3039
Background and Purpose—The complexity and heterogeneity of stroke, as well as the associated comorbidities, may render neuroprotective drugs less efficacious in clinical practice. Therefore, the development of targeted therapies to specific patient subsets has become a high priority in translational stroke research. Ischemic stroke with type 2 diabetes mellitus has a nearly double mortality rate and worse neurological outcomes. In the present study, we tested our hypothesis that rFGF21 (recombinant human fibroblast growth factor 21) administration is beneficial for improving neurological outcomes of ischemic stroke with type 2 diabetes mellitus.
Methods—Type 2 diabetes mellitus db/db and nondiabetic genetic control db/+ mice were subjected into permanent focal ischemia of distal middle cerebral artery occlusion, we examined the effects of poststroke administration with rFGF21 in systemic metabolic disorders, inflammatory gatekeeper PPARγ (peroxisome proliferator-activated receptor γ) activity at 3 days, mRNA expression of inflammatory cytokines and microglia/macrophage activation at 7 days in the perilesion cortex, and last neurological function deficits, ischemic brain infarction, and white matter integrity up to 14 days after stroke of db/db mice.
Results—After permanent focal ischemia, diabetic db/db mice presented confounding pathological features, including metabolic dysregulation, more severe brain damage, and neurological impairment, especially aggravated proinflammatory response and white matter integrity loss. However, daily rFGF21 treatment initiated at 6 hours after stroke for 14 days significantly normalized systemic metabolic disorders, rescued PPARγ activity decline, inhibited proinflammatory cytokine mRNA expression, and M1-like microglia/macrophage activation in the brain. Importantly, rFGF21 also significantly reduced white matter integrity loss, ischemic brain infarction, and neurological function deficits up to 14 days after stroke. The potential mechanisms of rFGF21 may in part consist of potent systematic metabolic regulation and PPARγ-activation promotion-associated antiproinflammatory roles in the brain.
Conclusions—Taken together, these results suggest rFGF21 might be a novel and potent candidate of the disease-modifying strategy for treating ischemic stroke with type 2 diabetes mellitus.
Visual Overview—An online visual overview is available for this article. (Stroke. 2018;49:3039-3049. DOI: 10.1161/ STROKEAHA.118.022119.)
Key Words: diabetes mellitus ◼ inflammation ◼ mice ◼ stroke ◼ white matter
© 2018 American Heart Association, Inc.
T
he complexity and heterogeneity of stroke, as well as the associated comorbidities, may render neuroprotective drugs less efficacious in clinical practice.1 Therefore,devel-opment of targeted therapies to specific patient subsets has be-come a high priority in translational stroke research.2 Diabetic
patients are 2 to 6× more susceptible to ischemic stroke; ≈30% of stroke patients are diabetic and >90% of them comprise type 2 diabetes mellitus (T2D).3 T2D stroke patients have
nearly double mortality rates, worse neurological outcomes, and respond less favorably to tPA (tissue-type plasminogen
Fibroblast Growth Factor 21) Improves Neurological
Outcomes Following Focal Ischemic Stroke of Type 2
Diabetes Mellitus Male Mice
Yinghua Jiang, MD; Ning Liu, PhD; Qingzhi Wang, MD; Zhanyang Yu, PhD; Li Lin, PhD;
Jing Yuan, MD; Shuzhen Guo, PhD; Bum Ju Ahn, PhD; Xiao-Jie Wang, PhD; Xiaokun Li, MD;
Eng H. Lo, PhD; Xiaochuan Sun, MD; Xiaoying Wang, PhD
DOI: 10.1161/STROKEAHA.118.022119
Stroke is available at https://www.ahajournals.org/journal/str
Received May 14, 2018; final revision received September 30, 2018; accepted October 10, 2018.
From the Department of Neurosurgery, The First Affiliated Hospital of Chongqing Medical University, China (Y.J., X.S., ); Neuroprotection Research Laboratory, Departments of Radiology and Neurology, Massachusetts General Hospital and Harvard Medical School, Boston (Y.J., N.L., Q.W., Z.Y., L.L., J.Y., S.G., B.J.A., E.H.L., X.W.); The Third Affiliated Hospital of Zhengzhou University, China (N.L.); Department of Neurology, The First Affiliated Hospital of Zhengzhou University, China (Q.W., J.Y.); and Key Laboratory of Biotechnology and Pharmaceutical Engineering, School of Pharmaceutical Sciences, Wenzhou Medical University, Zhejiang, China (L.L., X.-J.W., X.L.).
The online-only Data Supplement is available with this article at https://www.ahajournals.org/doi/suppl/10.1161/STROKEAHA.118.022119. Correspondence to Xiaochuan Sun, MD, Department of Neurosurgery, The First Affiliated Hospital of Chongqing Medical University, No. 1 Youyi Road, Yuzhong District, Chongqing 400016, China, Email [email protected] or Xiaoying Wang, PhD, Neuroprotection Research Laboratory, Departments of Radiology and Neurology, Massachusetts General Hospital, 149 13th St, Charlestown, MA 02129, Email [email protected]
activator) therapy because of the higher risk of hemorrhagic transformation.4,5
All diabetic stroke patients present poststroke hyper-glycemia and other metabolic disorders.6 From a
transla-tional perspective, development of new compounds with multiple pharmacological functions, particularly targeting T2D stroke-related metabolic dysregulation, ischemic brain damage, and detrimental proinflammation, called disease-modifying approaches, would be novel and more effective for treating ischemic stroke with T2D.7 FGF21 (fibroblast
growth factor 21) might be one of the optimal therapeutic candidates.
FGF21 is an endocrine member of the FGF family. It has a potent and central role in glucose and lipid metabo-lism, as well as in energy balance.8 Human FGF21
(molec-ular weight, 19.5 kD) is highly homologous to mouse FGF21 (≈75% identity).9 The very low heparin-binding affinity
makes FGF21 capable of crossing the blood-brain barrier by simple diffusion.10 FGF21 exerts potent and multipleiotropic
metabolic actions,11,12 with benefits that have been translated
from rodents to obese humans with T2D without causing side effects, such as mitogenicity and hypoglycemia.13,14
Growing experimental findings have demonstrated FGF21 is also a mediator of adaptive responses to tissue injury and repair in various pathological conditions.12,15,16 In the present
study, we propose that FGF21 is a novel therapeutic candidate with both adaptive and protective pharmacological actions that protect against both metabolic and stroke stresses in T2D, that result in improvement of neurological outcomes. Using T2D animal model, db/db mice, and nondiabetic ge-netic control db/+ mice, we tested our hypothesis that rFGF21 (recombinant human FGF21) administration is beneficial for improving neurological outcomes of T2D stroke mice. Our experimental results are highly supportive to the hypothesis and demonstrate that rFGF21 might be developed as a novel and potent disease-modifying approach for treating ischemic stroke with T2D.
