©
DOI: 10.1534/genetics.104.036509
Infrequent Co-conversion of Markers Flanking a Meiotic Recombination
Initiation Site in
Saccharomyces cerevisiae
Lea Jessop,* Thorsten Allers
†and Michael Lichten*
,1*Center for Cancer Research, National Cancer Institute, Bethesda, Maryland 20892 and†Institute of Genetics,
University of Nottingham, Queen’s Medical Centre, Nottingham NG7 2UH, United Kingdom
Manuscript received September 18, 2004 Accepted for publication December 13, 2004
ABSTRACT
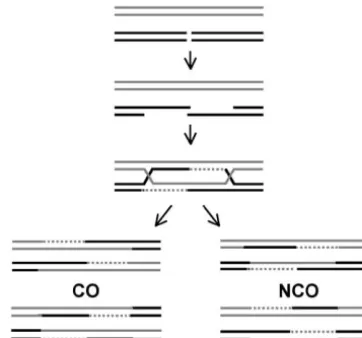
To study the mechanism of meiotic recombination inSaccharomyces cerevisiae, we examined recombination in an interval where the majority of events are initiated at a single hotspot for DNA double-strand breaks (DSBs), with little or no expected contribution by outside initiation events. This interval contained infrequently corrected palindromic markers 300 bp to the left and 600 bp to the right of the DSB hotspot. Conversion of single markers occurred frequently, while conversion of both markers occurred rarely, and many of the tetrads in which both markers converted were the products of multiple events. These data indicate that most meiotic recombination intermediates are asymmetrically positioned around the initiating DSB, with a short (⬍300 bp) tract of heteroduplex DNA (hDNA) to one side and hDNA on the other side frequently extending 600 bp or more. One consequence of this asymmetry is the preferential concentra-tion of crossovers in the vicinity of the initiating DSB.
I
NSaccharomyces cerevisiaeand other organisms, recom- that make up the hDNA segregate at the first mitotic division after meiosis (reviewed byPetes et al.1991). bination between homologous chromosomes isre-quired to ensure proper segregation of chromosomes If the mismatch is repaired, the marker will either be restored to parental (4:4) segregation or show full con-during meiosis (reviewed by Petronczkiet al. 2003).
Recombination is initiated by DNA double-strand breaks version (6:2 or 2:6) segregation. In this article, the term “gene conversion” encompasses both PMS and full con-(DSBs) made by the meiosis-specific endonuclease Spo11
(Bergerat et al. 1997;Keeney et al. 1997). DSB sites version. Gene conversion is often, but not always, associ-ated with an exchange of flanking markers. Gene con-tend to occur in clusters 100–200 nt wide (de Massyet
versions not associated with an exchange of flanking al. 1995; Liu et al. 1995; Xu and Petes 1996); these
markers are referred to as noncrossovers (NCOs); events clusters will be referred to as DSB hotspots. After DSB
that involve an exchange of flanking markers are re-formation, break ends are resected to generate 3⬘
single-ferred to as crossovers (COs). strand tails (Sun et al. 1991; Nag and Petes 1993).
The double-strand break repair (DSBR) model of It has been proposed that these single strands invade
Szostaket al.(1983), as modified bySunet al.(1991), homologous sequences and initiate DNA synthesis,
ulti-predicts that both NCOs and COs result from the resolu-mately forming joint molecules that contain two
Holli-tion of a dHJ intermediate (Figure 1). COs are produced day junctions flanking a tract of heteroduplex DNA
by cutting the two junctions in the opposite orientation, (hDNA;Szostak et al. 1983). Evidence in support of
while cutting in the same orientation gives rise to NCOs. this view comes from the isolation of double Holliday
This model makes two significant predictions. First, junction (dHJ) intermediates from meiotic yeast cells
hDNA will flank the DSB in both COs and NCOs, lead-(CollinsandNewlon1994;SchwachaandKleckner
ing to frequent bidirectional conversion and hDNA 1994) and the detection of hDNA in these intermediates
present in two of the four meiotic products, which in (AllersandLichten2001b).
S. cerevisiae are present as spores in a tetrad ascus. In Gene conversion is a signal of the presence of hDNA.
one spore, hDNA will be present to the left of the DSB, If unrepaired, a marker in asymmetric hDNA will
segre-and in the other spore, hDNA will be to the right. The gate in a 5:3 or a 3:5 ratio of parental alleles among
second prediction is specific to CO products. The loca-the eight strands of DNA present in loca-the four haploid
tion of the Holliday junctions (HJs) in the dHJ interme-products of meiosis. These are postmeiotic segregation
diate as well as the direction of their resolution will (PMS) events, so named because the two DNA strands
dictate the point where unconverted markers appear to exchange linkage. If a single marker is included in hDNA, and if the two HJs are resolved in the opposite
1Corresponding author: National Cancer Institute, Bldg. 37, Room
orientations, but without bias, then in half of the CO
6124, 37 Convent Dr. MSC 4255, Bethesda, MD 20829-4255.
E-mail: [email protected] products the exchange point will be to the right of the
and 670 bp to the right of theHIS4DSB hotspot, with only 3% of events being bidirectional.Gilbertsonand Stahl(1996) observed more frequent co-PMS for mark-ers 130 bp to the left and 190 bp to the right of the ARG4 DSB hotspot (55% of events that converted a marker in ARG4 were bidirectional), but the location of hDNA in these products did not fit a central predic-tion of the DSBR model—that both recombinant prod-ucts should contain hDNA. Instead, prodprod-ucts showing PMS for both markers contained hDNA in only a single spore, and the majority of these products contained hDNA in a configuration most consistent with initiation at a site other than theARG4DSB hotspot. Furthermore, the majority of PMS-associated crossovers mapped to the left of both markers in an interval that also contained
Figure1.—DSBR model for meiotic recombination (Szos- theDED81DSB hotspot. taket al.1983;Sunet al.1991). DSBs form and are resected
To detect hDNA and determine the timing of
inter-to give 3⬘ single-strand tails. These tails invade homologous
mediate and recombinant product formation during
sequences and initiate DNA synthesis (broken lines) to form
a dHJ intermediate. Both NCO and CO products are formed meiosis,AllersandLichten (2001a) used a physical
by junction cleavage; in every case, the two products contain assay to directly detect hDNA molecules. This assay ex-hDNA on opposite sides of the DSB. amined ectopic recombination between dispersed
re-porter constructs inserted at HIS4 andLEU2. Hetero-duplex DNA associated with NCOs appeared at least 30 marker and in half it will be to the left (discussed in min before hDNA associated with COs. Mutants that Gilbertson and Stahl 1996; for an illustration, see accumulate dHJs, such as those lacking the meiosis-Figure 5). This can be determined only in tetrads where specific transcription factor Ntd80 or the polo-like ki-the relevant marker does not undergo full conversion nase Cdc5, displayed a decrease in the formation of CO (i.e., shows PMS). Both predictions can be tested by products but accumulated NCO products (Allersand examining patterns of gene conversion and crossing Lichten2001a;Clyneet al.2003), indicating that
for-over in appropriately marked strains. mation of most NCO products does not depend upon
TABLE 1
Diploid yeast strains
MJL2834 HIS4 leu2-R::URA3-tel-ARG4 MATa
his4::URA3-tel-ARG4 LEU2 MAT␣
rad50-KI81::KanMX6 rad50-KI81::KanMX6
MJL2870 HIS4 leu2::ura3-EcPal⫹104-tel-ARG4 MATa
his4::URA3-tel-arg4-EcPal⫹9 LEU2 MAT␣
MJL2902 HIS4 leu2::URA3-tel-ARG4 MATa
his4::ura3-EcPal⫹104-tel-arg4-EcPal⫹9 LEU2 MAT␣
MJL2936 his4::URA3-tel-ARG4
STE50-natMX-RRP7 his4::ura3-EcPal⫹104-tel-arg4-EcPal⫹9
PacI RHQ1-hphMX-FUS1 SphI
LEU2 leu2-R
MATa
MAT␣
MJL2957 his4::URA3-tel-ARG4 RHQ1-hphMX-FUS1 LEU2 MATa
STE50-natMX-RRP7 his4::ura3-EcPal⫹104-tel-arg4-EcPal⫹9 leu2-R MAT␣
MJL2959 his4-R::URA3-tel-ARG4
STE50-natMX-RRP7 his4::ura3-EcPal⫹104-tel-ARG4
PacI RHQ1-hphMX-FUS1 SphI
LEU2 MATa
leu2-R MAT␣
MJL2961 his4::URA3-tel-ARG4 MATa
his4::URA3-tel-ARG4 MAT␣
rad50-KI81::KanMX6 rad50-KI81::KanMX6
MJL2976 his4-R::URA3-tel-ARG4
STE50-natMX-RRP7 his4::URA3-tel-arg4-EcPal⫹9
PacI RHQ1-hphMX-FUS1 SphI
LEU2 MATa
leu2-R MAT␣
MJL3010 MATa
MAT␣
rad50-KI81::KanMX6 rad50-KI81::KanMX6
MJL2960 his4::URA3-tel-ARG4
STE50-natMX-RRP7 his4::ura3-EcPal⫹104-tel-arg4-EcPal⫹9
PacI RHQ1-hphMX-FUS1 SphI
LEU2 MATa
leu2-R MAT␣
rad50-KI81::kanMX6 rad50-KI81::kanMX6
All strains are homozygous forura3⌬(hindIII-smaI) arg4⌬(eco47III-hpaI) lys2 ho::LYS2(AllersandLichten2001a).PacI/SphI andEcoRI (his4-R) restriction site polymorphisms are described inmaterials and methods.
