0095-1137/97/$04.00
1
0
Copyright
q
1997, American Society for Microbiology
Protease-Defective, gp120-Containing Human Immunodeficiency
Virus Type 1 Particles Induce Apoptosis More Efficiently than Does
Wild-Type Virus or Recombinant gp120 Protein in Healthy
Donor-Derived Peripheral Blood T Cells
MASANORI KAMEOKA,
1TAKURO KIMURA,
1YONG-HUI ZHENG,
1SATOKO SUZUKI,
1KOH FUJINAGA,
1RONALD B. LUFTIG,
2ANDKAZUYOSHI IKUTA
1*
Section of Serology, Institute of Immunological Science, Hokkaido University, Kita-ku, Sapporo 060, Japan,
1and Department of Microbiology, Immunology and Parasitology, Louisiana State University Medical Center,
New Orleans, Louisiana 70112
2Received 6 June 1996/Returned for modification 30 August 1996/Accepted 4 October 1996
Apoptosis and syncytium formation are two mechanisms by which human immunodeficiency virus type 1
(HIV-1) impairs uninfected CD4
1T-cell function and are mainly involved in the progression of the disease to
AIDS. Previously, we showed that gp120-containing, protease-deficient HIV-1 (L-2) particles generated
syn-cytia by particle-mediated fusion with uninfected cultured CD4
1T cells. Here, we present evidence that such
L-2 particles can induce apoptosis in 40 to 50% of T cells which were enriched from HIV-1-negative healthy
donor-derived peripheral blood mononuclear cells (PBMC-Ts). Activation of PBMC-Ts with
phytohemagglu-tinin, concanavalin A, or ionomycin after incubation with L-2 particles resulted in the loss of proliferative
capacity and gradual induction of apoptosis over 3 days. Wild-type strain LAI particles or recombinant gp120
were markedly less efficient (
<
15%) at inducing such apoptosis. Western blot (immunoblot) analysis revealed
that L-2 particles contained a larger amount of Env gp120 than LAI particles. Either preincubation of
PBMC-Ts with a Fas antagonist or preincubation of L-2 particles with soluble CD4 blocked most of the
apoptosis. This suggests that L-2-like particles can play a major role in HIV-1-induced apoptosis of uninfected
bystander cells.
Human immunodeficiency virus type 1 (HIV-1), the
etiolog-ical agent of AIDS, induces depletion of CD4
1T cells, which
is one of the consequences of the pathogenesis of AIDS.
Re-cent findings have suggested that this depletion occurs in part
by apoptosis attributable directly to HIV-1 replication (10, 15)
or indirectly to the action of HIV-1-encoded proteins such as
soluble Env gp120 (2, 21) or Tat (11, 27). The latter type of
apoptosis is believed to be mediated by the Fas-Fas ligand
(FasL) interaction (8). Previous reports have provided
evi-dence that increased levels of virus load and the number of
HIV-1 transcripts are closely correlated with disease
progres-sion (3, 22, 24). In addition, the ongoing dynamics of HIV-1
replication in vivo have been demonstrated by quantifying the
changes in viral genotype and phenotype that occur during the
turnover of CD4
1T cells (23, 26). These studies suggest a
direct role of HIV-1 infection in the killing of infected T cells.
However, the overall number of HIV-1 genome-positive T
cells in the blood of infected asymptomatic individuals is only
less than 0.1% (3, 22). Nevertheless, the vast majority of T cells
in peripheral blood can be induced to undergo apoptosis upon
ex vivo activation with mitogen or anti-CD3 antibody (6, 25).
Furthermore, recent in situ observations revealed that HIV-1
can induce apoptosis in a patient’s uninfected CD4
1T cells as
a bystander effect (4). Thus, it is likely that HIV-1-mediated
apoptosis in uninfected T cells can be regulated by indirect
mechanisms.
In previous studies we established that most subclones
de-rived from surviving cells after MT-4 cells, a CD4
1human
T-cell line, had been infected with wild-type strain LAI or
NL432 derived from infectious molecular clone pNL432
pro-duced infectious, but weakly cytopathogenic, HIV-1 particles
(19, 28). The accessory genes of these particles, such as vif, vpr,
and vpu, had defective genomes (9, 18). Essential genes of
about 10% of the subclones produced with LAI virus had
defective phenotypes, and in particular, one of these, named
L-2, produced protease-defective noninfectious particles (28).
Surprisingly, the L-2 particles were found to induce syncytia by
virus-to-cell fusion (20). Therefore, we developed a model that
suggests that such L-2-like particles might be found among
defective particles in late-stage asymptomatic HIV-1-infected
patients, and if formed, they could play a significant role in the
pathogenesis of AIDS (12). In this study we used L-2 particles
to show that after adsorption, activation-induced apoptosis
with mitogens is greatly enhanced in T-cell-enriched
periph-eral blood mononuclear cells (PBMC-Ts) from
HIV-1-nega-tive healthy donors relaHIV-1-nega-tive to that induced by wild-type LAI or
soluble recombinant gp120 (rgp120).