Methods
Details of materials and experimental procedures are available in the
online-only Data Supplement. The data that support the findings of this study are available from the corresponding author on reasonable request.
Focal Ischemia Stroke Animal Model and Experimental Groups
Ten weeks old male BKS.Cg-Dock7m +/+ Lepr db/J homozy-gous (db/db) mice, weighing 40 to 50 g, and their age-matched nondiabetic counterparts BKS-Cg-Dock7m +/+ Lepr db/J heter-ozygous (db/+) mice, weighing 25 to 30 g, were purchased from Jackson Laboratory. Focal ischemic stroke was achieved with coagulation of the distal middle cerebral artery, followed by 90 minutes bilateral common carotid artery ligation as previously
described with slight modifications.17 Animals were
subcutane-ously administrated with either 1.5 mg/kg recombinant FGF21 or saline initiated at 6 hours after stroke, followed by twice daily subcutaneous injections (1.5 mg/kg per time) with an interval of 10 to 12 hours for up to 14 days. All experiments were performed after protocols approved by the Massachusetts General Hospital Institutional Animal Care and Use Committee in compliance with the NIH Guide for the Care and Use of Laboratory Animals. All
experimental assessments were performed by investigators who were blind to the experimental groups.
Measurements of Body Weight, Blood Glucose, HbA1c (Glycated Hemoglobin), Insulin, and Adiponectin Levels
Details are provided in the online-only Data Supplement.
Neurobehavioral Assessments
The following sensorimotor function deficits were assessed before and poststroke day 1, 3, 5, 7, and 14. (1) Adhesive removal test result was represented by the time-to-contact and time-to-remove of each
forepaw that was recorded, respectively as previously described.18
(2) Grip strength test result was quantified as percentage of each
individual preischemic baseline as previously described.19 (3) Foot
fault test result was quantified as percentage of foot faults referring
to the total steps as previously described.20 Additionally, cognitive
function deficit was assessed at poststroke day 14 by Y-maze test and quantified with the percentage of alternations as we previously
described.21
Reverse Transcription and Real-Time Polymerase Chain Reaction Assay
Peri-infarct cortical tissues or corresponding cortical tissues in sham-operated group were collected at 7 days after stroke and stored at −80°C until processing. Total RNA was extracted using RNeasy Lipid Tissue Mini Kit (Qiagen), and real-time polymerase chain re-action was performed as we previously described. Further details are
provided in the online-only Data Supplement.
Measurement of PPARγ (Peroxisome
Proliferator-Activated Receptor γ) Activity With Electrophoretic Mobility Shift Assay (EMSA)
We performed electrophoretic mobility shift assay by following a
standard protocol as we previously described.22 Further details are
provided in the online-only Data Supplement.
Measurements of Brain Infarction and Axon Myelination Level
At 14 days after stroke, brain sections were stained with hematox-ylin & eosin for quantifying ischemic brain infarction size as we
pre-viously described.23 Axon myelination level was detected with the
luxol fast blue staining and quantified as previously described with
slight modification.24 Briefly, myelination levels of external capsule
were evaluated using quantitative scoring method. It assigned 4 for mice with the sham operation (having normal myelin levels in ex-ternal capsule area), whereas a fully demyelinated exex-ternal capsule area was given a score of 0. Sections were measured and scored by 2 independent individuals who were blinded to the experimental group assignment.
Immunohistochemistry Analysis
Immunohistochemistry was conducted using the standard protocol as
we described previously.21 Further details are provided in the
online-only Data Supplement.
Statistical Analysis
All data are expressed as mean±SEM. For parametric and continuous variable measurements, we use ANOVA followed by Tukey-Kramer post hoc tests. For nonparametric ordinal data such as neurobehav-ioral tests, we use nonparametric Kruskal-Wallis followed by post
hoc Mann-Whitney U test tests. Overall, P<0.05 was considered
significant.
Results
rFGF21 Redresses Hyperglycemia, Insulin Resistance, and Adiponectin Decline in Diabetic db/db Mice After Focal Stroke
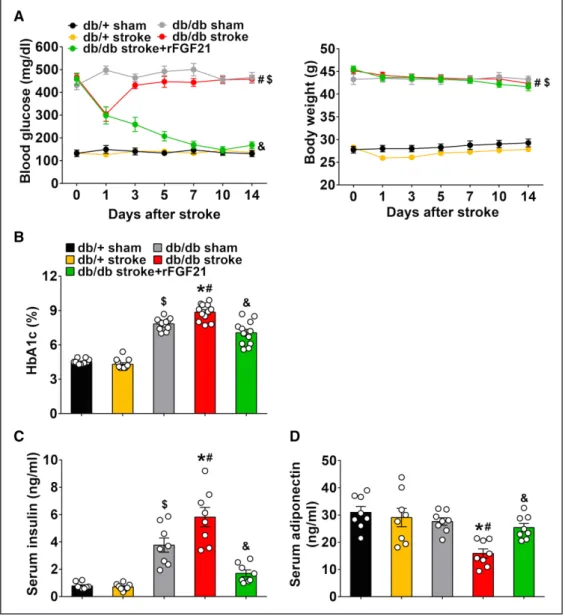
After a stroke, db/db mice maintained stable hyperglycemia (but body weight was not altered), had an elevated blood HbA1c level, hyperinsulinemia, and lowered serum adiponec-tin level (Figure 1). However, rFGF21 treatment significantly reduced blood glucose level by ≈63% reduction (Figure 1A) and lowered HbA1c level from 8.9% to 7.1% (Figure 1B). The poststroke hyperinsulinemia was also significantly di-minished by rFGF21 treatment (71% reduction; Figure 1C). Moreover, the stroke-induced lower serum adiponectin level was significantly rescued by rFGF21 treatment (60% increase; Figure 1D). These data suggest that rFGF21 has potent ther-apeutic effects for metabolic improvement in hyperglycemia, poststroke hyperinsulinemia, and hypoadiponectinemia of
db/db stroke mice. In a separate experiment, we also tested rFGF21 effects in nondiabetic db/+ mice. There was no sig-nificant difference in both blood glucose levels and body weight of nondiabetic db/+ mice after ischemic stroke be-tween rFGF21 treated and nontreated group, and rFGF21 did not cause hypoglycemia, indicating a relative safe profile of rFGF21 administration after stroke (Figure I in the online-only Data Supplement).
