The Effect of Gap Length on Double-Strand Break Repair in Drosophila
Dena M. Johnson-Schlitz and William R. Engels
1Department of Genetics, University of Wisconsin, Madison, Wisconsin 53706 Manuscript received April 13, 2006
Accepted for publication May 14, 2006
ABSTRACT
When a double-strand break has a gap between the broken ends, the missing information can be restored through synthesis from a homologous template. Here we address the question of how long such a gap can be before this process fails. We measured the frequency of homologous repair in the Drosophila germ line following the creation of gaps of specific sizes ranging from 3.8 to 210 kb. We found that gaps of #11 kb can be repaired with approximately the same efficiency as breaks with no gap at all. However, a gap of 44 kb was repaired only rarely, and one of 210 kb was not repaired at a measurable frequency. We conclude that DNA gap repair is a length-limited process, but that this limitation is critical only for gaps
?11 kb.
A
cell faces a crisis with the occurrence of a double-strand break (DSB). It must either repair the break, perhaps imperfectly, or face cell cycle arrest. One indication of the importance of DSB repair is that defects in the process have been associated with ele-vated cancer risk in humans ( Jackson2002; Rothkamm and Lobrich2002; Maserand DePinho2003; Valerie and Povirk2003; Bryant2004).The mechanisms for DSB repair are often classified according to whether or not a homologous template, usually the sister chromatid or the homolog, is used. Pathways that do not utilize a template, such as non-homologous end joining (Mooreand Haber1996) or single-strand annealing (Fishman-Lobell et al. 1992; Prestonet al.2002), entail a greater risk of mutation but may be available in circumstances where templated repair is not. There are at least two pathways for ho-mologous repair: one with the potential for crossing over and one without. Crossing over in meiosis is thought to depend on which of these two pathways is followed (Allers and Lichten 2001; Hunter and Kleckner 2001; Borneret al.2004; Mazinaet al.2004). In mitotic cells there is evidence for competition among multiple pathways of homologous and nonhomologous mecha-nisms to repair the same pool of DSBs (Prestonet al. 2006).
Homologous repair is necessary to restore the missing information when the DSB includes a gap. Studies in Drosophila (Nassifet al.1994; Covenyet al.2002) and yeast (Paqueset al.1998) have shown that gaps as long as 10 kb can be repaired with only a two- to fourfold reduction in efficiency relative to breaks with little or
no missing sequence. Here we examine much larger gaps, up to 210 kb, to determine whether homolo-gous repair is limited by the length of gap that must be filled in.
MATERIALS AND METHODS
Drosophila stocks and crosses: Drosophila crosses were carried out with standard methods (Ashburner1989).
Ge-netic symbols can be found in FlyBase (Drysdaleet al.2005).
PCR tests and flanking DNA sequencing:DNA extraction for all PCR tests was done with individual flies as described (Gloorand Engels1992). Primers included D0 and G0 as
described (Prestonand Engels1996). These primers span
theP-insertion site at 50C and yield an amplicon of 145 bp if there is no insertion or deletion at this position. Gap lengths of the seven deletion chromosomes used here were determined by C. Flores, using thermal asymmetric interlaced PCR (Liu
and Whittier 1995) to amplify a segment spanning the
deletion and comparing its sequence with the Drosophila genome sequence.
RESULTS
We made use of a series of chromosomes in which the same P-element insertion is precisely juxtaposed with deletions of various sizes (Prestonet al.1996). These chromosomes, listed in Table 1, were formed by a pro-cess called ‘‘hybrid element insertion’’(HEI) (Grayet al. 1996; Preston et al. 1996), which involves two sister copies of a transposable element and an insertion site on the homologous chromosome. The result is a re-combinant chromosome with the transposable element flanked by a duplication or deletion. Figure 1A shows how such a chromosome can be used to generate a gap relative to the homolog. Each transposition event repre-sents an opportunity for gap repair, since thePelement leaves behind a double-strand break (Engelset al.1990; 1Corresponding author:Department of Genetics, University of Wisconsin,
425G Henry Mall, Madison, WI 53706. E-mail: [email protected]
Beall and Rio 1997). This break can be repaired by copying from the sister chromatid, by copying from the homolog, or by end joining. Our experiment cannot detect homologous repair from the sister chromatid, but it can detect the other two events and distinguish between them. When the homolog is used, there is a gap that must be filled in. The size of the gap is equal to the length of the flanking deletion.
