Heroin-Conditioned Immunomodulation in Female Rats: Examining the Role of Estrogen
By: Taylor N. Weckstein Honors Thesis
Department of Psychology & Neuroscience University of North Carolina at Chapel Hill
Donald Lysle, Thesis Advisor Todd Thiele, Reader
Acknowledgements
Abstract
Opioids suppress normal immune status, impairing user response to pathogens. Using well-controlled animal models, our laboratory has established that heroin and heroin-paired cues inhibit production of pro-inflammatory biomarkers in male rats when they precede an endotoxin challenge with lipopolysaccharide (LPS). However, whether this phenomenon exists in female rats requires further scientific examination. There are well documented sex-based differences in opioid and immune responses in male and female clinical populations. For example, evidence suggests estrogen inhibits the production of interleukin-1β (IL-1β), a pro-inflammatory cytokine necessary for the expression of heroin-conditioned immunomodulation in male rats. To address this critical research gap, the present studies examine the effect of heroin and context-heroin associations on peripheral immune status in female rats. We focus investigations on peripheral nitric oxide (NO) production, a critical pro-inflammatory agent necessary for pathogen
resistance. Experiment 1 established that male and female rats have comparably impaired LPS-induced NO production when heroin (1 or 3 mg/kg, s.c.) precedes endotoxin challenge.
males. Specifically, exposure to heroin-paired stimuli is sufficient to elicit suppression of LPS-induced NO production in female rats. Estrous stage had no effect on the heroin-conditioned immune response. These data are the first to establish that naturally cycling female rats exhibit both heroin-induced and -conditioned immunomodulation. This research helps to inform sex-specific interventions to lessen the negative health consequences associated with heroin use.
Introduction
of other pro-inflammatory molecules at the site of infection (Lysle & How, 2000; Bogdan, 2001). In the laboratory we mimic bacterial infection through LPS administration, which readily induces NO production by the enzyme inducible nitric oxide synthase (iNOS). We have reliably demonstrated in male rats, that when heroin precedes LPS administration indices of NO production are blunted. Thus, heroin exposure negatively impacts host immunity and increases susceptibility to disease.
Sex-based Differences in the Opioid and Immune Response
Conventionally, animal models investigating the health consequences associated with opioid use have focused on male subjects. However, women experience distinct, sex-specific risks and responses (both behavioral and physiological) in relation to opioid use (Jamison, Butler, Budman, Edwards & Wasan 2010). The National Health and Nutrition Examination Survey 1999−2012 indicates that women are more likely to be prescribed opioids analgesics than men (7.2% vs 6.3%) and women have been found to be at a greater risk of misusing prescription opioids (Koons, Greenberg, Cannon & Beauchamp, 2018). Furthermore, heroin use has disproportionately increased among women in recent years, with the incidence of heroin use among females in the United States doubling between 2002-2004 and 2011-2013 (Jones, Logan, Gladden & Bohm, 2015). As such, there is a pressing need to examine sex-based differences in opioid use, as there are important clinical ramifications.