rFGF21 Attenuates Neurological Function Deficits and Reduced Ischemic Brain Lesion Size in Diabetic db/db Mice After Focal Stroke
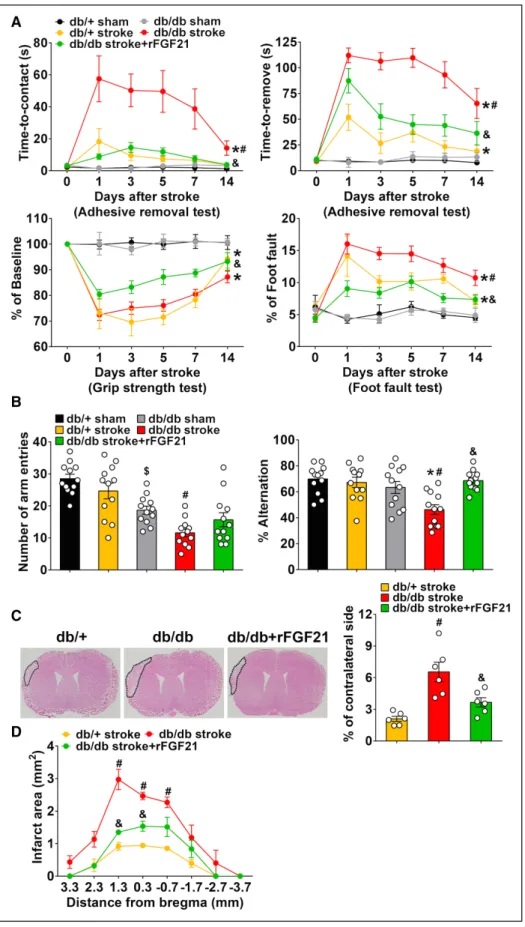
As expected, both db/+ stroke mice and db/db stroke mice showed significant deficits up to 14 days after stroke. The sensorimotor function deficits in db/db stroke mice were significantly worse than the nondiabetic db/+ stroke mice. Most importantly, rFGF21 treatment dramatically attenuated
Figure 1. rFGF21 (recombinant human fibroblast growth factor 21) redresses hyperglycemia, insulin resistance, and adiponectin decline in diabetic db/db mice after focal stroke. Mice were continuously monitored for 14 d after stroke. A, Blood glucose levels and body weight changes. B, Blood HbA1c (glycated hemoglobin) levels. C, Serum insulin concentrations. D, Serum adiponectin concentrations. Data are expressed as mean±SE, n=8 per group (serum insulin and adiponectin) or 12 per group (blood glucose, HbA1c, and body weight). *P<0.05 stroke group vs sham control group; #P<0.05 db/db stroke group vs db/+ stroke group; &P<0.05 db/db stroke+rFGF21 group vs db/db stroke group; $P<0.05 db/+ sham group vs db/db sham group.
Figure 2. rFGF21 (recombinant human fibroblast growth factor 21) attenuates neurological function deficits and brain infarct size in diabetic db/db mice after focal stroke. A, Sensorimotor functions were assessed at 0, 1, 3, 5, 7, and 14 d poststroke. Adhesive removal test consists of time-to-contact the adhesive tape (seconds) and time-to-remove the adhesive tape (seconds). Other 2 tests were grip strength test and foot fault test. B, Y-maze test was performed at 14 d poststroke, the results were recorded as total number of arm entries and alternations. Data are expressed as mean±SE, n=12 per group. C, At 14 d after stroke, representative H&E staining images of coronal sections from db/+ mice, db/db mice, and db/db mice treated with rFGF21 that shows infarct area delineated by black dashed lines, and the infarct volume was quantified. D, Infarct area in 8 consecutive coronal sections with 1 mm interval. Data are expressed as mean±SE, n=6 per group. *P<0.05 stroke vs sham; #P<0.05 db/db stroke vs db/+ stroke; &P<0.05 db/db stroke+rFGF21 vs db/db stroke.
deficits in all 3 sensorimotor function tests of db/db stroke mice (Figure 2A).
The db/db sham mice were prone to motionlessness or moved slowly in Y-maze test. At 14 days after stroke, the fur-ther reduced total number of arm entries and lowered alter-nation ratio in db/db mice were all significantly rescued by rFGF21 treatment (Figure 2B). These results suggest a potent therapeutic potential of rFGF21 treatment for improving neu-rological functions after ischemic stroke in T2D.
Ischemic brain infarct size was measured at 14 days after stroke. The db/db stroke mice had a significantly larger in-farct (6.6% of contralateral side) compared with the db/+ stroke mice (2.1% of contralateral side), excitingly rFGF21 significantly reduced infarct size of db/db stroke mice (44.1%
reduction; Figure 2C). We also found the third coronal section (+1.3 mm from bregma) represented the location of maximal cortical change in infarction after rFGF21 treatments; thus the third coronal section was selected for histological analysis in this study (Figure 2D). There was no mortality in all stroke groups.