MATERIALS AND METHODS
Cells and viruses.The culture fluid of MOLT-4 cells persistently infected with the LAI strain of HIV-1 was used as the source of wild-type virus particles. The culture fluid of L-2 cells, a subclone obtained from MT-4 cells that is persistently infected with the LAI strain (28) and that contains pol-defective proviral DNA (17), was used as the source of noninfectious HIV-1 (L-2) particles. Thus, the L-2 particles contain uncleaved Pr55gag and cleaved gp120, and all pol-encoded
enzymes, i.e., protease, reverse transcriptase, and integrase, are defective. The L-2 or wild-type LAI particles were collected from culture fluids by centrifuga-tion at 100,0003g for 30 min as described previously (7).
Preparation of PBMC-Ts.Peripheral blood mononuclear cells (PBMCs) were obtained by centrifugation on Ficoll-Hypaque (Pharmacia Biotech, Uppsala, Sweden) gradients of heparinized venous peripheral blood from HIV-1-negative healthy donors. PBMC-Ts were obtained from PBMCs by use of a nylon wool
* Corresponding author. Phone and fax: 81-(11)-707-6837. E-mail:
[email protected].
41
on May 15, 2020 by guest
http://jcm.asm.org/
magglutinin (PHA; 10mg/ml), concanavalin A (ConA; 10mg/ml), or ionomycin (1mg/ml) in 10% FCS–RPMI 1640 medium containing 25 U of recombinant interleukin-2 (rIL-2) for 3 days as described previously (6). As a control, PBMC-Ts were cultured for 3 days in 10% FCS–RPMI 1640 medium to which no stimulant or rIL-2 was added.
Lymphocyte proliferation assay.PBMC-Ts were treated as described above with optimal concentrations of PHA, ConA, or ionomycin in 96-well microplates. After cultivation for 2 days, proliferation rates were determined by adding 1mCi of [3H]thymidine ([3H]TdR; NEN-DuPont, Boston, Mass.) to each microculture, essentially as described previously (25). Following a 16-h incubation, the cells were harvested by using FILTERMATE 196 (Packard Instrument Company, Meriden, Conn.), and the level of [3
H]TdR incorporation into the cells was measured with a beta counter (MATRIX 96; Packard Instrument Company). Results represent the mean counts per minute for triplicate microcultures.
p24 ELISA.The amount of virus inoculum was measured by an enzyme-linked immunosorbent assay (ELISA) for the Gag p24 antigen according to the proto-col described for the kit (INTRACEL Corporation). Both mature p24 and immature uncleaved Gag precursor p55 and p40 are detected by this kit.
Measurement of cell mortality.Evaluation of viable and dead cell numbers was performed by a trypan blue dye exclusion assay (6). The percent cell mor-tality was calculated as follows: 1003[dead cell number/(viable cell number1 dead cell number)].
Measurement of apoptosis.Apoptosis was measured by three methods. First, quantitative analysis of apoptosis was performed by staining cells with propidium iodide and performing flow cytometric analysis as described previously (11). Briefly, the cells were fixed with cold 70% ethanol. After standing on ice for 30 min, the cells were centrifuged, resuspended in RNase A in phosphate-buffered saline (500mg/ml), and incubated at 378C for 20 min. After centrifugation, the cells were again resuspended in propidium iodide in phosphate-buffered saline (50mg/ml) and incubated on ice for 10 min. Finally, the cells were analyzed for their DNA content by using the Becton Dickinson FACScan system. Second, morphological changes of the nuclei were visualized with a fluorescence micro-scope (BH2-RFCA; Olympus, Tokyo, Japan) after staining with propidium io-dide as described above, except that the cells were fixed with 4% paraformalde-hyde for 2 h. Third, DNA fragmentation was determined by agarose gel electrophoresis of extracted cellular DNA as described previously (1).
Measurement of cell surface antigens.Expression of cell surface antigens was measured by flow cytometry. Briefly, 106PBMC-Ts were incubated with either anti-CD3 monoclonal antibody (MAb) (OKT-3; Ortho Diagnostic Systems, Rari-tan, N.J.) or anti-Fas MAb (CH-11; Medical and Biological Laboratories Co., Ltd., Nagoya, Japan) at a concentration of 10mg/ml at 48C for 30 min. After incubation, the cells were washed and stained with either FITC-conjugated rabbit anti-mouse IgG (Dako A/S) or an FITC-conjugated goat affinity-purified F(ab9)2fragment to mouse IgM (mchain) (Cappel) for 30 min. These cells were analyzed by using the Becton Dickinson FACScan system. Ratios of CD41and CD81T cells were determined by using anti-CD4 (OKT-4; Ortho Diagnostic Systems) and anti-CD8 (OKT-8; Ortho Diagnostic Systems) MAbs, followed by FITC-conjugated rabbit anti-mouse IgG (Dako A/S).
