Copyright 0 1996 by the Genetics Society of America
Complementation Mapping
of
Skeletal
and
Central Nervous System
Abnormalities in Mice
of the
piebald
Deletion Complex
Timothy
P. O'Brien, Danika L.
Metallinos,' Hsiupei Chen, Myung K. Shin
and
Shirley M.
Tilghman
Department of Molecular Biology and The Howard Hughes Medical Institute, Princeton University, Princeton, New Jersey 08544 Manuscript received December 19, 1995
Accepted for publication February 9, 1996
ABSTRACT The sJ5DftM6 $6Pu6 S I A q and ?4Pub
piebald deletion alleles belong to a set of overlapping deficiencies on
the distal portion of chromosome 14. Molecular analysis was used to define the extent of the deletions.
Mice homozygous for the smallest deletion, s15DffM6, die shortly after delivery and display alterations in the central nervous system, including hydrocephalus and a dorsally restricted malformation of the spinal cord. These mice also display homeotic transformations of vertebrae in the midthoracic and lumbar regions. Homozygous s~~~~~ mice contain a point mutation in the piebald gene, survive to weaning, and display no central nervous system or skeletal defects, arguing that the s'5D"M6 phenotype results from the loss of genes in addition to piebald. A larger deletion, ?jfi6, exhibits additional cartilage malformations
and defects in the anterior axial and cranial skeleton. The skeletal defects in both s15DffMb and s'6w mice
resemble transformations associated with the targeted disruption of Hox genes and genes encoding the
retinoic acid receptors, which play a role in the specification of segmental identity along the anteroposter- ior axis. Complementation analysis of the s'5D'fMb and s'6R* phenotypes, using two additional deletions,
localized the gene(s) associated with each phenotype to a defined chromosomal region.
0
NE approach to understanding the molecular mechanisms controlling mammalian develop- ment relies upon studying the phenotypic conse- quences of altered gene activity. Genetic screens pro- vide a powerful method to reveal genes involved in specific developmental processes. Although widely suc- cessful in other organisms, practical considerations have limited the use of such screens in the mouse. One notable exception was the specific locus test (SLT), a large scale mutagenesis effort carried out over the last 40 years at the Oak Ridge National Laboratory and Harwell, England (RUSSELL 1951). In the SLT a large number of radiation- and chemically induced alleles were recovered surrounding seven viable recessive mu- tations resulting in phenotypes that could be scored visually. The mutations ranged in size from point muta- tions to large deletions covering as much as 10 cM of DNA (reviewed in RINCHIK and RUSSELL 1990).One of the target genes in the SLT was the piebald
( s ) gene on the distal portion of chromosome 14.
It has recently been established that s encodes the endothelin-B receptor (EDNRB), a G protein-coupled seven-transmembrane domain receptor that responds to the endothelin family of small peptide growth fac-
tors ( HOSODA et al. 1994). Animals carrying the severe
spontaneous piebald lethal (s') allele, a null mutation
Cmresponding author: Shirley M. Tilghman, Department of Molecu- lar Biology, Princeton University, Princeton, NJ 08544.
E-mail: [email protected]
'
Present address; Department of Molecular and Cell Biology, Univer- sity of California, Berkeley, C A 94720 and School of Veterinary Medi- cine, University of California, Davis, CA 95616.at s, almost completely lack melanocytes and invari- ably die as juveniles of megacolon resulting from a deficiency in enteric ganglia (LANE 1966; HOSODA et al. 1994). The SLT utilized a weak s allele, which is fully viable because the enteric ganglia are unaffected. However, they exhibit a spotted coat, due to a partial loss of melanocytes.
As homozygotes, the induced alleles of piebald have been grouped into two classes: those that survive to weaning and are phenotypically indistinguishable from s', and those that are not recovered at weaning. When DNAs isolated from mice in these phenotypic classes were examined with molecular markers surrounding s, it was evident that the alleles from the first (SI) class were not deleted for any of the molecular markers, whereas those in the second class were typically deleted for one or more of the markers. Thus, the more severe phenotypes likely result from the deletion of essential gene(s) in addition to s (METALLINOS et al. 1994).
