Molecular Nature
of
Spontaneous Mutations in Mouse Lactate
Dehydrogenase-A Processed Pseudogenes
Kayoko
M.
Fukasawa,* Masako Tanimura; Ikuya Sakai," Farida
S.
Sharief," Fu-Zon Chung*
and Steven S.-L. Li*"
"Laboratory of Genetics, National Institute of Environmental Health Sciences, National Institutes of Health, Research Triangle Park, North Carolina 27709, and ?Center for Demographic and Population Genetics, University of Texas, Houston, Texas 77030
Manuscript received June 1 1 , 1986
Revised copy accepted September 15, 1986
ABSTRACT
The presence of at least ten mouse LDH-A pseudogenes was demonstrated in the genomic blot analysis, and four different processed pseudogenes have thus far been isolated and characterized. In this report, the nucleotide sequences of two different mouse lactate dehydrogenase-A processed pseudogenes, M 1 1 and M14, were determined and compared with the protein-coding sequences of the mouse and rat LDH-A functional genes. In the pseudogene M1 1 , the sequence of 64 nucleotides from codon no. 257 to 278 was tandemly duplicated. In the pseudogene M14, the sequence of 22
nucleotides from codon no. 68 to 75 was replaced by an inserted repetitive sequence of 242 nucleotides homologous to a mouse truncated R element. The pattern of nucleotide substitutions accumulated in mouse LDH-A pseudogenes M 1 1 and M 14, as well as that of pseudogene M 10 identified previously, was analyzed, and the substitution frequencies of the C or G at the CG dinucleotide were found to be high.
PROCESSED pseudogene is a DNA sequence
A
that is derived from the processed mRNA inter- mediate of a functional gene through retrovirus-like mechanism and that is either transcriptionally inactive because of the absence of promoter or incapable ofproducing a functional protein because of the pres- ence of frameshifts or nonsense mutations (LITTLE
1982; LUEDERS et al. 1982; NISHIOKA, LEDER a n d LEDER 1980; PROUDFOOT 1980; SHARP 1983). Thus, spontaneous mutations occurring in such pseudogenes a r e apparently not subject to functional (selective) constraints and may become randomly fixed in pop-
ulations. Processed pseudogenes have been found in
many mammalian gene families (LI 1983; WILDE
1986), and they provide a repertoire for investigating
the molecular nature of spontaneous mutations in mammals.
Lactate dehydrogenase (LDH; EC 1.1.27) catalyzes the interconversion of lactate and pyruvate with nic- otinamide adenine dinucleotide as coenzyme (EVERSE and KAPLAN 1973; HOLBROOK et al. 1975). In mam- mals, LDH isozymes A4 (muscle), B4 (heart) and C4
(testis) a r e encoded by three structural genes origi- nated from an ancestral gene during the course of evolution (MARKERT, SHAKELEE and WHITT 1975; LI
et al. 1983). T h e expression of these three LDH genes is developmentally regulated and tissue-specific. In order to understand the structural and evolutionary relationships of the LDH-A, B and C genes and to
' To whom correspondence should be addressed.
study the mechanism of gene regulation, we have undertaken the investigation of protein structure a n d
gene organization of mammalian LDH isozymes. We
have reported the primary structure of LDH-A4 iso- zymes from mouse and human as well as LDH-C4 isozymes from mouse a n d rat (LI et al. 1985; PAN et al. 1983; TSUJIBO, TIANO and LI 1985). We have also described the nucleotide sequences of LDH-A cDNA and processed pseudogenes, and the exon-intron or-
ganization of LDH-A functional genes from mouse and human (AKAI et al. 1985; LI et al. 1985; TSUJIBO,
TIANO a n d LI 1985). In this paper, we present the
nucleotide sequences of two newly-identified mouse LDH-A processed pseudogenes, the molecular nature
of spontaneous mutations in three mouse LDH-A pseudogenes, and the complexity of LDH-A gene- related sequences in the mouse genome.
MATERIALS AND METHODS
Molecular characterization of mouse LDH-A pseudo-
genes: The mouse (Mus musculus C57/B10) genomic library
was kindly provided by M. EDGELL (WEAVER et al. 1981),
and the genomic clones containing LDH-A gene-related sequences were isolated as previously described (BENTON
and DAVIS 1977; Rigby et al. 1977). The DNAs purified from clones XM 1 1 and XM 14 were analyzed by restriction mapping and Southern-blotting (SOUTHERN 1975; MANIA- TIS, FRITSCH and SAMBROOK 1982). The isolated DNA fragments containing LDH-A pseudogenes were further cleaved and subcloned into M13 mplO/mpll phages, and their nucleotide sequences were determined by the dideoxy chain termination method (MESSING, CREA and SEEBERG
198 1 ; SANGER, NICKLEN and COULSON 1977).
178 K. M. Fukasawa et al.
Analysis of genomic complexity: T h e high-hl, DNAs were isolated f r o m t h e liver of mouse (C57BLj6J and DBA/ 25) a n d were cleaved with restriction endonucleases EcoRI,
BamHI and HindIII. After electrophoresis on 0.75% aga-
rose gel, t h e D N A fragments were transferred t o a nitrocel- lulose filter a n d hybridized with t h e ”P-labeled mouse
LDH-A cDNA probe (AKAI et al. 1985).
RESULTS
Nucleotide sequences of mouse LDH-A pseudo- genes M l l and M14: Six mouse genomic clones con- taining LDH-A gene-related sequences were isolated from Charon 4A genomic library (WEAVER et al.
1981). Clone
AM15
possesses mouse LDH-A func- tional gene, and its exon-intron organization was de- scribed (FUKASAWA and LI 1986; LI et al. 1985). Clone AM 10 contains an LDH-A processed pseudogene and cloneAM
16 possesses a truncated LDH-A pseudo- gene; their nucleotide sequences were reported (AKAIet al. 1985; FUKASAWA et al. 1986). In this investiga- tion, clones
AM1
1 andAM12
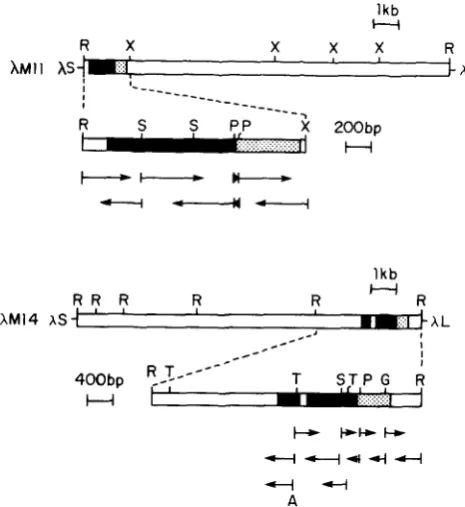
exhibited identical re- striction patterns, presumably containing the same LDH-A gene-related sequence; clone AM14 has a quite different restriction map. Figure 1 indicates the restriction endonuclease maps of the clonesAM
1 1(AM1
2) andAMI
4, along with their DNA sequencing strategies. T h e nucleotide sequences of approximately1.8 and 2.5 kb from clones
AM1
1 and AM14, respec- tively, are given in Figure2.