Sex differences also extend to the immune response. In general, females exhibit a more
robust innate immune response than males (Schwartz, 2012). For example, in both animal and
clinical models, females consistently exhibit increased glial activation and produce higher levels
of pro-inflammatory cytokines following exposure to an immune challenge such as LPS (Doyle
& Murphy, 2017; Karshikoff et al., 2015). Furthermore, high levels of estrogen have been shown to inhibit interleukin-1β (IL-1β) production following exposure to an LPS immune challenge
(Chakrabarti et al., 2014; Lewis et al., 2008). IL-1β is an important pro-inflammatory cytokine
processes (Goshen et al., 2007). Relevant to the current study, our laboratory has consistently shown the importance of DH IL-1 signaling in the acquisition and expression of heroin-conditioned immunomodulation in male rats (Szczytkowski, Lebonville, Hutson, Fuchs & Lysle, 2013; Paniccia et al., 2018; Lebonville et al., 2016). Specifically, both siRNA mediated knockdown of DH IL-1β gene expression (Szczytkowski et al., 2013) and antagonism of the active IL-1 receptor, IL-1R1 (Paniccia et al, 2018), significantly disrupts the context-heroin association driving the conditioned suppression of peripheral NO production in male rats (Szczytkowski et al., 2013). Presently, it is unknown if females exhibit opioid-induced and conditioned immunomodulation. If they do not, it is possible that estrogen interferes with the integral DH IL-1 signaling necessary for the heroin-conditioned immune response in female rats. Current Study
In Experiment 1 we investigated heroin-induced immunomodulation in male and female rats. We hypothesized that heroin would similarly suppress LPS-induced NO production in both sexes. To investigate this hypothesis, we used a two-factor, independent measures design testing the effect of sex (male or female) and heroin dosage (saline, 1 mg/kg, or 3 mg/kg s.c.) on indices of nitric oxide production following exposure to an immune challenge. Results from Experiment 1 were used to inform conditioning procedures in follow-up experiments.
posited that this effect may be modulated according to the estrous cycle—with heroin-conditioned effects minimized among animals in high estrous stages, such as proestrous or diestrus. High levels of estrogen have been found to inhibit the production of IL-1β, a protein necessary for the expression of heroin-conditioned immunomodulation. In order to test our hypothesis, we used independent measures design in which animals received either heroin (1 mg/ kg, s.c.) or saline in a standard conditioning chamber. On test day, animals were either re-exposed to the CS or remained in home cage as controls. Immediately following CS exposure or home cage stay, animals were challenged with LPS to stimulate an immune response, and tissue was collected 6 hours later. To quantify NO production, we employed real-time RT qPCR to assess splenic iNOS gene expression and the Griess reagent assay to examine plasma nitrate/nitrite levels. To account for estrous phase, we collected vaginal smears at the point of tissue collection.
The current studies aim to fill a major research gap and expand our understanding of how heroin and context-heroin associations drive major health consequences in females. This is the first step required to inform sex-specific treatment interventions for both current and recovering heroin users.
Materials and Methods
Animals
unlimited access to food and water. Rats were handled regularly throughout the five-day habituation period before the conditioning procedure. All animal handling and procedures were conducted in compliance with regulations by the University of North Carolina at Chapel Hill Institutional Animal Care and Use Committee (IACUC).
Drug administration
Lipopolysaccharide (produced from E. coli, serotype O55:B5, Sigma) was dissolved in 0.9% sterile saline to a final concentration of 1 mg/mL. This dose of LPS has been shown to elicit sickness and stimulate NO production (Lysle & Ijames, 2002). Heroin (diacetylmorphine) was obtained from the National Institute on Drug Abuse (Drug Supply Program, Bethesda, MD) and dissolved in 0.9% sterile saline solution to a concentration of 1 mg/ml and 3 mg/mL. These doses are sufficient for robust opioid-induced and -conditioned suppression of NO production (Lysle & How, 2000; Paniccia et al., 2018). To control for heroin administration and repeated injections, conditioning control were given a vehicle of 0.09% sterile saline. All syringes were stored at 4 °C until use. Solutions were allowed to come to room temperature prior to injection. All injections were administered subcutaneously (s.c.).
Procedure
hours later. Spleen tissue was separated into approximately 100 mg for mRNA extraction, and stored in RNAlater (Ambion, ThermoFisher Scientific, Waltham, MA), and blood plasma was collected to assess nitrate/nitrite concentrations.