Figure 1B shows the detailed mating scheme and lists eight types of offspring classified according to the mark-ers on chromosome 2. Gap formation and repair occur in the germ cells of males and are detected among the progeny. The top row (A–D) shows the expected geno-types if no repair, transposition, or excision events have occurred. Note that category A includes deletions on both homologs. Some of the deletions we tested—those of$10 kb—encompassed essential genes, thus render-ing category A inviable. For that reason we used the average number of offspring in categoriesB,C, andD
to estimate the Mendelian expectation for each class. CategoriesA9–D9are similar to the corresponding geno-types in the top row except for the presence or absence of theP{w*} element, as indicated by eye color. These genotypes represent potential repair events (categoryA9), transposition events (categoriesB9andC9), or excision events without complete templated repair (category
D9). Offspring in categoryA9were tested further by PCR as described (Table 2) to determine whether successful templated repair had occurred as opposed to end join-ing or other events that have been observed at this site (Preston and Engels 1996; Preston et al. 2002) or incomplete repair such as that described by McVeyet al. (2004a,b). As described in Table 2, the observed ratio from these PCR results was then applied as a correction factor to determine the frequency of templated repair. Recombinants betweenalandspwere also scored, but
these events were rare (203/72,601) and were not included in our calculations.
Each cross in Figure 1 was set up with a single male and three to four females at room temperature, and offspring were scored up to day 21. The use of single-male crosses has the advantage of allowing accurate quantitative measurement of the frequency of gap re-pair and a robust statistical analysis. Thus, each indi-vidual male parent provides a statistically independent estimate of repair frequency.
Note that excision of theP{w*} element followed by end joining or incomplete repair can result in offspring phenotypically similar to category A9. To avoid error from these false positives, we extracted DNA from most of theA9offspring and tested them by PCR as described in Table 2. Such false positives are expected primarily from the no-deletion controls and the three smaller deletions that do not encompass any essential genes. In the case of the four larger deletions, all end-joining events and most incomplete repair events will render genotype A9 inviable, and no false positive will occur. Therefore, as expected, the PCR correction given in Table 2 is substantial for the no-deletion controls and the three smaller deletions, but is nonexistent for the four largest deletions.
We tested eight chromosomes with deletions (i.e., gap lengths) ranging from 0 to 210 kb. An average of 96 individual male parents were testcrossed per chromo-some for a total of 72,398 progeny scored in the ex-periment. The results are shown graphically in Figure 2 and are available numerically in more detail in Tables 1 and 2.