NCO recombinants, suggesting that, in addition to uni- tributing to conversion of the palindromic markers. During allelic recombination, repression of DSBs at sites directional NCO products that arise by the SDSA
path-way, bidirectional NCO products may also arise via the near the DSB hotspot present in the interval minimized contributions from other initiation sites. Data from DSBR pathway. Moreover, althoughMerkeret al.(2003)
did observe tetrads with patterns of bidirectional hDNA these two configurations are in agreement, indicating that flanking heterologies present in the ectopic system predicted by the DSBR model, their complete data set
contains a greater number of tetrads that are most do not substantially bias the outcome of recombination. We find that bidirectional conversion of markers flank-readily explained as being the products of multiple
events. ing the DSB hotspot is infrequent and infer from these
Each of the genetic studies summarized here was data that most meiotic recombination events involve at done in a system where events producing recombinants most a short tract of hDNA to one side of the DSB, could have initiated at more than one DSB hotspot, while hDNA on the other side can frequently extend making it difficult to unambiguously identify the initia- beyond 600 bp. This asymmetry, in combination with a tion site for a given conversion event. In some cases, possible resolution bias, results in a substantial concen-many events were excluded in the effort to consider tration of crossovers in the vicinity of the initiating DSB. only events initiated by DSBs at a single hotspot. To
reduce these complications, we examined
recombina-MATERIALS AND METHODS
tion in an interval where most, if not all, events are initiated at a single DSB hotspot. To facilitate detection
Strains and media:All diploid strains used in this study are
of hDNA in the products, we placed poorly corrected SK1 derivatives (Kane and Roth 1974). Table 1 lists their palindromic markers on each side of the DSB hotspot. genotypes. The recombination interval examined contains a
1.1-kbHindIII-SmaIURA3fragment and a 2.3-kbEco47III-PstI
Additional flanking markers allowed NCO and CO
ARG4fragment inserted in a 64-bpClaI-SalI deletion ofHIS4 products to be distinguished as well as the position of
or at theEcoRV site ofLEU2. The open reading frames (ORFs)
the exchange point to be mapped. We examined both
ofURA3andARG4are arranged in a divergently transcribed
ectopic and allelic recombination events involving this configuration. The junction between the promoter regions of interval. During ectopic recombination, flanking heter- URA3 and ARG4 contains a 65-bp insert (5⬘-CAGCTGTCC
CACACACACCACCCACACACACACCACACCCACACCACA
con-CCACACCCACTCTGCAG) derived from yeast teleomere se-quences (Whiteet al.1993). The palindrome inarg4-EcPal⫹9
is at⫹9 of theARG4open reading frame (AllersandLichten 2001a) and the palindrome inura3-EcPal⫹104 was inserted at an XbaI site created at⫹104 of the URA3open reading frame. The palindromes contain duplicatedEcoRI sites, which allows physical detection. Strains used to study allelic recombi-nation contained a hygromycin B resistance cassette (hphMX) inserted 5130 bp upstream of the start of theHIS4ORF and the nourseothricin resistance gene (natMX) inserted 3804 bp downstream ofHIS4(Hoffmannet al.2005). Thehis4-R re-striction site mutant was constructed by filling in the EcoRI site 187 bp downstream of theClaI site inHIS4to create an
XmnI site. The PacI to SphI restriction site was created by inserting anSphI linker (GGCATGCC) into thePacI site 223 bp upstream of the HIS4 translation start. Details of strain construction will be supplied upon request.
Genetic procedures and media were as described (Allers andLichten2001a). Tetrads were dissected on solid media containing 2% peptone, 1% yeast extract, 4% glucose, 0.004% adenine, 2% agar, pH 5.5, and spores were germinated at 30⬚. Spores carrying hphMX or natMX were selected by replica plating spore colonies to YPD plates containing 300g/ml hygromycin B (Roche) or 100g/ml nourseothricin (Werner BioAgents).
CO determination:A subset of the ectopic COs generates
the same marker configuration as an allelic CO in the interval Figure2.—The recombination assay systems used in this betweenHIS4andLEU2. However, an ectopic CO leads to a study. (A) The 3.5-kbURA3-ARG4insert. Coding sequences deletion of the region betweenHIS4andLEU2in one of the are represented by gray and black boxes and noncoding se-products. Because this region contains no essential genes, quences are represented by gray and black lines. Inserted haploid recombinants containing this deletion are viable. This betweenURA3and ARG4is 65 bp of telomeric DNA (TEL, region contains theBIK1andFUS1genes;bik1⌬fus1⌬strains diagonally hatched box) that creates a strong meiotic DSB efficiently mate with wild-type strains, but not with otherbik1⌬ hotspot (see text) indicated by an arrow. Palindromic markers fus1⌬strains (Trueheart et al.1987). Spore colonies from (lollipops) are inserted at⫹104 of the URA3 open reading strains MJL2902 and MJL2870 were replica plated to YPD frame and at ⫹9 of the ARG4 open reading frame. These plates and cross stamped with a lawn of either of two tester markers areⵑ200–340 and 520–660 bp from the center of the strains, H417 (MATa ura3 ade1 trp5 lys2 met13-4 can1 cyh2 DSB hotspot, respectively. (B) Ectopic recombination system. ⌬[his4-LEU2]::URA3) or H418 (same as H417, but MAT␣), TheURA3-ARG4interval was inserted atHIS4(box with wavy which are deleted for the region betweenhis4andLEU2. After hatching) on one copy of chromosomeIIIand atLEU2 (check-4 hr on YPD, the plate was replica plated to minimal media ered box) on the homolog. The two insert sites atHIS4and plus 20g/ml histidine and 30g/ml lysine and incubated LEU2 are 17.9 kb apart in the SK1 strains used here. The at 30⬚overnight. If the original spore colony wasbik1⌬fus1⌬, marker configuration shown here is present in strain MJL2902, it did not mate with the tester strain and so did not grow on where the insert atHIS4 containsura3-EcPal⫹104and
arg4-selective media. If the spore colony wasBIK1 FUS1, it did mate EcPal⫹9. In strain MJL2870, the insert atLEU2contains
arg4-with the tester strain and the resulting diploid grew on selective EcPal⫹9and the insert atHIS4containsura3-EcPal⫹104. (C) media. Allelic recombination was examined betweenURA3-ARG4 in-Pulsed-field gel electrophoresis:Samples for pulsed-field gel tervals inserted at HIS4 on both copies of chromosomeIII. electrophoresis were prepared and run as previously described Homologs are distinguished bynatMX(horizontally hatched (Bordeet al.1999). box) orhphMX(vertically hatched box) insertions,ⵑ4.4 and
Random spore analysis:Separate patches of strains MJL2959 8.4 kb from the center of the DSB hotspot, respectively. and MJL2976 were sporulated on plates containing 2%
potas-sium acetate, 0.22% yeast extract, 0.05% glucose, and
com-plete amino acid mixture (drop-out mix; Abdullah and forEcoRI (New England Biolabs) to determine the location Borts2001) forⵑ24 hr. Ascus-cell mixtures were resuspended of meiotic COs from MJL2959 and MJL2976. To map DSBs, in 500l 1msorbitol, 10 mmEDTA, 50 mmKPO4(pH 7.5), DNA from meiotic cultures (GoyonandLichten1993) was 1%-mercaptoethanol, and 1 mg/ml zymolyase 80T (ICN). digested with 40 unitsXhoI (Roche) or with 10 units ofEcoRI Following incubation at 37⬚for 15 min, the spores were pel- (New England Biolabs) in the recommended buffer supple-leted and resuspended in 1 ml 0.1% Tween-80. Spores were mented with 100mspermidine. Digests were displayed on then sonicated until⬎90% were single spores as determined agarose gels, gel blots were probed with a32P-labeledARG4 by microscopy. Appropriate dilutions were plated onto fresh fragment (ARGD, ⫹165–⫹1413, relative to theARG4 open YPD plates containing either 300g/ml hygromycin B or 100 reading frame), and band intensities were quantified as de-g/ml nourseothricin. After 2 days, at least 200 colonies were scribed (AllersandLichten2001b).