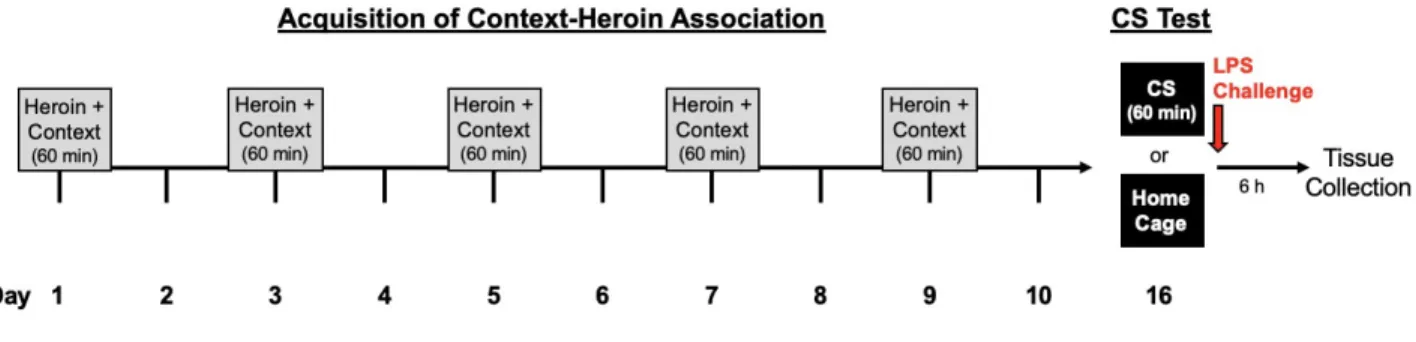
Experiment 2. Using a conditioning procedure commonly employed within our lab, (Szczytkowski et al., 2013; Paniccia et al., 2018), animals were given a subcutaneous injection of heroin (1 mg/kg, s.c., US) or vehicle (saline control) in a standard conditioning chamber (BRS/LVE, Laurel, MD, USA; H 26.7 cm X D 24.1 cm X W 30.5 cm). The conditioning chambers (CS) were distinct from the home cage environment—with metal grid floors and cedar bedding, located in sound attenuation chambers containing a house fan to produce consistent background noise. Importantly, the conditioning chambers were located in a room adjacent from the animal colony. Animals were randomly assigned to heroin or saline treatment group, and all procedures were counterbalanced. Animals received five, 60-minute context-heroin pairings separated by 48 hours (see Figure 1). All experimental procedures occurred in the dark phase of the light-dark cycle, and cues kept constant between sessions.
Figure 1. Our laboratory’s established heroin-conditioning procedure. In this experiment, female rats were either conditioned with heroin or saline during the acquisition phase.
RNA extraction, cDNA synthesis, and Real Time RT-qPCR
cDNA synthesis was done according to the manufacturer’s instructions for the Advantage for RT-PCR Kit (ClonTech, Takara, Mountain View, CA) using the Veriti 96 Well Fast Thermal Cycler (Applied Biosystems, ThermoFisher Scientific). Input RNA was normalized to a 1 g reaction per sample. cDNA was diluted 1:5 for the PCR reaction.
RT-qPCR was performed using the TaqMan™ Fast Advanced Master MixKit (Applied Biosystems, ThermoFisher Scientific) according to the manufacturer’s protocol. Two genes were analyzed using the TaqMan™ Gene Expression Assays (FAM): inducible nitric oxide synthase 2 (iNOS/NOS2, Assay ID: Rn00561646_m1, Thermo- Fisher Scientific), and the reference gene, L13A. Reactions were carried out a 384-well plate in triplicate. Each individual reaction contained 1.5 μL of of sample cDNA, 3 μL of PCR grade water, 0.5 μL of AP BioSystems TaqMan Fast Advanced Master Mix, and 5.0 μL of gene specific Expression Assay Mix. A no template control was run to ensure reaction purity. Plates were run in the QuantStudioTM 6 Flex RealTime PCR System (Applied Biosystems, ThermoFisher Scientific). Data were collected using the QuantStudio™ RealTime PCR Software with a PCR Run Method as follows: 50 °C for 2 min for PCR product contamination degradation, hold at 95 °C for 20 s for polymerase activation, and 45 PCR cycles of 95° C for 1 s and 60° C for 20 s with data collection at the end of each cycle. Data were analyzed using the Comparative CT (ΔΔCT) Method. iNOS was normalized to the reference gene (L13A), and the normalized to the ultimate control (average of all ΔCT values).