Sequence comparison of clonesAM
1 1 andAM
14 with the mouse LDH-A func- tional gene (clone AM 15) indicated that clonesAM
1 1and AM 14 contained different mouse LDH-A proc- essed pseudogenes lacking all seven introns present in the LDH-A functional gene. Both pseudogenes
M11
and
M
14 contain two direct repeats of 12 base pairs (bp) and 8-bp, respectively; one located just upstream from the5’
end and the other just downstream from the poly(A) tail. The nucleotide sequences upstream to their 5‘ direct repeat are very different from that of the promoter region of mouse LDH-A functional gene (FUKASAWA and LI 1986). Therefore, the LDH- A processed pseudogenesM
1 1 andM
14 are transcrip- tionally inactive because of the absence of a promoter. Furthermore, the “coding” sequences of both pseu- dogenes are incapable of producing LDH-A func- tional polypeptide chains as described below.Evolutionary relationships: In order to infer the evolutionary relationships among the mouse proc- essed pseudogenes and the mouse and rat functional genes, and also to infer the pattern of spontaneous mutations, we obtained an alignment of these se- quences as shown in Figure 3. From this alignment, the number of nucleotide differences between each sequence pair can be easily inferred, and the results are shown in Table 1. It is clear that all of the three mouse pseudogenes are more closely related to the mouse functional gene (M15) than to the rat func-
I k b H
R X X I x x R
A M I I AS- .I,
I.
AIR 200bp
- H
l k b
R R R R R - R
AM14 AS] I A L
---’ I I
I
W b k
- - - l ~ 4 + +
* c ( A
FIGURE 1 .-Restriction endonuclease maps of the mouse LDH- A processed pseudogenes MI 1 and M14 and their nucleotide se- quencing strategies. The overall maps of the mouse genomic clones AM11 and AM14 for restriction endonucleases R (EcoRI) and X ( X b a I ) were derived from single/double digestion as well as partial cleavage and Southern blot analyses with LDH-A cDNA probes from mouse and human. The “coding” sequences are indicated by solid blocks. The 5’ and 3’ untranslated regions are dotted. AS and AL indicate the short (10.5 kb) and long (20 kb) arm of Charon 4A
phage, respectively. The isolated EcoRI-XbaI fragment of 1.8 kb from clone AM1 1 was further cleaved by P (PstI) and S (SauSAI), and subcloned into M13 mplO or mpl1 phages. The isolatedEcoR1 fragment of 4.2 kb from clone AM14 was further cleaved by G
(BglII), P (PstI), S (SauSA), T (TagI), and/or A (AluI), and subcloned into M13 mplO or mpl1 phages. The ssDNA from M13 phages exhibiting positive hybridization with mouse and human LDH-A cDNA probes were purified and the nucleotide sequences of the inserted DNAs were determined by dideoxy chain termination methods.
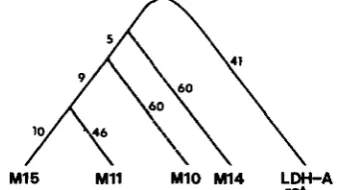
tional gene (MATRISIAN et al. 1985). Therefore, all of these pseudogenes appear to have arisen after the mouse-rat split. Further, the number of nucleotide differences between
M
15 andM
1 1,M
10 or M 14 are56, 79 and 84, respectively. For this reason, we pro- pose the branching order shown in Figure 4. Using the maximal parsimony principle, we have also in- ferred the number of nucleotide substitutions in each branch of the phylogenetic tree.
A
M11 gaattctacatcagggacccaaagc
ccaggagtttcctaaattaaactcaaacactaaagagtcataagttcaaaccaaggtaagccagactgggagttcataggactggcct a a a a a a t
Eco R I
*
G T G T C T G G A G C C A C T G T C G E C G A G C C C A A G C C A T G C T G C ~ C C C A T C G T G E A C T A G C G G T E T C A A A A G A T T C A A A G T C C A A
GATGGCAACCCTCAAGGACCAGCTGATTGTGA4TCTTCTTAAGGAAGAGCAGGCTCCCCAGGACAAGATTACAGTTGTTGGGGTTGGTGCTGTTGGCATG
GCTTGTGCCATCAGTATCTTA4TGAAGGGCTTGGCGGATGAGCTTGCCCTTGTTGACGTCATGG~GACAAACTCAAGGGCGAGATGATGGATCTCCAGC
ATGGCAGCCTCTTCCTTAAAACACC4AAAATTGTCTTCAGCAAAGACTACTGTGTAACTGCGAACGCCAAGCTGGTCATCATCACAGGGGTGTCTGTCAG
C AAGAGGGGG4GAGCCAACTCAACCTGGTCCAACGAAACATGAACATCTTCAAGTTCATCACTGCCAACATTGTGAAGCACAGTCCACACTTCGAGCTGC
TGATTGTCTCAAATCCAGTGGATATCTTGACCTACGTGGCTTGG~AATCAGTGGCTTTCCCAA4AGCGGAGTTATTGGAAGTGGTTGCAGTCTGGATTC
4GCTCGGTTCTGTTACCTfATGGAAG4AAGGCTGGGGGTTCACCTGAGCTGTCATGGCTGGGTCCTGGGAGAACATGGCAACTCCAGTGTGCCTGTGTGG
49TGGTGTGA4CGTTGCCGGTGTCTCCCTGAAGTCTCTGAACCCAGAACTGGGCACTGATGCAGAC~GGAACAGTGGAAGGAGGTTCAAAAGCAGGTGT
GGACAGTGCCTCCAAGGTGATCA4GCTGAGAGGTTACACATCCTGGGCCATTGGCCTCTCTGTGGCAGACTTGGCCGAGAGCATAATGAAGAACCTTAGG
CGGGTGC4TCCCATTTCCACC4TGATT4AGGGACTTGGTCGAGAGCATAATGAACCTTAGGTGGGTGC4TCCCATTTCCACCATGATTAAGGGTCTCTAT
GGA4TCAGTGATGACGTCTTCCTCAGTGTCCCATGTATCCTGGGACAAAATAGAATCTCAGATGTTGTGAAGGTGACTCTGACTCCCGAGGACGAGGCCC
GCCTGAAGA4GAGTGCAGACACCCTCTGGGGAATCGAGAAGGAGCTGCAGTTCTAAAGTCTTCTGAGTGTCCTAGCACTTCACTGTCCAGGCTGCAGCAG
GGTTTCTATGGAGACC4CACCCTTCTCTTCTGAACTGTGGTTAGTACAGTGGTTTTGAGGTGGTGTGGGGAAACATCTCACTCCCCACAGCTCTGTTGCC
AAGTGGTACTTGTGTAGTGGTAACCTGGTTAGTGTGAC4GTCCCGCTGTCTCCGAGACACACTGCCAACTGTGCAGGCTTCAATTACCCTGTGAGCCTGC
TGCATTGCTGCCCTGCGCACCCTCACCAAACATGCCTAGGCAACGAGTTCCCAGTTAGTCATAACCTGGCTCCAGTGTGTACGTCCATGATGCATATCTT
GTGCATGA4TGTTGTACAGGATATTTTATGTATTATATGTGTCTGTAGTGTGCATTGCAATGTTATGTGAGATGTAAAATCTGCATATGGAT~TGGAAC
t l
#257
X277U257 W277
#331
tA4GGACCCAAGTGTCATGCCAATGAAAACACCAAATAAACCTTG~CAGTGAAAAAAAAAAAAAGAGA a a a a a t t t c t a g a s i g n a l Poly (A) Xba I
B
H I a Eco R I
...