This study focuses on slZnttMb and ?6Pub, two overlap
ping chromosomal deletions at s. Phenotypic analyses demonstrate that mice homozygous for either the
or ?6Pub deletion allele die shortly after birth and
display central nervous system (CNS) abnormalities. In addition, changes in the identity of thoracic and lumbar vertebrae are observed in s15Dt1Mb homozygotes, while the ?6Pub homozygotes exhibit cartilage malformations
and defects in the anterior axial and cranial skeleton. With the goal of identifying the genes associated with the developmental defects, molecular and genetic anal- ysis was performed to map the distinct phenotypes to defined DNA intervals.
s15DttMb
448 T. P. O'Brien et al.
A
C
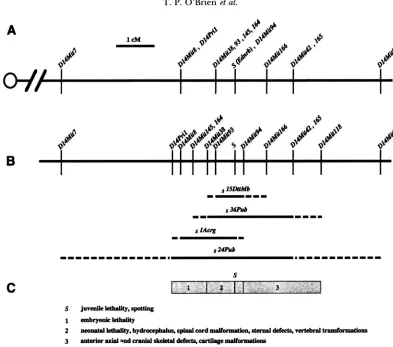
FIGURE 1 .-The piebald deletion complex. (A) Genetic map. Centromere is represented on the left. Molecular markers mapped relative to s (METALLINOS et al. 1994 and data not shown) are indicated with their distance from s in cM. (B) Molecular map. The order of the molecular markers determined on the basis of their presence or absence in the deletion alleles is shown on the first line. The confirmed extent of each deletion is depicted by the closed line. The dashed line represents the region containing the deletion breakpoints. Molecular markers at the end of the dashed line are present in the deletion allele. (C) Functional map. The shaded boxes represent the regions associated with a specific developmental defect(s). The numbers in each box match the listed defect(s) associated with that interval. The extent of a functional interval is defined by the deletion breakpoints based on the complementation analysis. The phenotype associated with the loss of s (Ednrb) gene function is represented as a solid black line and marked S.
MATERIALS AND METHODS
Mice: The induced s alleles were generated at the Oak
Ridge National Laboratory as described previously (RUSSELL 1951) and maintained as heterozygotes by crossing to (101/ R1 X C3Hf/R1) F, mice. The presence of the mutant allele was verified through subsequent test crosses to the Oak Ridge s/s stock. The s ~ ~ " " ~ ' ,
F"'*
and87m
heterozygous stocks ob-tained from Oak Ridge were expanded by matin with SSL/ Le s/s mice from The Jackson Laboratory. The SI' IMb/s, ?61'"b/ s and ?71w/s mice were then crossed to CAST/Ei/s heterozy- gotes to introduce polymorphisms useful for the molecular analysis (METALLINOS et al. 1994). The P/CAST/Ei and P/s
progeny of each of those crosses were intercrossed to enerate the embryos analyzed in Tables 1-3. The dAqand
8
'
"'
alleles were obtained from Oak Ridge as heterozygous stocks with s following the test cross. These mice have been maintained as a closed colony.Embryos and fetuses collected for the histological and skeletal analysis were obtained from timed matings where noon of the day for the detection of a vaginal plug was designated 0.5 dpc.
Genotype analysis: Mice were genotyped using D l 4Mit mi-
crosatellite (CA), markers (DIETRICH et al. 1992, 1994) by PCR using conditions previously described (METALLINOS et
8
F
al. 1994). The ,'"""s ?m and
8
"
mice and intercross progeny were scored using D14Mit8, which maps 1.3 cM proximal to s and generates reaction products that distin- guish the s (194 bp), CAST/Ei (209 bp) and 101/C3H (202bp) chromosomes. This marker is absent in
8'^';
therefore, com ound heterozygotes between and either s ~ ~ " ' ~ ~ ~ ' or?""'were identified by the presence of only the 101/C3H- specific reaction product from the s'5DffMb or ?6pu6 chromo-
some. Progeny from the s~~~~~~~ and
.?"'
