2]Methods. FFU, focus-forming units.
FIG. 5. Analysis of HIV-1 RNA in infected cells. The indicated HIV recep-tor-negative cell lines were infected with VSV-G-pseudotyped HIV-1 as de-scribed in the legend to Fig. 4B. Forty-eight hours later, total RNA was extracted from infected cells and analyzed by ribonuclease protection assay. Arrows indi-cate the predicted migration of the 317-nucleotide undigested probe (P) which spans the major 5⬘splice donor site, resulting in two protected fragments of 262 and 213 nucleotides that correspond to unspliced (U) or spliced (S) HIV-1 RNA, respectively. The numbers below the lanes indicate the levels of each of the RNA species, determined by PhosphorImager analysis and expressed in arbitrary units. M, molecular size markers.
on November 9, 2019 by guest
http://jvi.asm.org/
decipher to what degree this is a reflection of the level of unspliced Gag mRNA, p55Gag processing, particle assembly and egress, or protein stability. Each of these properties is either likely to be or is demonstrably variable among the cell lines tested. Conversely, the level of the unspliced viral RNA (Fig. 5) could be, in turn, affected by the level of Gag protein which might either stabilize viral RNA by packaging into viral particles or reduce the apparent level by the budding of virions from the cell.
The level of processed p24CAprotein in the supernatant of most of the infected rodent cells was even more dramatically reduced than in cell lysates (Fig. 6C). With the exception of CHO cells, infected rodent cell supernatants contained 1.9 to 6.3 ng of p24 per ml, values which were approximately 100- to 500-fold lower than those observed with human cells. In fact, all of the rodent cells contained 4- to 130-fold-lower concen-trations of processed p24 in culture supernatants than the corresponding cell lysates, whereas p24 concentrations were approximately equal in the lysates and supernatants of infected human cells (Fig. 6C). Thus, in addition to reduced levels of intracellular p55Gagprocessing, rodent cells display a signifi-cant defect in viral egress.
Furthermore, the level of intracellular p55Gagprocessing did not correlate with differences in the extent to which processed p24CAwas released from infected cells (Fig. 6A, B, and C). Specifically, p55Gagwas less efficiently processed in HOS cells than it was in either HeLa or CEMx174 cells, but processed p24 was released into the cell supernatant at least as efficiently as by the other human cell lines (Fig. 6B and C). Conversely, the p55Gagprecursor was processed in CHO cells with similar efficiency to that in HeLa and CEM cells and more efficiently than that in HOS cells, yet p24 was released five- to sevenfold less efficiently than from these human cell lines. Finally, p55Gag was more efficiently processed in XC cells than in HOS cells, yet the processed p24 was secreted from the XC cells almost 100-fold less efficiently (Fig. 6B and C). Overall, it appears that the level of intracellular processing of p55Gag is a separate phenotype from the degree to which the processed protein is secreted from the cell.
[image:6.612.57.289.72.683.2]In addition to defects in Gag expression, processing, and
FIG. 6. Reduced levels of Gag expression, processing, and assembly into infectious particles in rodent cell lines. The same cells that generated the infec-tious virus measured in Fig. 4B as well as the corresponding supernatant samples were analyzed for viral Gag protein content. (A) Equivalent quantities (10g) of cell lysate were separated on acrylamide gels and subjected to Western blot
analysis with a p24-specific monoclonal antibody. Note that the upper panel, which displays the p160Gag-Polprecursor, represents a longer exposure than the
lower panel. (B) Aliquots of cell lysates containing equivalent amounts of p24 (2.5 ng) as measured by p24 ELISA were analyzed by Western blotting as in panel A. (C) Viral p24 in cell lysates and supernatants as measured with a commercial ELISA kit. (D) The quantity of infectious virus produced relative to the total amount of supernatant p24 was calculated for each cell line. (E) Relative proportions of the total supernatant p24 that could be pelleted by ultracentrifugation through a sucrose cushion. Numbers refer to the percentage of the starting material that was recovered in each fraction and their sum.