The top bars in Figure 2 show estimates of the fre-quency of gap formation and homologous repair among all gametes receiving the relevant (al1
sp1
) chromosome from the males shown in Figure 1. Gap formation and
TABLE 1
Progeny counts from the eight primary classes shown in Figure 1B
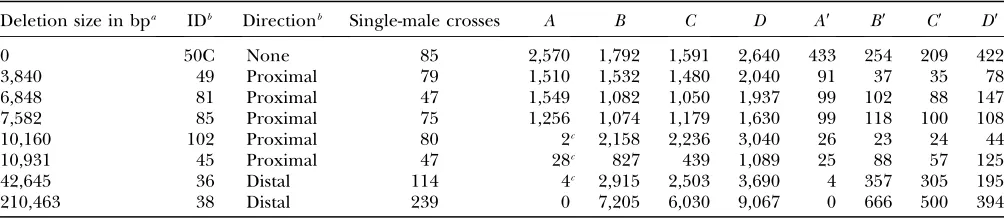
Deletion size in bpa IDb Directionb Single-male crosses A B C D A9 B9 C9 D9
0 50C None 85 2,570 1,792 1,591 2,640 433 254 209 422
3,840 49 Proximal 79 1,510 1,532 1,480 2,040 91 37 35 78
6,848 81 Proximal 47 1,549 1,082 1,050 1,937 99 102 88 147
7,582 85 Proximal 75 1,256 1,074 1,179 1,630 99 118 100 108
10,160 102 Proximal 80 2c 2,158 2,236 3,040 26 23 24 44
10,931 45 Proximal 47 28c 827 439 1,089 25 88 57 125
42,645 36 Distal 114 4c 2,915 2,503 3,690 4 357 305 195
210,463 38 Distal 239 0 7,205 6,030 9,067 0 666 500 394
aDeletion lengths are based on a standard reference sequence and will vary slightly with stock. b
Abbreviations for deletion chromosome names and directions are as reported (Prestonet al.1996). Standard nomenclature
would be,e.g.,Df(2R)50C-49.
c
The chromosomes with 10-, 11-, 43-, and 210-kb deletions are missing one or more essential genes encompassed by the de-letion. Therefore, most individuals in category A do not survive from those crosses. A few exceptions were expected, however, resulting from coincidental occurrence of both gap repair andP-element transposition to place a copy of theP{w*} element on one of the other paternal chromosomes. We tested 20 of these exceptions by PCR similar to the test done for categoryA9
repair can occur in the premeiotic germ cells, where a single event can yield multiple gametes. Our use of single-male crosses allowed us to take this clustering into account to calculate unbiased standard errors as de-scribed (Engels1979; Prestonet al.2006).
The bottom bars show an alternative measure of gap repair that takes account of variation inP-element mo-bility among the deletion-bearing chromosomes. All of
these chromosomes have the sameP-element insertion, but its mobility can be affected by the flanking deletions that place one side of the element in a different DNA context. Since the Pelement carries a visible eye-color marker [mini-white(Drysdaleet al.2005), denotedw*], the same cross that yielded an estimate of the gap repair frequency also gave us an estimate of the P-element mobility. That is, the transposition rate is proportional to the number ofal spprogeny carrying a copy of theP
element, as described in more detail in Tables 1 and 2. Therefore, our alternative measure of repair efficiency is proportional to the number of transpositions (i.e., opportunities for gap repair) for each successful gap repair where the homolog serves as the template. Since it is a reciprocal measure of repair efficiency, the bars are drawn going downward in Figure 2.
We see from Figure 2 that gaps of length up to 11 kb were all repaired efficiently. In fact, only the 10-kb gap category had a repair frequency that differed signifi-cantly from any of the others in this group when comparisons were made using permutation tests with the single-male replicates. The chromosome carrying this 10-kb deletion was also unusual in having a trans-position frequency of only 1%, which was less than half that of any of the other chromosomes in the study (Tables 1 and 2). If this chromosome is discounted, our results show no evidence that gaps up to 11 kb inhibit gap repair. The difference between gaps of up to 11 kb
vs. the two larger gaps is even clearer when one com-pares the ratios of repair opportunities to successful templated repair events (Figure 2, bottom).
In contrast, the chromosome with a 43-kb deletion showed a markedly reduced repair frequency (Figure 2) even though its transposition rate of 11% was compara-ble to that of the other chromosomes (Tacompara-bles 1 and 2). Even more striking was the complete lack of detectable homologous repair of 210-kb gaps. To maximize the power of our test with this chromosome, we performed 239 single-male crosses and scored 23,862 progeny but recovered no cases of homologous repair. We conclude that successful filling in of gaps is severely inhibited for gaps of 43 kb and essentially absent for gaps of 210 kb.