RT-PCR of FasL transcript.The levels of expression of FasL in the PBMC-Ts with or without adsorption of L-2 particles were semiquantified by using reverse transcriptase (RT) PCR. Briefly, cellular RNA extracted from PBMC-Ts which had been mock treated or adsorbed with L-2 particles for 18 h as described above was reverse transcribed by using oligo(dT)16as a primer. The cDNA was sub-jected to PCR with oligonucleotide primers specific to FasL mRNA (TGTTTC AGCTCTTCCACCTACA [positions 387 to 408] and AGAGGTGAGTTAAA AGGTGTCC [positions 1268 to 1247]) (1) and with glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA (GATGCTGGCGCTGAGTACGTCG [po-sitions 325 to 346] and GTGGTGCAGGAGGCATTGCTGA [po[po-sitions 521 to 500]) (29) as a control. The PCR products of FasL and GAPDH were separated by agarose gel electrophoresis and were blotted onto a Hybond N1membrane (Amersham International plc, Little Chalfont, United Kingdom), followed by hybridization with individual32P-labeled internal oligonucleotide probes as de-scribed previously (22). The hybridized bands were visualized with an imaging analyzer (Bas-1000; FUJIX, Tokyo, Japan).
Blocking assays of L-2 particle-mediated apoptosis.PBMC-Ts were preincu-bated at 378C for 2 h with 1mg of Fas antagonist (anti-Fas MAb ZB-4; Medical
detected with an anti-Gag p24 MAb (MAb V107) which could recognize both mature (p24) and immature (p40 and p55) types of HIV-1 capsid proteins (7). Each band of either Env gp120 or the Gag protein visualized on the X-ray film was scanned by laser densitometry (FAST SCAN; Molecular Dynamics, Sunny-vale, Calif.), and a relative ratio of Env gp120/total Gag protein (Env/Gag) was calculated for each lane.
RESULTS
Decreased proliferation of PBMC-Ts occurs after
adsorp-tion with L-2 particles more effectively than it does with
wild-type HIV-1 or rgp120 cross-linking.
PBMC-Ts were prepared
from HIV-1-negative healthy donors. The fractions contained
.
95% CD3
1T cells by fluorescence-activated cell sorter
anal-ysis (data not shown). These PBMC-Ts were incubated for 18 h
with either defective L-2 or wild-type LAI particles.
Alterna-tively, PBMC-Ts were treated with rgp120 for 1 h, followed by
incubation for 17 h with anti-HIV-1 polyclonal IgG for the
cross-linking of CD4 molecules. The efficiency of this
anti-HIV-1 polyclonal IgG in cross-linking CD4 was confirmed by
membrane fluorescence, and capping of CD4-rgp120 with
anti-HIV-1 polyclonal IgG on the surface of PBMC-Ts was
de-tected (data not shown). Cells were then stimulated with PHA,
ConA, or ionomycin for another 2 days. As a control, cells were
mock treated and similarly stimulated with PHA, ConA, or
ionomycin.
Cell proliferation rates as measured by [
3H]TdR
incorpora-tion for 16 h revealed that the levels of incorporaincorpora-tion of
[
3H]TdR in mock-treated or LAI-adsorbed cells were about
the same for all three stimulants (Fig. 1A). In contrast, the
level of incorporation of [
3H]TdR in PBMC-Ts adsorbed with
L-2 particles and then stimulated with PHA, ConA, or
iono-mycin was significantly (70 to 90%) reduced (Fig. 1A).
rgp120-cross-linked PBMC-Ts showed about the same level of
[
3H]TdR incorporation, independent of whether 100 ng, 1
m
g,
or 10
m
g of rgp120 per ml was added (Fig. 1B).
Increase in PBMC-T mortality after adsorption with L-2
particles.
Concomitant with the significant decrease in
prolif-erative response observed in PBMC-Ts after adsorption with
L-2 particles followed by mitogenic stimulation (Fig. 1A and
B), we found that by 3 days after such treatment there was also
a large (four- to fivefold) increase, to about 40% over the
background level, in the percent PBMC-T mortality (Fig. 1C).
In contrast, PBMC-Ts adsorbed with wild-type HIV-1 LAI
particles (Fig. 1C) or cross-linked with rgp120 at
concentra-tions of
#
1
m
g/ml (Fig. 1D) exhibited almost no increase (0 to
10%) after stimulation with PHA, ConA, or ionomycin.
Fur-thermore, even at the highest concentration (10
m
g/ml), rgp120
cross-linking induced a slight increase (
,
15% over the
back-ground level) compared with L-2 treatment (Fig. 1C).
Apoptosis of PBMC-Ts after adsorption with L-2 particles.
In order to confirm that the increased PBMC-T mortality
noted above arose as a result of induction of apoptosis by L-2
particles, the PBMC-Ts were examined by flow cytometry.
