JOURNALOFVIROLOGY,Sept. 1993,p.5260-5268 0022-538X/93/095260-09$02.00/0
Copyright© 1993,American Society for Microbiology
Identification, Sequence, and Transcriptional Mapping of lef-3,
a
Baculovirus Gene
Involved
in
Late and
Very
Late Gene
Expression
YONGHONG LI,' A. LORENAPASSARELLI,1ANDLOIS K. MILLER'2*
Departmentsof Genetics' andEntomology,2*Universityof Georgia, Athens, Georgia 30602 Received 2 April 1993/Accepted 4 June 1993
Atrans-actinggenerequired for lateviralgeneexpression in transient expressionassays wasidentifiedin the genome ofAutographa
californica
nuclearpolyhedrosis
virus. A genomic library of A. californica nuclearpolybhedrosisvirusDNAlackingaclonespanning the regionfrom43to48mapunits(mu)wasunabletoactivate geneexpressionfromareporterplasmidwhenthereportergene wasdriven byabaculovirus lateor verylate promoter in transient expression assays. The genomic region responsible for activating reporter gene
expressionwasfurthermappedto43.4to45.2muof the viralgenome. Thenucleotidesequenceof thisregion
wasdetermined and showntocontainseveral smallopenreading frames (ORFs) andonemajor ORF, named
lef-3, for late expression factor 3. The kf-3 ORFwaspredictedtoencodeapolypeptideof 385 aminoacids and
withamolecularmassof44,529daltons. No homologtothelef-3 ORFwasfoundinexistingdata bases. Further
analysis showed that only the lef-3 ORF in the region from 43.4 to 45.2 mu was necessary for late gene activation in theassaysused.Thetemporal regulation oflef-3 transcriptionwasstudied by Northern (RNA) blot hybridization; lef-3 wasfound tobe anearly genethatwastranscribedprimarily as a 2.0-kb mRNA. Primerextensionanalysisand Si nucleaseprotectionassaysrevealed that lef-3 transcription initiated about 280
bpupstreamof the firstATG codon andterminatednear apolyadenylation signal, 130bpdownstream of the
lastcodonof thekf-3 ORF.
Although baculoviruses are now widelyused asvectors for theexpression ofa diversityofheterologousgenes, the
fundamental nature of the RNA polymerase(s) and other factorsinvolved intranscriptionfromlate andverylateviral promoters remains largely unknown. On the basis of the differentialsensitivityofearlyand late baculovirus
transcrip-tion to a-amanitin in nuclear run-on assays (10, 16), it is
generally thoughtthatanRNApolymerase switching mech-anism isinvolved in the transition between thesetwophases
oftranscription. This view is furthersupported by biochem-ical evidence of avirus-induced RNApolymerase activity appearing bytheonsetof the latephaseof infection(8, 43)
andbymolecular evidence that theprimarydeterminantsof late andverylatepromotersarelocatedatthetranscriptional initiation site, invariablyaTAAGsequence, andwithin the sequencesspecifyingtheuntranslated leaderregions (24, 25, 32, 41, 42).Thenatureof thenewpolymeraseand thenature of the factors thatrecognizeordistinguishtheseunusual late andverylatepromotersremaintobe determined.
During infection, the transition from the early to the late
transcriptional phasesisdependentonproteinsynthesisand DNAreplication.In thepresenceofaphidicolin, aninhibitor of both hostand viral DNApolymeraseactivities(23), early
butnotlateorverylategene transcription isobserved (34, 36).Amutantdefective in the viralproliferatingcellnuclear
antigen (pcna) homolog is delayed in late gene expression (6),andamutantdefective in the DNA helicase motif ofp143
is blocked in late gene expression (21). The basis for this
dependence of lategene expression on DNAreplication is
notknown,noris it known whetherthisdependence would also be observed in transientexpression assayswith naked viral andreporterDNAs. Transientexpressionofareporter
*Corresponding author.
gene under the control of late and very late promoters requirestheexpressionofmanyviralgenes(27, 28).For the
polyhedrin promoter, expression fromreporterplasmids is
independent of homologous repeat (hr) sequences (33), whichhave been assigned the functions of enhancers of early
geneexpression (11, 13)andoriginsof DNAreplication (17, 29). Expression from polyhedrin promoter-based reporter plasmidsis also knowntorespond authenticallytopromoter mutations when theplasmids are cotransfected with intact
Autographa californica nuclear polyhedrosis virus (AcM NPV) DNA in transientexpression assays(25, 33).
Werecently developedamethod foridentifyingAcMNPV genesthatarerequiredintransientassaysfortheexpression
ofareportergene underthe control of the latevp39 (major capsid protein gene)orverylatepolyhedringenepromoters (27). The method uses an overlapping set (library) of 12 clonesthatcoverstheentire AcMNPVgenomeand thatcan substitute for intactAcMNPV DNA intrans-activatingthe
expression ofreporter genes under late andvery late pro-moter control. Removal of some of the clones from the
libraryresults in thereductionortotalelimination of expres-sion fromthereporterplasmids. By removingonecloneata time and substituting smaller plasmid clones for thelarger clone, we found it possibleto identify preciselythe genes requiredfortrans-activating reportergene expression.
Thus far,we have identified four genes,ie-1, ie-n, lef-1, andlef-2,thatarenecessaryand sufficienttosubstitute for 2 of the 12library clones, coveringtheregionfrom 83.7to8.7 map units (mu) of the AcMNPV genome, in supporting
transient expression from late andvery late reporter
plas-mids (27, 28). ie-1 and ie-n were previously identified as
genes involved in the trans-activation of some early viral promoters intransient expression assays (3, 12, 14),sothe role ofthesetwogenesin the transientexpressionassayfor late gene expression may simply be toactivate earlygene
5260
Vol. 67, No. 9
on November 9, 2019 by guest
http://jvi.asm.org/
LATE GENE EXPRESSION FACTOR 3 OF AcMNPV 5261
expression. However,lef-1 and lef-2 have no apparent effect on expression from the early pcna (also known as eti) promoter,which serves as a control in our assays, and are therefore more likely to be involved in late gene expression as components of viral transcriptional or replication com-plexes. In this study, we used the transient expression assay forlate expression factor identification to define a third gene located between 43.4 and 45.2 mu of theACMNPVgenome. The sequence and transcriptional regulation of this gene
(lef-3)weredetermined.
