ALCOHOL DEHYDROGENASE POLYMORPHISM IN MAIZE- SIMPLE AND COMPOUND LOCI'
DREW SCHWARTZ A N D TORU END02
Department of Botany, Indiana University, Bloomington
Received November 22, 1965
OMPOUND loci have previously been described in maize ( LAUGHNAN 1949; STADLER and EMMERLING 1956). The evidence for the duplicate nature of the locus usually comes from recombination experiments in which the two com- ponents of the complex are separated by rare crossovers. It is ordinarily difficult if not impossible to recognize the compound nature of a locus strictly on the basis of phenotype. If the action of the two genes in the complex are different enough to control different phenotypes, they are usually considered to be separate loci, even though they may have arisen by duplication of a single gene. The distinction between the single or compound form of a locus is easier in those cases where genetic differences in protein structure are analyzed, since single amino acid re- placements can be detected electrophoretically if the replacement alters the net charge of the protein (PAULING, ITANO, SINGER and WELLS 1949). A single cistron will specify a single protein form, whereas the compound locus composed
of two cistrons will specify two distinguishable forms of the protein, isozymes, if the cistrons differ in having codons which code for amino acids of unequal charge. Such a condition has been found to exist for the alcohol dehydrogenase (ADH) gene in maize. The gene which specifies this enzyme occurs singly or in duplicate. The cistrons which make up the compound locus specify different alcohol dehy- drogenase isozymes (EC 1.1.1.1). The two allelic forms found in the complex also occur singly.
MATERIALS AND METHODS
Alcohol dehydrogenase in maize occurs in the developing kernel, scutellum of the mature kernel, and plumule and root of the very young seedling. In most of the work reported in this paper the scutellum was used as the enzyme source, since enzyme concentration is highest in that tissue. A simple procedure was adopted for the electrophoretic analysis of the scutellar enzyme. Kernels were soaked i n water at 30°C for 24 hours. Each scutellum was excised from the kernels with a scalpel and squashed on a 6 x 6 mm square of Whatman # 3 mm filter paper. The filter paper squares were inserted in the starch gels for electrophoretic separation of the scutellar proteins. The electrophoretic procedure was similar to that described previously (SCHWARTZ 1960) f o r our esterase studies. Ethanol was used as the enzyme substrate. The ADH bands were developed by a modified staining technique for lactate dehydrogenase (DEWEY and CONKLIN 1960). The gels were immersed in a solution composed of the following: 15 ml 1 . 0 ~ Tris-HC1 buffer p H 8.0, 1.5 ml 0.5 M KCN solution, 1.5 ml 0 01 M nicotinamide adenine dinucleo-
'
This work was supported by National Science Foundatlon Grant GB 283371 0 D. SCHWARTZ A N D T. E N D 0
tide solution, 1.5 ml 0.01 M nitro-blue tetrazolium solution, 0.9 ml ethanol (95%), 1.5 ml 0.01 M phenazine methosulfate solution, and 128.0 ml distilled water.
The zymograms were developed at room temperature in the dark. The ADH bands started to appear within G-hr and development was allowed to progress until the bands had reached the desired intensity.
RESULTS A N D DISCUSSION
Three electrophoretically distinguishable allelic forms of an alcohol dehydro- genase have been found in maize. These forms specify isozymes with different migration rates. The isozymes migrate to the anode in starch gel electrophoresis at pH 8.5. They are designated AdW,
AdhF,
and Adhc, with the isozyme specified by theS
allele showing the slowest migration rate, and the isozyme specified by theC
allele the most rapid (Figure 1 a, b, c).