crosses with sIAq were genotyped using D14Mit8 as well as D14Mit94. The latter marker cosegregates with s (Figure 1A) and generates reaction products that distinguish the s (109 bp), CAST/Ei (79 bp) and 101/C3H (107 bp) chromosomes. This marker is absent in both s ' ~ " ~ ~ ~ ~ and Therefore, com ound heterozygotes between slAq and either s ~ " " ~ ~ ~ and2'''
dis- played only the 101/C3H-specific reaction product from the sIAq chromosome.Genetic mapping: The microsatellite (CA), markers
Analysis of piebald Deletion Mice
TABLE 1
Distribution of genotypes in offspring from -'IS and s36m heterozygous intercrosses
Genotype No. of
Age No. of litters PUPS
+/+
15DttMb/+ 15DttMb/15DttMb3-4 wk 42 208 74 (36) 134 (64) 0
18.5 dpc 16 118 26 (22) 61 (52) 31 (26)
+/+
36Pub/+
36Pub/36Pub3-4 wk 45 223 69 (31) 154 (69) 0
18.5 dpc 9 64 15 (23) 33 (52) 16 (25)
Values in parentheses are percentages.
an EcoRI restriction fragment length polymorphism (RFLP) between the parental strains, SSL/Le and CAST/Ei.
Molecular anah/sis of the induced s alleles: The genetic mark- ers were scored in genomic DNA preparations from ?"*, d f D f f m
and .?6n' homozygous animals or sIAq/CAST/Ei, and ?'""/ CAST/Ei heterozygotes to define the molecular limits of the deletions. The presence of D14Mitll8in ?6m homozygotes was confirmed by Southern blot analysis using a radiolabeled probe derived from sequences outside the D14Mit118 ( C A ) , repeat. The presence of D14Pnl in d A n R was confirmed by Southern blot analysis of dA*/CAST/Ei DNA, using the EcoRI
RFLP.
The deletion of the Ednrb gene in 515D11Mb and ?6Pub was determined by PCR and Southern blot analysis using DNA from homozygous animals. Primers specific for the extracel- lular (5'-GGAGCAAGCTGTAACATGCAATCGCG3' forward
and 5'-GATCTCCCACAGCCAGACTGGCGATG3' reverse),
transmembrane (5'-ATATCGAGCTGTTGCTTCTTGGAG3'
forward and 5'-CTGCATGAAGGCTGTTTCTGAAAGG3' reverse) and intracellular (5"TCGTGT'ITGTGCTGCTGGT-
GCCA-3' forward and 5'-TITGGCAAAGGTTTCAT"GT-
T"-3' reverse) domains of the protein were employed. A 1421-bp cDNA fragment representing the entire coding re- gion of the Ednrb gene was used as a probe for Southern blot analysis. The blot was stripped and rehybridized with a P-actin cDNA probe to demonstrate that equivalent amounts of DNA from each sample were analyzed.
Histological and skeletal analysis Specimens were rinsed in phosphate-buffered saline (PBS), fixed in Bouin's solution, dehydrated in ethanol, cleared in xylene and embedded in Paraplast (Oxford). Serial 8-pm sections were stained with Mayer's hematoxylin (Sigma) and eosin (Sigma).
Skeletons were prepared essentially as described in KESSEL
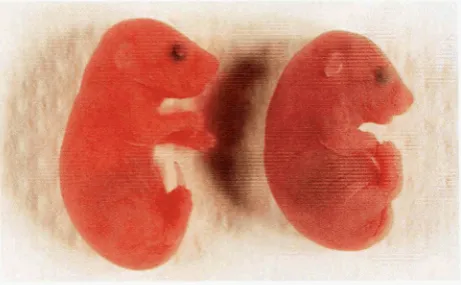
FIGURE 2.-characteristic morphology of 515,ulfM* mutants.
A control littermate (left) and 5lzDflMb homozygote (right) at
18.5 dpc shortly after delivery by Cesarean section.
449
and GRUSS (1991). Fetuses (18.5 dpc) or neonates were skinned, eviscerated and fixed in 100% ethanol for 4 days then placed in 100% acetone for 3 days and rinsed with water. They were stained for 10 days in a solution consisting of 2 ~010.14% alcian blue 8GX (Sigma) in 70% ethanol, 1 vol 0.12% alizarin red S (Sigma) in 95% ethanol, 8 vol 100% glacial acetic acid and 50 ~0170% ethanol. After staining, specimens were macer- ated in a solution of 1 % KOH in 20% glycerol at 37" overnight, then placed at room temperature until adequate tissue was removed for viewing (-1 -2 weeks). The specimens were then cleared, stored and photographed in a solution consisting of a 2:2:1 mixture of ethanokglycero1:benzyl alcohol.