GT membrane (Bio-Rad, Richmond, CA) in the same buffer. ⵑ130 bp, ura3-EcPal⫹104 is 200–340 bp from a DSB Prior to hybridization, DNA on the membrane was denatured andarg4-EcPal⫹9is 520–660 bp from a DSB.
by setting the membrane on filter paper saturated with 1.5m
We examined ectopic recombination in two diploid
NaCl and 0.5mNaOH for 15 min; the blot was then probed
strains that differ only in their marker configuration.
as described (Liu et al. 1995) with a 32P-labeled fragment
⫺307–⫺80 nt relative to theARG4open reading frame. Size MJL2902 contains bothura3-EcPal⫹104andarg4-EcPal⫹9
markers on these gels contained digests of mitotic DNA from inserted atHIS4(Figure 2B). In MJL2870,ura3-EcPal⫹104 S1898 [MAT␣ ura3⌬(hindIII-smaI) arg4⌬(eco47III-hpaI) lys2 is present atLEU2andarg4-EcPal⫹9is atHIS4. Cross-ho::LYS2 rad50-KI81::KanMX6 cyh2-z leu2::URA3-tel-ARG4].
overs between the inserts occur inⵑ30% of tetrads in both strains (Table 2). In MJL2902 the gene conversion frequency (both PMS and full conversion events) of
RESULTS
ura3-EcPal⫹104is 22%, somewhat greater than the 15% Recombination in theURA3-ARG4interval is initiated observed in MJL2870 (Table 2). The gene conversion from a single hotspot for DSBs:To determine the loca- frequency forarg4-EcPal⫹9is 14% in MJL2902 and 18% tion of hDNA in the products of meiotic recombination, in MJL2870. In both strains, roughly half of the gene we modified the system used previously byAllersand conversion events are associated with a CO (Table 2). Lichten(2001a) so that we could examine events initi- Although unidirectional conversion of these palin-ated at a single DSB hotspot. This modified system con- dromes is frequent, co-conversion rarely occurs. In this sists of a 3.5-kb insert containing theURA3 andARG4 work, we will refer to any tetrad with either PMS or full genes with 65 bp of yeast telomere sequences inserted conversion ofura3-EcPal⫹104in one spore colony and betweenURA3andARG4to create a strong DSB hotspot either PMS or full conversion ofarg4-EcPal⫹9 in any of (Figure 2A). This sequence induces a strong meiotic the four spore colonies as having undergone co-conver-DSB hotspot when inserted elsewhere (White et al. sion, without prejudgment as to whether these are the 1993). The location and frequency of DSBs in theURA3- products of single or multiple events. Only 6% (50/ ARG4interval inserted atHIS4on both copies of chro- 840) of tetrads from MJL2902 and only 5% (37/744) mosomeIIIwere mapped in arad50Sdiploid (MJL2961, of tetrads from MJL2870 show such co-conversion (Ta-Table 1), where DSBs are neither processed nor re- ble 2). On the basis of the gene conversion frequencies paired (Alani et al. 1990; Cao et al. 1990). Southern of the two markers, co-conversion as a result of two analysis reveals frequent DSBs in the vicinity of the telo- independent events would be expected to occur in 3% meric sequences (Figure 3A). In arad50Sdiploid hetero- of tetrads from both strains.
Figure3.—DSBs in theURA3-ARG4 in-terval. (A and B) Symbols are as in Figure 2. Southern blot of DNA isolated from a meiotic culture of MJL2961, arad50Sstrain homozygous for the insertion atHIS4. (A) DNA from 0 and 7 hr after induction of meiosis was digested withXmnI (X) and probed withARG4sequences as described inmaterials and methods. Size markers (lane not shown) are aBstEII digest of bac-teriophage DNA. (B) For higher-resolu-tion mapping of the DSB hotspot, DNA was digested withPpuMI (P) andBanII (B) and probed as described in materials and methods. Arrows indicate the location of DSBs. Size markers (lane not shown) are
MseI (728 bp),MspAI (665 bp), andEarI (278 bp) digests of DNA from a mitotic culture of S1898 (seematerials and meth-ods). The 862-bp size marker is aBanII/
PpuMI digest of 0 hr DNA of MJL2961. (C) Pulsed-field gels of undigested DNA from meiotic cells, using electrophoresis condi-tions and a probe (YCL075w) to reveal DSBs on the left arm of chromosomeIII. (Top lane) Strain MJL2961; (bottom lane) strain MJL3010. Traces are of phosphorimager signals for the relevant portion of each lane (black line, MJL2961; gray line, MJL3010); peak intensities of the full-length chromo-someIIIbands (marked with asterisks) are 2.1⫻103psl and 1.9⫻103psl for MJL2961 and MJL3010, respectively (psl are units that represent phosphor signal intensity). About 30% of chromosomes III suffer breaks in the left arm in both MJL2961 and MJL3010; in MJL2961, two-thirds of these breaks map to thehis4::URA3-ARG4insert. The trace for MJL3010 is broken to account for the absence of an insert atHIS4. The position of the recombination interval relative to the centromere is indicated on the schematic; size markers (lane not shown) were aHindIII digest of bacteriophageDNA andDNA concatamers.
Figure 1, of a DSB from the central hotspot, and thus ARG4 interval on other DSB sites along the left arm of chromosomeIII, we performed pulsed-field gel analysis may be the product of multiple events. The remaining
five tetrads contain hDNA in a single spore with parental of meiotic DNA from MJL2961, which contains the URA3-ARG4 interval at HIS4 on both homologs, and contributions on opposite strands,i.e., atrans
configu-ration. Four of these are nonrecombinant for flanking from MJL3010, which does not contain the interval. DSBs at sites near the hotspot were reduced as much markers and are thus compatible with
topoisomerase-mediated unwinding of dHJ intermediates (Gilbert- as fourfold relative to MJL3010 (Figure 3C), so that the majority of the DSBs in a 20-kb region centered around sonandStahl1996). Thus, of the 23 co-PMS tetrads
from MJL2902, only 8 show the hDNA pattern predicted his4::URA3-ARG4 occur at the hotspot. This strong re-pression of nearby breaks made it possible to examine by the classic DSBR model (Figure 1). Similarly, 17
tetrads from MJL2870 show co-PMS. Six are most likely recombination in strains containing allelic his4::URA3-ARG4inserts, with the expectation that most events were multiple events, as described for MJL2902. Five tetrads
show the pattern of hDNA predicted by the DSBR model initiated by the DSB hotspot within the URA3-ARG4 interval.
and three of these are CO associated.