The enzyme behaves as a dimer. In plants homozygous for any one of these alleles, only a single
ADH
band is observed in one of three positions, dependingon the allele. In heterozygotes an allodimer or hybrid enzyme with an inter- mediate migration rate is observed in addition to the two autodimers (Figure lc, e, f). This is analogous to the situation observed for the
E,
(SCHWARTZ 1960) andE,
(SCHWARTZ 1964) esterases in maize as well as for a number of other enzyme systems. The analogy with theE,
esterase system extends beyond hybridFICUFIE 1.-Zymograms of scutellar alcohol dehydrogenase isozymes from homozygous and heterozygous genotypes. (a) and (d) = AdhF/AdhF, (b) = AdP/AdhS, (c) = AdhC(')/AdhF,
M A I Z E ALCOHOL DEHYDROGENASE 71 1
enzyme formation. The three North American alleles of the pH 7.5 esterase, E l F , E,’. and E l S form a series in that the FS allodimer composed of monomers specified by the two extreme alleles,
ElF
and ElS has a migration rate identical to the NN autodimer formed by the intermediate allele, E,”. This also holds for the other four E , alleles in the second series (SCHWARTZ,FUCHSMAN
and MCGRATH 1965). In the case of the Adh alleles, the SC allodimer isozyme formed in AdN/ AdhC heterozygotes migrates at the same rate as the FF isozyme found in AdhFhomozygotes (Figure IC, d ) . This suggests that the magnitude of the charge differ- ence between S and
F
is the same as that betweenF
and C.The genes which specify the three ADH isozymes are considered to be allelic, since in heterozygotes the two genes always segregate at meiosis, with the one exception to be noted later in the paper. For example, if a plant carrying AdhS
and AdhC is crossed with a plant containing only the AdhF genes, two classes of progeny are found: AdhF/AdP heterozygotes which form the
FF,
FS, and SS isozyme bands, and AdhC/AdhF heterozygotes which form the CC, CS, and SS bands, 92 and 88 respectively in a typical cross. This is the case regardless of the gene combination in the heterozygotes. If the cistrons specifying the three iso- zymes were closely linked but not allelic, each chromosome would have to carry a null allele at two of the three loci. All plants examined showed at least one of the three isozymes.The exception to segregation of allelic types mentioned above came from a strain of maize from Colombia, South America. This strain carries a compound locus which contains two alleles, AdhF and AdhC (AdhFC). The duplicate nature of the gene was deduced from the observation that the two alleles are inherited as a unit and do not segregate from each other. I n crosses of AdhFC/AdhS by Adhs/ Adhs, all the progeny which form isozymes containing the C monomer also form isozymes with the F monomer. Segregation could readily be detected from these crosses, since it would result in progeny of the constitution AdhC/AdhS which would form only the CC, CS, and SS isozymes. No recombinations separating the two alleles in the complex have been observed. The backcrosses yielded 255 ker- nels which carried all three alleles (AdhS/AdhE) and 228 kernels that formed only the SS isozyme (AdhS/AdhS). Self-pollination of the Adhs/Adhz plants gave rise to three classes of progeny, 95 AdhS/AdhS, 146 AdhS/Adh”, and 74
AdNy/AdhE.
Zymograms from kernels heterozygous for the AdhE duplication and an AdN
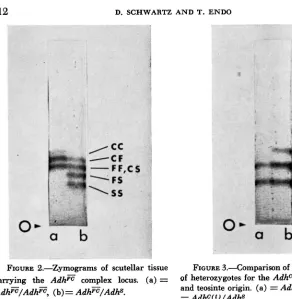
allele should show three isozyme bands, FF, FC, and CC. Only two distinct bands are seen with a very faint third band appearing if zymograms are allowed to develop for a prolonged period (Figure 2a). Kernels heterozygous for Adhz and
712 D. SCHWARTZ A N D T. ElVDO
I
0.
I.-a
b
FIGURE 2.-Zymograms of scutellar tissue FIGURE 3.4omparison of banding patterns c a w i n g the A d h E complex
AdhF/Atfh=, (b) = Adhz/Adhs.
(a) = of heterozygotes for the Adhc alleles of maize and teosinte origin. (a) = AdhC(m)/Adhs, (b) = Adhc(t)/Adhs.
which the complex
AdhE
locus was recovered was also segregating for anAdhC
allele in simplex form not associated withAdttF.