ctattcatgtctctatgaaaccaaaggaattgagatatcaagttcttcaaatttttt tgtttatgtttaaattaaatttaaaagcatactctcaactgagggggagggggaggaggagagggagagggagaggaggagacttttacccagcactccctcatqgtcaagaacagagacagatgagattctctqcatcttgggtcttgcaactgaggaaagccatcacgttccagcctcagagaaagtgaaaacaggc~
C G T C c A G C C i C 4 A A A G A T T E A A A G T C C A 4 ~ C C G C A A C C C ~ C A A G G A C C A ~ C T G A T T G T G ~ A T C T T C T T A ~ G G 4 A G A A C A ~ G T C C C C C A A ~ A C A A G A T T A ~ 4TTGTTGGGGTTGGTGCTGTTGTCATGGCTTGTGC TATCAGT4TCTTAATGAAGGACTT4GTGGATGAGCTTGCCCTTGTTGATGTTCTTGGAAAACAAG
CTAA4GGGAGAGATGATGTATCTCGAGCATGGA~tgggaggagattccctttgtcctatgaaggctctatggtccagtgtgggggaatggcaggg
caggagtcaggagtgggtgggttggtgagcaggggagggtggaggaaatggagggggtgtttttcggagggcaaatgaggaaagaggataacatttgaga
tgqaaataaagaaaotatccaataaaagcaaaaaacaaaacaaaacaaaacaaacaaacaaacaaaaaaccatc AAATTGTCTCCAGC4AAGACTATTG
TGT4ACTGCA4ACTCCA4GCT6GTCATTATCACACTGTTGGCCTGTC4GCATAGG~G4AAGCCGACTCAGTTTGGTCCACAGA4ACGCGAACACTTTAAG
TTCATCATTGCTAACATTGTGAAGTACCTACCACACTGCA4GCTGCTTATTGTCTAATTCCAGTGGATATCTTGACCTATATGGCTTGGAAA4TCAGCGG
CTTTCCCAAAATCCGAGTTATTGGAAGTGGTTGCAGTCTGGATTCAGCTCAGTTCTGTTACCTGATGGGAGAAAGGCTGGGGGTTCAGGCGATG4GCTGT
CACGGCT65GTCCTGGGAGAACATGGCGACTCCAGTGTGCCTGTGTGGAGTGGTGTGAACGTTGCCGGCATGTCCCTGAAGTCTCTGAACCCAGAATTGG
GCACTGATGCAGAC4AGGAGCAGTGGAAGGAGGTTCACAAGCAGGAGGTGGACAGTTCCTACG4GGTGATCAAGCTGAAACTGGGCCATTGGCCTTTCTG
T~GCAGACTTGTTCGAGAGC4TAATGAAGAACCTTAGGCGGGT~CATCCCATTTCCACC4TGATT~GGGTCTCTATGGAATCAATGATGATGTCTTCCT
C4GTGTCTCATGTATCCTGGGACAAAATGGAATCTCAGATGTTGTGAAGGTGACACTGACTCCTGAGGAAGAGGGCCGCTTGAAGAAGAGTGCAGATACA
CTCTGGGGAATCCA~AA4GAGTAGTTCTAGTCTTCCCAGTGTCCTAACACTTCACCGTCCAGGCTGCAGCAGGGTTTCTATGGAGACCACAATACTTTTC
ATCTGAGTGGTGGTTAGT4CAATTGTATTGAGATGGTCTGGGGGA4AAAAAAAGCCCTCCATTCCCACAGCTCTGCCCTGCTGCCA4GTGGTACTTGTGT
AGT6GTAACCTGGT4TTGTGAT4GTCCCACTGTCTCCAAGAC4CATTGCCAACTGTGTTGCAGGCCTTGCTGCACAACATCACCAAACTTGCCTAGGCCA
ATGAGTTCCCAGTTAGTCATAACCTGGCTCCAGTGTGTAAGTCCATTATGCATATCTTGTTCTACAGGAT4TTTTATGTATTCATATGTGTCTCTAGTAT
11
Y67
- - 9 1 5
Y331
AC4TTGTACTATGTGAAATGTAAGATCTGCATACGGATGACGGAACCAATCCACTCAAGTGTCATGCCAAGGAAAACACCAAATATACCTTGAACAGTGA
s i g n a l
4A44AAAATaa a qc ttacacttgtacataacagaaacaagcaagtcaagtaaagaaaaatattttggagatgagctttattatggaacagaaacta gccttaattttaccacttatttatttatttatttatttattttttctatgacaatttcatgcaggtatataatgtattagggtcagtgtaagcctctatt
actc~ttcttggctcctatcattgtgaaggaataactttttgttccaaaatgtatctctcctttatcgtctttttgttttgttatcttcacctttgcttt
taaatgactttctgagttacttagggttgtttgtgtqatcacggatgggatcttatttactggacaacaggcagctcactgtgcctatgctactgaaga~
a a t g a c t c t c c a t c t c c a g c a a c c g g t a a c t a t c a a t a g c t c t t c a g g g a g t a g t g g t g c c t t a t a a c a t c t t c t t c a t t c a t g t t g g g a c t t t g a t g g a
t c c a a t c t t a t g c a g t c c a a a t g c a g a c a a
...