DISCUSSION
Several differences between this work and previous studies should be mentioned: We examined repair events in the germ cells of Drosophila males as opposed to the female germ line (Covenyet al. 2002) or yeast cells (Paqueset al.1998). In addition, there is evidence that the genomic position of a DSB can influence the relative usage of repair pathways (W. R. Engels and C. Preston, unpublished results). However, we believe the critical difference between the present study and previous investigations is that much larger gaps were examined: 210 kb vs. 10 kb. Only by testing repair of
Figure1.—(A) Gap formation in parental males. A
P-trans-posable element at cytological position 50C on chromosome 2 is immediately adjacent to a deletion (Prestonet al.1996).
Excision of this element in the male germ cells is activated by aP-transposase gene present on chromosome 3 (not in-cluded in the diagram). The result of this excision is a gap relative to the homolog equal to the length of the deletion. Males of this kind are used in the cross shown below to mea-sure the frequencies of gap repair and other events. (B) Mat-ing scheme for measurMat-ing repair frequency. See FlyBase (Drysdaleet al.2005) for all genetic symbols not explicitly
defined. The male parent (top right) is equivalent to the males indicated in part A. ThePelement carrying a mini-white gene and designatedP{w*} is located at cytological position 50C and is mobilized in the germ line by a third-chromosomal transposase source designated D2–3(99B) as described by Robertson et al. (1988). Deletions are denoted by angle
brackets (‘‘,d.,’’ etc.). In categoriesA,D, andD9the deletion symbol is shown with a question mark to indicate that the presence of the deletion was not confirmed in those cases. The female parents are heterozygous for a specific deletion, Df(2R)50C-25, abbreviated ,#25., which was selected be-cause it extends in both directions from the 50C insertion point and encompasses essential genes on both sides. It was obtained via two-step HEI as described by Preston et al.
(1996). The symbol ‘‘TMx’’ refers to one of the balancer chro-mosomesTMSorTM2(Drysdaleet al.2005). In categories
these large gaps were we able to detect the length lim-itation of DSB repair.
Our results provide strong evidence that homologous gap repair is a length-limited process, but other inter-pretations should be considered. In particular, note that the two chromosomes that produce gaps of 43 and 210 kb have deletions extending in the rightward direction from the originalP insert whereas the other chromo-somes in this study extend leftward (Table 1). If this directional difference were somehow affecting gap re-pair, it would offer an alternative explanation for our main results. However, two lines of evidence argue against a directional effect being an important factor in the observed repair differences. First, our data show that the 43-kb deletion was repaired significantly more often than the 210-kb deletion (P¼0.01), even though both are in the same direction. In addition, previous data showing symmetrical conversion tracts following excision of this same P insert (Preston and Engels 1996) argue against direction being crucial. That is, we saw no evidence of a directional effect, at least for the
relatively short conversion tracts (,3 kb) observed in that study.