on November 9, 2019 by guest
egress from rodent cells, the small amount of p24 in the su-pernatant media of most infected rodent cell lines was pre-dominantly not in the form of infectious virus particles. In fact, as shown in Fig. 6D, virus secreted from the human cell lines contained 24 to 40 IU/pg of p24, whereas mouse- and rat cell-derived virus contained 0.2 to 0.8 IU/pg. The BHK-21-derived virus was more infectious (approximately 3 IU/pg) than that derived from mouse and rat cells, while the CHO-derived virus exhibited a similar infectivity to that produced by human cells. However, neither the degree to which p55Gagwas processed within the cell nor the relative efficiency with which processed p24 was released into the supernatant was predictive of the infectivity of the virus produced by a given cell line.
To examine whether apparent differences in the infectivity of virus preparations derived from rodent and human cells were due to the presence of nonparticulate p24CA, filtered culture supernatants from infected NIH 3T3, Rat2, and HeLa cells were layered above a 20% sucrose cushion and subjected to ultracentrifugation. Thereafter, pelleted and nonpelleted p24CAproteins were measured by ELISA. As can be seen in Fig. 6E, despite the widely different infectivities of virus prep-arations derived from Rat2 and HeLa cells (Fig. 6D), remark-ably similar proportions (approximately 80%) of the total su-pernatant p24CAcould be pelleted through 20% sucrose. Virus preparations derived from NIH 3T3 contained a somewhat smaller proportion of pelletable p24CA, but this difference was relatively slight (less than twofold). In fact, when the infectivity of NIH 3T3- and Rat-2 cell-derived virus was compared with that derived from HeLa cells, based only on pelletable p24CA, values were approximately 50- and 30-fold lower in the case of the murine and rat cell lines, respectively.
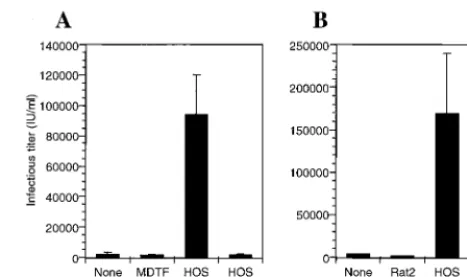
Fusion with uninfected human cells enhances the
produc-tion of infectious virions from infected rodent cells. The
re-duced ability of rodent cells to produce infectious HIV-1 par-ticles could be due to a lack of essential cellular cofactors that are required for virus assembly and egress. Alternatively, ro-dent cells could express a dominant inhibitor of these pro-cesses. To address this question, HIV-1-infected rodent cells were generated by using HIV-1IIIB(VSV-G) and subsequently fused, using PEG, with uninfected human HOS cells. As con-trols, the infected rodent cells were also fused with uninfected autologous cells, or were cocultivated with human cells, but not fused. As can be seen in Fig. 7, fusion of infected MDTF or Rat2 cells with uninfected human HOS cells resulted in a dramatic increase (50- to 200-fold) in the level of infectious virus production compared to that of unfused cells or cells that were fused with MDTF or Rat2 controls. While fusion with human cells did not restore levels of virus production to those observed if the HOS cells had been infected directly, we esti-mated visually that approximately 10% of the cells in the mixed cultures actually fused after PEG treatment. This being true, the observed 50- to 200-fold increase in viral production is quite close (within 1 order of magnitude) to the result that would be predicted if the human cell phenotype was com-pletely restored in rodent-human cell heterokaryons. Further-more, and as can be seen in Table 1, the predominant effect of fusion with human cells was to increase the apparent infectivity of the supernatant p24CA(approximately 35-fold) rather than to increase the total amount of p24CAsecreted (approximately 2- to 3-fold). Minor or no effects on either total p24CA pro-duction or virus infectivity were observed when only rodent cells were fused. These data suggest that the small number of rodent or human heterokaryons efficiently produced highly infectious virus, and the simplest explanation for these results is that human cells express a factor or factors required for
infectious HIV-1 production that are either lacking or non-functional in rodent cells.