In theseAdhC
homozygotes, a weak band at the CC position is seen. TheAdhC/AdhF
orAdhC/AdhS
hetero- zygotes both show only a faint band at the CC position and two strong bands at the FC andFF,
or SC and S S positions, respectively (Figure 3a). The allele which forms the weak band has been designatedAdhc(")
to indicate its maize origin and to distinguish it from the other allele designatedAdhc(t),
which was recovered from teosinte. The CC autodimer specified by theAdhcct)
allele migrates at the same rate as the autodimer formed by theAdhc(")
allele. Furthermore, the allo- dimers formed in heterozygotes betweenAdhcct)
andAdhc(")
wi"& the other alleles have identical migration rates. The alleles differ as far as we can deter- mine only in thatAdhc(t)
produces isozyme bands of normal intensity in com- parison withAdhF
andAd@
(Figure 3). The intensity of the C"C" autodimer is low in all plant parts where the dehydrogenase is found, as in endosperm, scutellum, and plumules and roots of young seedlings. Even when the C"C" auto- dimer is too faint to be seen in the zymogram, the presence of theAdhc('")
allele can be detected inAdhS/AdhCcm)
orAdhF/AdhCfm)
heterozygotes because of the appearance of allodimers involving a Cm monomer as SC" orFC"
bands.MAIZE ALCOHOL DEHYDROGENASE 713
activity at particular positions in the gel, the deviation from the expected pattern in Adhc@' heterozygotes could result from a deficiency in the number of C" polypeptides synthesized, or a lower specific activity of C" as compared to S or F. If in an AdhC'"'/Adhs heterozygote only one half as many C" as S monomers are produced, random association would result in lCC:4CS:4SS dimers. On the other hand, if equal numbers of Cm and S monomers were synthesized, but the C"' polypeptide is inactive while the S component in SC" dimers is fully active such that a n SC" dimer has one half the activity of an SS dimer, then the SC" and SS bands would be of equal intensity. since twice as many SCm as SS dimers would be formed. A densitometer scan of the isozyme bands in the zymogram shows the CmCm band to be much less than one fourth the intensity of the SC" and SS bands (Figure 4 ) . This would argue against decreased synthesis of C" polypeptides as the factor responsible f o r the deviation in band intensities.
If
more than 50% reduction in synthesis of C monomers were postulated to account for the faintness of the CnCm band, the intensities of the SS and SCm bands would not be equal. Figure 4 shows these two bands to be of about equal intensity. These results are more consistent with a decreased specific activity of the Cm polypeptide. Previous work with the E , esterase in maize established that some alleles, designated as the prime forms, show different relative activities in comparison with the standard forms, in different tissues (SCHWARTZ 1962). A similar situa- tion has been found for the alcohol dehydrogenases. The relative activity of theAdhS allele is much lower in the endosperm than in the scutellum. In scutellar tissue heterozygous for AdhS and AdhF the two alleles show about equal activity
as determined by the observation that the intensities of the SS and FF isozyme bands are the same (Figure l f )
.