When a gap is too large to be filled in from the homolog, there are at least two other options that would allow continuation of the cell cycle. If the cell is in late S-phase or G2, templated repair from the sister chro-matid is possible. In that case, only the length of the
P{w*} element must be filled in (Figure 1). Another pos-sibility is nonhomologous end joining (NHEJ), which does not depend on the stage of the cell cycle. In fact, several NHEJ products were observed in the present study among the offspring in categoryA9in cases where the PCR test did not indicate templated repair. In some cases NHEJ was inferred from a PCR fragment larger or smaller than that expected from templated repair. In other cases, the mutant phenotype of the CP1 gene (Grayet al.1998) implied loss of sequences flanking the insertion, as would be consistent with end joining. For example, the 4-kb deletion yielded 91 offspring of cat-egoryA9(Table 1). Of these, 39 had theCP1mutant phe-notype, suggesting an aberrant repair event of some
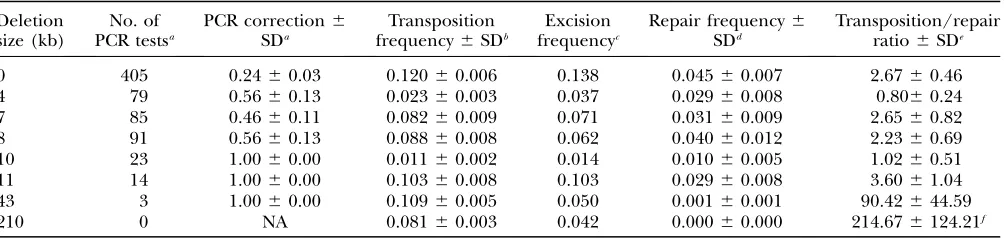
TABLE 2
Estimates ofP-element mobility and gap repair frequency
Deletion size (kb)
No. of PCR testsa
PCR correction6
SDa
Transposition frequency6SDb
Excision frequencyc
Repair frequency6
SDd
Transposition/repair ratio6SDe
0 405 0.2460.03 0.12060.006 0.138 0.04560.007 2.6760.46
4 79 0.5660.13 0.02360.003 0.037 0.02960.008 0.8060.24
7 85 0.4660.11 0.08260.009 0.071 0.03160.009 2.6560.82
8 91 0.5660.13 0.08860.008 0.062 0.04060.012 2.2360.69
10 23 1.0060.00 0.01160.002 0.014 0.01060.005 1.0260.51
11 14 1.0060.00 0.10360.008 0.103 0.02960.008 3.6061.04
43 3 1.0060.00 0.10960.005 0.050 0.00160.001 90.42644.59
210 0 NA 0.08160.003 0.042 0.00060.000 214.676124.21f
a
DNA was extracted from gap repair candidates in phenotypic category A9(Figure 1B) and amplified with primers D0 and G0 described previously (Prestonet al.1996). The correction factor was calculated as the proportion of PCR tests that yielded the
145-bp fragment expected from gap repair templated by the homolog. The standard deviation was obtained from the single-male independent replicates as described (Engels1979). Note that in the case of the lethal-bearing deletions, all 40 PCR tests indicated
complete homologous repair, as opposed to end joining or a partial repair of the kind observed in other systems (McVeyet al.
2004a, McVeyet al.2004b).
b
Transposition frequency was estimated asððB9 1C9Þ=ðB1B9 1C1C9ÞÞfrom the phenotypic categories shown in Figure 1B. Note that this estimate does not include a correction for transpositions to theal1
sp1
or X chromosomes. It can, therefore, be considered proportional to the absolute transposition rate. Its standard error was computed from the single-male independent crosses as described (Engels1979).
cExcision frequency was computed from the phenotypic categories in Figure 1B asð
D9=ðD1D9ÞÞ.
dRepair frequency was estimated asð3A9=ð
B1B9 1C1C9 1D1D9ÞÞ ðPCR correctionÞ. The first term is the phenotypic repair frequency computed from the offspring counts shown in Table 1, and the second term comes from the third column of this table. Standard errors for the phenotypic repair frequency were computed from the single-male replicates as described (Engels1979),
and the standard errors for the corrected repair frequency were computed from the large-sample approximation for the standard error of a product of two random variables,xandy: sðxyÞ ¼ ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffix2s2
y1y2s2x p
. In this case,xandyrefer to the two factors in the ex-pression for repair frequency,i.e., the phenotypic repair frequency and the PCR correction. This approximation requires that xandynot be strongly correlated. Inspection of a scatterplot of the phenotypic repair frequencyvs. the PCR correction for in-dividual male crosses showed no evidence of such a correlation.
e
The transposition/repair ratio was computed from columns 4 and 6 of this table. The standard error was approximated by the large-sample method:sðx=yÞ ¼ ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffix2s2
y=y 41s2
x=y 2 p
. In this case,xandyrefer to columns 4 and 6.
f
Since no repair events were observed for the 210-kb gap, we used as the denominator the upper limit of a 95% confidence interval. This procedure amounts to using the phenotypic repair frequency that would have been computed if we had observed –ln(0.05)¼3 repair events instead of zero. The resulting repair frequency and its (binomial) standard error were 0.000386
kind, as was confirmed in 30/30 tested by PCR. In addition, the PCR tests showed that 5 of the 52 CP11 offspring had no amplicon with primers D0 and G0, also indicating end joining or another aberrant event.