on May 15, 2020 by guest
http://jcm.asm.org/
tially, the cells were adsorbed with L-2 or LAI particles or
cross-linked with rgp120 (10
m
g/ml) for 18 h, and then this step
was followed by stimulation with PHA, ConA, or ionomycin
for an additional 3 days. The stimulated PBMC-Ts were
ana-lyzed for their DNA contents, after staining with propidium
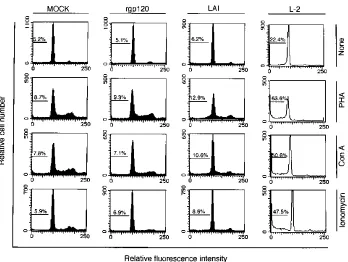
iodide, by flow cytometric analysis (Fig. 2). These results were
compared with those for cells that had been mock treated. As
seen in the flow cytometric histogram, most of the cell
popu-lation showed a DNA peak at the G
0or G
1phase in the
absence of stimulant, while a small percentage (10 to 15%) of
the population shifted to the S or the G
2or M phase after
PHA, ConA, or ionomycin stimulation. As a measure of
apo-ptosis, we used the percentage of cells with hypodiploid DNA
content. Thus, for the mock treatment, there was a 6 to 10%
background spontaneous apoptosis for particle- or
non-rgp120-treated PBMC-Ts. In stark contrast to these results, the
histogram of PBMC-Ts after adsorption with L-2 particles and
mitogenic treatment reveals a major increase in the percentage
of cells with hypodiploid DNA (Fig. 2, L-2 panels). After
subtraction of the background percentage (mock-treated
cells), there was about a 50% increase in apoptotic cell
num-bers after PHA stimulation and a 40% increase after ConA or
ionomycin stimulation. Unstimulated cells showed a 17%
in-crease in apoptosis after L-2 particle exposure. Under the
same assay conditions, but with wild-type LAI adsorption or
rgp120 treatment, there was only a minimum increase (
,
5%)
in the apoptosis of PBMC-Ts after adsorption (Fig. 2, rgp120
and LAI panels). In fact, for LAI or rgp120, there was even
some movement of cells into the G
2-M boundary, as had been
seen with mock treatment.
As an additional assay procedure, propidium iodide-stained
PBMC-Ts which had been adsorbed with L-2 particles and
stimulated with PHA, ConA, or ionomycin were then
exam-ined by immunofluorescence microscopy to determine if there
was any observable nuclear destruction characteristic of
apo-ptosis. As noted in Fig. 3, relative to mock-treated cells, a
substantial degree of nuclear disintegration was observed for
L-2-treated PBMC-Ts after mitogen stimulation. In addition,
degradation of DNA into oligonucleosome-sized fragments
was observed only in L-2 particle-adsorbed PBMC-Ts after
mitogenic stimulation but not after mock treatment (data not
shown). In contrast, no observable nuclear destruction of
PBMC-Ts adsorbed with LAI particles or treated with rgp120
was observed (data not shown). Results similar to those in Fig.
2 and 3 were obtained for PBMC-T samples from four
differ-ent healthy donors.
Semiquantitative analysis of envelope proteins in L-2 and
LAI particles.
On the basis of our findings that L-2 particles
could induce syncytium formation (by fusion from without) of
MOLT-4 cells (20) and apoptosis of PBMC-Ts (Fig. 2 and 3),
we examined the possible mechanism by which L-2 particles, in
comparison with wild-type LAI virus, exhibited these enhanced
functions. One possibility is a different content of surface
gp120 between L-2 and wild-type LAI particles. Therefore,
comparative Western blot analysis of these particles was
per-formed. Serial dilutions of L-2 and wild-type LAI particles,
prepared similarly as for the induction of apoptosis and
ad-justed to the same protein content by Gag p24 antigen capture
ELISA, were subjected to Western blotting with an anti-gp120
MAb. The result clearly showed that there was a larger amount
of gp120 for the same starting amount of L-2 Gag protein than
was found for wild-type LAI particles (Fig. 4A). As an internal
control, the same polyvinylidene difluoride membrane was
re-acted with anti-Gag p24 MAb, and the results presented in Fig.
FIG. 1. Proliferation rates and percent cell mortalities of PBMC-Ts after adsorption with HIV-1 particles (L-2 and LAI) or rgp120, followed by mitogenic stimulation. PBMC-Ts (23106/ml) were adsorbed with 6 ng of defective L-2 (h) or wild-type LAI particles (3) per ml as well as mock treated with medium (■) for 18 h at 378C (A and C). Also, PBMC-T cells were untreated (■) or treated with 100 ng (s), 1mg (o), or 10mg (h) of rgp120 per ml for 1 h at 378C, followed by cross-linking with anti-HIV-1 polyclonal IgG for 17 h at 378C (B and D). After incubation, the cells were stimulated with PHA (10mg/ml), ConA (10mg/ml), or ionomycin (1mg/ml). Then, the [3H]TdR incorporation rate at 2 days after stimulation (A and B) or the percent cell mortality at 3 days after stimulation (C and D) was measured as described in Materials and Methods. The data obtained with PBMC-Ts from one donor are presented here as representative for data obtained with cells from all donors. Standard deviations from the mean (error bars) were calculated from triplicate assays. Essentially similar results were obtained for six different healthy donor-derived PBMC-T samples.
on May 15, 2020 by guest
http://jcm.asm.org/
4B were observed. The relative ratio of Env/Gag was
calcu-lated by laser densitometry scanning to be about fourfold
larger in L-2 particles (ratio of 0.8) than LAI particles (ratio of
0.2). The result in Fig. 4B also indicates a similar standard
deviation from the mean (error bars) calculated from the Env/
Gag ratio in each lane.