RESULTS
Molecular analysis of the induced piebald alleles: Three of the induced alleles at piebald used in this study had been shown previously to contain deletions of DNA markers surrounding s
(?'""',
slAog and s'~~~), while twoof the alleles (s15D'1Mb and ?7p"b) were not deleted for
any of the markers tested (METALLINOS et al. 1994). The extent of the deletions was further defined by testing DNA for the presence of additional microsatellite mark- ers (DIETRICH et al. 1994) that mapped to the s region, based on their segregation in a 613-animal inter subspe- cific backcross (METALLINOS et al. 1994; Figure 1A). D14Mit145, D14Mitl64 and D14Mit93 cosegregated with D14Mit38, which had been mapped 0.5 cM proxi- mal to s. D14Mit94 could not be separated from s in 613 animals analyzed, while D l 4 M i t l 6 6 m a p p e d 0.8 cM distal to s, and D14Mit165 cosegregated with D14Mit42 at a position 1.5 cM distal to s (Figure 1A).
These markers were then use to analyze DNA derived
CAST/Ei and S ' ~ ~ ~ / C A S T / E ~ heterozygotes. This analy- sis established that s15Dt1Mb was also a deletion that re- moved both D14Mit93 and D14Mit94.
As
D14Mit145, D14Mit164 and Dl4Mit38 were present in s15D1tMb DNA, D14Mit93 was established as the most distal marker in the group and the proximal boundary of the deletion (Figure 1B). The amplification of Dl4Mit145 and D14Mit164, but not D14Mit38in s;'6""b homozygous DNAordered these markers, as indicated in Figure lB, and defined the proximal boundary of the deletion. Finally, the d A o g chromosome was deleted for all the
450 T. P. O'Brien et al.
. .. . . . _ . . ..-. - _ -
s15Dt1Mb FIGURE
3.-Hydrocephalus in s'5DLmb homozygotes. (A) Frontal section through the head of a control littermate and (B) a
mutant at 18.5 dpc. There is a moderate expansion (arrows in B) of the superior part of the body of the right lateral ventricle (lv) and the third ventricle (tv). ( C ) Sagittal section through a control littermate and (D) a s15Dfmb mutant at 18.5 dpc
reveals that the medulla oblongata (m) has been repositioned such that it occludes (arrowhead in D) the subarachnoid space
(ss) along a region just below the thin layered roof of the fourth ventricle (rfv). fv, fourth ventricle; c, cerebellum.
markers that mapped at 0.5 cM, along with D14Mit8, which maps 1.3 cM proximal to s. However a YAGde- rived DNA marker, D14Pri1, that cosegregated with D14Mit8 was present on the slAWg chromosome (Figure 1B). s~~~~~ was confirmed as the largest proximal dele- tion, as it was deleted for both D14Mit8 and D14Pril.
The distal breakpoint of s15D1Mb was localized be- tween Dl4Mit94 and D14Mit166, while that of ?hpub
extended beyond D14Mit42 to D14Mit118. The distal breakpoint of d A m g occurred between Ednrb and