MATERIALS AND METHODS
Cells and virus. Spodoptera
frugiperda
IPLB-SF21(SF-21) cells (39) were used as host cells for propagating AcM-NPV; the L-1 variant of AcMNPV (20) was used as the wild-type strain. The cells were maintained as described before (26) in TC100 medium (GIBCO BRL, Grand Island, N.Y.) supplemented with10% fetal bovine serum and 0.26% tryptosebroth.
Transient expression assays. SF-21 cells(2 x 106cells per 60-mmplate)werecotransfectedwitha reporterplasmidand either AcMNPV DNA or clonedsegmentsrepresentingall or
most of the viral genome. The reporter plasmids were
pETCAThr5 (27), pCAPCAT (37), andphcwt(33),in which the chloramphenicol acetyltransferase (CAT) gene was un-der thecontrolofearly
(eti),
late(capsidprotein genevp39),and very late (polyhedrin gene) viral promoters, respec-tively. For each transfection, 2 ,g of the reporter plasmid wascombined with0.5 ,g of viral DNA or 0.5 ,g of each of the clones from the AcMNPV library. When one of the cloned DNAswasomitted, 0.5 pgofplasmidoran equiva-lentamountofagel-purifiedDNAfragmentwassubstituted inthe transfection mixture. The total amount of DNA used in eachcotransfection was 8.0pg; salmon sperm DNA was used tobalance DNA concentrations whenever necessary. The transfection was performedby the calcium phosphate method (26). The cells were collected 24 h after transfec-tion for the early promoter and 48 h after transfection for both the late and the very late promoters andweredisrupted
bythreerepeatedfreeze(liquid nitrogen)-thaw (37°C)cycles. Cell debris was pelleted, and 1/10 of the supernatant was
used to assayfor CATactivity as described previously (9,
33).
DNA cloning and sequencing. The DNA fragment
corre-sponding to theAcMNPV genome from the EcoRI site at 43.4 mu to theSstI site at 45.4 mu (see Fig. 2A) was cloned into thepBluescriptSKvector(Stratagene,LaJolla,Calif.).
TheEcoRI-MluIfragmentof thisclone was then subcloned into the EcoRI and SmaI sites of thepBluescript KSvector
(Stratagene) byfillingin theM1uI site of the fragmentwith the large fragment of DNA polymerase I (Klenow) and
ligating the fragment to the vector. The resulting clone, pSDEM2,wasusedtogeneratetwo setsof nested deletions
with exonuclease III and mung bean nuclease(15) byuseof
the
KpnI
orSstIsitesasdeletionblockpoints,inaccordance with protocols recommended by Stratagene. pSDEM2 and the setsof deletion cloneswere sequencedfrom the T3 and T7 primer sites by the dideoxynucleotidechainterminationmethod (35)with asequencingkit (US Biochemical, Cleve-land, Ohio) and double-stranded DNA plasmid templates
(38). Sequenceswere compiled and analyzed byuse of the programsof Pustell and Kafatos(31)and the merger program ofIntelliGenetics. The deduced openreadingframes(ORFs)
within thesequencedregionwere comparedwith the
trans-lated sequences from GenBank (release 73) and U-EMBL (release 31) by use of the FASTA program (30) and the BLAST (1) network service at the National Center for Biotechnology Information.
RNA isolation. Total RNA was isolated from mock- or AcMNPV-infected SF-21 cells at various timespostinfection (p.i.) by the guanidinium isothiocyanate method(7).Avirus inoculum(10PFU percell) was allowed to adsorb to SF-21 monolayers (5 x 106 cells per 100-mm plate) at room temperature.The inoculum wasremovedand replaced with fresh medium after a 1-h adsorption period. The infected cells were then incubated at 27°C; the beginning of the incubation was considered zero time p.i. For inhibition of DNA synthesis, aphidicolin (5
,ug/ml)
was included in the medium added to 0 h. For blocking ofprotein synthesis,cycloheximide(100 ,ug/ml)wasaddedtothe medium 30 min
before inoculation.
Northern (RNA) blot
analysis.
Total RNA (10 ,ug) from mock- or AcMNPV-infected cellswas denatured by glyox-alation (22), electrophoretically separated in 1.0% agarose, and transferred to Zeta-Probeblottingmembranes (Bio-Rad Laboratories, Richmond, Calif.). The 1-kb DNA ladder(GIBCO BRL)usedasthe size markerwasalsoglyoxalated.
Blots were hybridized at 50°C to a strand-specific RNA
probe(hybridizationbuffer:50% formamide,1.5xSSPE[1x
SSPEis 180 mMNaCl, 10 mMNa2HPO4,1 mM
EDTA],
0.1 mg of tRNA perml, and 0.5% salmon sperm (DNA). For construction of a lef-3-specific probe, deletion plasmidpKHD31, generated for sequencing of the lef-3 gene, was
linearized with ClaI and transcribed in the presence of
[32P]UTP
with T3 RNApolymerase.Primerextension
analysis.
Asynthetic23-meroligonucle-otide (5'-GCTCATCAATTIACCACTAATCC-3') near the Nterminus oflef-3 was radiolabeled atthe 5' end with T4
polynucleotide kinase, annealedto10p,g of totalRNA,and extended withMoloney murine leukemiavirusreverse
tran-scriptase under the same conditions as those described earlier(6). Theextensionproductswere resolvedon an8% polyacrylamide-urea gel along with a DNA
sequencing
ladder generated from plasmid pSDEM2 by use of theradiolabeledoligonucleotide primerdescribed above. S1 nuclease protection analyses. The termini oflef-3
tran-scriptsweredeterminedby S1nucleaseprotection analyses (40) with 10,ug of total cell RNA per reaction. The DNA
fragment used as a probe to map the 5' end oflef-3 was
prepared bydigesting plasmid pKHD31 (one of the nested deletionplasmids generatedforsequencingof thelef-3 gene)
with ClaI, dephosphorylating the ends with calf intestinal
phosphatase, radiolabeling the 5' ends with T4
polynucle-otide kinase, and recleaving the plasmid with SstI, a site
within the vector. The resulting 984-bp fragment was
iso-lated andgelpurified.The DNAfragmentusedtomapthe 3' end oflef-3was madeby
digesting plasmid
pSDEM2withClaI, radiolabeling the 3' ends with the Klenow DNA
polymerase fragment, recleaving the plasmid with EcoRI,
and gel purifying the resulting 1,408-bp fragment. DNA-RNAhybridizationwascarriedoutwith80%formamide-40 mMpiperazine-N,N'-bis(2-ethanesulfonic acid)
(PIPES)-0.4
MNaCl-1mMEDTAat50°Covernightfor 5' mappingand 60%formamide-40mMPIPES-0.4 MNaCl-1 mM EDTAat 52°Covernightfor 3' mapping.Nucleotide sequence accession number. The sequence
re-ported in this paper was
deposited
in GenBank under the accession number L18873.VOL. 67,1993
on November 9, 2019 by guest
http://jvi.asm.org/
5262 LI ET AL. A.
C CKi
.