Detection of Fas and FasL expression in PBMC-Ts
ad-sorbed with L-2 particles.
In order to determine the
mecha-nism by which apoptosis is induced in PBMC-Ts after
adsorp-tion with L-2 particles, the cells were initially analyzed by flow
cytometry for cell surface expression of Fas antigen. PBMC-Ts
adsorbed with L-2 particles expressed an enhanced level of Fas
antigen (Fig. 5A). Also, adsorption with wild-type LAI
parti-cles or rgp120 cross-linking (10
m
g/ml) led to a similar level of
enhanced Fas antigen expression, irrespective of the absence
[image:4.612.133.481.67.329.2]FIG. 2. Flow cytometric histogram of L-2 particle-adsorbed PBMC-Ts. PBMC-Ts from a healthy donor were adsorbed with L-2 particles, as described in the legend to Fig. 1. As controls, PBMC-Ts were adsorbed with LAI particles, treated with rgp120 (10mg/ml), or mock treated. After incubation for 18 h at 378C, the cells at a density of 106/ml were unstimulated (None) or stimulated with a final concentration of 10mg of PHA or ConA per ml or 1mg of ionomycin per ml for 3 days. All the cells were then stained with propidium iodide and characterized for their DNA content by flow cytometry. The percentages of apoptotic cells are indicated in each panel. Note that only for L-2 particles was a decreased G2-M peak seen after mitogen stimulation. Essentially similar profiles were obtained with four different healthy donor-derived PBMC-T samples.
FIG. 3. Fluorographic examination of L-2-adsorbed PBMC-Ts after mito-genic stimulation. PBMC-Ts were adsorbed with L-2 particles as described in the legend to Fig. 1. As a control, the same cells were mock treated with medium. The cells were then stimulated with PHA, ConA, or ionomycin as described in the legend to Fig. 1. As a control, the cells were treated with medium (None). Finally, cells were labeled with propidium iodide and then observed under a fluorescence microscope. Cell fragments are indicated by white arrowheads.
FIG. 4. Quantitative analysis of particle-associated HIV-1 proteins. (A) Threefold serial dilutions of L-2 or LAI particles were subjected to Western blot analysis as described in Materials and Methods. HIV-1 Env gp120 or Gag proteins were detected with MAbs to each protein, followed by visualized with enhanced chemiluminescence Western blotting detection reagents. The numbers above each lane indicate the p24 content of each particle preparation, which was estimated with a p24 antigen capture ELISA kit. (B) Each band of Env gp120 or Gag protein was scanned by laser densitometry and the relative ratio of Env/Gag was calculated. Error bars represent standard deviations from the Env/Gag ratio in each of the three lanes.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:4.612.63.295.511.668.2]of the significant apoptosis observed as a result of these
treat-ments. Next, we also analyzed the level of expression of FasL
mRNA in PBMC-Ts at 18 h after adsorption with L-2 particles,
since strong cell aggregation which seemed similar to that after
mitogenic stimulation was induced only in L-2
particle-ad-sorbed PBMC-Ts and not in LAI-adparticle-ad-sorbed,
rgp120-cross-linked, or mock-treated PBMC-Ts (data not shown). The
re-sults obtained by RT-PCR indicated that FasL expression was
enhanced in PBMC-Ts after adsorption with L-2 particles
com-pared with that in mock-treated cells (Fig. 5B), as shown by a
higher relative ratio of FasL to GAPDH PCR products (Fig.
5C).
Blocking of L-2 particle-mediated apoptosis by Fas
antago-nist or soluble CD4.
To further confirm the contribution of the
Fas-FasL interaction in L-2 particle-mediated apoptosis, a
blocking assay was performed by using as the Fas antagonist a
MAb to Fas antigen that does not cause apoptosis. PBMC-Ts
were preincubated with the Fas antagonist for 2 h and were
then adsorbed with L-2 particles for 18 h prior to PHA, ConA,
or ionomycin stimulation. The percent cell mortality assayed
by trypan blue dye exclusion and the cell proliferation rate
measured by the [
3H]TdR incorporation assay were
deter-mined as described above. The results in Fig. 6A indicate that
the Fas antagonist leads to a blocking effect, from about 45 to
15% cell mortality. In parallel with this blocking, the
prolifer-ation rate of L-2 particle-adsorbed PBMC-Ts was partially (60
to 80%) restored in the presence of the Fas antagonist (1
m
g/ml) (Fig. 6B).
Since, a priori, we suspected that gp120-containing L-2
par-ticles bind to T cells via CD4 molecules, we designed a
block-ing experiment to determine if CD4 was functionally important
in the observed L-2-induced apoptosis of PBMC-Ts. L-2
par-ticles were preincubated for 2 h with soluble CD4 prior to
adsorption with PBMC-Ts. After the 18-h incubation with L-2
particles, PBMC-Ts were stimulated with PHA, ConA, or
ionomycin for 3 days, as described above, and the percent cell
mortality and the level of [
3H]TdR incorporation were
mea-sured. It can be seen in Fig. 6 that soluble CD4 gives a blocking
effect on both the level of cell mortality, from 45 to 15% (Fig.