E c o ~ I P o l y ( A )+
-
101- 1
100
200 300 400
500
600
700
800
900 1000 1100 1200
1300
1400
1500
1600
-
201-
in1- 1 100
200
300 400
500
600
700
800
900 1000 1100
1200 1300
1400 1500 1600
1700
1600 1900 2000
2100
2200
FIGURE 2.-Nucleotide sequences of the mouse LDH-A pseudogenes M I 1 and M I 4 and their flanking regions. The nucleotide sequences flanked by direct repeats of the mouse LDH-A pseudogenes M 1 1 and M 1 4 are numbered from the 5 ’ to 3’ end, and the number of the last nucleotide in each row is given at the right-hand side. The flanking sequences of LDH-A pseudogenes M 1 1 and M 1 4 , as well as the insertion sequence of 242 nucleotides in pseudogene M 1 4 , are given in lower case, and the direct repeats are denoted by arrows. The numbers (#) of some codons, the presumptive polyadenylation signal and poly(A) tail are also indicated.
180
M 1 5
M I 0
Mll
M14
r a t
M 1 5
Mlll M l 1 M l J
r a t
Ml5 MlO M l l
M I 4
r a t
M15 M1O M l l M14
r a t
M15 M10 M 1 1 M14
r a t
M15
M l O
M 1 1 M14
r a t
M 1 5 M l O M l l M14
r a t
MI5
M i l l
MI 1
M14
r a t
M l 5
M l l l
Ml I
Ml J
r a t
K. M. Fukasawa et al.
1 10 20 30
Het A 1 ~ T h r L ~ u l y s A s p t i l n L e i i I 1 eVa I AsnLeiiLeuLysGluGluGInAl aProGl nAsnLys I IeThrVal Val GlyVal GlyAl aVal tilyMetAl aCysAl d I 1 eSer I 1 eLeu
4 l ~ l ; C A A C C C l C A A l ~ ~ A ~ C 4 ~ C l ~ A T l t i l t i A A T C T l C ~ l A A G G A A G A G C A G G C T C C C C A G A A C A A G A T T A C A G T T G T T t i G t i t i T T t i G T G C T t i l T G t i C A T t i G C T T t i T t i C C A T C A G T A T C I T A
...
.CA....
TCT.....
.G.. T.....
...
G...C C -
...
.C.....
A... .TC....
.A.....
.A.-.......
.T......
.T.....
...
G...
A....
TC...
Met LjsAsgLeuAl aAspGluLeuAlaLeuVa1 AspValMet GluAspLysLeuLysGlyGl UMetMetAspLeuGlnHi sGly SerLeuPheLeuLysThrProLys I leva1 SerSer
Al(;AAGGACTTtiGCL;GATGAGCTTGCCCTTGTTGACGTCATtiGAAGACAAACTCAAGGGCGAGATGATGGATCTCCAGCATGGCAGCCT~ T KCTTAAAACACCAAAAATTtiTCTNAtiC
...
A...
A...
T...
G..A...
T...
T...
G...
...
G...T.......
.A.T.....
T. .TT...
..A...
.G. .A...
..A.....
.T....
.G.....
..A+++++++++++++++++++,+,,...
...
1...
T...
A...
T..G..A...
A...
T...
T...
G...
80 90 100 110
...
T..G..
G.A...A...G...
C...
A...
G...
A...
A...
T.G...
C....
..G.....
..C...
..A.-.. .T.T.T.....
.AA.....
A.....
.A.....
...
.T.....
A.....
.ACT.TT... .T......
.T-.-. .A. .A....
.A...
.GTT....
.CA.....
.C.....
.T.....
...
T..TA....
G...
A...
A...
..TT...
120 130 1411 1 qn
...
4 I1 50 60 70
LysAspTyrCysVal ThrAl aAsnSerLysLeuVal I1 e I 1 eThrAl aGlyAl aArgGl nG1 nG1 uGlyGl uSerArgLeuAsnLeuVa1 G1 nArgAsnVa1 Asn I1 ePheLysPhe I 1 e
MAGACTACTtiTGTAACTGCGAACTCCAAGCTGGTCATTATCACCGCGGGGGCCCGTCAGCAAGAGGGGGAGAGCCGGCTCAACCTGGTCCAGCGAAACGTGAACATCTTCAAGTTCATC
_-.
I 1 eProAsn I leva1 LysTyrSerProHi sCysLysLeuLeu 11 eVal SerAsnProVal Asp1 1 eLeuThrTyr Val A I aTrpLys I 1 eSeTGlyPheProLysAsnAryVa1 I1 eGly
ATTCCCAACATTGTCAAGTACAGTCCACACTGCAAGCTGCTGATCGTCTCCAATCCAGTGGATATCTTGACCTACGTGGCTTtiGAAAATCAGTGGCTTTCCCAAAAACCGAtiTAATTtiGA
...
A...TC...G...
.tiT...
T..-..A...
G..A...T...
.C.G.....
.G.. .C.....
.T.G.....
.T....
.A.....
.G.G...
.T......
..
.G.T.....
.G.....
.CTA.....
.T. .T...
.-A.T.....
TA.....
.C.....
.T.....
.T......
...
A. .TG...
.G. .A.....
.G....
.A....
.C.....
A. .C.....
.G....
.C....
.C.....
A...
.T......
160 170 180 190
SerGlyCysAsnLeuAspSerAl aArgPheArgTyrLeuMetGlyGluArgLeuGlyVa1 H i sAl aLeuSerCysHi sGlyTrpVal LeuGlyGl uHi sGlyAspSerSerVa1 ProVal
AGTGGTTGCAATCTGGATTCAGCGCGGTTCCGTTACCTGATGGGAGAGAGGCTGGGGGTTCACGCGCTGAGCTGTCACGGCTGGGTCCTGGGAGAACATGGCGACTCCAGTGTGCCTGT ti
...
.C.....
.T.CA.. .TA.....
.A.....
.CA.....
...
G...
T...
T...
A...A...
CT.---........
T.......
A...
...
G...
T.A....
T...
A...
G...A...
...
.G. .T.....
..A.....
.A....
.TC.A.....
.G.....
.G.....
200 210 220 230
TrpSerGly Val AsnVal A1 aGly Val SerLeuLysSerLeuAsnProGl uLeuGly ThrAspAl aAspLysGl uG1 nTrpLysG1 uVal H i sLysGl nVal Val AspSerAl aTyrGl U
TGGAGTGGTGTGAATGTTGCCGGCGTCTCCCTGAAGTC TCTTMCCCAGAACTGGGCACTGACGCAGACAAGGAGCAGTGGAAGGAGGTTCACAAtiCAGGTGGTGGACAGTtiCCTACGAG
...