D14Mit94, placing Dl4Mit94 distal to Ednrb. Finally, the absence of both D14Mit42 and D14Mit165 in s24pub establishes this deletion as the largest among the group on both proximal and distal ends. These analy- ses establish the minimum size of s~~~~~~~ at 0.5 cM,
Analysis of piebald Deletion Mice
~ 1 5 D t t M b / ~ l S D t t M b
451
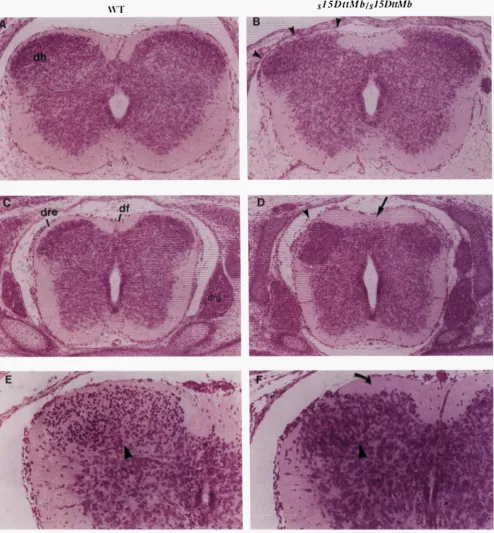
FIGURE 4.-Organization of the dorsal region of the developing spinal cord is erturbed 111 the sz5D'1Mb homozygotes. (A)
Transverse section through the cervical spinal cord of a normal littermate and a &'ItMb mutant (B) at 14.5 dpc. The cellular
boundary of the dorsal horn (dh) is extended both dorsomedially and dorsolaterally (arrowheads in B) at the expense of the
surrounding white matter (funicular zone). (C) The spinal cord at the thoracic level in a 14.5 dpc control littermate and a
mutant (D). The clearly demarcated dorsal root entrance zone (dre) in the control animal is eclipsed by cells (arrowhead
in D) in the mutant, while the dorsal margin of the dorsal horn is flattened against a thickened overlying funicular zone (region
between arrowhead and arrow in D). The dorsal funiculus ( d f ) is not clear in the mutant while additional cells are scattered throughout the region (arrow in D). (E) Transverse sections through the cervical spinal cord of a control littermate and a s'5D'1Mb
mutant ( F ) at 16.5 dpc demonstrates the altered morphology of cells of the dorsal horn that extend dorsolaterally and remain
flattened along the thickened funicular zone (curved arrow in F)
.
In addition, a smaller, darker staining population of furtherdifferentiated cells (arrowhead in E) in the control animal is not apparent in the dorsal horn of the mutant (arrowhead in F).
452 T. P. O'Brien et al.
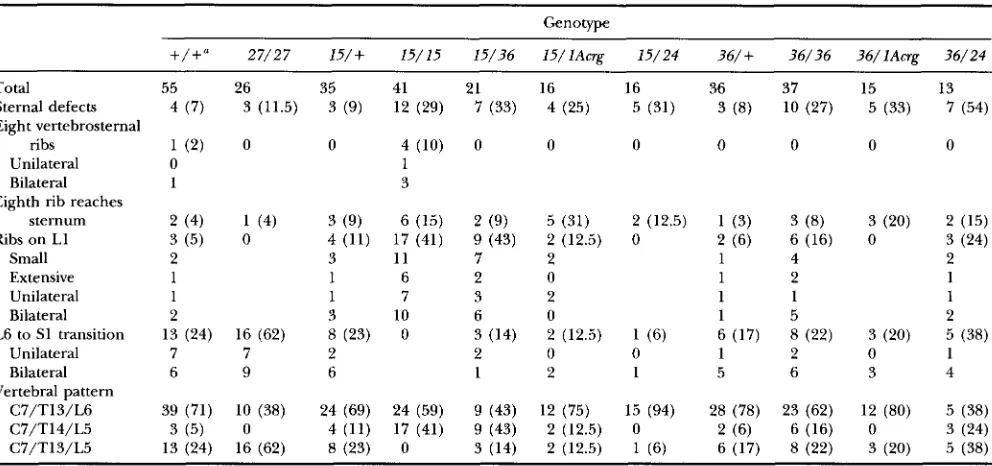
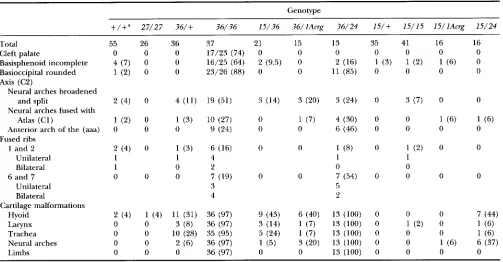
TABLE 2
Incidence of skeletal abnormalities associated with d5-* homozygotes
Genotype
27/27 15/+ 15/15 15/36 1 5 / l A w ~ 15/24 36/+ 36/36 36/lAcrp 36/24
Total 55 26 35 41 21 16 16 36 37 15 13
Sternal defects 4 ( 7 ) 3 (11.5) 3 (9) 12 (29) 7 (33) 4 (25) 5 (31) 3 (8) 10 (27) 5 (33) 7 (54)
Eight vertebrosternal
ribs 1 (2) 0 0 4 (10) 0 0 0 0 0 0 0
Unilateral 0 1
Bilateral 1 3
Eighth rib reaches
sternum 2 (4) 1 (4) 3 (9) 6 (15) 2 (9) 5 (31) 2 (12.5) 1 (3) 3 (8) 3 (20) 2 (15)
Ribs on LI 3 (5) 0 4 (11) 17 (41) 9 (43) 2 (12.5) 0 2 (6) 6 (16) 0 3 (24)
Small 2 3 11 7 2 1 4 2
Extensive 1 1 6 2 0 1 2 1
Unilateral 1 1 7 3 2 1 1 1
Bilateral 2 3 10 6 0 1 5 2
Unilateral 7 7 2 2 0 0 1 2 0 1
Bilateral 6 9 6 1 2 1 5 6 3 4
C7/T13/L6 39 (71) 10 (38) 24 (69) 24 (59) 9 (43) 12 (75) 15 (94) 28 (78) 23 (62) 12 (80) 5 (38)
C7/T14/L5 3 (5) 0 4 (11) 17 (41) 9 (43) 2 (12.5) 0 2 (6) 6 (16) 0 3 (24)
C7/T13/L5 13 (24) 16 (62) 8 (23) 0 3 (14) 2 (12.5) 1 (6) 6 (17) 8 (22) 3 (20) 5 (38)
+,
s or Cast/Ei; 27,s'"4.