Co-conversion is infrequent during allelic recombina- To distinguish COs from NCOs, strains in which one copy of chromosomeIIIwas marked withnatMX(which tion:When sequences containing a strong DSB site are
inserted at HIS4, they cause a reduction in DSBs at confers nourseothricin resistance) and the homolog was marked withhphMX(which confers hygromycin B resis-nearby sites (Wu and Lichten 1995; Fan et al. 1997;
URA3-TABLE 2
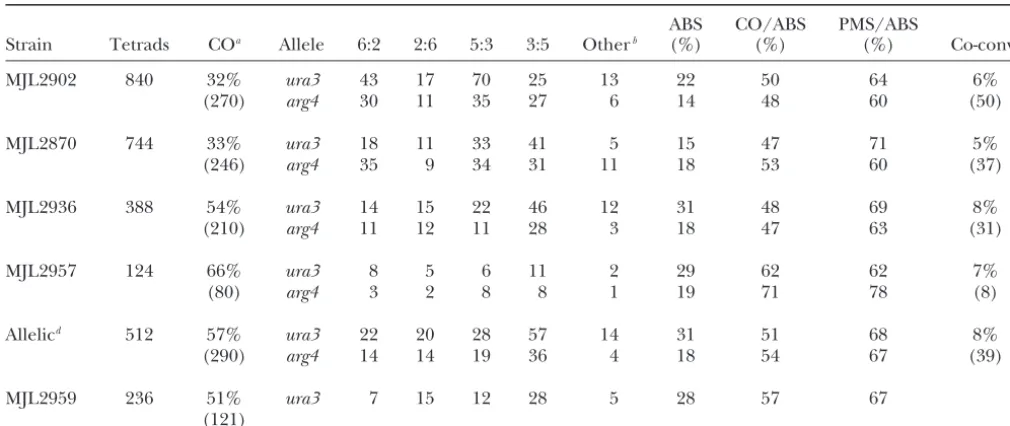
Aberrant segregation and co-conversion
ABS CO/ABS PMS/ABS
Strain Tetrads COa Allele 6:2 2:6 5:3 3:5 Otherb (%) (%) (%) Co-convc
MJL2902 840 32% ura3 43 17 70 25 13 22 50 64 6%
(270) arg4 30 11 35 27 6 14 48 60 (50)
MJL2870 744 33% ura3 18 11 33 41 5 15 47 71 5%
(246) arg4 35 9 34 31 11 18 53 60 (37)
MJL2936 388 54% ura3 14 15 22 46 12 31 48 69 8%
(210) arg4 11 12 11 28 3 18 47 63 (31)
MJL2957 124 66% ura3 8 5 6 11 2 29 62 62 7%
(80) arg4 3 2 8 8 1 19 71 78 (8)
Allelicd 512 57% ura3 22 20 28 57 14 31 51 68 8%
(290) arg4 14 14 19 36 4 18 54 67 (39)
MJL2959 236 51% ura3 7 15 12 28 5 28 57 67
(121)
ura3, ura3-EcPal⫹104; arg4, arg4-EcPal⫹9; ABS, total aberrant segregation events/total tetrads; CO/ABS, fraction of tetrads showing aberrant segregation that is crossover associated; PMS/ABS, fraction of tetrads showing aberrant segregation where the palindromic marker shows PMS; co-conv, fraction of tetrads showing co-conversion.
aThe interval for the ectopic crosses MJL2902 and MJL2870 isURA3–ARG4. For MJL2936, MJL2957, and MJL2959 the interval
isnatMX–hphMX. The number in parentheses is the total number of tetrads with a CO in the specified interval.
b7:1, 1:7, 8:0, 0:8, and aberrant 4:4, 5:3, 3:5, 6:2, or 2:6 segregation patterns (see supplementary material at http://genetics.org/
supplemental/ for details). Calculations of ABS frequencies assume that these were produced by two independent conversion events.
cThe number in parentheses is the number of tetrads that showed co-conversion as defined in the text. dData from MJL2936 and MJL2957 were summed.
of the DSB hotspot. The chromosome marked bynatMX similar to that seen for the ectopic recombination events: unidirectional conversion is frequent while co-carriedura3-EcPal⫹104andarg4-EcPal⫹9(Figure 2C).
DSB mapping in arad50Sstrain with this marker config- conversion is not. Gene conversion ofura3-EcPal⫹104 occurs in 31% of tetrads and 18% of tetrads have a gene uration (supplementary Figure 1 at http://www.genetics.
org/supplemental/) confirmed that most breaks in an conversion atarg4-EcPal⫹104.Roughly half of the gene conversion events are CO associated (Table 2).
Co-con-ⵑ24-kb region occurred at the URA3-ARG4 DSB
hot-spot. The next closest DSB site was at thenatMXinsert, version of both ura3-EcPal⫹104 and arg4-EcPal⫹9 oc-curs in 8% of tetrads; the frequency expected for two 4.4 kb to the left of theURA3-ARG4DSB hotspot, with
ⵑ2.4% of chromosome IIIs broken; no DSBs were de- independent events is 6% of tetrads.
A total of 24 tetrads show co-PMS for the two palin-tected at thehphMXinsertion or for at least 13 kb to
the right of theURA3-ARG4DSB hotspot (supplementary dromic markers, and 9 of these are clearly multiple events (for criteria, see discussion of MJL2902 above). Figure 1 at http://www.genetics.org/supplemental/). In
addition, frequencies of DSBs at theURA3-ARG4DSB Of the remaining 15 tetrads, only 6 have the pattern of hDNA predicted by the DSBR model, and 4 of these hotspot were not substantially altered either by the
pres-ence of the palindromic markers or by the prespres-ence of are CO associated. The 9 other co-PMS tetrads show PMS for both palindromic markers in a single spore; of the drug-resistant insert markers (data not shown).
To examine patterns of allelic gene conversion, ge- these, only 2 contain hDNA in thetransNCO configura-tion that might be produced by topoisomerase-resolu-netic data from two diploid strains, MJL2936 and
MJL2957, were pooled. These two strains differ only by tion of a dHJ intermediate. Thus, as was the case for ectopic recombination, the majority of allelic co-PMS an SphI restriction enzyme site polymorphism (RSP)
upstream ofHIS4that is absent in MJL2957. This RSP products do not have structures predicted by the DSBR model (Figure 1).
was used for physical mapping of exchanges (see below)
and did not have a significant effect on conversion of Exchange is preferentially located near the DSB:In COs that are associated with PMS, as well as those that ura3-EcPal⫹104 or arg4-EcPal⫹9 (Table 2; chi-square
Figure 4.—Position of the ex-change point among CO products. Symbols are as in Figure 2. (A) In the ectopic recombination system, the exchange point can be located in one of three intervals defined by flanking heterology (wavy and checkered boxes) and by the two palindromic markers (lollipops). The size (in kilobases) and fre-quency of exchanges mapped to the interval (percentage of crossover tetrads) are given below the sche-matic; crossover intensity (centi-morgans per kilobase) in each inter-val is given in the bar graph above. The average crossover intensity for chromosome III is 0.48 cM/kb (Cherryet al.2002). Values reflect combined data from MJL2902 and MJL2870. The frequency of ex-change (crossover tetrads/total tet-rads) in intervals I, II, and III is 3%, 14%, and 8% for MJL2902 and 3%, 17%, and 7% for MJL2870. An addi-tional 3% of combined tetrads had an exchange that could have oc-curred in either interval I or interval II; an additional 4% of combined tetrads had an exchange that could have occurred in either interval II or interval III. (B) In the allelic re-combination system, intervals I and III are subdivided by restriction en-zyme site polymorphisms forEcoRI (E) andSphI (S) as described in the text. “Genetic” refers to the fre-quency of tetrads with exchange that mapped to intervals I, II, and III; values reflect combined data from MJL2936 and MJL2957. An additional 5% of tetrads had an exchange that could have occurred in either interval I or interval II, and an additional 3% of tetrads had an exchange that could have occurred in either interval II or interval III. “Molecular” refers to the frequency of tetrads expected to have an exchange in the subintervals Ia, Ib, IIIa, and IIIb, as calculated from physical analysis of DNA from random spores (see text). The bar graph shows the crossover intensity (centimorgans per kilobase) in each of the five intervals. Values for intervals Ia, Ib, IIIa, and IIIb were calculated from molecular data and, for interval II, from tetrad analysis. (C) Physical analysis of DNA isolated from either nourseothricin-resistant (NOU) or hygromycin-B-resistant (HYG) spore colonies of MJL2976. The schematic represents the marker configuration in this strain: one of the homologs is marked bynatMX(horizontally hatched rectangle) and contains
TABLE 3 A substantial fraction of ectopic crossovers occurred without associated gene conversion. About one-third of
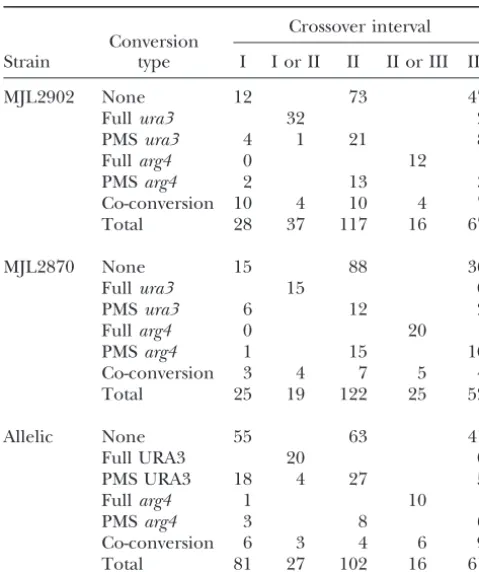
Crossover distributions
total ectopic crossovers (161/508) have an exchange point in interval II without associated gene conversion,
Crossover interval
Conversion and 22% (110/508) of total ectopic crossovers have an Strain type I I or II II II or III III exchange point in intervals I or III without associated
gene conversion. Possible origins for the crossovers
MJL2902 None 12 73 47
Fullura3 32 2 where the exchange point is separated from the most
PMSura3 4 1 21 8 likely initiation (DSB) site by an unconverted marker
Fullarg4 0 12 will be discussed below.