We also tested the effects of rFGF21 in neurological out-comes and ischemic brain lesion size of nondiabetic db/+ mice. rFGF21 treatment did not alter neurological functions up to 14 days after stroke (Figure IIA in the online-only Data Supplement), while slightly reduced brain infarction size in nondiabetic db/+ mice (12.5% reduction), but it was not statis-tically significant (Figure IIB and IIC in the online-only Data Supplement).
rFGF21 Enhances Nuclear Transcriptional Factor PPARγ DNA-Binding Activity in Diabetic db/db Mice After Focal Stroke
The PPARγ DNA-binding activity was significantly decreased at 3 days after stroke in both db/+ stroke (36.6% reduc-tion) and db/db stroke mice (41.1% reducreduc-tion). Importantly, rFGF21 significantly promoted the PPARγ DNA-binding ac-tivity of db/db mice (34% increase; Figure 3). These results demonstrate the dampened PPARγ activity in db/db mouse brains after stroke can be significantly elevated by poststroke administration of rFGF21.
rFGF21 Suppresses Proinflammatory Cytokine Expression in Perilesion Cortex of Diabetic db/db Mice After Focal Stroke
At 7 days after stroke, mRNA expression of inflammatory genes in the cortical perilesion area was examined by real-time polymerase chain reaction. We did not detect any sig-nificant differences between db/+ sham and db/db sham mice (Figure 4). However, in 4 selected proinflammatory genes, ischemic stroke only significantly increased TNF-α (tumor necrosis factor-α) and CCL (C-C motif chemokine ligand) 3 mRNA expression in db/+ mice. While in db/db stroke mice, all 4 proinflammatory genes TNF-α, IL (interleukin)-1β, TLR (Toll-like receptor) 4, and CCL3 mRNA expressions exhibited significantly higher levels, suggesting more dominant proin-flammatory response in the ischemic brain of db/db stroke mice. Importantly, rFGF21 treatment significantly reduced the stroke-elevated proinflammatory gene mRNA expressions, demonstrating a potent inhibiting effect for proinflammation in the ischemic brain of T2D db/db mice (Figure 4A). In con-trast, ischemic stroke significantly increased 4 anti-inflamma-tory cytokines or trophic factors mRNA levels for IL-4, IL-10, IGF (insulin like growth factor) 1, and TGF (transforming growth factor)-β in both db/+ and db/db mice. Interestingly, there was a significant lower mRNA expression of IL-4 and IL-10 in the db/db stroke mice compared with db/+ stroke mice. Importantly, rFGF21 treatment to the db/db stroke mice significantly upregulated all 4 selected mRNA expressions of IL-4, IL-10, IGF-1, and TGF-β (Figure 4B). These data sug-gested there might be an impairment of anti-inflammatory and neurorepair responses in the db/db mouse brains after stroke, which may be significantly restored by rFGF21 treatment. Figure 3. rFGF21 (recombinant human fibroblast growth factor 21)
enhances nuclear transcriptional factor PPARγ (peroxisome proliferator-activated receptor γ) DNA-binding activity in diabetic db/db mice after focal stroke. At 3 d poststroke, DNA-binding activity of transcriptional fac-tor PPAR-γ was measured by electrophoresis mobility shift assay (EMSA) in nuclear fractions. A, Representative image of EMSA gel. Loaded sam-ples were lane 1–3: db/+ sham, lane 4: db/+ stroke, lane 5: db/db sham, lane 6: db/db stroke, and lane 7: db/db stroke plus rFGF21. B, Quantifi-cation by densitometry of specific PPARγ DNA-binding bands. Data are expressed as mean±SE, n=7 per group. *P<0.05 stroke vs sham; &P<0.05 db/db stroke+rFGF21 vs db/db stroke.
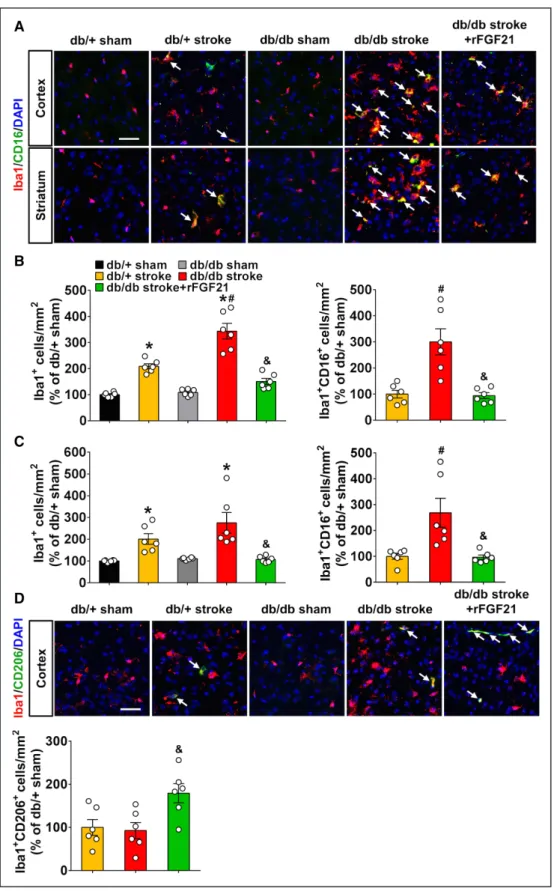
rFGF21 Suppresses Proinflammatory Microglia/ Macrophage Activation in Perilesion Area of Diabetic db/db Mice After Focal Stroke
At 7 days after stroke, immunohistochemistry was performed to determine microglia/macrophage activation by counting Iba-1 positive cells, and M1-like proinflammatory microglia/ macrophage by counting Iba-1/CD16 double positive cells. In the perilesion cortex, ischemic stroke significantly increased the Iba-1 positive cells and Iba-1/CD16 double positive cells; this increase was significantly higher in db/db mice than db/+ mice. However, rFGF21 almost completely eliminated the increases (Figure 5A and 5B) In the subcortex striatum areas, Iba-1 positive cell number was significantly increased in both db/+ and db/db stroke mice, but significantly higher
Iba-1/CD16 double positive cells were only detected in db/ db stroke mice. The increase in Iba-1/CD16 double posi-tive cells can be significantly reduced by rFGF21 treatment (Figure 5C). Next, we determined M2-like anti-inflammatory microglia/macrophage by counting Iba-1/CD206 double pos-itive cells. There were very low levels of Iba-1/CD206 double positive cells, and no significant differences could be detected between groups. However, rFGF21 treatment significantly increased the Iba-1/CD206 double positive cells in perilesion cortex of db/db stroke mice (Figure 5D). These results further demonstrated a distressed proinflammatory response in the brain of db/db stroke mice; suppressing M1-like microglia/ macrophage activation might be one of the anti-detrimental neuroinflammation mechanisms of rFGF21.