Why is homologous gap repair unable to handle longer gaps? Paques et al. (1998) suggested that the DNA polymerase used in DSB repair may lack processi-vity compared to the polymerase used for chromosome replication or break-induced replication (Kraus et al. 2001). A similar suggestion was made by McVeyet al. (2004a) to explain partial repair events. Time con-straints in the cell cycle might also prevent polymerase from completing extremely long tracts. Nonhomolo-gous end joining is available as an alternative to homol-ogous pathways for repair of the same breaks. Indeed, the relative usage of these pathways varies in a compen-satory fashion (Preston et al. 2006). The inability of
homologous gap repair to handle extremely long gaps provides another rationale for the existence of multiple, but not necessarily redundant, mechanisms for DSB repair.
Carlos Flores conceived the idea of usingPinserts adjacent to very
long deletions to test for very long gap repair. He also determined the lengths of the deletions by sequencing and provided comments on the manuscript. Christine Preston produced and characterized the de-letion chromosomes and helped with the manuscript. This work was supported by the National Institutes of Health (GM30948).
LITERATURE CITED
Allers, T., and M. Lichten, 2001 Differential timing and control of
noncrossover and crossover recombination during meiosis. Cell
106:47–57.
Ashburner, M., 1989 Drosophila, A Laboratory Manual.Cold Spring
Harbor Laboratory Press, Cold Spring Harbor, NY.
Beall, E. L., and D. C. Rio, 1997 Drosophila P-element transposase
is a novel site-specific endonuclease. Genes Dev.11:2137–2151.
Borner, G. V., N. Klecknerand N. Hunter, 2004
Crossover/non-crossover differentiation, synaptonemal complex formation, and regulatory surveillance at the leptotene/zygotene transition of
meiosis. Cell117:29–45.
Bryant, P. E., 2004 Repair and chromosomal damage. Radiother.
Oncol.72:251–256.
Coveny, A. M., T. Drayand G. B. Gloor, 2002 The effect of
heter-ologous insertions on gene conversion in mitotically dividing
cells inDrosophila melanogaster.Genetics161:249–258.
Drysdale, R. A., M. A. Crosby, W. Gelbart, K. Campbell,
D. Emmertet al., 2005 FlyBase: genes and gene models. Nucleic
Acids Res.33:D390–D395.
Engels, W. R., 1979 The estimation of mutation rates when
premei-otic events are involved. Environ. Mutagen.1:37–43.
Engels, W. R., D. M. Johnson-Schlitz, W. B. Egglestonand J. Sved,
1990 High-frequency P element loss in Drosophila is
homolog-dependent. Cell62:515–525.
Fishman-Lobell, J., N. Rudinand J. E. Haber, 1992 Two alternative
pathways of double-strand break repair that are kinetically
separa-ble and independently modulated. Mol. Cell. Biol.12:1292–1303.
Gloor, G., and W. Engels, 1992 Single-fly DNA preps for PCR.
Dros. Inf. Serv.71:148–149.
Gray, Y. H., M. M. Tanakaand J. A. Sved, 1996 P-element-induced
recombination inDrosophila melanogaster: hybrid element
inser-tion. Genetics144:1601–1610.
Gray, Y. H., J. A. Sved, C. R. Prestonand W. R. Engels, 1998
Struc-ture and associated mutational effects of the cysteine proteinase
(CP1) gene of Drosophila melanogaster. Insect Mol. Biol.7:291–
293.