PMSarg4 2 13 3
The flanking intervals in the allelic recombination
Co-conversion 10 4 10 4 7
system are much larger than those in the ectopic system
Total 28 37 117 16 67
(Figure 4, A and B). Interval I is between natMXand ura3-EcPal⫹104(4.1 kb) and interval III is between
arg4-MJL2870 None 15 88 36
Fullura3 15 0 EcPal⫹9 andhphMX (7.5 kb). To allow a more direct
PMSura3 6 12 2 comparison of exchange distributions, we used RSPs
Fullarg4 0 20 flanking theURA3-ARG4interval to subdivide intervals
PMSarg4 1 15 10
I and III. Interval Ia lies betweennatMXand theEcoRI
Co-conversion 3 4 7 5 4
RSP (3.1 kb). Interval Ib is from theEcoRI RSP to
ura3-Total 25 19 122 25 52
EcPal⫹104(1.0 kb). Similarly, intervals IIIa and IIIb are from arg4-EcPal⫹9 to the SphI RSP (2.6 kb) and the
Allelic None 55 63 41
Full URA3 20 0 SphI RSP tohphMX(4.9 kb), respectively (Figure 4B).
PMS URA3 18 4 27 5 This creates anEcoRI-URA3-ARG4-SphI region (intervals
Fullarg4 1 10 Ib, II, and IIIa in Figure 4B) that more closely
approxi-PMSarg4 3 8 6
mates theURA3-ARG4 region of homology examined
Co-conversion 6 3 4 6 9
in ectopic crosses. To show that the two RSPs have no
Total 81 27 102 16 61
effect on conversion of the palindromic markers, we
Intervals I, II, and III are illustrated in Figure 4. Allelic data analyzed tetrads from MJL2957 and MJL2959, two are pooled from MJL2936 and MJL2957, as in Table 2. In strains related to MJL2936. MJL2957 lacks the
SphI RSP
tetrads with full conversion or aberrant 4:4 segregation of
present in MJL2936 and these two strains show similar ura3-EcPal⫹104, COs in intervals I and II cannot be
distin-gene conversion frequencies (Table 2). MJL2959, which
guished; in tetrads with full conversion or aberrant 4:4
segrega-tion ofarg4-EcPal⫹9, COs in intervals II and III cannot be carries only ura3-EcPal⫹104, is heterozygous for the
distinguished. EcoRI site inHIS4.This strain and MJL2936 show similar
gene conversion frequencies for the palindromic marker (Table 2). Thus, neither of the RSPs markedly affects gene verted markers appear to exchange linkage) can be conversion ofura3-EcPal⫹104orarg4-EcPal⫹9.
(102/512) contain crossovers where the exchange maps to the central interval II (Table 3), an exchange density that is more than fivefold greater than the exchange density seen in flanking intervals (Figure 4B). The simi-lar exchange-density maps seen in both ectopic and allelic configurations indicates that the preference for DSB-proximal exchange (COs where the exchange point is between the two palindromic markers) is not due to a restriction of branch migration imposed by the extensive flanking heterology present during the ectopic recombination.
The above conclusion assumes that gene conversion of the RSPs or co-conversion of a RSP with a palindromic marker did not appreciably contribute to recombinants detected by the molecular analysis. We believe that both types of events are rare. For MJL2959, nourseothricin-resistant spore colonies that co-convert theEcoRI RSP andura3-EcPal⫹104will yield a 7.1-kb fragment in the molecular analysis shown in Figure 4D. This band is not detected (⬍0.3% of total DNA). The same 7.1-kb band among hygromycin-B-resistant colonies could be the product of either a full gene conversion event in which the hphMXmarked chromosome gains an SphI
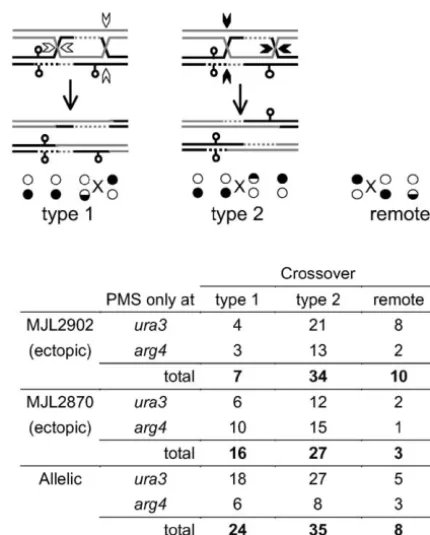
restric-Figure5.—Resolution of the dHJ intermediate. The
inter-tion site or an exchange in interval IIIa or II that is
mediate diagrammed here is for the marker configuration in
associated with co-conversion of ura3-EcPal⫹104 and MJL2902, or the allelic cross, and can be resolved in two ways theEcoRI RSP. This product is present at a level corre- to give rise to CO-associated gene conversion ofarg4-EcPal⫹9. sponding toⵑ1% of tetrads (Figure 4D). If this is taken Type 1 resolution (open arrowheads) cuts the HJ on the con-tinuous stands that do not contain newly synthesized DNA
as the maximum frequency for either gene conversion
and leads to a DSB-distal exchange to the right of arg4-of theSphI RSP or co-conversion ofura3-EcPal⫹104and
EcPal⫹9. Type 2 resolution (solid arrowheads) cuts the HJs
the EcoRI RSP, the results presented in Figure 4B are
on the strands containing newly synthesized DNA and their
not substantially altered. equivalent strands on the other duplex leading to a
DSB-We note that, in the allelic configuration, 11% of COs proximal exchange to the left ofarg4-EcPal⫹9. “Remote” refers to tetrads where an unconverted palindromic maker separates
are estimated to occur in interval Ia and 4% in interval
the crossover from the palindromic marker in hDNA. The
IIIb (Figure 4B). Moreover, one-third (96/290) of COs
table lists the number of tetrads with unidirectional PMS
con-in thenatMX-hphMXregion show exchange in intervals version for eitherura3-EcPal⫹104orarg4-EcPal⫹9with type I and III without associated gene conversion of the two 1, type 2, or remote resolution. The solid and open circles palindromic markers. Possible origins for these cross- represent the genotype of the tetrad colonies where the paren-tal configuration was 䊊䊊䊊䊊/䊉䊉䊉䊉. Circles that are
half-overs will be discussed below.
white represent PMS.Xmarks the exchange point. Contribution of different dHJ resolution modes to
exchange point location:According to the DSBR model (Figure 1), the exchange point in CO products should
gene conversion ofura3-EcPal⫹104, the exchange point always be located at the end of a hDNA tract that starts at
will be either distal (to the left of the marker, type I the DSB itself. In the case of unidirectional conversions,
resolution) or DSB proximal (to the right of the marker, which compose the majority of events that we observed,
type 2 resolution). If the two types of resolution occur one HJ should be located DSB proximal to the
con-at equal frequencies, then an equal number of DSB-verted marker while the other should be DSB distal to
proximal and DSB-distal COs are expected to accom-the converted marker on accom-the side of accom-the converted
pany PMS for a marker. We examined tetrads with CO-marker opposite from the DSB (Figure 5). If we consider
associated PMS of eitherura3-EcPal⫹104orarg4-EcPal⫹9 an intermediate in which only thearg4-EcPal⫹9marker
and determined the position of the exchange point is located in hDNA, COs produced by cutting the stands
relative to the unconverted markers. In both ectopic opposite the newly synthesized DNA (type 1 resolution
and allelic crosses, PMS-associated DSB-proximal COs in Figure 5) results in the exchange point being located
(type 2 resolution) were present in excess of DSB-distal DSB distal, to the right ofarg4-EcPal⫹9. If the junctions
COs (type 1 resolution), but only in one case (MJL2902) are resolved by cutting the strands containing the newly
did proximal vs. distal values differ significantly from synthesized DNA (type 2 resolution in Figure 5), the
equivalence (Fisher’s exact test,P ⬍0.005). However, exchange point will be located proximal to the DSB, to
(96/142) display type 2vs. type 1 resolution, a value and direction of mismatch correction and that gene significantly different from equivalence (Fisher’s exact conversion patterns for such markers accurately reflect test:P⬍0.005) and similar to that calculated for gene underlying patterns of hDNA.