6A), and partial restoration of the proliferation rate (50 to
85%) (Fig. 6B), an effect analogous to that seen with the Fas
antagonist.
DISCUSSION
We have presented evidence that protease-defective,
gp120-containing HIV-1 particles induce a significant level (40 to
50%) of apoptosis in healthy donor-derived PBMC-Ts after
subsequent mitogenic stimulation with PHA, ConA, or
iono-mycin (Fig. 1 to 3). Wild-type HIV-1 particles and monomeric
rgp120 are markedly less effective in inducing such apoptosis (5
to 15%) (Fig. 1 and 2). Although several reports indicated that
FIG. 5. Expression of Fas and FasL by adsorption with L-2 particles. (A) Expression of Fas antigen on the surface of PBMC-Ts after adsorption with L-2 or LAI particles or treatment with rgp120. PBMC-T cells were adsorbed with L-2 or LAI particles, treated with 10mg of rgp120 per ml, or mock treated as described in the legend to Fig. 1. After the treatments, the levels of expression of Fas antigen on the surfaces of PBMC-Ts were measured by flow cytometry (see Materials and Methods). (B) Enhanced level of expression of FasL mRNA in PBMC-Ts after adsorption with L-2 particles. PBMC-Ts were adsorbed with L-2 particles for 18 h, as described in the legend to Fig. 1. As a control, mock-treated PBMC-Ts were used. After incubation, total cellular RNA was extracted and subjected to RT-PCR. Then, the PCR products of FasL and GAPDH mRNA were hybridized with individual32P-labeled internal oligonucleotide probes, followed by visualization of hybridized bands by imaging analyzer, as described in Materials and Methods. (C) The relative ratio of FasL mRNA levels/GAPDH mRNA levels was calculated.
FIG. 6. Blocking of L-2 particle-mediated apoptosis by Fas antagonist or soluble CD4. PBMC-Ts were mock treated (■) or adsorbed with L-2 particles for 18 h (3), as described in the legend to Fig. 1. The same cells were preincubated with Fas antagonist for 2 h and were subsequently adsorbed with L-2 particles (s). In addition, PBMC-Ts were also adsorbed with L-2 particles, which had been preincubated with soluble CD4 for 2 h, and were then further incubated for 18 h as described in the legend to Fig. 1 prior to mitogenic stimulation (h). After incubation, the cells were stimulated with PHA, ConA, or ionomycin as de-scribed in the legend to Fig. 1. The percent cell mortality at 3 days after stimu-lation (A) and the [3H]TdR incorporation rate at 2 days after stimulation (B) were measured as described in the legend to Fig. 1. Error bars represent standard deviation from the mean for three different samples from healthy donors.
on May 15, 2020 by guest
http://jcm.asm.org/
anti-CD3 antibody (25); and (iii) induction of apoptosis occurs
primarily in uninfected bystander T cells (4). It is also known
that in the blood of asymptomatic carriers, greater than
99.99% of the virus in plasma is not infectious (24). These
observations are not inconsistent with our results; i.e.,
apopto-sis in patients can be induced by adsorption to healthy
PBMC-Ts by very low levels of L-2 like particles, since a final
concentration of 2.0 ng of L-2 particles per 10
6PBMC-Ts was
enough for apoptosis induction (data not shown). In the case
of LAI particles, even 12 ng per 10
6PBMC-Ts was not effective
for apoptosis induction (data not shown). These results suggest
the importance of the total gp120 content per virion and its
conformation for the induction of apoptosis induction. Since
our preliminary data on the sequence of the L-2 provirus
revealed that the mutations occur at least in vpr, env gp41, and
nef, in addition to the pol protease, specific mutations might be
involved in this L-2 Env function.
A recent result noted by Maldarelli et al. (14) has shown that
rapid induction of apoptosis can occur through cell-to-cell
transmission of HIV-1 without virus replication and depends
on a mechanism that requires a CD4-Env association. We
believe that the L-2 particle-induced apoptosis that we
ob-served is similar to this effect for several reasons: (i) the L-2
particles are replication defective (17); (ii) L-2 particles
pos-sess large amounts of gp120 (Fig. 4) and efficiently induce
syncytium formation in MOLT-4 cells (20); (iii) L-2 particles
induce apoptosis in 40 to 50% of PBMC-Ts, while under the
same assay conditions wild-type LAI virus does not lead to a
significant degree of apoptosis; and (iv) L-2 particle-mediated
apoptosis was blocked by soluble CD4 (Fig. 6). In addition, we
detected aggregations of PBMC-Ts adsorbed with L-2 particles
but not cells adsorbed with LAI particles of rgp120 (data not
shown). This may reflect the larger amount of surface gp120
found on L-2 or a stronger fusion activity of L-2 particles (by
fusion from without) (20). In our system, such cell aggregations
seemed to be closely related to the induction of apoptosis in
PBMC-Ts, as LAI-adsorbed or rgp120-treated PBMC-Ts did
not show significant levels of apoptosis.