Hunter, N., and N. Kleckner, 2001 The single-end invasion: an
asymmetric intermediate at the strand break to
double-holliday junction transition of meiotic recombination. Cell106:
59–70.
Jackson, S. P., 2002 Sensing and repairing DNA double-strand
breaks. Carcinogenesis23:687–696.
Kraus, E., W. Y. Leungand J. E. Haber, 2001 Break-induced
repli-cation: a review and an example in budding yeast. Proc. Natl.
Acad. Sci. USA98:8255–8262.
Liu, Y. G., and R. F. Whittier, 1995 Thermal asymmetric interlaced
PCR: automatable amplification and sequencing of insert end fragments from P1 and YAC clones for chromosome walking.
Genomics25:674–681.
Maser, R. S., and R. A. DePinho, 2003 Take care of your
chromo-somes lest cancer take care of you. Cancer Cell3:4–6.
Mazina, O. M., A. V. Mazin, T. Nakagawa, R. D. Kolodnerand S. C.
Kowalczykowski, 2004 Saccharomyces cerevisiae Mer3 helicase
stimulates 39-59heteroduplex extension by Rad51; implications for
crossover control in meiotic recombination. Cell117:47–56.
McVey, M., M. Adams, E. Staeva-Vieira and J. J. Sekelsky,
2004a Evidence for multiple cycles of strand invasion during
re-pair of double-strand gaps in Drosophila. Genetics167:699–705.
Figure2.—Frequency of homologous gap repair. Top bars
show the frequency of homologous repair of gaps whose lengths are indicated above. The numerator for this measure is the number of progeny whose paternal second chromo-some has undergone loss of thePelement and restoration of the deleted sequence. Such progeny are recognized pheno-typically as w, al1
, Cy1
, sp1
McVey, M., D. Radutand J. J. Sekelsky, 2004b End-joining repair
of double-strand breaks inDrosophila melanogasteris largely DNA
ligase IV independent. Genetics168:2067–2076.
Moore, J. K., and J. E. Haber, 1996 Cell cycle and genetic
require-ments of two pathways of nonhomologous end-joining repair of double-strand breaks in Saccharomyces cerevisiae. Mol. Cell.
Biol.16:2164–2173.
Nassif, N., J. Penney, S. Pal, W. R. Engelsand G. B. Gloor, 1994
Ef-ficient copying of nonhomologous sequences from ectopic sites via
P-element-induced gap repair. Mol. Cell. Biol.14:1613–1625.
Paques, F., W. Y. Leungand J. E. Haber, 1998 Expansions and
con-tractions in a tandem repeat induced by double-strand break
repair. Mol. Cell. Biol.18:2045–2054.
Preston, C. R., and W. R. Engels, 1996 P-element-induced male
re-combination and gene conversion in Drosophila. Genetics144:
1611–1622.
Preston, C. R., J. A. Svedand W. R. Engels, 1996 Flanking
dupli-cations and deletions associated withP-induced male
recombina-tion in Drosophila. Genetics144:1623–1638.
Preston, C. R., W. Engelsand C. Flores, 2002 Efficient repair of
DNA breaks in Drosophila: evidence for single-strand annealing
and competition with other repair pathways. Genetics161:711–
720.
Preston, C. R., C. C. Floresand W. R. Engels, 2006 Differential
usage of alternative pathways of double-strand break repair in
Drosophila. Genetics172:1055–1068.
Robertson, H. M., C. R. Preston, R. W. Phillis, D. Johnson
-Schlitz, W. K. Benzet al., 1988 A stable genomic source of
Pelement transposase inDrosophila melanogaster.Genetics118:
461–470.
Rothkamm, K., and M. Lobrich, 2002 Misrepair of
radiation-induced DNA double-strand breaks and its relevance for
tu-morigenesis and cancer treatment (review). Int. J. Oncol.21:
433–440.
Valerie, K., and L. F. Povirk, 2003 Regulation and mechanisms of
mammalian double-strand break repair. Oncogene22:5792–5812.