DISCUSSION
[image:7.612.314.548.72.210.2]In this study, we have characterized the ability of a number of rodent cell lines to support HIV-1 replication. Clearly, the expression of functional CycT1, CD4, and coreceptor mole-cules in most rodent cells was insufficient to constitute a per-missive environment for HIV-1 replication. In fact, multiple additional blocks to HIV replication were observed among rodent cells. In most cases, these blocks were sufficiently severe to render HIV-1 replication undetectable. Nevertheless, CHO cells were found to be exceptional among the rodent cell lines examined in that at least some degree of viral replication could be observed. Upon more detailed analysis, the differences ob-served between NIH 3T3, Rat2, CHO, and HOS cells during single-cycle virus replication assays (Fig. 4) correlated well with the viral growth phenotypes shown in Fig. 2. Cumulatively, viral entry and production defects in NIH 3T3 and Rat2 cells would be predicted to result in a roughly 105-fold (per cycle) replication defect as compared to human cells. Given these findings, it is not surprising that little or no viral replication is
[image:7.612.312.551.571.670.2]FIG. 7. Fusion of HIV-1-infected rodent cells with uninfected human cells enhances infectious particle release. MDTF (A) or Rat2 (B) cells were infected with VSV-G-pseudotyped HIV-1 as described in the legend to Fig. 4. The following day, the cells were washed, trypsinized, and replated, either alone (None) or along with an equal number of MDTF, Rat2, or HOS cells. Coculti-vated cells were fused with PEG, except where indicated, and subsequent infec-tious virus production was analyzed after 16 h.
TABLE 1. The effect of fusion with human cells on p24CAand
infectious virus production by HIV-1-infected rodent cellsa
Infected cell (⫻fusion partner)
p24 in supernatant
(ng/ml)b
Infectious virus titer (IU/ml)c
Infectivity (IU/pg of p24)
MDTF 4.1 (1.0)d 700 (1.0) 0.17 (1.0)
MDTF⫻MDTF 3.1 (0.8) 1,000 (1.4) 0.31 (1.8) MDTF⫻HOS 11.4 (2.8) 68,000 (97.1) 5.92 (34.8)
Rat2 11.2 (1.0) 4,000 (1.0) 0.35 (1.0)
Rat2⫻Rat2 10.5 (0.9) 2,600 (0.7) 0.24 (0.7) Rat2⫻HOS 19.8 (1.8) 240,000 (60.0) 12.12 (34.6)
aSupernatant fluid was harvested from HIV-1-infected rodent cells after PEG
fusion with homologous rodent cells or with human HOS cells.
bViral p24CAantigen was determined by ELISA.
cInfectious virus titer was determined by using the focal immunoassay as
described in Materials and Methods.
dNumbers in parentheses indicate values relative to unfused cells, which were
arbitrarily set at 1.0.
on November 9, 2019 by guest
http://jvi.asm.org/
decrease in YU2 enveloped HIV-1 entry into CD4 CCR5 NIH 3T3 compared to GHOST cells is in good agreement with a previous result obtained with the JRFL envelope (32). At present, the molecular basis for this entry defect in mouse and rat fibroblasts is unclear. It is likely that the block is at the level of envelope-receptor binding or membrane fusion, since no such defects were observed when using VSV-G-pseudotyped virus stocks. In addition, CD4⫹CXCR4⫹Rat2 cells (and to a
lesser extent NIH 3T3 cells) are substantially resistant to for-mation of syncytia when cocultivated with HIV-1IIIB-express-ing cells (P.D.B., unpublished data). InterestHIV-1IIIB-express-ingly, previous evidence has suggested that glycosylation might influence the activity of coreceptors. Most recently, the introduction of mu-tations at potential glycosylation sites in the CXCR4 molecule was shown to result in a coreceptor that could be used by HIV-1 strains that are ordinarily CCR5 tropic (8). Other re-ports have indicated that deglycosylation of cell surfaces by pretreatment with inhibitors resulted in higher levels of CD4-independent entry by a particular HIV-2 strain (38, 44). It is quite possible that species or cell-type-dependent differences in coreceptor glycosylation and/or multimerization (28) could determine the apparent differences in activity observed when CXCR4 and CCR5 are expressed on different cell lines. Alter-natively, it is conceivable that additional species or cell-type-specific molecules play a role (either positive or negative) in mediating HIV-1 entry.