A.....
.CA......
.T.....
.G.....
.T.. .C.....
.G......
A. .TAC.....
A...C.....
.T...G......
T...A......
A...
C.A.....
C...
A.G...
G...
T.......
T...
A.......
T...
...
.C. .C.....
..G....
.GC.G.....
.G. .T.....
.T..G.....
.T.....
A....
.A240 250 260 270
Val 11 eLysLeuLysGlyTyrThrSrTrpAla IleGlyLeuSerVal A1 aAspLeuA1 aG1 user 11 eMet LysAsnLeuArgArgVal H i sPro I 1 eSerThrMet I leLysGl yLeu
GTGATCAAGCTGAAAGGTTACACATCCTGGGCCATTGGCCTCTCTGTGGCAGACTTGGCTGAGAGCATAATGAAGAACCTTAGGCGGGTGCATCCCATTTCCACCATGATTAAGGGTCTC
AG
...
G...
C...G...
AA.T...
C...
A...
T...
...
.G.....
.TC.....
.T.....
...
...
.T.....
.TTC....
...
c c280 290 300 310
TyrGiy I 1 eAsnGl uAspVal PheLeuSerVal ProCys IleLeuGlyGl nAsnGlyI 1 eSerAspVal Val LysVal ThrLeuThrProGl uG1 uG1 u A I aArgLeuLysLysSerAl a TATGGAATCAATGAGGATGTC TTCCTCAGTGTCCCATG1ATCCTGGGACAAAATGGAATCTCGGATGTTI;TGAAGGTGACACTGACTCCTGAGGAAGAtiGCCCGCCTtiAAGAAGAGCGCA
...
T...
G...
A..G...
C...
C...
T.A....
G...T..C...
A...
A...
T...
C...
C...
T......
...
...
T...
T...
A...
G....
T...
T......
G...
C...
A...
C..G...
T...3 20 331
AspThrLeuTrpGly I leGlnLysGluLeuG1 nPheTer
:;AC4CCClCTtiGGGAATCCAGAAGGAGClGCAGTTCrM
....
C......
G.....
T..A......
A...-.--...
G. . T
...
FIGURE 3.-Sequence comparison of the mouse LDH-A processed pseudogene M10, M11 and M 1 4 with the protein-coding sequence of the mouse (M 15) and rat LDH-A functional genes. Different nucleotides are given, while identical nucleotides are indicated by dots; deletions are denoted by hyphens (-). The plus sign (+) indicates the sequence of 22 nucleotides of codon no. 68 to 75, which was replaced by an inserted repetitive sequence of 242 nucleotides. The deduced amino acid sequence of mouse LDH-A functional gene is given above its nucleotide sequence.
a deletion of 1 1 nucleotides, 8 single-base deletions, and a replacement of 22 nucleotides from codon no. 68 to 75 by a sequence of 247 nucleotides flanked by two perfect direct repeats of 5-bp. As indicated in Figure 5 , this inserted repetitive sequence (IRS) was found to be homologous to the 3’ end of the 475
Mouse LDH-A Pseudogenes
TABLE 1
Numbers of nucleotide differences between LDH-A sequences
M 1 5 M11 M I 0 M14
Mouse M 15 -
Mouse M 11 56
-
Mouse M10 79 83
Mouse M 14 84 96 103
Rat LDH-A 65 93 96 93
-
-
previously identified genomic clone AM 10 (FUKASAWA
et al. 1986), a mouse LDH-A processed pseudogene and a B1 repetitive element are flanked by perfect direct repeats of IO-bp, and this B1 sequence starts at 14 nucleotides 3’ to the presumptive polyadenylation signal (PROUDFOOT and BROWNLEE 1976) of the pseu- dogene. T h e “coding” sequence of pseudogene M 10 possesses a deletion of four nucleotides and two single- base deletions. In the genomic clone AM16 (AKAI et
al. 1985), the LDH-A processed pseudogene con- tained a truncated DNA sequence from codon No. 266 to the poly(A) tail of mRNA (data not shown).
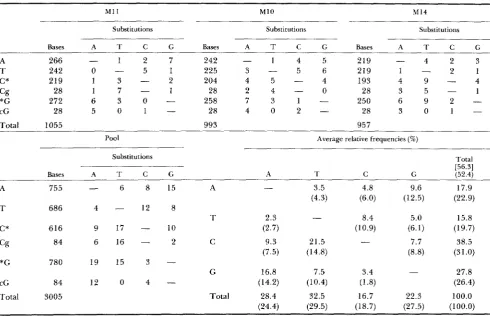
We have inferred the pattern of nucleotide substi- tutions in the pseudogenes, following the procedure of GOJOBORI, LI and GRAUR (1982). Using the phylo- genetic tree shown in Figure 4, we first inferred the ancestral sequence at each branch node and then inferred the direction of nucleotide substitutions in each pseudogene; deletions and insertions were ex- cluded from comparison. T h e results are shown in Table 2; the relative substitution frequencies were calculated according to the procedure of GOJOBORI, LI and GRAUR (1 982).
T h e three pseudogenes show very similar patterns of substitutions. For example, the frequencies of the two transitions C + T and G -+ A are the highest and the substitutions CG + CA and CG + T G are very high. Therefore, the data from the three pseudogenes are pooled together and used to calculate the average relative substitution frequencies shown in the last ma- trix in Table 2. T h e substitution pattern in these three pseudogenes is quite similar to that of the study by LI, Wu and Luo (1984). First, the proportion of transitions is 56.3% in the three LDH-A pseudogenes
M15 M l l M10 M14 LDH-A
rat
FIGURE 4.-A phylogenetic tree for the mouse LDH-A gene (M 15), mouse pseudogenes M 1 1, M 10 and M 14, and the rat LDH- A gene (MATRISIAN et al. 1985). The branching order is inferred from the distance matrix in Table 1.