Oxygen-exposed NO rapidly degrades into nitrate and nitrite products (Lundberg, Weitzberg, & Gladwin, 2008). Thus, in order to quantify NO production, we assessed
concentrations of NO degradation products using the Griess Reagent Assay (Szczytkowski & Lysle, 2007). We diluted 6 μl of plasma in 44 μl of d H2O, 10 μl of nitrate reductase (1.0 U/ml), 20 μl of a 0.31 M phosphate buffer (pH =7.5), 10 μl of 0.86 mM NADPH (Sigma-Aldrich Inc., Milwaukee, WI), and 10 μl of a 0.11 mM flavin adenine dinucleotide in a 96-well plate. We incubated the mixture in the dark for 90 minutes. Subsequently, we added Griess Reagent (1:1 volume solution of 1% sulfanilamide in 5.0% phosphoric acid and 0.1% N-(1-napthyl)ethyl-enedamine dihydrochloride in distilled water) and let the samples sit for approximately 10 minutes. We compared absorbance values taken at 550 nm to a standard curve to determine concentration.
Estrogen Swabs
Figure 2. Vaginal smears show cell distribution by estrus stage and hormone levels. Pictures taken on Leica DM 6000 microscope. Band hormone levels taken from Miller and Takahashi (2014).
Statistical Analysis
Because nitric oxide rapidly degrades in the plasma when exposed to oxygen, we used two separate measures to asses peripheral NO production: splenic iNOS mRNA expression and plasma nitrate/nitrite. Both nitrate/nitrite concentrations and iNOS CT values were analyzed using analysis of variance (ANOVA) in in SPSS Statistics (IBM, Armonk, NY). A Grubb’s test was used to determine statistically significant outliers within group, which were removed from analyses. A Tukey’s HSD post-hoc test was used to probe significant interactions. Homogeneity of variance was determined using Levene’s Test. The alpha level was set at p = 0.05.
Splenic iNOS:
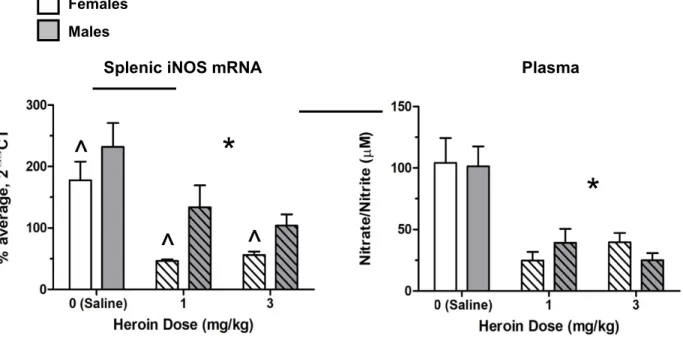
Experiment 1. A 2 X 3 ANOVA of L13A splenic mRNA levels revealed a significant effect of sex on gene expression—with females exhibiting overall higher levels of splenic L13A (F(2, 28) = 4.52, p = 0.042). However, there was no effect of drug on L13A, validating it as an adequate reference gene. Results of a 2 X 3 ANOVA on splenic iNOS mRNA revealed a significant main effect of drug (F (2, 30) = 24.29, p = 0.000) and sex (F (2, 30) = 21.16, p = 0.000). A Tukey’s HSD post-hoc test confirmed that animals administered heroin (either 1 mg/kg or 3 mg/kg) had significantly lower iNOS gene expression relative to saline controls (p<0.05¿. Overall, female rats had significantly lower splenic iNOS mRNA levels than male rats (p < 0.05, see Figure 3).