Figure 4. rFGF21 (recombinant human fibroblast growth factor 21) suppresses proinflammatory cytokine expression in perilesion cortex of diabetic db/ db mice after focal stroke. Peri-infarct tissues were collected at 7 d poststroke for real-time polymerase chain reaction assay. A, Relative fold changes of proinflammatory cytokines/chemokines mRNA expression including TNF-α (tumor necrosis factor-α), IL (interleukin)-1β, TLR (Toll-like receptor) 4, and CCL (C-C motif chemokine ligand) 3. B, Relative fold change of anti-inflammatory cytokines/tropic factors mRNA expression including IL-4, IL-10, IGF (insulin like growth factor) 1, and TGF (transforming growth factor)-β. Data are expressed as mean±SE, n=5 per group. *P<0.05 stroke vs sham; #P<0.05 db/db stroke vs db/+ stroke; &P<0.05 db/db stroke+rFGF21 vs db/db stroke.
Figure 5. rFGF21 (recombinant human fibroblast growth factor 21) suppresses proinflammatory microglia/macrophage activation in perilesion area of diabetic db/ db mice after focal stroke. Mice were sacrificed at 7 d poststroke and brain sections were stained for Iba-1 or Iba-1/CD16 double or Iba-1/CD206 double positive cells. A, In cortical and striatal peri-infarct area, representative immunofluorescence images of Iba-1 and CD16 double positive cells. B, Number of Iba-1 positive (Iba1+) or Iba-1 and CD16 double positive cells (Iba1+ CD16+) in cortical peri-infarct area were quantified as % of db/+ sham control mouse brains per square
mil-limeter. C, Numbers of Iba-1 positive (Iba1+) or Iba-1 and CD16 double positive cells (Iba1+ CD16+) in striatal peri-infarct area were quantified as % of db/+ sham
control mouse brains per square millimeter. D, In cortical peri-infarct area, representative immunofluorescence images of Iba-1 and CD206 double positive cells. Numbers of Iba-1 and CD206 double positive cells (Iba1+ CD206+) were quantified as % of db/+ sham control mouse brains per square millimeter. Merged images
under high magnification represent colocalization (white arrow) of Iba-1 and CD16/32 or CD206, respectively. Scale bar=50 μm. Data are expressed as mean±SE, n=6 per group. *P<0.05 stroke vs sham; #P<0.05 db/db stroke vs db/+ stroke; &P<0.05 db/db stroke+rFGF21 vs db/db stroke.
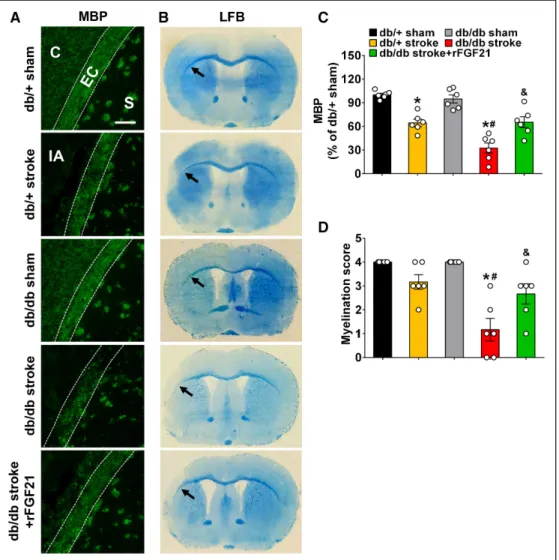
rFGF21 Ameliorates White Matter Injury in Diabetic db/db Mice After Focal Stroke
White matter integrity after stroke was examined at 14 days after stroke. Axonal injury was examined by immunohis-tochemistry of MBP (myelin basic protein) expression, and myelination score was determined by luxol fast blue staining at perilesion external capsule. Ischemic stroke significantly decreased more MBP expression (MBP loss) in db/db mice than db/+ mice, however, we only detected a significant de-crease of myelination score in db/db stroke mice. Importantly, rFGF21 significantly rescued MBP expression decrease (Figure 6A and 6C) and elevated myelination score in db/ db stroke mice (Figure 6B and 6D). These results suggest an aggravated white matter integrity loss in db/db stroke mice, where rFGF21 significantly promoted white matter integrity by attenuating the axonal injury while promoting myelination, which might be in part responsible to the improved neurolog-ical outcomes.
Discussion
The major findings of the present experiments can be summa-rized that (1) compared with nondiabetic db/+ mice, diabetic db/db mice presented some of confounding pathological fea-tures of ischemic stroke, including metabolic dysregulation, more severe brain damage, and neurological impairment, and in particular aggravated proinflammatory response and white matter integrity loss. (2) The db/db mice that received rFGF21 at 6 hours after stroke for 14 days showed significantly nor-malized systemic metabolic disorders, rescued anti-inflamma-tory gatekeeper PPARγ activity, inhibited proinflammatory cytokines mRNA expression and microglia/macrophage acti-vation after stroke in the perilesion cortex. (3) The rFGF21 ad-ministration also significantly reduced white matter integrity loss, ischemic brain infarction, and neurological function defi-cits up to 14 days after stroke. These results demonstrate that rFGF21 might be a novel and potent candidate of the disease-modifying approach for treating ischemic stroke with T2D.
Figure 6. rFGF21 (recombinant human fibroblast growth factor 21) ameliorates white matter injury in diabetic db/db mice after focal stroke. Mice were sac-rificed at 14 d poststroke and coronal brain sections were stained for MBP (myelin basic protein) expression by immunohistochemistry or stained with Luxol fast blue dye. A, Representative MBP immunofluorescence images. Loss of MBP displayed as dark area within the external capsule area adjacent to infarct core. B, Representative images of Luxol fast blue (LFB) stained brain sections. Black arrow indicates the myelination of external capsule after stroke in each group of mice. C, Quantification of MBP fluorescence intensity within the external capsule area adjacent to the infarct core. MBP levels were expressed as % of average fluorescence intensity of db/+ sham mice. Scale bar=20 μm. D, Myelination levels were semiquantified as myelination score. Data are expressed as mean±SE, n=6 per group. *P<0.05 stroke vs sham; #P<0.05 db/db stroke vs db/+ stroke; &P<0.05 db/db stroke+rFGF21 vs db/db stroke. C indicates cortex; EC, external capsule (the area between 2 dashed white lines); IA, infarct area; and S, striatum.