Finally, we were interested in determining the mechanism
behind the observed apoptosis in PBMC-Ts adsorbed with L-2
particles. Our experiments suggest that it involves the surface
expression of Fas antigen and FasL mRNA (Fig. 5), which is
consistent with what has been found during the normal positive
and negative selections of T cells during ontogeny of the
im-mune response (13). Alderson et al. (1) have also recently
reported that previously activated T cells undergo apoptosis
when stimulated through CD3–T-cell receptor, and this is
ev-idently mediated by Fas-FasL interactions. In our assay system,
an enhanced level of expression of Fas antigen was observed
on the cell surface of all PBMC-T cells adsorbed with L-2 or
LAI particles, as well as cells treated with rgp120 (Fig. 5A).
However, only PBMC-T cells adsorbed with L-2 particles, but
not with LAI particles or rgp120 protein, showed an enhanced
ACKNOWLEDGMENTS
This work was supported in part by a grant-in-aid for scientific
research on priority areas from the Ministry of Education, Science,
Sports and Culture; a grant-in-aid for AIDS research from the Ministry
of Health and Welfare; and a special grant-in-aid for the promotion of
education and science at Hokkaido University provided by the
Minis-try of Education, Science, Sports and Culture of Japan.
REFERENCES
1. Alderson, M. R., T. W. Tough, T. Davis-Smith, S. Braddy, B. Falk, K. A. Schooley, R. G. Goodwin, C. A. Smith, F. Ramsdell, and D. H. Lynch.1995. Fas ligand mediates activation-induced cell death in human T lymphocytes. J. Exp. Med. 181:71–77.
2. Banda, N. K., J. Bernier, D. K. Kurahara, R. Kurrle, N. Haigwood, R. P. Sekaly, and T. H. Finkel.1992. Crosslinking CD4 by human immunodefi-ciency virus gp120 primes T cells for activation-induced apoptosis. J. Exp. Med. 176:1099–1106.
3. Embretson, J., M. Zupancic, J. L. Ribas, A. Burke, P. Racz, K. Tenner-Racz, and A. T. Haase.1993. Massive covert infection of helper T lymphocytes and macrophages by HIV during the incubation period of AIDS. Nature 362: 359–362.
4. Finkel, T. H., G. Tudor-Williams, N. K. Banda, M. F. Cotton, T. Curiel, C. Monks, T. W. Baba, R. M. Ruprecht, and A. Kupfer.1995. Apoptosis occurs predominantly in bystander cells and not in productively infected cells of HIV- and SIV-infected lymph nodes. Nature Med. 1:129–134.
5. Foster, S., P. Beverley, and R. Aspinall. 1995. gp120-induced programmed cell death in recently activated T cells without subsequent ligation of the T cell receptor. Eur. J. Immunol. 25:1778–1782.
6. Groux, H., G. Torpier, D. Monte, Y. Mouton, A. Capron, and J. C. Ameisen. 1992. Activation-induced death by apoptosis in CD41T cells from human immunodeficiency virus-infected asymptomatic individuals. J. Exp. Med. 175:331–340.
7. Ikuta, K., C. Morita, S. Miyake, T. Ito, M. Okabayashi, K. Sano, M. Nakai, K. Hirai, and S. Kato.1989. Expression of human immunodeficiency virus type 1 (HIV-1) gag antigens on the surface of a cell line persistently infected with HIV-1 that highly expresses HIV-1 antigens. Virology 170:408–417. 8. Katsikis, P. D., E. S. Wunderlich, C. A. Smith, L. A. Herzenberg, and L. A.
Herzenberg.1995. Fas antigen stimulation induces marked apoptosis of T lymphocytes in human immunodeficiency virus-infected individuals. J. Exp. Med. 181:2029–2036.
9. Kishi, M., Y.-H. Zheng, M. K. Bahmani, K. Tokunaga, H. Takahashi, M. Kakinuma, P. K. Lai, M. Nonoyama, R. B. Luftig, and K. Ikuta.1995. Naturally occurring accessory gene mutations lead to persistent human im-munodeficiency virus type 1 infection of CD4-positive T cells. J. Virol. 69:7507–7518.
10. Laurent-Crawford, A. G., B. Krust, S. Muller, Y. Riviere, M.-A. Rey-Cuille, J.-M. Bechet, L. Montagnier, and A. G. Hovanessian.1991. The cytopathic effect of HIV is associated with apoptosis. Virology 185:829–839. 11. Li, C. J., D. J. Friedman, C. Wang, V. Metelev, and A. B. Pardee. 1995.
Induction of apoptosis in uninfected lymphocytes by HIV-1 tat protein. Science 268:429–431.
12. Luftig, R. B., and K. Ikuta. 1994. Are defective, HIV protease-deficient particles the real culprit in AIDS? A specific type of defective particle produced after persistent infection of lymphoid cells may be the trigger in pathogenesis. ASM News 60:417–419.
13. Lynch, D. H., F. Ramsdell, and M. R. Alderson. 1995. Fas and FasL in the homeostatic regulation of immune responses. Immunol. Today 16:569–574. 14. Maldarelli, F., H. Sato, E. Berthold, J. Orenstein, and M. A. Martin. 1995. Rapid induction of apoptosis by cell-to-cell transmission of human immu-nodeficiency virus type 1. J. Virol. 69:6457–6465.