Gow
I II i.\. .
L1
I I I
B.
46
.0
*.e
*@@@@
i4
dill -V, W- w<
0 *@
FIG. 1. LocalizationtoPstH5 ofthegene(lef-3) requiredfor late
geneexpression of AcMNPV. (A) Restrictionmapof theAcMNPV
genomefrom 33.0to57.0muandkey restriction sitesintheregion:
H, HindIll;P,PstI;S, SmaI; Bg, BglII.mu aregivenunder the lines
representingthe AcMNPVgenome, and below themu are arrows
representingthepositions of knowngenes.The three overlapping genomicclones of thelibraryin thisregionof thegenome, PstHI, PstH5, and HC9,areindicatedby bars, with theopen area
repre-sentingtheregionknowntobe included in the clone and the shaded
areas representing additional regions that may be included. (B) Autoradiogram of CAT transient expression assays. SF-21 cells
were cotransfected with a reporter plasmid (pETCAThr5, pCAP CAT,orphcwt)and additional viral DNAasindicated above each lane: wt, wild-type AcMNPV DNA; L, entire library of cloned
overlapping AcMNPV DNA fragments; L*, ACMNPV genomic library lacking PstH5;0,reporter DNAonly. Cm, chloramphenicol;
AcCm,various forms ofacetylated chloramphenicol.
RESULTS
Identification of late expression factor gene 3 Ief-3. The reporter plasmids and clone library used in the transient assays to define AcMNPV genes involved in late gene expression were described previously (27). In brief, three differentreporterplasmidswereused. All three containeda
portion of the hr5 sequence and the CAT gene under the control ofaviral promoter. Theearlypromoterof theviral pcna (also known as eti) homolog (6) was the promoter driving CAT expression in reporter plasmid pETCAThr5 (27)and servedessentiallyas acontrolinthese experiments; it isdependent onie-1 expression and is stimulated byie-n
expression (28). The vp39 promoter, including 462 bp up-streamof thetranslationalstartcodon(36, 37), servedasthe
late promoter driving CAT expression in reporter plasmid pCAPCAT,and thepolyhedrinpromoterservedasthevery late promoter driving CAT expression in reporter plasmid phcwt (33).
CATactivitywasobtained for all three reporterplasmids
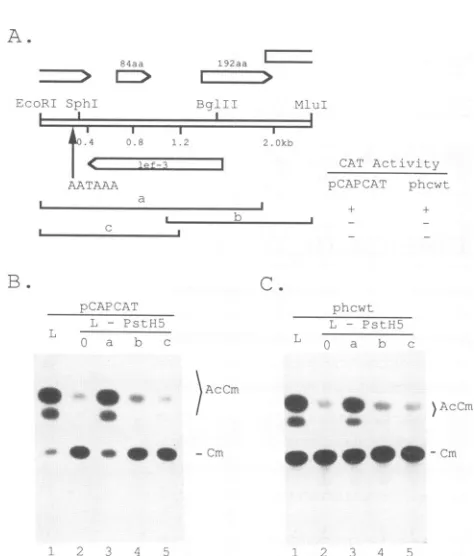
FIG. 2. Finemapping of lef-3 withinPstH5.(A) Diagram ofthe SmaI-D fragment(40.3 to49.4mu) clonedasplasmid pXmal7 (a)
and the derivedsubfragments (btoi).Therelative CATactivity for each transfection with the plasmidor asubfragment isgiven as a
plusor aminus, meaning that CATactivitywascomparabletothat of thepositiveorthenegative control, respectively. Restriction sites were SmaI(S),PstI(P),EcoRI(E), BglII (Bg), SstI (SI), SstII(SII), BamHI(B),SphI(Sp), MluI (M),andHpaI(H). (B) CATassaysfor cotransfections with pCAPCAT and the various DNAfragments, indicatedasletters above the lanes. The lanes labeledL - PstH5 (lanes 2to11) indicate that PstH5wasomittedfrom these transfec-tions. The lowercase letters above each lane (a to i) referto the plasmidorDNA fragmentshownin panelA and usedto replace PstH5.Lane 2(0)containsnoadditionalDNAsupplement. Lane 1 (L) contains the entire library of overlapping AcMNPV DNA clones.AcCm and Cmare asdefinedin thelegendtoFig. 1.
cotransfected with either wild-typeviral DNAor the
com-pletesetofoverlappingAcMNPV clones(Fig. 1B,lanes1, 2,
5, 6, 9, and10). Thus,aspreviously reported(27),theability
ofwild-typeAcMNPV DNA to trans-activate the reporter
plasmidscould bebasicallyreconstitutedbysubstitutingthe entire set of viral DNA fragments for intact viral DNA.
When the PstH5 clone,which extends from
approximately
37to50mu(Fig. 1A),wasomitted from thecotransfections,CATexpression from the earlypromoterwas not
substan-tially affected (Fig. 1B, compare lanes 2 and 4). However, the omission of the PstH5 clone from the cotransfections with the reporterplasmids carryingthe CAT gene under late and very late promoter control substantially reduced CAT
expression (Fig. 1B, compare lanes 6 and 8 and lanes 10 and
12). Thus,atleastonetrans-activatingfactor necessary for late and very late gene expression in these assays was
present exclusively in the PstH5 clone. The PstH5 clone
substantially overlaps two other clones of the genomic library,PstHlandHC9; theregion uniquetoPstH5extends fromapproximately43to48mu (Fig. 1A).
To delineate further the region containing the
PstH5-i
L-I-c
J. VIROL.
...
.I .I.
W&
46
0
00000000
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.51.285.69.359.2] [image:3.612.308.541.76.355.2]A.
EcoRI 43.4
L
BglII 44.6
M1uI 45.2
I
I 1 I 1I III 1 II II I
III I 111_ IIIIIIIIIIII111111 11 I1 TV
1.~~~
H 1 I IHI
l I 11 1111111111 III
II
I1iiil
u II1I
II- III VIAIliiiI II1le -3II I H il11 II III lifli I I
1 2 3
1'
2' 3'
B. 1
90 180 -270 _
360 a
450 T
540 ;
630
720
-M A T K
810 _
-L-R S L S G E S S G E P L I K R M A M A S S P K K I R E N Y K
900 _ IPTlA
R I S G K L M S K M T L S I D N E Y H Y T F R I M S D N K I
990 CAGAATATAC
Q E YY 1080AGTCAAATGATT
S Q M I
1170TTTGAAAATGA
F E N E 1260 GllX
V V Y K 1350AATATA1AkG
N I K G
1440CAACAAAT1I Q Q I F 1530GATATTCAAAT
A Y S N
G D
Q I
C%A
D G
N L
TCTGPC
S D
AATGGA
N G
TTCAA
L Q S Q S
XAAAATAC
N E Y G2CAATATTI
V N I
AAGaGAM
N D D
GAACAAT
D I N
TCAAACG,.