conversion of markers in the HIS4 gene itself (67%; Other studies: Although others have examined the
Fosset al.1999). pattern of gene conversion resulting from meiotic
re-We also observed tetrads where the exchange point combination, each of those studies used a system in is separated from the converted marker by an uncon- which more than one initiation site was present. Both verted marker, but still involves the gene-converted Schultes andSzostak (1990) and Gilbertson and chromatid. For example, two tetrads from MJL2902 Stahl (1996) examined gene conversion of markers show PMS for arg4-EcPal⫹9, with the exchange point flanking the ARG4 DSB hotspot, and both obtained located downstream of the unconvertedura3-EcPal⫹104 evidence for frequent co-conversion. While this has in interval I (Figure 5). Possible origins for this class of been interpreted as indicating that many of the events products will be discussed below. Its frequency (ⵑ4% that initiated at the ARG4 DSB hotspot were bidirec-of conversion tetrads) is not significantly greater than tional, the possibility that some co-conversion events that expected for independent events (ⵑ3% on the were initiated at a second DSB hotspot, locatedⵑ2 kb basis of crossover frequencies in nonconversion tetrads; away in theDED81promoter (Sunet al.1989), cannot P⫽0.15, one-tailed Fisher’s exact test). be eliminated. Furthermore, a substantial fraction of the co-conversions thatGilbertsonandStahl(1996) detected have marker configurations consistent with
DISCUSSION
derivation from events that initiated atDED81and pro-Co-conversion of markers flanking the DSB hotspot is ceeded unidirectionally through the two markers. Gil-infrequent:We examined the pattern of hDNA resulting bertsonandStahl(1996) used poorly corrected palin-from meiotic recombination initiated at a single DSB dromic markers and thus were able to determine the hotspot with little or no expected contribution from pattern of hDNA present in the recombination products initiation events occurring outside the interval. We find of a subset of tetrads. Only a minor fraction of the co-that bidirectional conversion occurs infrequently when PMS tetrads that they observed had the pattern of hDNA the markers examined areⵑ300 and 600 bp from the predicted for events initiated at the
ARG4DSB hotspot DSB hotspot. In both allelic and ectopic recombination,
and proceeded as predicted by the DSBR model in Fig-the fraction of tetrads displaying simultaneous
conver-ure 1. That is, only 4/108 co-PMS tetrads showed PMS sion of ura3-EcPal⫹104 and arg4-EcPal⫹9 was only
for a marker on one side of theARG4DSB in one spore slightly greater than that predicted for independent
and PMS for the marker on the other side of the DSB events (allelic, 8%vs. 6%; ectopic, 6% vs. 3%).
More-in a different spore. over, patterns of hDNA in tetrads with PMS for both
The pattern of co-conversion from recombination markers infrequently corresponded to those predicted
events initiated at theHIS4DSB hotspot has also been by the DSBR model (13/40 and 6/24 for ectopic and
examined.Porteret al.(1993) found that only a small allelic configurations, respectively) while a similar
frac-fraction of events initiated by breaks at this hotspot tion (14/40 and 9/24) were unambiguously the product
resulted in co-conversion of markers 670 bp to the right of multiple events. We therefore believe it likely that, at
and 900 bp to the left of the hotspot. We find that when this recombination hotspot, most co-conversion tetrads
hDNA to one side of the DSB is ⬎600 bp, hDNA on result from independent events rather than from true
the other side does not extend beyond 300 bp. Recently, bidirectional conversion and that the majority of
conver-Merkeret al.(2003) examined co-conversion of mark-sion events (at least 80% and possibly⬎90%) initiated
ers closer to the HIS4DSB hotspot—140 bp to the left at this DSB hotspot are unidirectional, involving gene
and 240 bp to the right. They also observed frequent conversion of only one of the two markers that flank
unidirectional conversion and concluded that initial the initiating DSB.
strand invasion was shorter than the length of resection. In the simplest terms, this finding would imply that
They observed a greater frequency of bidirectional con-heteroduplex DNA formation is similarly
unidirec-version than we did, although some of these events tional. Other interpretations that involve bidirectional
could have been produced either by two independent heteroduplex formation coupled with near-obligate
res-initiations or by events initiated elsewhere. In their toration of markers on one side of the break by a
mis-study, 30% (16/56) of tetrads classified as products of match correction system that isMSH2/MLH1
indepen-single events showed a marker pattern consistent with dent and that corrects palindrome mismatches at high
the distribution of hDNA predicted by the DSBR model. efficiency (Fosset al.1999;Hoffmannet al.2005) have
Since the palindromic markers used byMerker et al. been proposed. While such mechanisms are not
ex-(2003) are located closer to the DSB hotspot than the cluded by current data, we will assume in the remainder
markers used in our study, their detection of more fre-of this discussion that all molecules with a palindrome
inva-Figure6.—Models for meiotic recombination based on our findings as well as those of others. (A) Extensive hDNA is formed by extension of the single-strand tail and second end capture. One end of the resected DSB invades homologous sequences forming a short (⬍300 bp) tract of hDNA and initiates DNA synthesis. NCO products arise through a SDSA pathway where the newly synthesized DNA is displaced and anneals to the other end of the DSB. A final round of DNA synthesis completes the mature NCO product. If the second end of the DSB is captured prior to strand displacement, a dHJ is formed. This intermediate is resolved by the two HJs in opposite directions (open arrowheads) to form CO products; only one of the two resolution modes is shown. A minor pathway for NCO production by unwinding the dHJ intermediate cannot be excluded. (B) Extensive hDNA is formed during initial strand invasion. One of the resected ends invades the homolog to form a tract of hDNA ⬎600 bp. Capture of the second end of the resected DSB leads to formation of the dHJ that is resolved to form CO products as described above. Displacement of the newly synthesized strand in the intermediate shown in step 2 does not form an NCO gene conversion product.
sion is sometimes⬎140 bp, while our data indicate that dromes, which contains the initiating DSB, without asso-ciated conversion of either marker. These findings sup-invasion is most often⬍300 bp.
Relationship between DSB end resection and hDNA port the suggestion that the average amount of hDNA formed during meiotic DSB repair is significantly less formation:5⬘–3⬘resection from theARG4DSB isⵑ450
nt or more in two-thirds of broken chromosomes (Sun than the amount of single-strand DNA formed by resec-tion, a conclusion also supported by electron-micros-et al.1991). AtHIS4, DSBs are resected byⵑ600 nt (Nag
andPetes 1993). We have also examined resection at copy-based measurements of interjunction distances in dHJ intermediates (BellandByers 1983). Below, we the DSB site in our interval and find that the majority
of breaks are resected ⬎500 bp (data not shown). On consider two ways in which this could occur, focusing on events in whicharg4-EcPal⫹9is converted.
the basis of these measures, most of the DSBs at the
hotspot within the URA3-ARG4 interval should be re- Initial strand invasion is short: Figure 6A presents a model in which the single-strand tail containing ura3-sected beyondura3-EcPal⫹104,which isⵑ300 bp from
the DSB. We expect that many are also resected beyond EcPal⫹104invades the homolog and initiates DNA syn-thesis that extends past the site ofarg4-EcPal⫹9. Hetero-arg4-EcPal⫹9, 600 bp from the DSB. Tetrads with
conver-sion ofarg4-EcPal⫹9must have been produced by events duplex DNA will thus be present atarg4-EcPal⫹9in CO products and in NCO products, the latter irrespective where hDNA extended at least that distance. In the
majority of such tetrads, ura3-EcPal⫹104 is not con- of whether they are formed by SDSA (steps 7 and 8 in Figure 6A) or by the unwinding of a dHJ intermediate verted. This indicates that, in single events where hDNA
extends at least 600 bp from the initiating DSB in one (step 6 in Figure 6A). Forura3-EcPal⫹104to be present in hDNA, at least in CO products, the initial strand direction, it rarely extends 300 bp in the other direction.