Reduced abundance of unspliced HIV-1 RNA and Gag
pro-teins in rodent cells.Following viral entry, the HIV-1 life cycle
appeared to proceed efficiently, independently of the cell spe-cies or type, up to and including early gene expression. There-after, profound defects in the ability of many rodent cell lines to produce infectious HIV-1 were observed. While the levels of spliced HIV-1-specific transcripts were approximately equal in all cell lines examined, the abundance of the unspliced, genomic transcript was significantly reduced in rodent cells (Fig. 5). This phenotype was more profound than that recently reported (32) and is similar to the predicted consequence of a defect in Rev function (45). However, it has been clearly shown that Rev is functional in rodent and even avian cells (31). Thus, the reduced abundance of genomic HIV-1 RNA in rodent cell lines is likely due either to oversplicing or to a reduced stability of the unspliced transcript in rodent compared to human cells. In fact, HIV-1 transcripts have been previously shown to splice significantly more efficiently in mouse than in human cells (31). Since unspliced retroviral transcripts also serve as mRNAs for the translation of Gag and Gag-Pol polyprotein precursors, a reduced level of unspliced HIV-1 RNA in rodent cells might be predicted to result in lower levels of expression of these proteins. In fact, comparative quantitation of the absolute level ofgag gene expression proved to be complex (Fig. 6). There were clear quantitative differences among the cell lines in
proportion of the total p24 that was present in the culture supernatant. With the exception of the hamster cell line CHO, all rodent cells secreted uniform and low levels of p24 (⬍7 ng/ml) into culture supernatants, values that were up to 500-fold lower than those observed with human cells. This defect in p24 secretion was not universally linked to defects in intracel-lular Gag processing, since p55Gag processing in the rat cell line XC was at least as efficient as that in the human cell line HOS, but XC cells nevertheless displayed a profound defect in mediating p24CAegress. In marked contrast to the other ro-dent cell lines used in single-cycle replication assays, CHO cell cultures secreted levels of p24CAthat were only 10-fold lower than that which was typical of human cell lines. The reduced quantity of p24CAin CHO cell supernatants compared to that in human cells was partly due to a small reduction in overall Gag protein expression, possibly due to the reduced abun-dance of unspliced HIV-1 RNA. However, the generalized rodent cell defect in p24CAegress was at least partly conserved in CHO cells, because p24 release was reduced approximately sevenfold compared to that in human cells.
Among rodent cells, CHO cells were also exceptional in that the infectivity of the p24CAreleased into the culture superna-tant was similar to that derived from human cells. Conversely, the majority of p24CAin the supernatant of the other cell lines was not in the form of infectious particles (Fig. 6). Thus, in most rodent cell lines, residual p24 secretion is composed of a small number of infectious particles as well viral proteins in a noninfectious, but largely particulate form. These results are in contrast to those of recent study that indicated that the small amount of p24CAreleased by NIH 3T3 cells was almost as infectious as that derived from human cells (32). The reasons for this discrepancy are unclear at present, but the two studies are otherwise largely consistent in reporting a profound, but not absolute, block in the assembly of infectious HIV-1 parti-cles in rodent cells.