M14 14s R consensus
M14 IRS R consensus
M14 IRS R consensus
M14 IRS R consensus
M14 I R S
----,
CATCAGTGGGAGGAGATTCCCTTTGTCCTATGAAGGCTCTATGGTCCAGT
***** ********** ***** * * ** * **** *****
CATCACTGGGAAGAGAGGCCCTTGGACTTGTAAACTTTATATGCCCCAGT
GTGGGGGAATGGCAGGG-CAGGAGTCAGGAGTGGGTGGGTTGGTGAGCA~~ ACAGGGGAACGCCAGGGCCAAAAAGTGGGAGTGGGTGGGTAGGGGAGTNGG
**a*** * ***** ** * ************* ** ***
GGAGGGTGGAGGAAATGGAGGGGGTGTTTTTCGGAGGGCAAATGAGGAAA GCAGGG-GGAGGGTAT---GGGGGACTTTT---
**** ***** ** ***** ****
GAGGATAACATTTGAGATGGAAATAAAGAAAATATCCAATAAAAGCAAAA
-GGGATAGCATTTGAAATGTAAATGAGGAAAATATCTAATAAAAA
***** ******* *** **** * ********* *******
-
AACAAAACAAAACAAAACAAACAAACAAACAAAAAACCATCA
50
100
150
200
242
FIGURE 5.-Comparison of mouse inserted repetitive sequence present in pseudogene M14 with the consensus sequence of mouse R repetitive elements. The nucleotide sequence of the inserted repetitive sequence (IRS) present in mouse LDH-A pseudogene M14 is compared with the 3’ end of consensus sequence of mouse R repeat family described previously (GEBHARD et al. 1982). The identical nucleotides are indicated by the asterisk (*) and deletions are denoted by dashes. N means A, G, C or T. The perfect 5-bp direct repeats flanking the IRS are indicated by arrows.
and 59.2% in the study of LI, WU and LUO (1984). T h e proportion of transitions excluding CpG dinucle- otides is 52.4% in the LDH-A pseudogenes and 54.4% in LI, Wu and LUO (1984). Second, the relative sub- stitution frequencies at C or G is high; it is 66.3% in the LDH-A pseudogenes and 64.3% in LI, Wu and LUO (1984). Third, for each type of nucleotide the rate of transition is the highest while the rate of transversion between complementary bases is the low- est. For example, for the G nucleotide, the rate of the transition G + A is 16.8% in the LDH-A pseudogenes and 16.0% in LI, Wu and LUO (1984); the rate of the non-complementary transversion G + T is 7.5% in the LDH-A pseudogenes and 7.0% in LI, Wu and LUO (1 984); the rate of the complementary transver- sion G + C is 3.4% in the LDH-A pseudogenes and 5.5% in LI, Wu and Luo (1984).
182 K. M. Fukasawa et al. TABLE 2
Frequencies and relative frequencies of nucleotide substitutions in mouse LDH-A processed pseudogenes
M11 M 1 0 M 1 4
Substitutions Substitutions Substitutions
A T C G Bases A T C G Bases A T C G
Bases
A 266
-
1 2 7 242-
1 4 5 219 - 4 2 32 1
T 242 0 - 5 1 225 3 - 5 6 219 1
-C* 219 1 3 - 2 204 4 5 - 4 193 4 9 - 4
28 1 7 - 1 28 2 4 - 0 28 3 5 - 1
6 9 2
-*G 272 6 3 0 - 258 7 3 1 - 250
cG 28 5 0 1 - 28 4 0 2 - 28 3 0 1
-Total 1055 993 957
c g
Pool
Substitutions
A
T
C*
c g
*C
CG Total
Bases A T C G
755
-
6 8 15686 4 - 12 8
616 9 17 - 10
84 6 16 - 2
780 19 15 3 -
84 12 0 4
-3005
Average relative frequencies (%)
Total
[56.3]
A T C G (52.4)
T 2.3
(2.7)
(7.5)
C 9.3
G 16.8
(14.2)
Total 28.4
(24.4)
3.5 (4.3)
-
21.5 (14.8)
7.5 (10.4)
32.5 (29.5)
4.8 (6.0)
(10.9) 8.4
-
3.4 (1.8) 16.7 (18.7)
9.6 17.9
(12.5) (22.9)
5.0 15.8
(6.1) (1 9.7)
7.7 38.5
(8.8) (31.0)
- 27.8
(26.4)
22.3 100.0
(27.5) (100.0)
~~
The substitutions are from the nucleotides in the left column to the nucleotides in the row. The C* indicates the C not followed by G and the *G indicates the G not preceded by C. [
1:
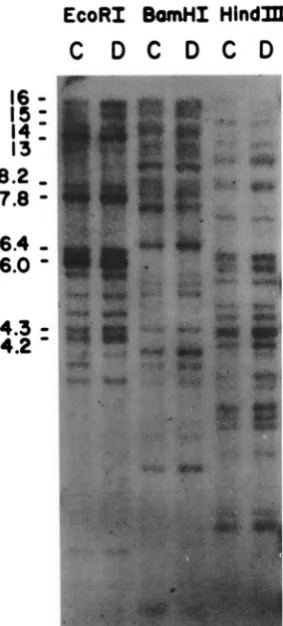
proportion of transitions; ( ): excluding CpG.kb fragment of clone M 16 probably corresponds to a weak hybridization band located between those of mouse LDH-A pseudogene M11 (14 kb) and func- tional gene (1 3 kb). T h e other strong hybridization bands of EcoRI 16, 15, 8.2, 6.4, 6.0 and 4.3 kb are probably derived from other LDH-A pseudogenes whose genomic clones have not yet been isolated. In short, at least ten different LDH-A pseudogenes are present in mouse genome.
DISCUSSION
The analysis of nucleotide sequences of mouse LDH-A processed pseudogenes has revealed various types of DNA alterations including base substitutions, deletions, tandem duplication and insertion of repet-
itive elements. T h e rate and the pattern of nucleotide substitutions in mammalian processed pseudogenes have been extensively analyzed and were elegantly used to illustrate the molecular evolution of mamma- lian genes and pseudogenes (GOJOBORI, LI and GRAUR 1982; Li 1983; Li, Gojobori and NEI 1981; LI, WU and LUO 1984; Wu et aE. 1986). The relatively higher rate of base substitutions at CG dinucleotide was
found in mouse LDH-A pseudogenes as well as other mammalian pseudogenes (GOJOBORI, LI and GRAUR 1982). The high frequency of polymorphism at MspI and Tag1 restriction sites containing the CG dinucle- otide in human genomic DNA (BARKER, SCHAFER and WHITE 1984) further supports the proposal that the methylated cytosine residue is a mutational “hotspot” in mammalian DNA. The “coding” sequence of pseu- dogene M 1 1 contains a tandem duplication, while the pseudogene M 16 possesses only a truncated “coding” sequence and 3 untranslated region. T h e pseudogene M 10 was followed by a B 1 repetitive element (KRAYEV
et al. 1980, 1982) at its poly(A) addition site, and the “coding” sequence of the pseudogene M14 was in- serted by a truncated R repetitive element (GEBHARD
Mouse Pseudogenes
least 10 mouse LDH-A pseudogenes in addition to the LDH-A functional gene. T h e chromosomal locations of these mouse LDH-A pseudogenes remain to be determined, while the mouse LDH-A functional gene has been mapped genetically to chromosome No.