Additional data consistent with short hDNA tracts are invasion must be⬎300 bp. The frequent failure of this marker to be included in hDNA when arg4-EcPal⫹9 provided by the finding that many (ⵑ30% of ectopic
palin-resection. This conclusion—that the length of resection been suggested that nicks or small gaps at the end of the tract of DNA synthesis might direct junction resolution does not determine the length of strand invasion—was
also reached byMerkeret al.(2003). (Foss et al. 1999). Such nicked or gapped structures (step 3 in Figure 6, A and B) have been proposed as a Strand invasion is long:Figure 6B presents a model in
which initial strand invasion is⬎600 bp and incorpo- substrate for crossover formation by the Mus81/Mms4 nuclease (Osman et al. 2003; Hollingsworth and rates arg4-EcPal⫹9 in hDNA. Parental segregation of
ura3-EcPal⫹104would occur if the tract of DNA synthe- Brill2004). However, this nuclease recognizes the free 5⬘-end at a flap junction or gap and nicks within the sis using the homolog as a template was⬍300 bp. This
latter mechanism is consistent with the structure of the duplex five nucleotides upstream on the flap-containing strand (Bastin-Shanoweret al.2003). While this would single end invasion (SEI) intermediates reported by
Hunter and Kleckner (2001) in that the 3⬘-end of resolve the DSB-distal junction (the HJ to the right of the palindrome in hDNA in Figure 5) in a manner these three-arm intermediates does not show extensive
DNA synthesis. Such intermediates could not produce consistent with our observations, we predict that the DSB-proximal junction (the HJ between the palin-NCO gene conversions of arg4-EcPal⫹9 by an SDSA
mechanism (steps 7 and 8 in Figure 6B), although these dromes in Figure 5) would be much more than five nucleotides from the free 5⬘-end created by resection could still be produced by topoisomerase-mediated dHJ
unwinding (step 6 in Figure 6B), as suggested byGil- (which is at least 500 bp from the DSB; data not shown) and thus should not be a substrate for Mus81/Mms4 bertsonand Stahl(1996). Topoisomerase-mediated
unwinding of dHJ intermediates would produce a single endonuclease cleavage. Furthermore, our observation of a substantial number of type 1 resolutions would meiotic product with markers flanking the DSB site in
atransheteroduplex configuration (Figure 6A, step 6). indicate that nick-directed cutting cannot be the only mechanism for Holliday junction resolution.
Such bidirectional conversion products are detected,
but only in a small minority of total conversion tetrads We observed a few crossover tetrads where the point of exchange was separated from the marker showing (Gilbertson and Stahl 1996; Merker et al. 2003;
HoffmannandBorts2005; this study). Since the ma- PMS by an unconverted marker. The low frequency at which such crossovers are recovered among gene jority of the NCO events that we and others (Porteret
al.1993;Merkeret al.2003) observe appear to include conversion tetrads (only slightly more than expected for two independent events) indicates that neither can only one marker in heteroduplex, our data do not
ad-dress the likelihood of SDSAvs.dHJ unwinding. Argu- be a major pathway for meiotic crossover formation at this locus. However, in a substantial fraction of crossover ments disfavoring dHJ unwinding as the predominant
mechanism of NCO formation are provided by recent tetrads, neither marker displayed conversion and the point of exchange was separated from the DSB hotspot findings that both SEI and dHJ intermediates are on a
crossover-predominant pathway (Hunterand Kleck- by an unconverted marker (in Table 3, 22% of ectopic crossovers and 33% of allelic crossovers, corresponding ner2001;Bo¨ rneret al.2004) and that NCOs form at
wild-type levels in mutants that fail to make either SEIs to 7% and 19% of total tetrads, respectively). There are several possible ways in which such crossovers, which we or dHJs (Bo¨ rneret al.2004) as well as in mutants that
accumulate unresolved dHJs (AllersandLichten2001a; will term remote crossovers, might arise. Using rad50S strains, we did not detect enough DSBs at sites other Clyneet al.2003).
Modes of dHJ resolution: According to the DSBR than the URA3-ARG4 DSB hotspot to account for the number of crossovers observed (see Figure 3 and supple-model, COs are formed by dHJ resolution in which the
two HJs are cut in opposite directions. Two possible CO- mental Figure 1 at http://www.genetics.org/supplemen tal/). However, contributions from DSBs or other le-producing resolution modes are possible. In one (type
1 in Figure 5), strands ending 3⬘ at the original DSB sions not detected in rad50S mutants cannot formally be excluded. Alternatively, remote COs might arise from (and their cognate strands on the other partner) are
cut; in the other (type 2 in Figure 5), strands ending repair of DSBs formed at the URA3-ARG4hotspot ac-companied by bubble migration, strand-displacement-5⬘ at the original DSB are cut, either near or within
regions of newly synthesized DNA (dotted lines in Fig- mediated crossing over, or other mechanisms that move strand exchange structures away from an initiation site ure 5). Thus, the strands cut during formation of a
crossover can be inferred from the position of the ex- without creating intervening hDNA (Allersand Lich-ten2001b;Smith2001;Younget al.2002). Such appar-change point relative to the hDNA formed, and any bias
in resolution will be revealed as an unequal recovery of ent remote COs can also be produced by the DSBR mechanism outlined in Figure 1, if dHJ resolution is the two types of crossovers. The modest bias that we
observe toward type 2 resolution (Figure 5) is in the directed toward type 1 resolution (Figure 5) and mis-matches in hDNA are repaired to restore parental allele same direction as the bias inferred for recombination
in both the HIS4 and ARG4 genes (Gilbertson and ratios (see Foss et al. 1999 for discussion); however, single remote crossovers are produced by this mecha-Stahl 1996; Foss et al. 1999; references within). Its
near-Alani, E., R. PadmoreandN. Kleckner, 1990 Analysis of wild-type
unidirectional and does not include markers on both
and rad50 mutants of yeast suggests an intimate relationship
sides of the initiating DSB. between meiotic chromosome synapsis and recombination. Cell
61:419–436.
Conclusion:On the basis of our findings and those of
Allers, T., andM. Lichten, 2001a Differential timing and control
others, we suggest the following mechanism for meiotic
of noncrossover and crossover recombination during meiosis.
recombination inS. cerevisiae (Figure 6A). A 3⬘ single- Cell106:47–57.
Allers, T., andM. Lichten, 2001b Intermediates of yeast meiotic
strand tail from one end of the DSB invades homologous
recombination contain heteroduplex DNA. Mol. Cell8:225–231.
sequences and initiates DNA synthesis. This initial
inva-Bastin-Shanower, S. A., W. M. Fricke, J. R. MullenandS. J. Brill,
sion is almost always ⬍300 bp and frequently is⬍140 2003 The mechanism of Mus81-Mms4 cleavage site selection
distinguishes it from the homologous endonuclease Rad1-Rad10.
bp; on the other hand, extension of the tail can often
Mol. Cell. Biol.23:3487–3496.
extend ⬎600 bp. The extended tail can be displaced
Bell, L. R., andB. Byers, 1983 Homologous association of
chromo-and anneal with the other end of the resected DSB, somal DNA during yeast meiosis. Cold Spring Harbor Symp.
Quant. Biol.47(Pt 2): 829–840.
yielding a NCO recombinant by SDSA; in such
recombi-Bergerat, A., B. de Massy, D. Gadelle, P. C. Varoutas, A. Nicolas nants hDNA is present on only one side of the DSB
et al., 1997 An atypical topoisomerase II from Archaea with
(Figure 6, A or B). Double Holliday junction intermedi- implications for meiotic recombination. Nature386:414–417. Borde, V., T.-C. WuandM. Lichten, 1999 Use of a recombination
ates will be formed if the second end of the resected
reporter insert to define meiotic recombination domains on
chro-DSB is captured by the initial invasion intermediate
mosomeIIIofSaccharomyces cerevisiae.Mol. Cell. Biol.19:4832–
(step 3 in Figure 6). By nature of the short initial inva- 4842.
Bo¨ rner, G. V., N. KlecknerandN. Hunter, 2004
Crossover/non-sion, very little hDNA is formed at the initial strand
crossover differentiation, synaptonemal complex formation, and
invasion step. The majority of hDNA is formed during
regulatory surveillance at the leptotene/zygotene transition of
second-strand capture. Resolution of the dHJ yields a meiosis. Cell117:29–45.
Borts, R. H., W. Y. Leung, W. Kramer, B. Kramer, M. Williamson CO product in which most of the hDNA is located to
et al., 1990 Mismatch repair-induced meiotic recombination
re-one side of the initiating DSB.
quires thePMS1gene product. Genetics124:573–584.
A second result of the short initial invasion is that Cao, L., E. AlaniandN. Kleckner, 1990 A pathway for generation
and processing of double-strand breaks during meiotic
recombi-one of the HJs in the dHJ intermediate will always be
nation inS. cerevisiae.Cell61:1089–1101.
positioned close to the DSB, while the other HJ position
Cherry, J. M., C. Ball, S. Chervitz, K. Dolinski, S. Dwightet al.,
will be dispersed. Unbiased resolution of these dHJ in- 2002 Saccharomyces genome database (http://genome-www.
stanford.edu/Saccharomyces/).
termediates as COs (equal numbers of type 1 and type
Clyne, R. K., V. L. Katis, L. Jessop, K. R. Benjamin, I. Herskowitz 2 resolution in Figure 5) will produce a population of
et al., 2003 Polo-like kinase Cdc5 promotes chiasmata formation
products in which half contain exchange points near the and cosegregation of sister centromeres at meiosis I. Nat. Cell
Biol.5:480–485.