The most unique biological feature of FGF21 is its ability to exert potent and multipleiotropic metabolic effects,11,12 such
as improvements in hyperglycemia, insulin resistance, lipid profiles, and adiponectin production in obese and diabetic animals.25–27 We found db/db stroke mice presented
hyper-glycemia, elevated poststroke hyperinsulinemia, and hypoa-diponectinemia, each of these metabolic disorders has been demonstrated independently to be associated with poor clin-ical outcome after stroke.28 Importantly, rFGF21 treatment,
even when initiated at 6 hours after stroke remarkably elimi-nated these metabolic dysregulations. These potent metabolic modulation effects of rFGF21 may be one of its beneficial mechanisms for improving neurological outcomes after stroke in the db/db mice.
It is important to determine whether this endocrine regu-lator rFGF21 is beneficial or might cause detrimental hypogly-cemia to the ischemic stroke control of nondiabetes mellitus. Experimental results showed there were no hypoglycemia and body weight decline during the 14 days treatment period of rFGF21 after stroke. Furthermore, rFGF21 treatment did not significantly alter neurological functions but slightly reduced brain infarct size. These results suggest that rFGF21 is rela-tively safe and does not induce hypoglycemia. However, the neutral results of rFGF21 treatment in long-term outcomes of nondiabetes mellitus stroke mice might be in part because of the relatively smaller infarction and mild neurological deficits of the distal middle cerebral artery occlusion model, and rel-atively smaller sample size applied in the current study.29 The
precise roles and mechanisms of rFGF21 in ischemic stroke outcomes of nondiabetes mellitus need to be fully investigated in future experiments, including other ischemic stroke animal models. Importantly, in contrast to the protective effects of rFGF21 in T2D db/db stroke mice, these results obtained from nondiabetic mice are also supportive to our hypothesis that rFGF21 is a novel and specific disease-modifying approach for treating ischemic stroke with T2D.
We next explored the effects and potential mechanisms of rFGF21 in the brain of diabetic stroke mice. It has been known that a very low heparin-binding affinity makes FGF21 capable of crossing the blood-brain barrier by simple diffusion.10 Our
previous study showed subcutaneous injection with rFGF21 significantly increased its concentration in cerebrospinal fluid, and phospho-FGFR (fibroblast growth factor receptor) 1 lev-els in mouse brain tissues, demonstrating rFGF21 could cross blood-brain barrier and functionally activate FGF21-FGFR1 signaling in the brain.21 Interestingly, FGF21 has been
re-ported as a key mediator involving in the physiological and pharmacological actions of PPARγ.30 PPARγ is a nucleus
re-ceptor that transcriptionally regulates gene expressions, espe-cially functions as a master gatekeeper of cytoprotective stress response in brain injury and repair after stroke mainly by sup-pressing proinflammatory and neurodegenerative responses.31
Because ischemic stroke in diabetic animals may exacerbate or sustain the proinflammatory response,5,32 thus we measured
early transcriptional regulator PPARγ activity at 3 days after stroke, yet examined its downstream signaling pathways mi-croglia/macrophage activation, and expression of inflamma-tory factors in the brain at late 7 days after stroke in T2D db/ db mice. However, the precise temporal causal relationship
between PPARγ activity and inflammatory factor expression remains to be carefully elucidated.
We found a significant decrease of PPARγ DNA-binding activity at 3 days after stroke in perilesion cortex of T2D mice, but it was significantly elevated by poststroke admin-istration of rFGF21. Although as a transcriptional factor, how the rFGF21-activated PPARγ regulate its targeting signaling pathways remain to be elucidated; accumulating experimental evidence have supported the critical roles of PPARγ activity for improving the chances of cellular survival and recovery of homeostatic equilibrium after ischemic stroke.31
PPARγ functions as a major transcriptional factor that inhibits proinflammatory responses after ischemic stroke.31
Experimental evidence has demonstrated that in acute and subacute phase after ischemic stroke, the higher proinflamma-tory or lower anti-inflammaproinflamma-tory cytokine expression profiles represent a relatively more detrimental environment in the ischemic brain that might contribute to the secondary brain tissue damage and neurorepair impairment after stroke.32 Our
experimental data showed highly elevated M1-like proinflam-matory microglia/macrophage activation in the brain of T2D db/db mice after stroke, which is consistent with the increased mRNA expression of proinflammatory cytokines. Importantly, the potent effects of rFGF21 on inhibition of proinflamma-tory cytokine mRNA expression and M1-like microglia/mac-rophage activation, and elevation of anti-inflammatory gene mRNA expression, facilitate a strong antineuroinflamma-tion role, and may be one of key underlying mechanisms for improving neurological outcomes in T2D stroke. The antineu-roinflammation action of rFGF21, might be in part because of PPARγ activation promotion.
However, we can not exclude the possibility that rFGF21 might be able to suppress systemic inflammation and brain infiltration of inflammatory cells in diabetic stroke mice, thereby contribute to the promotion of neurological function recovery. Since in our previously published study, we showed rFGF21administration significantly reduced mRNA levels of proinflammatory cytokines IL-1β, IL-6, and TNF-α in white adipose tissue, and completely eliminated the elevated blood plasma cytokine levels of TNF-α and IL-1β, demonstrating a potent antisystemic inflammation effect of rFGF21 in the high-fat diet consumption induced obese mouse model.21
However, this systemic anti-inflammatory effects and mecha-nisms of rFGF21 in focal ischemic stroke of T2D warrant fur-ther investigation.
Interestingly, white matter injury induced by ischemic stroke inside and beyond stroke lesions is one of the clinically pathological features of T2D, which has been considered one of the key mechanisms for worse stroke damage and poorer recovery.33 A previous experimental study showed PPARγ
ac-tivation by rosiglitazone treatment that facilitated microglial polarization toward the beneficial M2 phenotype, was consid-ered at least in part, a contributor to the improvement of white matter integrity after cerebral ischemia in mice.34 Importantly,
we found rFGF21 significantly promoted white matter integ-rity by attenuating the axonal injury and increase myelination levels, which might be partially contributed to rFGF21 treat-ment-mediated PPARγ activation and one of important under-lying mechanisms for the improved neurological outcomes.