S N V
AATT1CA
N C T TICAAAGA
FKD F K D AAA,GA= K E C ATTGTAAAG
I V K
OCGAGTIGP D V
E V F TACATGAAT
Y M N
AAOGAAAG
N AK
GGAGAAGAGTGC3AI
M E E G K C Y 3AA-GAAA-IIGAAACTGCA
E M E I E T A
rACAAAmATCTACAAAAA
Y K F I Y K K
'mAAGGGAATICIGTC
V Q V E C S V
kAATTAAAAGCAATGAA
K Y L K D N E W N V V N S T
INTCAA=A
I N I S R S N
AATTArTAAATTTAAAATGTT
D I S L N Y V K Y K F T P M S D Y L T N K H I N S G L Y K V V F E
AATGAACAATAAATTCCAGAA
N A K T L I N L F K N
AA TCACATATAAAGTGT
N Q I F T I Y S I K C AAA
R I E L C E A K E S E
MTC ATGTAAATGMTGT
K H V A S Y N V N V L
1620
K S E L E E N D M G D N K F I V Q F K S D E L N I A D S D D
17100
C S T S S D L G K W N K S V F Y V N T N K K T E A D S L Q K
1800 _
L C A D F N Q I S M L L E D N L I K V T I Y V T V E N G E N
1890
H N M N V L G L L K Y D E D E N E Y K F L 1980I
2070_ 2160
2250
2340AITITTAATTC
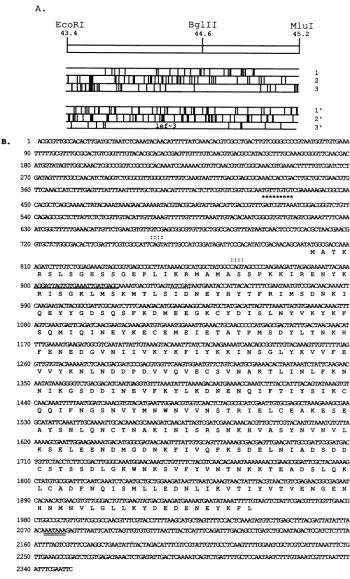
FIG. 3. Distributionof translational termination codons and sequence of theregionfrom 43.4to45.2muofAcMNPV,containinglef-3. (A)Restriction map of theregion sequenced. Stopcodons within all sixpossible readingframesareindicated below the mapbyvertical lines. Thelef-3ORF extends in thecounterclockwise direction.(B)Nucleotide sequence andpredictedamino acid sequence for the AcMNPVlef-3 ORF. The nucleotide sequence fromMluI (45.2 mu)toEcoRI(43.4mu)ispresentedwithnumberingtothe left of the first nucleotide in each line. Thepredicted amino acid sequence forlef-3is shown below the nucleotide sequence with the standardsingle-lettersymbolfor each amino acid. CAGT motifsnearthe N terminus oflef-3arenotedwithacolon(:)abovethe sequence. The actualtranscriptionalinitiationsite determinedempiricallyisindicatedwith asterisks above the sequence. The underlined sequence represents theregioncomplementarytothe primer sequence used for 5'-endmapping. The potential polyadenylation signal (AATAAA) is doublyunderlined. The ClaI site used in Northern blotanalysisandS1nuclease assaysis underlinedwith abrokenline.
5263
r
rG
m
11
I1 DG
r
CG 2 ::A rc
3
7IT,
II
DG NG
m 3G
IC .A
DNA
Al
3c rr
713 W
rz
Al
m
;A
LIO zN
rT
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.140.490.69.648.2]5264 LI ET AL.
A.
I I
I
B.
__-,'st
n b
(-_D
40
-tFIG. 4. Assignmentoftrans-activati Schematic representation of ORFsand
midsusedtoreplace PstH5 intheCATa
amino acid (aa) residues are represer
relative CAT activity for each cotrans
phcwt and various DNAs is given asa
assay withreporter plasmid pCAPCA'
done with pCAPCAT as areporter pla indicated above each lane. Lane 1inclu
(L), while lanes 2to5included clonesL
DNA was added in lane 2, while lane,
plasmidsathroughc, diagrammatically aspanelB,exceptthatCAT activitywa,
fections withreporterplasmidphcwt.A inthe legendtoFig. 1.
specific trans-activating function,
the PstH5 clone could be replace
plasmid, pXmal7, which contains
AcMNPV (40.3 to 49.4 mu) and
region unique to PstH5 (Fig. 1A
pXmal7 was equivalent to PstH5
trans-activate the vp39 promoter
remainder of thelibraryclones(Fig. 3).Twosubfragments of pXmal7 e: BglIIsiteat44.6mutothe SmaIsit
2A, rowsbandc)wereunabletofi PstH5 (Fig. 2B, lanes 4 and 5).
extending from the SstI site at40.3
2A, row d) or SstII (Fig. 2A, ro
trans-activate lategeneexpression
lanes 6 and 7). A PstI-SstI
subfral
(Fig. 2A, rowf)aswellas an EcoR
to 45.4mu) (Fig.2A, rowg) alsote
expression assays (Fig. 2B, lanes E these fragments was mapped with
zymes (Fig. 2A). Further subfragm testedfor their abilitytosubstituteI
(Fig. 2B, lanes 10 and 11), and th(
c
waslocalized to the EcoRI-MluI(43.4to45.2mu) segment of
AcMNPV(Fig.2A,rowh).Asmallfragmentextending from
the SphI site to the HpaI site, however, was unable to restore expression when substituted for the PstH5 clone
(Fig. 2B, lane 11).
Nucleotide sequenceofkf-3 and the flanking regions. The EcoRI-MluI
fragment
that was found to restore late geneexpression
tothe AcMNPVlibrary
in the absence ofPstH5was cloned and sequenced. The nucleotide sequence is -- presented in Fig. 3B. Computer analysis revealed a
major
--
- ORF in the counterclockwise direction and predicted apolypeptideproductof 385 amino acids and withamolecular
massof44,529daltons. Thepredictedinitiation codon is ina favorable context (AATATGG) for efficient translation in accordance with Kozak's rules (PuNNATGPu) (18) and
other
abundantly expressed
AcMNPV genes(26).