DSB site, and half contain exchange points at dispersed
Collins, I., andC. S. Newlon, 1994 Meiosis-specific formation of
locations. As a result, exchange points will appear to be
joint DNA molecules containing sequences from homologous
concentrated near the DSB site. Biased junction resolu- chromosomes. Cell76:65–75.
de Massy, B., V. RoccoandA. Nicolas, 1995 The nucleotide
map-tion by preferentially cutting strands that contain newly
ping of DNA double-strand breaks at theCYS3initiation site of
synthesized DNA near the junction (type 2 resolution
meiotic recombination inSaccharomyces cerevisiae. EMBO J.14:
in Figure 5) will result in a further preferential location 4589–4598.
of the exchange point near the initiating DSB. Thus, one Fan, Q. Q., F. Xu, M. A. WhiteandT. D. Petes, 1997 Competition between adjacent meiotic recombination hotspots in the yeast
consequence of unidirectional or near-unidirectional
Saccharomyces cerevisiae.Genetics145:661–670.
hDNA formation will be a genetic map in which cross- Foss, H. M., K. J. HillersandF. W. Stahl, 1999 The conversion over distributions closely parallel those of DSBs (Wu gradient atHIS4ofSaccharomyces cerevisiae. II. A role for mismatch repair directed by biased resolution of the recombinational
inter-andLichten1994) and crossover events appear to be
mediate. Genetics153:573–583.
highly localized, even when the resolution of that map is Gilbertson, L. A., andF. W. Stahl, 1996 A test of the double-strand less than the average length of hDNA formed. Such break repair model for meiotic recombination inSaccharomyces
cerevisiae.Genetics144:27–41.
highly punctuated crossover maps are typically seen at
Goyon, C., andM. Lichten, 1993 Timing of molecular events in
mammalian recombination hotspots (reviewed byKauppi meiosis inSaccharomyces cerevisiae: stable heteroduplex DNA is
et al.2004). Although short gene conversion tracts may formed late in meiotic prophase. Mol. Cell. Biol.13:373–382.
Hoffmann, E. R., andR. H. Borts, 2005 Transevents associated
contribute to this tight crossover localization, it also
with crossovers are revealed inmlh1⌬andmsh2⌬strains in
Saccha-remains possible that it reflects an underlying
unidirec-romyces cerevisiae.Genetics169:1305–1310.
tionality in hDNA formation. Hoffmann, E. R., E. Eriksson, B. J. HerbertandR. H. Borts, 2005
MLH1andMSH2promote the symmetry of double-strand break We thank Eva Hoffman and Rhona Borts for strains and for
commu-repair events at theHIS4hotspot inSaccharomyces cerevisiae. Genet-nicating results in advance of publication. We also thank Eva Hoffman,
ics169:1291–1303. Rhona Borts, Tom Petes, Cyril Buhler, Robert Shroff, Gerry Smith,
Hollingsworth, N. M., andS. J. Brill, 2004 The Mus81 solution and three anonymous reviewers for discussions and comments that to resolution: generating meiotic crossovers without Holliday improved this manuscript. junctions. Genes Dev.18:117–125.
Hunter, N., andN. Kleckner, 2001 The single-end invasion: an asymmetric intermediate at the strand break to double-Holliday junction transition of meiotic recombination. Cell106:
LITERATURE CITED
59–70.
Kane, S. M., andR. Roth, 1974 Carbohydrate metabolism during
Abdullah, M. F., andR. H. Borts, 2001 Meiotic recombination
ascospore development in yeast. J. Bacteriol.118:8–14. frequencies are affected by nutritional states inSaccharomyces
are: recombination distributions in mammals. Nat. Rev. Genet. Schultes, N. P., andJ. W. Szostak, 1990 Decreasing gradients of gene conversion on both sides of the initiation site for meiotic
5:413–424.
Keeney, S., C. N. GirouxandN. Kleckner, 1997 Meiosis-specific recombination at theARG4locus in yeast. Genetics126:813–822.
Schwacha, A., andN. Kleckner, 1994 Identification of joint mole-DNA double-strand breaks are catalyzed by Spo11, a member of
a widely conserved protein family. Cell88:375–384. cules that form frequently between homologs but rarely between sister chromatids during yeast meiosis. Cell76:51–63.
Liu, J., T. C. WuandM. Lichten, 1995 The location and structure
Smith, G. R., 2001 Homologous recombination near and far from of double-strand DNA breaks induced during yeast meiosis:
evi-DNA breaks: alternative roles and contrasting views. Annu. Rev. dence for a covalently linked DNA-protein intermediate. EMBO
Genet.35:243–274. J.14:4599–4608.
Sun, H., D. Treco, N. P. SchultesandJ. W. Szostak, 1989
Double-Merker, J. D., M. Dominskaand T. D. Petes, 2003 Patterns of
strand breaks at an initiation site for meiotic gene conversion. heteroduplex formation associated with the initiation of meiotic
Nature338:87–90. recombination in the yeastSaccharomyces cerevisiae.Genetics165:
Sun, H., D. TrecoandJ. W. Szostak, 1991 Extensive 3⬘ -overhang-47–63.
ing, single-stranded DNA associated with the meiosis-specific
dou-Nag, D. K., and T. D. Petes, 1993 Physical detection of
hetero-ble-strand breaks at the ARG4 recombination initiation site. Cell duplexes during meiotic recombination in the yeastSaccharomyces
64:1155–1161.
cerevisiae.Mol. Cell. Biol.13:2324–2331.
Szostak, J. W., T. L. Orr-Weaver, R. J. RothsteinandF. W. Stahl,
Nag, D. K., M. A. WhiteandT. D. Petes, 1989 Palindromic
se-1983 The double-strand-break repair model for recombination. quences in heteroduplex DNA inhibit mismatch repair in yeast.
Cell33:25–35. Nature340:318–320.
Trueheart, J., J. D. BoekeandG. R. Fink, 1987 Two genes required
Osman, F., J. Dixon, C. L. DoeandM. C. Whitby, 2003 Generating
for cell fusion during yeast conjugation: evidence for a phero-crossovers by resolution of nicked Holliday junctions: a role for
mone-induced surface protein. Mol. Cell. Biol.7:2316–2328. Mus81-Eme1 in meiosis. Mol. Cell12:761–774.
White, M. A., M. DominskaandT. D. Petes, 1993 Transcription
Paques, F., andJ. E. Haber, 1999 Multiple pathways of
recombina-factors are required for the meiotic recombination hotspot at tion induced by double-strand breaks inSaccharomyces cerevisiae.
theHIS4locus inSaccharomyces cerevisiae.Proc. Natl. Acad. Sci. Microbiol. Mol. Biol. Rev.63:349–404.
USA90:6621–6625.
Petes, T. D., R. E. MaloneandL. S. Symington, 1991
Recombina-Wu, T.-C., andM. Lichten, 1994 Meiosis-induced double-strand tion in yeast, pp. 407–521 in The Molecular and Cellular Biology
break sites determined by yeast chromatin structure. Science263:
of the Yeast Saccharomyces: Genome Dynamics, Protein Synthesis, and 515–518.
Energetics, edited by J.Broach, E.Jonesand J.Pringle. Cold Wu, T. C., andM. Lichten, 1995 Factors that affect the location Spring Harbor Laboratory Press, Cold Spring Harbor, NY. and frequency of meiosis-induced double-strand breaks in
Sac-Petronczki, M., M. F. SiomosandK. Nasmyth, 2003 Un me´nage charomyces cerevisiae.Genetics140:55–66.
a` quatre: the molecular biology of chromosome segregation in Xu, F., andT. D. Petes, 1996 Fine-structure mapping of meiosis-meiosis. Cell112:423–440. specific double-strand DNA breaks at a recombination hotspot
Porter, S. E., M. A. WhiteandT. D. Petes, 1993 Genetic evidence associated with an insertion of telomeric sequences upstream of that the meiotic recombination hotspot at the HIS4 locus of theHIS4locus in yeast. Genetics143:1115–1125.
Saccharomyces cerevisiaedoes not represent a site for a symmetrically Young, J. A., R. W. Schreckhise, W. W. SteinerandG. R. Smith, processed double-strand break. Genetics134:5–19. 2002 Meiotic recombination remote from prominent DNA
Sambrook, J., andD. W. Russell, 2001 Molecular Cloning: A Labora- break sites inS. pombe. Mol. Cell9:253–263. tory Manual. Cold Spring Harbor Laboratory Press, Cold Spring