Defective infectious virus production in rodent cells is
res-cued by factors present in human cells.Importantly, the block
in infectious virus production in mouse and rat cells could be substantially relieved by fusion with uninfected HOS cells (Fig. 7). Multiple and complex hypotheses can be devised to account for this result. It is possible, for example, that human cells could express a factor that blocks the function or expression of a rodent cell-specific inhibitor of infectious HIV-1 production. However, the simplest, and most likely scenario, is that human cells provide one or more cofactors that enhance the assembly and/or release of infectious HIV-1 virions. This is the first evidence of which we are aware of a species-restricted positive cofactor(s) involved in the egress of infectious retrovirus par-ticles. At present, we can only speculate as to its identity and mechanism of action. It has been reported that infected HIV-1 NIH 3T3 cells contain electron-dense vesicle-associated
on November 9, 2019 by guest
rial (32), which may be aggregates of the p55Gagprecursor, and very few budding virions. Conceivably, therefore, cellular fac-tors could be required for specific targeting of viral proteins to the plasma membrane prior to assembly and budding. Alter-natively, mistargeting of the Gag precursor could be entirely secondary to a particle assembly defect. Clearly, the pheno-types described herein are not readily explained by a lack of Vif target proteins in HIV-1-producing rodent cells. The func-tional targets of Vif proteins are likely to be dominant-negative inhibitors of viral infectivity (29, 41) and do not affect virion egress. Similarly, the potential lack of a functional cyclophilin in rodent cells is not an adequate explanation for this pheno-type. A genetically or pharmacologically induced lack of cyclo-philin in human cell-derived HIV-1 particles results in a phe-notype that is only manifested in subsequent target cells and is dramatically different from that described here (6, 43).
The data contained in this report provide compelling evi-dence for the existence of cellular factors that play a critical and specific role in the late stages of infectious HIV-1 particle production. The identification of such factors is likely to be critical for the successful development of murine models of AIDS and could provide additional targets for therapeutic intervention in HIV infection.
ACKNOWLEDGMENTS
We thank Therese Grdina for excellent technical assistance and Cecelia Cheng-Mayer, Theodora Hatziioannou, Mark Muesing, and Alexandra Trkola for gifts of cell lines and other reagents. Hybridoma 183 (clone H12-5C) and GHOST cell lines were obtained through the AIDS Research and Reference Reagent program, Division of AIDS, NIAD, NIH from Bruce Cheseboro and Vineet Kewalramani and Dan Littman, respectively.
This work was supported by the Donald A. Pels Charitable Trust, the Howard Hughes Medical Institute, and a grant from the National Institute of Allergy and Infectious Diseases (IR01AI42538) to B.R.C.
REFERENCES
1.Alonso, A., D. Derse, and B. M. Peterlin.1992. Human chromosome 12 is required for optimal interactions between Tat and TAR of human immu-nodeficiency virus type 1 in rodent cells. J. Virol.66:4617–4621.
2.Atchison, R. E., J. Gosling, F. S. Monteclaro, C. Franci, L. Digilio, I. F. Charo, and M. A. Goldsmith.1996. Multiple extracellular elements of CCR5 and HIV-1 entry: dissociation from response to chemokines. Science274:
1924–1926.
3.Bieniasz, P. D., R. A. Fridell, I. Aramori, S. S. Ferguson, M. G. Caron, and B. R. Cullen.1997. HIV-1-induced cell fusion is mediated by multiple re-gions within both the viral envelope and the CCR-5 co-receptor. EMBO J.
16:2599–2609.
4.Bieniasz, P. D., T. A. Grdina, H. P. Bogerd, and B. R. Cullen.1999. Analysis of the effect of natural sequence variation in Tat and in cyclin T on the formation and RNA binding properties of Tat-cyclin T complexes. J. Virol.
73:5777–5786.
5.Bieniasz, P. D., T. A. Grdina, H. P. Bogerd, and B. R. Cullen.1998. Recruit-ment of a protein complex containing Tat and cyclin T1 to TAR governs the species specificity of HIV-1 Tat. EMBO J.17:7056–7065.
6.Braaten, D., E. K. Franke, and J. Luban.1996. Cyclophilin A is required for an early step in the life cycle of human immunodeficiency virus type 1 before the initiation of reverse transcription. J. Virol.70:3551–3560.
7.Browning, J., J. W. Horner, M. Pettoello-Mantovani, C. Raker, S. Yurasov, R. A. DePinho, and H. Goldstein.1997. Mice transgenic for human CD4 and CCR5 are susceptible to HIV infection. Proc. Natl. Acad. Sci. USA94:
14637–14641.