ThisORF wasdesignatedlef-3, for late expression factor 3. Thecodingregion of lef-3 has a high A+T content (63.8% A+T; 36.2%
G+C).
No apparentsignificant homology tolef-3wasfound 2 ' w; s whendata bases (GenBank release 73; U-EMBL release31)
were searched by use of the Genetics Computer Group* * w w v -
>-
FASTA program(30)
and the BLAST(1)
network serviceat the National Center for Biotechnology Information, norwere anysignificant known sequence motifs found within the
lef-3 gene product, with the exception of five potential
N-glycosylation sites(IntelliGeneticsKeybank program,
re-: lease9), although no apparent signal sequence was found.
ing
function to the ORF. (A)Identification
of the ORFwithin theEcoRI-MluI
fragmentAcMNPV portions of plas- that is necessary for late and very late gene expression. In issay. ORFs of more than 80 addition tolef-3, sequence data revealed two smaller ORFs nted by open arrows. The in the minimal trans-activating fragment (EcoRI-MluI) that sfection with pCAPCAT or overlapped lef-3 and could potentially encode polypeptides
iplus or a minus. (B) CAT of 84 and 192 amino acids, as depicted in Fig. 4A. To T. All cotransfections were determine whether one of thesmaller ORFs was responsible smid. Additional DNAs are for trans-activating the late
promoter,
wetested the ability of ided the entire clone library several deletionplasmidstosubstitute for PstH5 in transient ackingPstH5. No additional expression plasmid containintrantie
s 3 to 5 contained deletion expression assays. Only the plasmid containing the entire shown in panel A. (C) Same lef-3 ORFwas functionally similar to PstH5 in providing a
Ls
assayed following cotrans- trans-activating factor for the vp39 promoter(Fig. 4B). This LcCM and Cm are as defined plasmid (Fig. 4A, deletion clone a) was also able tosubsti-tute for PstH5 in trans-activating the very late polyhedrin
promoter (Fig. 4C). Thetwo smallerdeletion plasmids (Fig. 4A, clones b andc) were unable tofunctionally substitute for we first determined that PstH5 when used together in thecotransfections, indicating -d in these assays by a that the two smaller ORFsand/or partial ORFs do not act in the SmaI-D fragment of concert (data not shown). Thus, lef-3 is necessary for encompasses the entire trans-activation, although wecannotpreclude thepossibility and Fig. 2A). Plasmid that the84-amino-acid-encoding ORF (Fig. 4A) is necessary in terms of its ability to for late gene expression in addition to lef-3. The data are in the presence of the consistent with prior experiments showing that the disrup-.2B, compare lanes1 and tion of either the BglII or the SphI site abolished trans-xtending from the unique activating function(Fig. 2, lanes 4, 5, and11); these sites are
;e
at 40.3 or49.4 mu(Fig. within the translated region and are upstream of thepoly(A) unctionally substitute for signal, respectively, of lef-3 (Fig. 4A) (see also the 3' However, subfragments mapping data below).3mu to theBamHI (Fig. Northern (RNA) blot
analysis.
To examine the temporal)w e) site were able to regulation of lef-3 transcription, we performed a Northern in these assays (Fig. 2B, blotanalysis with alef-3-specific RNA probe. The uniformly gment (42.0 to 45.4 mu) labeled, strand-specific RNA probe extended from
nucle-I-SstI subfragment (43.4 otide (nt) 940 to about nt 1840 in anorientation opposite that sted positive in transient of lef-3 mRNA (Fig. 3B and Fig. 5A). As shown in Fig. SB, and 9). The smallest of a primaryvirus-specific transcript of approximately 2.0 kb
i various restriction en- was observed in infected cells at 3 h p.i. The level of this
ients of this region were 2.0-kb RNA was highest at 6 h p.i., declined slightly at 12 h for PstH5 in these assays p.i., anddeclineddramatically but remained detectable at24
e trans-activating region and 48 h p.i. When cells were treated with the DNA J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.52.289.64.342.2]VOL.67,1993L~~~~~ATEGEN-E EXPRESSION FACTOR 3 OF AcMNPV 5265
A
B
M 3 6 12 A C 24 48
.RI ScI
*-.4
4AT :
Clal
II,
:: gIiIrI
4.11
30-o
2.0- P:
1.6-1.-0
-C
D
e0M 501-= em 489 404 00
331 OM
242 *a
190- eAm 147- Of 111- 4fb
67- *oa
P M 3 6 12 A C 24 48
4= -4 ao
1444 eMM4 _ _. -~
736 a.
476 "
FIG. 5. Transcriptional mappingoflef-3. (A)Location andnatureofprobesusedin the Northern blotanalysisand the 5' and 3' Si nuclease
protection assays, shown in relationship to lef-3 (represented by an open arrow), and key restriction enzyme sites within the region sequenced. Theorientation of the RNAprobeused in the Northern blotanalysis is indicatedbythe arrowheadatthe 3' end. Thepositions
of radiolabels in 5' and 3' probesare indicatedby asterisks. Thepotential poly(A) site (AATAAA) is indicatedbyaverticalarrow. The
transcriptionalmappingdataforlef-3aresummarizedbythefilledarrowatthetop,representingthesize, location,and orientation of thelef-3 transcript.skvectorrepresentspBluescript sequences. (B)Northern blotanalysisoflef-3transcripts.Total RNAwasisolated from either
mock-infected cells(laneM)orAcMNPV-infected cellsat3, 6, 12, 24,and 48 hp.i. (lanes3, 6, 12, 24,and48,respectively) and 12 hp.i.in
thepresence ofaphidicolin (lane A)orcycloheximide (lane C).The RNAwasdenatured,electrophoretically separated, andtransferredtoa
nylon blottingmembrane. The membranewashybridizedwith thelef-3strand-specificRNAprobeindicated inpanelA.Thesizes of DNA
markers, inkilobases, are indicatedonthe left. (C)5'Mappingoflef-3transcripts bySi nucleaseprotection analyses.A984-bp fragment,
5' 32plabeled attheCiaI-generatedend as shown inpanelA,was hybridizedto RNAsamplesasdescribed forpanelBand subjectedto
digestionwith Si nuclease. The Si nuclease-resistantproductswereresolvedona4.0% polyacrylamide gelandvisualizedby autoradiog-raphy. Molecularweightmarkers (laneM),innucleotides,werederived fromMspl-digestedpUC19. LaneP,undigested probe.Themajor protected fragmentisindicatedbyanarrowhead.(D)3'Mappingoflef-3transcripts bySi nucleaseprotection analyses.Theprobeused for
hybridization to total RNAwas a 1,408-bpEcoRI-CiaI fragment radiolabeled with 32Pat theCiIA-generated end as shown in panel A.