Although our experimental findings are both fundamen-tally and translationally significant, several limitations still exist. First, in this study, we used C57BLKS-Leprdb T2D mice (db/db T2D mice, Jackson Lab). We are aware that although the leptin receptor mutation does not reflect disease cause in humans, this model has already given us insight into glucose me-tabolism and identified novel pathways of its complications.35
However, there are variable pathogenic mechanisms between different T2D animal models36; testing rFGF21 in other animal
models of diabetes with ischemic stroke should be pursued in the future. Second, in the present study, we only tested rFGF21 effects in PPARγ activation, cytokine mRNA expression, and microglia/macrophage activation after focal stroke of T2D mice. However, our results suggested that rFGF21-mediated PPARγ activation might in part contribute to the proinflamma-tory inhibition in the brain of T2D mice. The causal relation and significance of the rFGF21-PPARγ activation in modulat-ing neuroinflammation of T2D stroke need to be defined by pharmacological and genetic approaches in future investiga-tion. Third, there are multiple pathological factors that dynam-ically and interactively participate in T2D stroke brain damage evolution and recovery processes.32 Thus, it would be
impor-tant to understand how rFGF21 may pharmacologically modu-late these individual pathological mechanisms of tissue injury and repair, which requires more investigations.37 Moreover,
we acknowledge that it is likely impossible to truly separate each mechanism or signaling pathway of rFGF21, however, the multitargeted property of rFGF21 might be a translational strength.15 Fourth, because of lack of available working
anti-body for immunohistochemistry to detect the active form of FGF21 receptor FGFR1 (phosphorylation-FGFR1), we were unable to define brain regions and cellular types for FGF21-FGFR1 activation that taking place, however, spatial and time profile of exogenous FGF21-mediated receptor activation and consequent biological signaling pathways in the context of neurovascular unit warrant further investigation.38 Fifth,
be-cause stroke patients while in hospital are treated with glucose-lowering medications, it would be important to have them as rFGF21 controls, which was missing in present study. New experiments including the glucose-lowering medication con-trols would be informative to gain a broader cross-comparison of clinically relevant antidiabetic agent. Last, this study was proposed as a proof-of-concept investigation, all translational aspects, include using FGF21 analog, testing, and comparing both male and female animals, should be further tested in a well-controlled preclinical translational setting.14,37
In conclusion, we demonstrate rFGF21 administration is beneficial for improving neurological outcomes in T2D stroke mice. The potential mechanisms are, at least in part, by sys-temic metabolic modulation, brain tissue PPARγ activation-mediated proinflammation inhibition, and whiter matter integrity promotion. rFGF21 might be developed as a novel and potent disease-modifying approach for treating ischemic stroke with T2D.
Sources of Funding
This study was in part supported by the National Institutes of Health grant RO1 NS099539 (Dr Wang).
Disclosures
None.References
1. Sandu RE, Buga AM, Uzoni A, Petcu EB, Popa-Wagner A. Neuroinflammation and comorbidities are frequently ignored fac-tors in CNS pathology. Neural Regen Res. 2015;10:1349–1355. doi: 10.4103/1673-5374.165208
2. Kernan WN, Launer LJ, Goldstein LB. What is the future of stroke preven-tion?: debate: polypill versus personalized risk factor modification. Stroke. 2010;41(suppl 10):S35–S38. doi: 10.1161/STROKEAHA.110.592022 3. Beckman JA, Paneni F, Cosentino F, Creager MA. Diabetes and vascular
disease: pathophysiology, clinical consequences, and medical therapy: part II. Eur Heart J. 2013;34:2444–2452. doi: 10.1093/eurheartj/eht142 4. Air EL, Kissela BM. Diabetes, the metabolic syndrome, and ischemic
stroke: epidemiology and possible mechanisms. Diabetes Care. 2007;30:3131–3140. doi: 10.2337/dc06-1537
5. Tureyen K, Bowen K, Liang J, Dempsey RJ, Vemuganti R. Exacerbated brain damage, edema and inflammation in type-2 diabetic mice sub-jected to focal ischemia. J Neurochem. 2011;116:499–507. doi: 10.1111/j.1471-4159.2010.07127.x
6. Paneni F, Beckman JA, Creager MA, Cosentino F. Diabetes and vascular disease: pathophysiology, clinical consequences, and medical therapy: part I. Eur Heart J. 2013;34:2436–2443. doi: 10.1093/eurheartj/eht149 7. Patrone C, Eriksson O, Lindholm D. Diabetes drugs and neurological
disorders: new views and therapeutic possibilities. Lancet Diabetes Endocrinol. 2014;2:256–262. doi: 10.1016/S2213-8587(13)70125-6 8. Woo YC, Xu A, Wang Y, Lam KS. Fibroblast growth factor 21 as an
emerging metabolic regulator: clinical perspectives. Clin Endocrinol (Oxf). 2013;78:489–496. doi: 10.1111/cen.12095
9. Murata Y, Konishi M, Itoh N. FGF21 as an endocrine regulator in lipid metabolism: from molecular evolution to physiology and pathophysi-ology. J Nutr Metab. 2011;2011:981315. doi: 10.1155/2011/981315 10. Hsuchou H, Pan W, Kastin AJ. The fasting polypeptide FGF21
can enter brain from blood. Peptides. 2007;28:2382–2386. doi: 10.1016/j.peptides.2007.10.007
11. Kim KH, Lee MS. FGF21 as a stress hormone: the roles of FGF21 in stress adaptation and the treatment of metabolic diseases. Diabetes Metab J. 2014;38:245–251. doi: 10.4093/dmj.2014.38.4.245