8.Chabot, D. J., H. Chen, D. S. Dimitrov, and C. C. Broder.2000. N-linked glycosylation of CXCR4 masks coreceptor function for CCR5-dependent human immunodeficiency virus type 1 isolates. J. Virol.74:4404–4413. 9.Chesebro, B., K. Wehrly, J. Nishio, and S. Perryman.1992.
Macrophage-tropic human immunodeficiency virus isolates from different patients exhibit unusual V3 envelope sequence homogeneity in comparison with T-cell-tropic isolates: definition of critical amino acids involved in cell tropism. J. Virol.66:6547–6554.
10. Choe, H., M. Farzan, Y. Sun, N. Sullivan, B. Rollins, P. D. Ponath, L. Wu, C. R. Mackay, G. LaRosa, W. Newman, N. Gerard, C. Gerard, and J. Sodroski.1996. The beta-chemokine receptors CCR3 and CCR5 facilitate infection by primary HIV-1 isolates. Cell85:1135–1148.
11. Clapham, P. R., D. Blanc, and R. A. Weiss.1991. Specific cell surface requirements for the infection of CD4-positive cells by human immunode-ficiency virus types 1 and 2 and by simian immunodeimmunode-ficiency virus. Virology
181:703–715.
12. Connor, R. I., B. K. Chen, S. Choe, and N. R. Landau.1995. Vpr is required for efficient replication of human immunodeficiency virus type-1 in mono-nuclear phagocytes. Virology206:935–944.
13. Cosset, F. L., Y. Takeuchi, J.-L. Battini, R. A. Weiss, and M. K. L. Collins.
1995. High-titer packaging cells producing recombinant retroviruses resis-tant to human serum. J. Virol.69:7430–7436.
14. Deng, H., R. Liu, W. Ellmeier, S. Choe, D. Unutmaz, M. Burkhart, P. Di Marzio, S. Marmon, R. E. Sutton, C. M. Hill, C. B. Davis, S. C. Peiper, T. J. Schall, D. R. Littman, and N. R. Landau.1996. Identification of a major co-receptor for primary isolates of HIV-1. Nature381:661–666.
15. Deng, H. K., D. Unutmaz, V. N. KewalRamani, and D. R. Littman.1997. Expression cloning of new receptors used by simian and human immunode-ficiency viruses. Nature388:296–300.
16. Doranz, B. J., J. Rucker, Y. Yi, R. J. Smyth, M. Samson, S. C. Peiper, M. Parmentier, R. G. Collman, and R. W. Doms.1996. A dual-tropic primary HIV-1 isolate that uses fusin and the beta-chemokine receptors CKR-5, CKR-3, and CKR-2b as fusion cofactors. Cell85:1149–1158.
17. Dragic, T., V. Litwin, G. P. Allaway, S. R. Martin, Y. Huang, K. A. Na-gashima, C. Cayanan, P. J. Maddon, R. A. Koup, J. P. Moore, and W. A. Paxton.1996. HIV-1 entry into CD4⫹cells is mediated by the chemokine receptor CC-CKR-5. Nature381:667–673.
18. Feng, Y., C. C. Broder, P. E. Kennedy, and E. A. Berger.1996. HIV-1 entry cofactor: functional cDNA cloning of a seven-transmembrane, G protein-coupled receptor. Science272:872–877.
19. Fujinaga, K., R. Taube, J. Wimmer, T. P. Cujec, and B. M. Peterlin.1999. Interactions between human cyclin T, Tat, and the transactivation response element (TAR) are disrupted by a cysteine to tyrosine substitution found in mouse cyclin T. Proc. Natl. Acad. Sci. USA96:1285–1290.
20. Garber, M. E., P. Wei, V. N. KewalRamani, T. P. Mayall, C. H. Herrmann, A. P. Rice, D. R. Littman, and K. A. Jones.1998. The interaction between HIV-1 Tat and human cyclin T1 requires zinc and a critical cysteine residue that is not conserved in the murine CycT1 protein. Genes Dev.12:3512– 3527.
21. Hart, C. E., C. Y. Ou, J. C. Galphin, J. Moore, L. T. Bacheler, J. J. Wasmuth, S. R. Petteway, Jr., and G. Schochetman.1989. Human chromosome 12 is required for elevated HIV-1 expression in human-hamster hybrid cells. Sci-ence246:488–491.