Molecular weight markers were derived from Taql-digested pUC19. Lane P, undigested probe. The nuclease-resistant fragment is
indicatedbyanarrowhead.
synthesis inhibitor aphidicolin,which inhibits late and very latetranscriptionbecause ofadependenceonDNA
replica-tion, the level of the 2.0-kb RNA decreased only a small
degree (Fig. SB, compare lanes 12 and A), indicating the
early nature of lef-3 transcription. Treatment of infected cells with the protein biosynthesis inhibitor cycloheximide significantlyelevated the levels oflef-3transcripts (Fig. SB, compare lanes 12 and C), a result also consistent with the classification oflef-3 as an earlygene. Aminortranscript,
slightly largerthan 2.0kb,wasalsoseenatalltimesp.i.;this
transcript had a temporal pattern similar to that for the 2.0-kbtranscript.
Mapping of the 5' and 3' ends of kf-3 transcripts. To
determine theinitiationsite(s)forlef-3transcriptioninSF-21 cells, we performed a primer extension analysis and nuclease protection assays. Examination of the sequence
nearthe 5' end of thetranscribedregionoflef-3revealedtwo
CAGTmotifs that are frequently found near the
transcrip-P M 3 6 12 A C 24 48
2 0
. . a
VOL. 67,1993
'V
--A
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.89.528.74.451.2]5266 LI ET AL.
T C G A M 3 6 12 A C 2448
loo Fw
A
I
:5 f A|
T/
l
a I
ok- ..'
..- -w
vo:
ldk. 3
W--f4
*4-i@D "*ft
.i. IOW
FIG. 6. Primer extension analysis of the 5' end of the lef-3
mRNA. A 5'32P-labeled oligonucleotide(S'-GCTCATCAATITCA CACTAATCCr-3') was hybridized to total RNA from mock-in-fected cells(M) orfrom infected cells harvested atthe hours p.i.
indicated above thelanes, and the RNAwas reverse transcribed.
The lanes designated A and C between the 12- and 24-h lanes indicated thatthe RNAwasisolatedat12 hp.i. in thepresenceof
aphidicolin (A)orcycloheximide (C). The radiolabeled oligonucle-otidewasalsoused as aprimer with aDNAtemplatecontaining lef-3 andwas extended in thepresenceofdideoxyribonucleotides ddATP (A), ddTTJP (T), ddGTP (G), and ddCTP (C). The primer
extension productswereresolvedon an8.0%polyacrylamide gel.
Thetranscriptional startsite is shown ina region specified by the
nucleotidesequenceof the DNA strand complementarytothelef-3 mRNA.
tionalstart sites of early baculovirusgenes(2, 4, 5, 14, 19).
One CAGT is located 41 bp upstream of the translational start codon, while theother CAGT is located within theN terminus of the translated region and is followed, 57 bp
downstream,byanATG codon inafavorablecontext(AAA ATGA)fortranslationalinitiation (Fig. 3B).A23-ntprimer, corresponding to the sequence from nt 900to nt 922, was
thussynthesized and radiolabeled at the 5' end for the primer extensionanalysis. Three major distinct extension products, none of which mapped to a CAGT motif, were observed (Fig. 6). The longest and most abundant primer extension productcorresponded to a transcriptional start site about 280 bp upstreamof the first ATG codon oflef-3. The temporal pattern for this extension product was consistent with that
observedin Northernblots; the most obvious feature was an increase in the level of the product in the presence of
cycloheximide (Fig. 6, laneC). The shortestprimer exten-sion product would place the transcriptional start site at nt 750, 3 bp upstreamof the first CAGT motif. No extension products corresponding to the second CAGT motif were
observed in these primer extension reactions when gel electrophoresis was carried out to resolve shorter products (data notshown).
Si
nuclease protection analyses were also performed to mapthe 5'end of thelef-3mRNA. A 984-ntDNAfragmentradiolabeledattheClaI sitewasused as a probe (Fig.SA),
and afragment of about 400 to 450 nts was protected when this probe washybridized to total RNA and subjected toSi
nuclease digestion (Fig. SC). The transcriptional start site
deduced from the size of this protected fragment corre-sponds to that defined by the longest primer extension products. No protected fragment of a size corresponding to the shortestprimer extension product in the primer exten-sionanalysiswasobserved. The shortest extension product maytherefore represent a prematurely terminated product. Formapping of the 3' end of the lef-3 mRNA, a 1,408-bp
EcoRI-ClaI fragment 3' labeled at the CiaI-generated end was used as a probe in
Si
nuclease protection analyses oftotalRNA(Fig. SA). Asshown inFig. 5D, anSi
nuclease-resistantfragment of about 1,100 to 1,200 nts was found in
AcMNPV-infectedbut notmock-infected cells. The size of this fragment suggests that the AATAAA motifoccurring 130 bp downstream of the last codon oflef-3 is a
polyade-nylationsignal.
Taken together, the 5' and 3' mapping data showed that
lef-3was transcribed froma 1.6-kbregionof the AcMNPV genome.Thesizewasingoodagreementwith Northern blot data, which revealedamajor2.0-kbtranscript.
DISCUSSION
We havefound another gene ofAcMNPVthatisrequired
intransient assays for substantialexpressionfrom a reporter gene under late and very late promotercontrol; removal of this gene from a library of clones covering the AcMNPV genome results in a dramatic reduction in the level of
expression from late and very late gene reporter plasmids.
This gene, knownasthe lateexpressionfactor3 gene(lef-3),
is located inaregion of theAcMNPVgenome(43.4to45.2
mu) that has not been previously characterized in any baculovirus system. It is the fifth gene to be identified as
functional in late baculovirus gene expression in this sub-tractive assay system. Like lef-1 and lef-2, lef-3 affects
expression from the late vp39 and very late polyhedrin
promotersbut does notaffectexpressionfrom theearlypcna
(etl)
promoter, although we cannot currently eliminate thepossibilitythat it affectsexpressionfrom otherearly
promot-ers. It is most likely that lef-3 is involved in late gene
expression, but whether it acts at the level ofreplication, transcription, ortranslation hasnotbeendetermined. Since
these threelefgenesaffect late geneexpression, it islikely
that the effect thatweobservedonverylate geneexpression
is duetotheir activation of late genes.