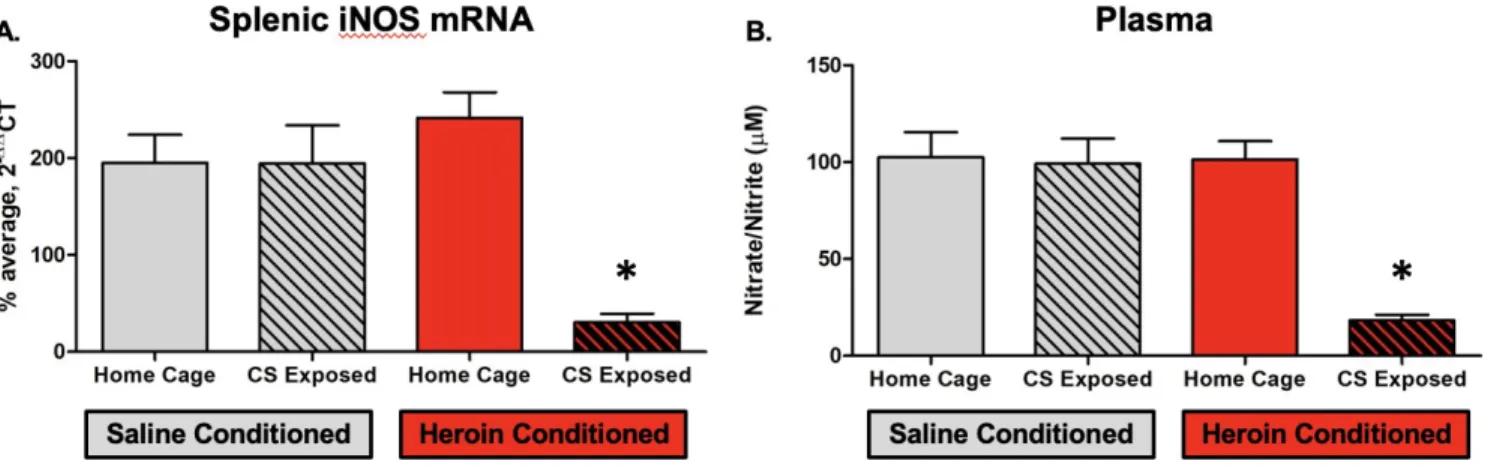
Experiment 2. A 2 X 2 ANOVA of L13A splenic mRNA levels revealed no significant differences between conditions, validating L13A as a reference gene. Comparatively, results of a 2 x 2 ANOVA of iNOS mRNA levels revealed significant main effects of heroin conditioning (F (1, 26) = 14.99, p = 0.001) and CS exposure (F (1, 26) = 31.44, p = 0.000), with a significant interaction between the two variables (F (1, 26) = 17.78, p = 0.000). A Tukey’s HSD post-hoc test confirmed that animals that were heroin-conditioned and CS exposed had significantly lower iNOS gene expression relative to their heroin-conditioned home cage counterparts and saline-conditioned controls (p=0.000¿ (see Figure 4).
Plasma Nitrate/Nitrite:
Splenic iNOS mRNA Plasma
*
^
^
^
*
Females Males
on plasma nitrate/nitrite levels (F (2, 30) = 21.26, p = 0.000). Consistent with our hypothesis, a Tukey’s HSD post-hoc test confirmed that animals administered either dose of heroin followed by LPS had significantly lower plasma nitrate/nitrite levels (p<0.05¿ relative to saline counterparts (see Figure 3).
Experiment 2. Results of a 2 x 2 ANOVA revealed significant main effects of heroin conditioning (F (1, 27) = 14.75, p = 0.001) and CS exposure (F (1, 27) = 16.41, p = 0.000), along with a significant interaction between two variables (F (1, 27) = 13.93, p = 0.001). A Tukey’s HSD post-hoc test confirmed that animals conditioned with heroin and exposed to the CS had significantly lower plasma nitrate/nitrite levels (p=0.000¿ relative to both their heroin-conditioned, home cage counterparts as well as saline conditioned controls (see Figure 4).