7.
It should be pointed out that the weak hybridization bands of mouse genomic blot probed with mouse LDH-A cDNA may represent LDH-B/LDH-C gene- related sequences whose genomic clones remain to be isolated and characterized. As described previously (CHUNG et al. 1985; TSUJIBO, TIANO and LI 1985),
the human genome contains multiple LDH-A gene- related sequences, and several processed pseudogenes have been isolated and characterized. T h e presence of multiple pseudogenes has also been reported for a number of other mammalian gene families such as cy- tubulins (LEMISCHKA and SHARP 1982), cytochrome t
(Wu et al. 1986) and glyceraldehyde 3-phosphate de- hydrogenase (BENHAM, HODGKINSON and DAVIES
1984; HANAUER and MANDEL 1984; PIECHACZK e t al.
1984;
Tso
et al. 1985).EcoRI
BomHI
HlndPfC D C D C D
t
FIGURE 6.-Genomic blot analysis of mouse LDH-A gene-re-
lated sequences. T h e genomic DNAs were cleaved by restriction
endonucleases EcoRl, BamHl or Hind111 and were electrophoresed
on 0.75% agarose gel. T h e DNA fragments of C57BL/6J (C) and DBA/2J (D) mice were Southern yansferred to nitrocellulose filter and hybridized with mouse LDH-A cDNA probe in 6 X SSC at 63" overnight and washed twice with 1 X SSC a t 42" for 30 min. T h e estimated sizes (kb) of the EcoRI fragments exhibiting strong hy- bridization are indicated.
(KLECKNER 198 I). Similarly, the interspersed repeti- tive elements in mammals might constitute a signifi- cant mutation liability because of the possible trans- position and rearrangement of the functional genes flanked by repetitive sequences.
I t has been proposed that the intronless pseudo- genes were formed by the integration into germline DNA of reverse-transcribed processed mRNA (SHARP
1983) and that formation of processed pseudogenes may share common pathways with retroviruses and transposable elements. In Drosophila, processed pseu- dogenes have not yet been reported, but several types of transposable elements have been isolated and char- acterized (SPRADLING and RUBIN 1981). T h e abun- dance of mammalian processed pseudogenes might be directly associated with an existing mechanism similar to the replication of retroviruses in mammalian germ- lines. Thus, the "housekeeping" genes actively ex- pressed in germline cells may be found to have more copies of processed pseudogenes than those expressed mainly in differentiated somatic cells. There are at
We thank M. H. EDCELL for the mouse genomic library, B. H. JUDD for his interest and support of this investigation, W. QUAT-
TLEBAUM for computer analysis of nucleotide sequences, F. JOHN-
SON, R. VOELKER, T. KUNKEL, J. DRAKE, C. LANGLEY, B. H. JUDD
and W.-H. LI for helpful discussion and reading the manuscript. This study was partly supported by National Institutes of Health grant GM30998 to W.-H. LI.
LITERATURE CITED
AKAI, K., K. YAGI, H. F. TIANO, Y.-C. E. PAN, M. SHIMIZU, K.
FONG, R. A. JUNGMANN and S. S.-L. LI, 1985 Isolation and characterization of a cDNA and a pseudogene for mouse lactate dehydrogenase-A isozyme. Int. J. Biochem. 17: 645-648. BARKER, D., M. SCHAFER and R. WHITE, 1984 Restriction sites
containing CpG show a higher frequency of polymorphism in
human DNA. Cell 3 6 131-138.
BENHAM, F. J.. S . HODCKINSON and K. E. DAVIES, 1984 A glyc- eraldehyde 3-phosphate dehydrogenase pseudogene on the
short arm of the human X chromosome defines a multigene
family. EMBO J. 3: 2635-2640.
BENTON, W. D. and R. W. DAVIS, 1977 Screening Xgt recombi- nant clones by hybridization to single plaques in situ. Science
A mutation affecting the lactate dehydrogenase locus-1 in the mouse-I. Genetical and electrophoretical characterization. Biochem. Genet. 19: 301-
309.
CHUNG, F.-Z., H. TSUJIBO, U. BHAITACHARYYA, F. S. SHARIEF and
S. S.-L. LI, 1985 Genomic organization of human lactate dehydrogenase-A gene. Biochem. J. 231: 537-541.
Lactate dehydrogenase: struc- ture and function. Adv. Enzymol. 37: 61-133.
Nucleotide sequence of the putative regulatory region of mouse lactate dehydrogenase- A gene. Biochem. J. 235: 435-439.
FUKASAWA, K. M., W. H. LI, K. YAGI, C.-C. LUO and S. S.-L. Lr,
1986 Molecular evolution of mammalian lactate dehydrogen-
ase-A genes and pseudogenes: association of a mouse processed pseudogene with a B1 repetitive sequence. Mol. Biol. Evol. 3:
GEBHARD, W., T. MEITINCER, J. HOCHTL and H. G. ZACHAU,
1 9 6 180-182.
CHARLES, D. J. and W. PRETSCH, 1981
EVERSE, J. and N. 0. KAPLAN, 1973
FUKASAWA, K. M. and S. S.-L. LI, 1986
184 K. M. Fukasawa et al.
1982 A new family of interspersed repetitive DNA sequences in the mouse genome. J. Mol. Biol. 157: 453-471.
Patterns of nucleo- tide substitutions in pseudogenes and functional genes. J. Mol.
T h e glyceraldehyde 3- phosphate dehydrogenase gene family: structure of a human cDNA and of an X chromosome linked pseudogene; amazing complexity of the gene family in mouse. EMBO J. 3: 2627- 2633.
HOLBROOK, J. J., A. LILJAS, S. J. STEINDEL and M. G. ROSSMANN, 1975 Lactate dehydrogenase. pp. 192-292. In: The Enzymes, Vol. 11, Edited by P. D. BOYER, Academic Press, New York.
A cluster of repetitive elements within a 700 base pair region in the mouse genome. Nucleic Acids Res. 11: 2 177-2 184. KANNO, T., K. SUW, M. KITAMURA, S. MIWA, S. ICHIYAMA and Y.
NISHIMURA, 1983 Lactate dehydrogenase A-subunit and B- subunit deficiencies: comparison of the physiological roles of LDH isozymes. Isozymes Curr. Top. Biol. Med. Res. 7: 131- 150.
KLECKNER, N., 198 1 Transposable elements in prokaryotes. Annu. Rev. Biochem. 1 5 341-404.