However, in heterozygous endosperm the rela- tive intensities of the FF, FS, and SS isozyme bands are very different from the distribution expected from equal activity of the two alleles. In the triploid endo- sperm of the constitution AdhF/AdhF/AdP, the isozymes should occur in a ratio of 4FF:4FS:ISS (assuming equal allele activity and random dimerization). How- ever, zymograms from such endosperm show that theFF
band is much more intense than the FS hybrid, and the SS band is extremely faint (Figure 5a). In endosperm with two doses of the AdhS allele, AdhF/AdhS/AdhS, the expected ratio is lFF:4FS:4SS. The observed ratio is close to 1FF:2FS:lSS (Figure 5b). Such results would be obtained if the AdhS allele in the endosperm were only one half as active as AdhF. I n AdhF/AdhS/AdhS endosperm equal numbers of F and S monomers would be formed giving a 1 :2:1 ratio for the dimer combinations. In contrast to the behaviour of the E , prime alleles which show full activity in young endosperm and reduced activity only in older tissue, the AdhS allele is less active than Adhp throughout the development of the endosperm. Reduced gene activity rather than reduced enzyme activity is proposed to explain these results, since in the heterozygous scutellum zymograms the FF and SS bands are of about equal intensity. The alternative explanation would require different specific activities for the same enzyme synthesized in the two tissues. This condition is certainly possible but less likely.714 D. SCHWARTZ A N D T. ENDO
FIGURE 4.--l)ensitometc~ scan of isozyme FIGURE 5.-Zymograms showing relative bands in zymograms of Adh"("l)/AdhS hetero- band intensities in heterozygous endosperm zygous scutellum. The minor unlabeled peaks from reciprocal crosses. (a) = AdhF/AdhF/ result from enzyme heterogeneity (see text). Adhs, (b) = AdhF/AdM/Adhs.
developed for prolonged periods. Zymograms developed from homozygous ma- terial which show a single intense band in the gel may also show very faint bands a t the positions to which the other auto- and allodimers migrate. For example, in zymograms from AdhS/AdkV material one can detect a weak band at the
FS
position and a still weaker band a tFF.
This condition is also seen in zymograms of heterozygotes. Another much faster migrating alcohol dehydrogenase band is also often observed in the zymograms, but it too occurs as a faint minor band. The position of this minor band varies with the genotype and there is a complete correlation between the migration rates of the major and the fast-migrating minor isozymes, as reported forADH
in Drosophila (GRELL,JACOBSON
and MURPHY 1965). Material homozygous for the Adh=duplication shows two faster migrating minor bands which correspond to those observed in the AdhP/Adho heterozygotes. The inheritance of the minor ADH isozymes is now under in- vest igat ion.The assistance of MRS. KAY JAMES is gratefully acknowledged.
SUMMARY
MAIZE ALCOHOL DEHYDROGENASE 715
and Adhcft’, appear to specify isozymes with identical migration rates but differ- ent specific activity. The AdhF and AdhCfm’ alleles are found associated in a com- pound locus as well as occurring singly. The enzyme behaves as a dimer in that hybrid enzymes are formed in heterozygotes.
LITERATURE CITED
DEWEY, M. M., and J. L. CONKLIN, 1960
GRELL. E. H., K. B. JACOBSON, and J. B. MURPHY, 1965
LAUGHNAN, J. R., 1949
PAULING. L., H. A. ITANO, S. J. SINGER, and I. C. WELLS, 1949
SCHWARTZ, D., 1960
Starch gel electrophoresis of lactic dehydrogenase
Alcohol dehydrogenase i n Drosophila
The action of allelic forms of the gene A in maize. 11. The relation of
Sickle cell anemia, a molecular
Genetic studies on mutant enzymes in maize: Synthesis of hybrid enzymes by heterozyg3tes. Proc. Natl. Acad. Sci. U S . 46: 1210-1215. - 1962 Genetic studies on mutant enzymes in maize. 111. Control of gene action in the synthesis of pH 7.5 esterase. Genetics 47: 1609-1615.
-
1964 A second hybrid enzyme in maize. Proc. Natl. Acad. Sci. U.S. 51: 602-605.Allelic isozymes of the pH 7.5 esterase
Relation of unequal crossing over to the inter- from rat kidney. Proc. Exptl. Biol. Med. 105: 492-494.
melanogaster: Isozymes and genetic variants. Science 149 : 80-82.
crossing over to mutat:on of A b . Proc. Natl. Acad. Sci. U.S. 35: 167-1 78.
disease. Science 110: 543-548.
SCHWARTZ, D., L. FUCHSMAN, and K. H. MCGRATH, 1965
STADLER, L. J., and M. H. EMMERLING, 1956 in maize. Genetics 52: 1965-1268.