Figure 3. Both male and female Lewis rats administered heroin (1 mg/kg or 3 mg/kg) have disrupted NO production to an LPS challenge, as indicated by significantly lower splenic iNOS mRNA expression (A) and plasma nitrate/nitrite levels (B). All group sizes were n = 6. * signifies a main effect of heroin compared to saline, while ^ signifies a main effect of sex.
Figure 4. Heroin-conditioned, CS exposed female Lewis rats exhibit an inhibited immune response to an LPS challenge, as indicated by significantly lower splenic iNOS mRNA expression (A) and plasma nitrate/nitrite levels (B). All group sizes were n = 8. * signifies a statistically significant difference (p < 0.05) from all other groups.
Analysis of Estrous Cycle
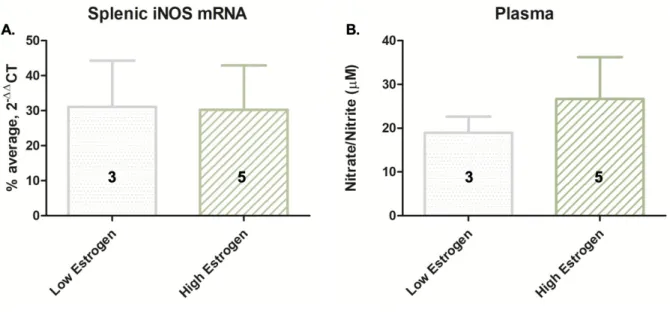
Figure 5. Among heroin-conditioned, CS exposed animals (n = 8), there was no significant difference in splenic iNOS mRNA expression (A) or plasma nitrate/nitrite levels (B) between high estrogen animals (in proestrus or diestrus) and low estrogen animals (in metestrus or estrus). Group sizes are displayed in bar graph.
Figure 6. iNOS mRNA expression (B) and plasma nitrate levels (C) by stage in the estrus cycle across all groups. Splenic iNOS mRNA loosely track with estradiol levels across stage (A). Group sizes are displayed in the bar graph.
Discussion
oxide production in comparably in both male and female rats (Figure 3). Accordingly, we hypothesized that using a 1 mg/kg dose, heroin-conditioned female rats re-exposed to the CS would exhibit conditioned immunosuppression, as indicated by lower levels of peripheral NO production. However, we posited that this effect would be moderated by the high estrogen levels among animals in proestrus and diestrus. Consistent with the primary hypothesis, heroin-conditioned animals exhibited lower levels of LPS-induced NO production relative to controls when exposed to the CS (Figure 4). That is similar to males, naturally cycling female Lewis rats formed the heroin-context association and exhibited conditioned immunosuppression of peripheral NO under the same experimental conditions.
There are numerous factors that may explain why we did not see an effect of estrus stage on peripheral NO production. First, estrogen swabs were collected at six hours following CS or home cage stay. Given that a full cycle occurs in 4-5 days, this variability may have impacted the accuracy of our categorizations (Cora et al., 2015). As the smearing protocol becomes more streamlined in our laboratory, estrous staging determinations will become more accurate. Furthermore, endogenous estrogen levels may not have been high enough to significantly interfere with IL-1β production given that low levels of estrogen do not inhibit IL-1β production (Correale et al., 1998). Further, any modulatory effect of estrogen on IL-1β may have been offset
in females due to higher baseline pro-inflammatory cytokine levels following exposure to an endotoxin challenge with LPS (Doyle &Murphy, 2017; Karshikoff et al., 2015).