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.86.259.69.495.2]LATE GENE EXPRESSION FACTOR 3 OF AcMNPV 5267 Analysesoflef-3 transcription showed thatlef-3 isanearly
genewithamajor transcript of 2.0 kbandaminortranscript
of slightlylarger than 2.0kb. The steady-statelevels of lef-3 transcripts are highest between 3 and 12 h p.i. and are
reduced dramatically by 24 h p.i. This temporal pattern is
similarto that observed forlef-1. Aswith other
AcAMNPV
earlygenes, transcription occurs in the presence of
cyclo-heximideand aphidicolin. Theelevation oflef-3 RNA levels
at 12 h p.i. in the presence of cycloheximide but not
aphidicolin suggests that the shutoff oflef-3 transcription requires an early gene product, although there are other
possible explanations (e.g., the presence of cycloheximide mayincrease thestability oflef-3 RNA).
Both primer extension and Si nuclease analyses of the 5'
end of the lef-3 transcript indicated that transcriptional initiation occurs approximately 280 nts upstream of the
translational initiation codon for the predicted 385-residue gene product. This result was surprising in two respects.
Firstly, a 280-nt leader is unusually long for an mRNA,
particularlyfor AcMNPV mRNAs;AcMNPV ORFs thatare
transcribed with long leaders generally have translated
ORFs in the leaders. NoATG-initiated ORFs were found
withinthelef-3leadersequence, nor werestable secondary
structures observed by computer analysis. Secondly, one
CAGTmotifwasfoundapproximately 60 bpupstreamofthe
translational initiation codon of the 385-codon ORF, and
another,with an upstreamTATAbox, was found
approxi-mately100bp downstreamof thepredicted initiationcodon
(CAGTmotifs havebeen foundatthestartsites ofsomebut
notall early baculovirus RNAs [2, 4, 5, 14, 19]). However,
both S1 nuclease and primer extension data indicated that
neitherof these CAGT motifswasusedas atranscriptional
initiation site. The site determined for lef-3 transcriptional
initiation mapped approximately 25 bp upstream of aCT
TATsequence,suggesting thatalatetranscriptmayinitiate
ontheopposite strand (i.e., fromATAAG) and overlap the
5'endof the lef-3 RNA. Whetherorhowthis late transcript
mightaffect lef-3 expression isnot yet known. The overlap shouldnotaffectour5'transcriptional mapping data because
the5' end mappedtosimilarlocations when RNA from cells
infectedinthepresenceof cycloheximide oraphidicolinwas
used in primer extension andS1 nuclease analyses. The
nature of the less abundant and slightly longer transcript
observedin the Northern blots wasnot specifically defined in thesestudies.
It is likely that lef-3 playsacentral role inlate baculovirus
gene expression. Only a few of the 12 clones of the AcM
NPVlibraryhavesuchastrongeffectonexpression fromthe
late andverylate promoters when removed from the
tran-sient expression assays (28). The four clones showing the
strongest effects uponremoval areIE15, BC5, PstH5, and
HC10. Wehave found that ie-1, ie-n, and lef-2 are able to substitute for IE15 (27), lef-1 is able to substitute for BC5
(28), and lef-3 is abletosubstitute for
PstH5
in the subtrac-tiveassaysystem(this paper). Wearecurrently determiningthefunctionalgenes mappingto the
HC10
region. ACKNOWLEDGMENTSWe thankS.Hilliard for technical assistance.
Thisworkwassupported inpartbyPublic HealthServicegrant
AI23719 from the National Institute of Allergy and Infectious Diseases.
REFERENCES
1. Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J.
Lipman. 1990. Basic localalignment searchtool. J. Mol. Biol. 215:403-410.
2.
Blissard,
G.
W.,
andG.
F. Rohrmann. 1989. Location,se-quence, transcriptional
mapping,
and temporal expression ofthe
gp64 envelope
glycoprotein
gene
of theOrgyia
pseudotsug-ata
multicapsid
nuclearpolyhedrosis
virus. Virology170:537-555.
3.
Carson,
D.D.,
L. A.Guarino,
and M. D. Summers. 1988.Functional
mapping
of an AcNPVimmediately earlygene whichaugments expression
of the IE-1 trans-activated 39K gene.Virology
162:444 451.4.
Carson,
D.D.,
M. D.Summers,
and L. A.Guarino.
1991.Molecular
analysis
of a baculovirusregulatory
gene. Virology182:279-286.
5.
Chisholm,
G.E.,
andD.J.
Henner. 1988. Multiple earlytran-scripts
andsplicing
of theAutographa
califomica
nuclearpolyhedrosis
virus IE-1
gene.
J.
Virol. 62:3193-3200.6.
Crawford,
A.M.,
and L. K.Miller.
1988. Characterizationof anearly gene
accelerating expression
of lategenesof thebaculo-virus
Autographa
califomica
nuclearpolyhedrosis
virus. J. Virol. 62:2773-2781.7.
Davis,
L.G.,
M. D.Dibner,
andJ.
F. Battey. 1986. Basicmethods in
molecularbiology.
Elsevier Science
Publishing,Inc., New York.
8.
Fuchs,
L.
Y.,
M.
S.Woods,
and R. F. Weaver. 1983. Viraltranscription
duringAutographa
califomica
nuclearpolyhedro-sis virus infection: a novel RNA
polymerase
inducedin infectedSpodoptera
frugiperda
cells. J. Virol. 48:641-646.9.
Gorman,
C.M.,
L. F.Moffat,
and B. H. Howard. 1982.Recombinant
genomes
whichexpress
chloramphenicol
acetyl-transferase in mammalian cells. Mol. Cell. Biol. 2:1044-1051.
10.
Grula,
M.
A.,
P. L.Buller,
and R. F. Weaver. 1981.a-Amanitin-resistant viral RNA
synthesis
in nuclei isolated from nuclearpolyhedrosis
virus-infected Heliothis zea larvae and Spodopterafrugiperda cells. J. Virol. 38:916-921.
11.
Guarino,
L.A.,
and W.Dong.
1991.Transientexpressionof anenhancer-binding protein
in insect cells transfected with theAutographa
califomica
nuclearpolyhedrosis
virus IE-1gene.J.Virol. 65:3676-3680.
12.
Guarino,
L.A.,
andM. D. Summers. 1986. Functionalmappingof a
trans-activating gene
required
forexpression
of abaculo-virus
delayed-early
gene.
J. Virol.
57:563-571.13.