15. Martin, S. J., P. M. Matear, and A. Vyakarnam. 1994. HIV-1 infection of human CD41T cells in vitro. Differential induction of apoptosis in these cells. J. Immunol. 152:330–342.
16. Matsushita, S., M. Robert-Guroff, J. Rusche, A. Koito, T. Hattori, H.
on May 15, 2020 by guest
http://jcm.asm.org/
Hoshino, K. Javaherian, K. Takatsuki, and S. Putney.1988. Characteriza-tion of a human immunodeficiency virus neutralizing monoclonal antibody and mapping of the neutralizing epitope. J. Virol. 62:2107–2114. 17. Nakano, T., K. Sano, T. Goto, S. Morimatsu, S. Ueda, K. Ikuta, S. Kato, and
M. Nakai.1994. Analysis of pol gene of a non-infectious HIV-1 clone, abstr. PA0043. In Proceedings of the 10th International Conference on AIDS and the International Conference on Sexually Transmitted Diseases. Yokohama, Japan.
18. Nakaya, T., K. Fujinaga, M. Kishi, S. Oka, T. Kurata, I. M. Jones, and K. Ikuta.1994. Nonsense mutations in the vpr gene of HIV-1 during in vitro virus passage and in HIV-1 carriers-derived peripheral blood mononuclear cells. FEBS Lett. 354:17–22.
19. Nishino, Y., M. Kishi, M. Sumiya, K. Ogawa, A. Adachi, K. Maotani-Imai, S. Kato, K. Hirai, and K. Ikuta.1991. Human immunodeficiency virus type 1
vif, vpr, and vpu mutants can produce persistently infected cells. Arch. Virol.
120:181–192.
20. Ohki, K., M. Kishi, Y. Nishino, M. Sumiya, T. Kimura, T. Goto, M. Nakai, and K. Ikuta.1991. Noninfectious doughnut-shaped human immunodefi-ciency virus type 1 can induce syncytia mediated by fusion of the particles with CD4-positive cells. J. Acquired Immune Defic. Syndr. 4:1233–1240. 21. Oyaizu, N., T. W. McCloskey, M. Coronesi, N. Chirmule, and S. Pahwa.
1993. Accelerated apoptosis in peripheral blood mononuclear cells (PBMCs) from human immunodeficiency virus type-1 infected patients and in CD4 cross-linked PBMCs from normal individuals. Blood 82:3075–3080. 22. Pantaleo, G., C. Graziosi, J. F. Demarest, L. Butini, M. Montroni, C. H. Fox,
J. M. Orenstein, D. P. Kotler, and A. S. Fauci.1993. HIV infection is active and progressive in lymphoid tissue during the clinically latent stage of
dis-ease. Nature 362:355–358.
23. Perelson, A. S., A. U. Neumann, M. Markowitz, J. M. Leonard, and D. D. Ho. 1996. HIV-1 dynamics in vivo: virion clearance rate, infected cell life-span, and viral generation time. Science 271:1582–1586.
24. Piatak, M., M. S. Saag, L. C. Yang, S. J. Clark, J. C. Kappes, K.-C. Luk, B. H. Hahn, G. M. Shaw, and J. D. Lifson.1993. High levels of HIV-1 in plasma during all stages of infection determined by competitive PCR. Sci-ence 259:1749–1754.
25. van Noesel, C. J. M., R. A. Gruters, F. G. Terptra, P. T. A. Schellekens, R. A. W. van Lier, and F. Miedema.1990. Functional and phenotypic evi-dence for a selective loss of memory T cells in asymptomatic human immu-nodeficiency virus-infected men. J. Clin. Invest. 86:293–299.
26. Wei, X., S. K. Ghosh, M. E. Taylor, V. A. Johnson, E. A. Emini, P. Deutsch, J. D. Lifson, S. Bonhoeffer, M. A. Nowak, H. H. Hahn, M. S. Saag, and G. M. Shaw.1995. Viral dynamics in human immunodeficiency virus type 1 infec-tion. Nature 373:117–122.
27. Westendrop, M. O., R. Frank, C. Ochsenbauer, K. Stricker, J. Dhein, H. Walczak, K.-M. Debatin, and P. H. Krammer.1995. Sensitization of T cells to CD95-mediated apoptosis by HIV-1 tat and gp120. Nature 375:497–500. 28. Yunoki, M., K. Maotani-Imai, H. Kusuda, M. Motoyama, S. Miyake, H. Imai, Y. S. Shin, S. Kato, K. Sano, C. Morita, M. Nakai, K. Hirai, and K. Ikuta.1991. Production of infectious particles from defective human immu-nodeficiency virus type 1 (HIV-1)-producing cell clones by superinfection with infectious HIV-1. Arch. Virol. 116:143–158.
29. Zhao, J., N. Araki, and S. K. Nishimoto. 1995. Quantitation of matrix Gla protein mRNA by competitive polymerase chain reaction using glyceralde-hyde-3-phosphate dehydrogenase as an internal control. Gene 155:159–165.