Ultimately, further research is necessary to examine potential sex-based differences in the neurobiological mechanisms underlying heroin-conditioned suppression of NO production. In male animal models, our laboratory has established that separately, both intact DH IL-1 and astrocyte signaling are responsible for the context-heroin association driving conditioned peripheral immunomodulation (Szczytkowski et al., 2013; Paniccia et al., 2018). Thus, future studies can investigate the role of DH IL-1 and astrocyte signaling in the heroin-conditioned immune effect in female rats. Importantly, DH astrocytes have been shown to express estrogen receptors, and estradiol has been shown to mediate astrocyte excitability through Ca2+¿−¿¿induced
References
Avital, A., Goshen, I., Kamsler, A., Segal, M., Iverfeldt, K., Richter‐Levin, G., & Yirmiya, R.
(2003). Impaired interleukin‐1 signaling is associated with deficits in hippocampal
memory processes and neural plasticity. Hippocampus, 13(7), 826-834.
Bannerman, D. M., Rawlins, J. N. P., McHugh, S. B., Deacon, R. M. J., Yee, B. K., Bast, T., ... &
Bogdan, C. (2001). Nitric oxide and the immune response. Nature immunology, 2(10), 907.
Chaban, V. V., Lakhter, A. J., & Micevych, P. (2004). A membrane estrogen receptor mediates intracellular calcium release in astrocytes. Endocrinology, 145(8), 3788–3795.
Chakrabarti, M., Haque, A., Banik, N. L., Nagarkatti, P., Nagarkatti, M., & Ray, S. K. (2014). Estrogen receptor agonists for attenuation of neuroinflammation and neurodegeneration. Brain research bulletin, 109, 22-31.
Cora, M. C., Kooistra, L., & Travlos, G. (2015). Vaginal cytology of the laboratory rat and mouse:
review and criteria for the staging of the estrous cycle using stained vaginal smears. Toxicologic pathology, 43(6), 776-793.
Correale, J., Arias, M., & Gilmore, W. (1998). Steroid hormone regulation of cytokine secretion by proteolipid protein-specific CD4+ T cell clones isolated from multiple sclerosis patients
and normal control subjects. The Journal of Immunology, 161(7), 3365-3374.
Dalla, C., & Shors, T. J. (2009). Sex differences in learning processes of classical and operant conditioning. Physiology & Behavior, 97(2), 229–238.
Doyle, H. H., & Murphy, A. Z. (2017). Sex differences in innate immunity and its impact on opioid pharmacology. Journal of neuroscience research, 95(1-2), 487-499.
Frago, L. M., Canelles, S., Freire-Regatillo, A., Argente-Arizón, P., Barrios, V., Argente, J., . . . Chowen, J. A. (2017). Estradiol Uses Different Mechanisms in Astrocytes from the
Hippocampus of Male and Female Rats to Protect against Damage Induced by Palmitic Acid. Frontiers in molecular neuroscience, 10, 330-330. doi:10.3389/fnmol.2017.00330
Goshen, I., Kreisel, T., Ounallah-Saad, H., Renbaum, P., Zalzstein, Y., Ben-Hur, T., ... & Yirmiya, R. (2007). A dual role for interleukin-1 in hippocampal-dependent memory processes. Psychoneuroendocrinology, 32(8-10), 1106-1115.
Jamison, R. N., Butler, S. F., Budman, S. H., Edwards, R. R., & Wasan, A. D. (2010). Gender differences in risk factors for aberrant prescription opioid use. The Journal of Pain, 11(4),
Jones, C. M., Logan, J., Gladden, R. M., & Bohm, M. K. (2015). Vital signs: demographic and substance use trends among heroin users—United States, 2002–2013. MMWR. Morbidity
and mortality weekly report, 64(26), 719.
Jones, M. E., Paniccia, J. E., Lebonville, C. L., Reissner, K. J., & Lysle, D. T. (2018). Chemogenetic manipulation of dorsal hippocampal astrocytes protects against the development of stress-enhanced fear learning. Neuroscience, 388, 45-56.