Guarino,
L.A.,
and M. D. Summers. 1986. Interspersedhomol-ogous
DNA of
Autographa
califomica
nuclear polyhedrosisvirus enhances
delayed-early
gene
expression.
J. Virol.60:215-223.
14.
Guarino,
L.
A.,
and M. D. Summers. 1987. Nucleotide
sequenceand
temporal expression
of a baculovirus regulatory gene. J.Virol. 61:2091-2099.
15.
Henikoff,
S.
1984.
Unidirectional
digestion
with exonucleaseIIIcreates
targeted
breakpoints
for
DNAsequencing.
Gene28:351-359.
16.
Huh,
N.E.,
and R. F. Weaver. 1990. Identifying the RNApolymerases
that
synthesize
specific
transcripts of theAu-tographa
califomica
nuclear
polyhedrosis
virus.
J.Gen. Virol.71:195-202.
17.
Kool, M.,
P. M. M. M.
Van denBerg, J.
Tramper, R. W.Goldbach,
and
J.
M.
Valk
1993. Location of two putativeorigins
of DNAreplication
ofAutographa
califomica
nuclearpolyhedrosis virus. Virology192:94-101.
18.
Kozak,
M. 1986. Bifunctional
messenger
RNAs
in eukaryotes.Cell 47:481-483.
19.
Krappa, R.,
andD.Knebel-Morsdorf.
1991. Identificationof thevery
early
transcribed baculovirus
gene
PE-38.
J. Virol.65:805-812.
20.
Lee,
H.
H.,
and L. K. Miller. 1978.
Isolation of genotypicvariants
of
Autographa
califomica
nuclear polyhedrosis
virus.J. Virol. 27:754-767.
21.
Lu, A.,
and
E. B. Carstens. 1991. Nucleotide
sequenceof a geneessential for
viral DNA
replication
in the
baculovirusAu-tographa
califomica
nuclear
polyhedrosis
virus.
Virology181: 336-347.22.
McMaster,
G.
K.,
and G. G. Carmichael.
1977. Analysis ofsingle-
and double-stranded nucleic acids on polyacrylamide
and
agarose gels by
using
glyoxal
and acridine orange. Proc.VOL.67, 1993
on November 9, 2019 by guest
http://jvi.asm.org/
5268 LI ET AL.
Natl. Acad. Sci. USA74:4835-4838.
23. Miller, L. K., J. E. Jewell, and D. Browne. 1981. Baculovirus induction of a DNA polymerase. J. Virol. 40:305-308. 24. Morris, T., and L. K. Miller. Unpublished results.
25. Ooi, B. G., C. Rankin, and L. K. Miller. 1989. Downstream sequences augment transcription from the essential initiation site of abaculovirus polyhedrin gene. J. Mol. Biol. 210:721-736. 26. O'Reilly,D. R., L. K. Miller, and V. A. Luckow. 1992. Baculo-virusexpression vectors: a laboratory manual. W. H. Freeman &Co., NewYork.
27. Passarelli, A. L., and L. K. Miller. 1993. Three baculovirus genesinvolved in late and very late gene expression:ie-1, ie-n, andlef-2. J. Virol. 67:2149-2158.
28. Passarelli,A.L., and L. K.Miller. Identification and character-ization oflef-1, a baculovirus gene involved in late and very late geneexpression. J.Virol. 67:3481-3488.
29. Pearson,M.,R. Bjornson, G.Pearson, and G. Rohrmann. 1992. TheAutographacalifomica baculovirus genome: evidence for multiple replication origins. Science 257:1382-1384.
30. Pearson, W.R., and D. J. Lipman. 1988. Improved tools for biological sequence comparison. Proc. Natl. Acad. Sci. USA 85:2444-2448.
31. Pustell, J.,and F. C. Kafatos. 1984. A convenient and adaptable packageof computer programs forDNAand protein sequence management, analysis and homology determination. Nucleic AcidsRes.12:643-655.
32. Qin, J.,A.Liu, and R. F.Weaver. 1989. Studiesonthe control region of the plO gene of the Autographa califomica nuclear polyhedrosis virus. J. Gen. Virol. 70:1273-1279.
33. Rankin, C.,B. G.Ooi,and L. K.Miller. 1988. Eight base pairs encompassing the transcriptional start point are the major determinant for baculovirus polyhedrin geneexpression. Gene 70:39-50.
34. Rice,W.C.,and L. K. Miller.1986.Baculovirus transcription in the presence of inhibitors and in nonpermissive Drosophila
cells. VirusRes.6:155-172.
35. Sanger, F., S. Nicklen,and A. R. Coulson.1977.DNA sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA74:5463-5467.
36. Thiem, S. M.,and L. K.Miller.1989.Identification, sequence, andtranscriptional mapping of themajor capsid protein gene of the baculovirusAutographa califomica nuclear polyhedrosis virus. J. Virol.63:2008-2018.
37. Thiem, S. M., and L.K.Miller. 1990.Differential gene expres-sion mediatedby late, very late, and hybrid baculovirus pro-moters. Gene 91:87-94.
38. Toneguzzo, F., S.Glynn,E.Leci,S.Muolsness,and A.Hayday. 1988. Use ofachemically modifiedT7polymerase for manual and automatedsequencing of supercoiledDNA.BioTechniques 6:460-469.
39. Vaughn, J.L.,R. H.Goodwin, G. J.Tompkdns,and P. McCaw-ley. 1977. The establishment oftwo cell lines from theinsect Spodoptera fmugiperda (Lepidoptera: Noctuidae). In Vitro (Rockville) 13:213-217.
40. Weaver,R.F.,andC.Weissman. 1979.Mapping ofRNAbya modification of theBerk-Sharp procedure; the 5' termini of 15S beta-globin mRNA precursor and mature lOS beta-globin mRNA have identical map coordinates. Nucleic Acids Res. 7:1175-1193.
41. Weyer, U.,and R. D.Possee. 1988. Functional analysis of the plO gene 5' leader sequence of the Autographa califomica nuclearpolyhedrosis virus. Nucleic AcidsRes.16:3635-3654. 42. Weyer, U.,and R. D.Possee. 1989.Analysis of the promoter of
theAutographacalifornicanuclearpolyhedrosis virus plO gene. J. Gen. Virol. 70:203-208.
43. Yang,C. L.,D.A. Stetler,and R. F. Weaver.1991. Structural comparison of theAutographacalifornica nuclear polyhedrosis virus-induced RNA polymerase and the three nuclear RNA polymerasesfromthehost, Spodoptera
ftugiperda.
VirusRes. 20:251-264.J. VIROL.