22. Hill, C. L., P. D. Bieniasz, and M. O. McClure.1999. Properties of human foamy virus relevant to its development as a vector for gene therapy. J. Gen. Virol.80:2003–2009.
23. Hofmann, W., D. Schubert, J. LaBonte, L. Munson, S. Gibson, J. Scammell, P. Ferrigno, and J. Sodroski.1999. Species-specific, postentry barriers to primate immunodeficiency virus infection. J. Virol.73:10020–10028. 24. Hwang, S. S., T. J. Boyle, H. K. Lyerly, and B. R. Cullen.1991. Identification
of the envelope V3 loop as the primary determinant of cell tropism in HIV-1. Science253:71–74.
25. Kinsella, T. M., and G. P. Nolan.1996. Episomal vectors rapidly and stably produce high-titer recombinant retrovirus. Hum. Gene Ther.7:1405–1413. 26. Kuhmann, S. E., E. J. Platt, S. L. Kozak, and D. Kabat.1997. Polymorphisms
in the CCR5 genes of African green monkeys and mice implicate specific amino acids in infections by simian and human immunodeficiency viruses. J. Virol.71:8642–8656.
27. Kwak, Y. T., D. Ivanov, J. Guo, E. Nee, and R. B. Gaynor.1999. Role of the human and murine cyclin T proteins in regulating HIV-1 tat-activation. J. Mol. Biol.288:57–69.
28. Lapham, C. K., M. B. Zaitseva, S. Lee, T. Romanstseva, and H. Golding.
1999. Fusion of monocytes and macrophages with HIV-1 correlates with biochemical properties of CXCR4 and CCR5. Nat. Med.5:303–308. (Erra-tum,5:590.)
29. Madani, N., and D. Kabat.1998. An endogenous inhibitor of human immu-nodeficiency virus in human lymphocytes is overcome by the viral Vif pro-tein. J. Virol.72:10251–10255.
30. Maddon, P. J., A. G. Dalgleish, J. S. McDougal, P. R. Clapham, R. A. Weiss, and R. Axel.1986. The T4 gene encodes the AIDS virus receptor and is expressed in the immune system and the brain. Cell47:333–348. 31. Malim, M. H., D. F. McCarn, L. S. Tiley, and B. R. Cullen.1991. Mutational
definition of the human immunodeficiency virus type 1 Rev activation do-main. J. Virol.65:4248–4254.
32. Mariani, R., G. Rutter, M. E. Harris, T. J. Hope, H.-G. Krausslich, and N. R. Landau.2000. A block to human immunodeficiency virus type 1 assembly in murine cells. J. Virol.74:3859–3870.
33. Miller, A. D., D. G. Miller, J. V. Garcia, and C. M. Lynch.1993. Use of retroviral vectors for gene transfer and expression. Methods Enzymol.217:
581–599.
34. Morgenstern, J. P., and H. Land.1990. Advanced mammalian gene transfer: high titre retroviral vectors with multiple drug selection markers and a complementary helper-free packaging cell line. Nucleic Acids Res.18:3587– 3596.
on November 9, 2019 by guest
http://jvi.asm.org/
Tahara, and A. Koito.1998. Disturbed CD4⫹T cell homeostasis and in vitro HIV-1 susceptibility in transgenic mice expressing T cell line-tropic HIV-1 receptors. J. Exp. Med.187:1439–1449.
41. Simon, J. H., N. C. Gaddis, R. A. Fouchier, and M. H. Malim.1998. Evidence for a newly discovered cellular anti-HIV-1 phenotype. Nat. Med.4:1397–
462.
47. Wimmer, J., K. Fujinaga, R. Taube, T. P. Cujec, Y. Zhu, J. Peng, D. H. Price, and B. M. Peterlin.1999. Interactions between Tat and TAR and human immunodeficiency virus replication are facilitated by human cyclin T1 but not cyclins T2a or T2b. Virology255:182–189.