Karshikoff, B., Lekander, M., Soop, A., Lindstedt, F., Ingvar, M., Kosek, E., ... & Axelsson, J. (2015). Modality and sex differences in pain sensitivity during human endotoxemia. Brain, behavior, and immunity, 46, 35-43.
Koons, A. L., Greenberg, M. R., Cannon, R. D., & Beauchamp, G. A. (2018). Women and the experience of pain and opioid use disorder: a literature-based commentary. Clinical therapeutics, 40(2), 190-196.
Lebonville, C. L., Jones, M. E., Hutson, L. W., Cooper, L. B., Fuchs, R. A., & Lysle, D. T. (2016).
Acquisition of heroin conditioned immunosuppression requires IL-1 signaling in the dorsal hippocampus. Brain, behavior, and immunity, 56, 325-334.
Lewis, D. K., Johnson, A. B., Stohlgren, S., Harms, A., & Sohrabji, F. (2008). Effects of estrogen
receptor agonists on regulation of the inflammatory response in astrocytes from young adult and middle-aged female rats. Journal of neuroimmunology, 195(1-2), 47-59.
Lysle, D. T., & Ijames, S. G. (2002). Heroin-associated environmental stimuli modulate the expression of inducible nitric oxide synthase in the rat. Psychopharmacology, 164(4), 416-422.
Lysle, D. T., & How, T. (2000). Heroin modulates the expression of inducible nitric oxide synthase. Immunopharmacology, 46(3), 181-192.
Lundberg, J. O., Weitzberg, E., & Gladwin, M. T. (2008). The nitrate–nitrite–nitric oxide pathway
in physiology and therapeutics. Nature reviews Drug discovery, 7(2), 156.
McCarthy, L., Wetzel, M., Sliker, J. K., Eisenstein, T. K., & Rogers, T. J. (2001). Opioids, opioid
receptors, and the immune response. Drug and alcohol dependence, 62(2), 111-123. Menachem-Zidon, O. B., Avital, A., Ben-Menahem, Y., Goshen, I., Kreisel, T., Shmueli, E. M.,
... & Yirmiya, R. (2011). Astrocytes support hippocampal-dependent memory and long-term potentiation via interleukin-1 signaling. Brain, behavior, and immunity, 25(5),
1008-1016.
Paniccia, J. E., Lebonville, C. L., Jones, M. E., Parekh, S. V., Fuchs, R. A., & Lysle, D. T. (2018). Dorsal hippocampal neural immune signaling regulates heroin-conditioned immunomodulation but not heroin-conditioned place preference. Brain, behavior, and immunity, 73, 698-707
Schwarz, J. M., & Bilbo, S. D. (2012). Sex, glia, and development: interactions in health and disease. Hormones and behavior, 62(3), 243-253.
Straub, R. H. (2007). The complex role of estrogens in inflammation. Endocrine reviews, 28(5), 521-574.
Srinivasan, D., Yen, J. H., Joseph, D. J., & Friedman, W. (2004). Cell type-specific interleukin-1β
signaling in the CNS. Journal of Neuroscience, 24(29), 6482-6488.
Szczytkowski, J. L., Lebonville, C., Hutson, L., Fuchs, R. A., & Lysle, D. T. (2013). Heroin-induced conditioned immunomodulation requires expression of IL-1β in the dorsal hippocampus. Brain, behavior, and immunity, 30, 95-102.
Szczytkowski, J. L., & Lysle, D. T. (2007). Conditioned effects of heroin on the expression of inducible nitric oxide synthase in the rat are susceptible to extinction and latent inhibition. Psychopharmacology, 191(4), 879-889.
Todd, C. S., Abed, A. M., Strathdee, S. A., Scott, P. T., Botros, B. A., Safi, N., & Earhart, K. C. (2007). HIV, hepatitis C, and hepatitis B infections and associated risk behavior in injection drug users, Kabul, Afghanistan. Emerging infectious diseases, 13(9), 1327.