12. Kharitonenkov A, DiMarchi R. FGF21 revolutions: recent advances illu-minating FGF21 biology and medicinal properties. Trends Endocrinol Metab. 2015;26:608–617. doi: 10.1016/j.tem.2015.09.007
13. Gaich G, Chien JY, Fu H, Glass LC, Deeg MA, Holland WL, et al. The effects of LY2405319, an FGF21 analog, in obese human subjects with type 2 diabetes. Cell Metab. 2013;18:333–340. doi: 10.1016/j.cmet.2013.08.005
14. Zhang J, Li Y. Fibroblast growth factor 21 analogs for treating met-abolic disorders. Front Endocrinol (Lausanne). 2015;6:168. doi: 10.3389/fendo.2015.00168
15. Kim KH, Lee MS. FGF21 as a mediator of adaptive responses to stress and metabolic benefits of anti-diabetic drugs. J Endocrinol. 2015;226:R1–16. doi: 10.1530/JOE-15-0160
16. Planavila A, Redondo-Angulo I, Villarroya F. FGF21 and cardiac physiopathology. Front Endocrinol (Lausanne). 2015;6:133. doi: 10.3389/fendo.2015.00133
17. Liauw J, Hoang S, Choi M, Eroglu C, Choi M, Sun GH, et al. Thrombospondins 1 and 2 are necessary for synaptic plasticity and func-tional recovery after stroke. J Cereb Blood Flow Metab. 2008;28:1722– 1732. doi: 10.1038/jcbfm.2008.65
18. Bouet V, Boulouard M, Toutain J, Divoux D, Bernaudin M, Schumann-Bard P, et al. The adhesive removal test: a sensitive method to assess sensorimotor deficits in mice. Nat Protoc. 2009;4:1560–1564. doi: 10.1038/nprot.2009.125
19. Doeppner TR, Kaltwasser B, Bähr M, Hermann DM. Effects of neural progenitor cells on post-stroke neurological impairment-a detailed and comprehensive analysis of behavioral tests. Front Cell Neurosci. 2014;8:338. doi: 10.3389/fncel.2014.00338
20. Choi YK, Maki T, Mandeville ET, Koh SH, Hayakawa K, Arai K, et al. Dual effects of carbon monoxide on pericytes and neurogenesis in trau-matic brain injury. Nat Med. 2016;22:1335–1341. doi: 10.1038/nm.4188 21. Wang Q, Yuan J, Yu Z, Lin L, Jiang Y, Cao Z, et al. FGF21 attenuates
high-fat diet-induced cognitive impairment via metabolic regulation and
anti-inflammation of obese mice. Mol Neurobiol. 2018;55:4702–4717. doi: 10.1007/s12035-017-0663-7
22. Liu N, Yu Z, Xiang S, Zhao S, Tjärnlund-Wolf A, Xing C, et al. Transcriptional regulation mechanisms of hypoxia-induced neu-roglobin gene expression. Biochem J. 2012;443:153–164. doi: 10.1042/BJ20111856
23. Mori T, Tan J, Arendash GW, Koyama N, Nojima Y, Town T. Overexpression of human S100B exacerbates brain damage and periin-farct gliosis after permanent focal ischemia. Stroke. 2008;39:2114–2121. doi: 10.1161/STROKEAHA.107.503821
24. Werneburg S, Fuchs HLS, Albers I, Burkhardt H, Gudi V, Skripuletz T, et al. Polysialylation at early stages of oligodendrocyte differenti-ation promotes myelin repair. J Neurosci. 2017;37:8131–8141. doi: 10.1523/JNEUROSCI.1147-17.2017
25. Kharitonenkov A, Shiyanova TL, Koester A, Ford AM, Micanovic R, Galbreath EJ, et al. FGF-21 as a novel metabolic regulator. J Clin Invest. 2005;115:1627–1635. doi: 10.1172/JCI23606
26. Lin Z, Pan X, Wu F, Ye D, Zhang Y, Wang Y, et al. Fibroblast growth factor 21 prevents atherosclerosis by suppression of hepatic sterol regu-latory element-binding protein-2 and induction of adiponectin in mice. Circulation. 2015;131:1861–1871. doi: 10.1161/CIRCULATIONAHA. 115.015308
27. Kim HW, Lee JE, Cha JJ, Hyun YY, Kim JE, Lee MH, et al. Fibroblast growth factor 21 improves insulin resistance and ameliorates renal injury in db/db mice. Endocrinology. 2013;154:3366–3376. doi: 10.1210/en.2012-2276
28. Chen MP, Tsai JC, Chung FM, Yang SS, Hsing LL, Shin SJ, et al. Hypoadiponectinemia is associated with ischemic cerebrovas-cular disease. Arterioscler Thromb Vasc Biol. 2005;25:821–826. doi: 10.1161/01.ATV.0000157784.25920.a7
29. Rosell A, Agin V, Rahman M, Morancho A, Ali C, Koistinaho J, et al. Distal occlusion of the middle cerebral artery in mice: are we ready to
assess long-term functional outcome? Transl Stroke Res. 2013;4:297– 307. doi: 10.1007/s12975-012-0234-1
30. Dutchak PA, Katafuchi T, Bookout AL, Choi JH, Yu RT, Mangelsdorf DJ, et al. Fibroblast growth factor-21 regulates PPARγ activity and the antidiabetic actions of thiazolidinediones. Cell. 2012;148:556–567. doi: 10.1016/j.cell.2011.11.062
31. Cai W, Yang T, Liu H, Han L, Zhang K, Hu X, et al. Peroxisome proliferator-activated receptor γ (PPARγ): a master gatekeeper in CNS injury and repair. Prog Neurobiol. 2018;163-164:27–58. doi: 10.1016/j.pneurobio.2017.10.002
32. Shukla V, Shakya AK, Perez-Pinzon MA, Dave KR. Cerebral ischemic damage in diabetes: an inflammatory perspective. J Neuroinflammation. 2017;14:21. doi: 10.1186/s12974-016-0774-5
33. Yu X, Song R, Jiaerken Y, Yuan L, Huang P, Lou M, et al. White matter injury induced by diabetes in acute stroke is clinically rele-vant: a preliminary study. Diab Vasc Dis Res. 2017;14:40–46. doi: 10.1177/1479164116675491
34. Han L, Cai W, Mao L, Liu J, Li P, Leak RK, et al. Rosiglitazone pro-motes white matter integrity and long-term functional recovery after focal cerebral ischemia. Stroke. 2015;46:2628–2636. doi: 10.1161/STROKEAHA.115.010091
35. McMurray F, Cox RD. Mouse models and type 2 diabetes: trans-lational opportunities. Mamm Genome. 2011;22:390–400. doi: 10.1007/s00335-011-9345-3
36. King AJ. The use of animal models in diabetes research. Br J Pharmacol. 2012;166:877–894. doi: 10.1111/j.1476-5381.2012.01911.x
37. Liu JJ, Foo JP, Liu S, Lim SC. The role of fibroblast growth factor 21 in di-abetes and its complications: a review from clinical perspective. Diabetes Res Clin Pract. 2015;108:382–389. doi: 10.1016/j.diabres.2015.02.032 38. Sa-Nguanmoo P, Chattipakorn N, Chattipakorn SC. Potential roles of
fibroblast growth factor 21 in the brain. Metab Brain Dis. 2016;31:239– 248. doi: 10.1007/s11011-015-9789-3