KRAYEV, A. S., K. G. SKRYABIN, A. P. BAYEV and G. P. GEORGIEV, 1980 T h e nucleotide sequence of the ubiquitous repetitive DNA sequence B1 complementary to the most abundant class of mouse fold-back RNA. Nucleic Acids Res. 8: 1201-1215. KRAYEV, A. S., T. V. MARKUSHEVA, D. A. KRAMEROV, A. P.
RYSKOV, K. G. SKRYABIN, A. A. BAYEV and G. P. GEORGIEV, 1982 Ubiquitous transposon-like repeats B1 and B2 of the mouse genome: B2 sequencing. Nucleic Acids Res. 10: 7461- 7475.
T h e sequences of an ex- GOJOBORI, T., W.-H. LI and D. GRAUR, 1982
EvoI. 18: 360-369.
HANAUER, A. and J. L. MANDEL, 1984
KALB, V. F., S. GLASSER, D. KING and J. B. LINGREL, 1983
LEMISCHKA, I. and P. A. SHARP, 1982
pressed rat a-tubulin gene and a pseudogene with an inserted repetitive element. Nature 300: 330-335.
W.-H., 1983 Evolution of duplicate genes and pseudogenes. pp. 14-37. In: Evolution of Genes and Proteins, Edited by M. NEI and R. K. KOEHN. Sinauer Associates, Sunderland, Mas- sachusetts.
S. S.-L., W. M. FITCH, Y.-C. E. PAN and F. S. SHARIEF, 1983 Evolutionary relationships of vertebrate lactate dehy- drogenase isozymes A4 (muscle), B, (heart) and C, (testis). J.
Biol. Chem. 258: 7029-7032.
S. S.-L., H. F. TIANO, K. M. FUKASAWA, K. YAGI, M. SHIMIZU, F. S. SHARIEF, Y. NAKASHIMA and Y.-C. E. PAN, 1985 Protein structure and gene organization of mouse lactate de- hydrogenase-A isozyme. Eur. J. Biochem. 149: 21 5-225. W.-H., T . GOJOBORI and N. NEI, 1981 Pseudogenes as a paradigm of neutral evolution. Nature 292: 237-239. W.-H., C.-I. WU and C.-C. LUO, 1984 Nonrandomness of
point mutations as reflected in nucleotide substitutions in pseu- dogenes and its evolutionary implications. J. Mol. Evol. 21: 58- 71.
LITTLE, P. F. R., 1982 Globin pseudogenes. Cell 28: 683-684. LEUDERS, K., A. LEDER, P. LEDER and E. KUFF, 1982
Association between a transposed alpha-globin pseudogene and retrovirus-like elements in the BALB/c mouse genome. Nature
Molecular Cloning, A Laboratory Manual. Cold Spring Harbor Laboratory,
295: 426-428.
MANIATIS, T., E. F. FRITSCH and J. SAMBROOK, 1982
Cold Spring Harbor, New York.
MARKERT, C. L., J. B. SHAKELEE and G. S. WHITT, 1975 Evolution of a gene. Science 189: 102-1 14.
MATRISIAN, L. M., G. RAUTMANN, B. E. MAGUN and R. BREATH- NACH, 1985 Epidermal growth factor or serum stimulation of rat fibroblasts induces a n elevation in mRNA levels for lactate dehydrogenase and other glycolytic enzymes. Nucleic Acids Res. 13: 71 1-726.
MESSING, J., R. CREA and P. H. SEEBERG, 1981 A system for shotgun DNA sequencing. Nucleic Acids Res. 9: 309-32 1. MONTGOMERY, E. A. and C. H. LANGLEY, 1983 Transposable
elements in Mandelian populations. 11. Distribution of three
copia-like elements in a natural population of Drosophila melan- ogaster. Genetics 104: 473-483.
Unusual a-globin- like gene that has clearly lost both globin intervening se- quences. Proc. Natl. Acad. Sci. USA 77: 2806-2809.
PAN, Y . 4 . E., F. S. SHARIEF, M. OKABE, S. HUANG and S. S.-L. LI, 1983 Amino acid sequence studies on lactate dehydrogenase C4 isozymes from mouse and rat testes. J. Biol. Chem. 258:
PIECHACZK, J. M., S. R. BLANCHARD. C. SABOUTY, L. MARTY DANI and P. JEANTEUR, 1984 Unusual abundance of vertebrate 3- phosphate dehydrogenase pseudogenes. Nature 312: 469-47 1. PROUDFOOT, N. J., 1980 Pseudogenes. Nature 286: 840-841. PROUDFOOT, N. J. and G. G. BROWNLEE, 1976 3’ non-coding
region sequences in eukaryotic mRNA. Nature 263: 21 1-214. RIGBY, P. W. J., M. DIECKMANN, C. RHODES and P. BERG,
1977 Labeling deoxyribonucleic acid to high specific activity in vitro by nick translation with DNA polymerase I. J. Mol. Biol. 113: 237-251.
DNA sequenc- ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci.
Conversion of RNA to DNA in mammals: Alu-like elements and pseudogenes. Nature 301: 47 1-472.
TEM-induced gene mutations at enzyme loci in the mouse. Environ. Mutagen. 1: 19-25.
Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol.
Drosophila genome organization: conserved and dynamic aspects. Annu. Rev. Ge- net. 15: 219-264.
Tso, J. Y., X.-H. SUN, T.-H. KAO, K. S. REECE and R. Wu, 1985 Isolation and characterization of rat and human glyc- eraldehyde 3-phosphate dehydrogenase cDNAx: genomic com- plexity and molecular evolution of the gene. Nucleic Acids Res. 13: 2485-2502.
Nucleotide se- quences of the cDNA and an intronless pseudogene for human lactate dehydrogenase-A isozyme. Eur. J. Biochem. 147: 9-1 5. WEAVER, S., M. B. COMER, C. L. JAHN, C. A. HUTCHISON, 111 and M. H. EDGELL, 1981 T h e adult beta-globin genes of the “single” type mouse C57Bl. Cell 24: 403-41 1.
WILDE, C. D., 1986 Pseudogenes. CRC Crit. Rev. Biochem. 1 9
Wu, C.-I., W.-H. LI, J. J. SHEN, R. C. SCARPULLA, K. J. LIMBACH Evolution of cytochrome c genes and pseu-
Communicating editor: W.-H. LI NISHIOKA, P., A. LEDER and P. LEDER, 1980
7005-7016.
SANGER, F., S. NICKLEN and A. R. COULSON, 1977
USA 7 4 5463-5467. SHARP, P. A., 1983
SOARES, E. R., 1979
SOUTHERN, E. M., 1975
98: 503-51 7.
SPRADLING, A. C. and G. M. RUBIN, 1981
TSUJIBO, H., H. F. TIANO and S. S.-L. LI, 1985
323-352.
and R. WU, 1986