15, s15Dt'Mb; 36, .?6m; lAcr , dAT; 24, ?4pu6. Values in parentheses are percentages."+/+,
Littermates from the s ' ~ " ' ~ * / + Xd6hb/+,
JDttMb/+
and .?6pu"/+ intercrosses used to generate the mutant offspringfor the skeletal analysis and genotyped as Cast/Ei/Cast/Ei, Cast/Ei/s and s/s using the D14Mit8 molecular marker. These control littermates were pooled as no significant differences were observed in the analysis of individual populations.
L6 to S1 transition 13 (24) 16 (62) 8 (23) 0 3 (14) 2 (12.5) 1 (6) 6 (17) 8 (22) 3 (20) 5 (38)
Vertebral pattern
The status of the piebald gene was examined by PCR analysis using primers directed against the coding re- gions from the extracellular, transmembrane or intra- cellular domains of EDNRB. These primers failed to generate specific products from s15DttMh, ?6p"b and slAng
homozygous DNA (data not shown). Southern blot analysis using an Ednrb cDNA confirmed the PCR results
additional information that both deletions remove the entire Ednrb coding region (data not shown). In con- trast, all of the molecular markers tested, including
Ednrb, were present in ?7Pub homozygous DNA. Se-
quence analysis of the ?7Pub allele revealed that it con-
tains a point mutation in a highly conserved amino acid in the Ednrb gene (M. K. SHIN and S. M. TILGHMAN, unpublished results).
and
d6fib
deletion mutants die at birth: The progeny from intercrosses of s~~~~~~~ or ?6pub heterozy-gotes were genotyped at 3-4 weeks after birth (Table 1). N o homozygous offspring were observed. However, litters delivered by Cesarean section at 18.5 dpc con-
25% frequency (Table 1). The mutant fetuses were alive, made gasping attempts to breathe and responded to a prodding stimulus. However, they rapidly became cyanotic and died within 30 min, presumably of respira- tory failure. The s15DttMb and offspring were consis- tently smaller, weighing on average 14 and 22% less than their littermates, respectively. They also exhibited several characteristic external features, including a pro- for s15DtlMb and
?@*
homozygous DNA and providedsJ5iD"Mb
tained s15DltMb and ?6fih homozygotes at the expected
nounced curvature of the body axis, a swelled dome- shaped head that is tilted downward, and forelimbs that appear depressed against the chest (Figure 2).
Central nervous system abnormalities in sJ5iD"Mb and
d6fib
mutants: Progeny from s15DttMh and inter-crosses were subjected to histological analysis spanning the period from midgestation to birth. No gross abnor- malities in the development of the heart or lungs were observed. However, coronal sections through the head of both slfDttMb and ?6pub homozygotes revealed congeni-
tal hydrocephalus, consistent with their swelled and dome-shaped appearance (Figure
2).
The dilation was most prominent in the lateral and third ventricles (Fig- ure 3, A and B).
Sagittal sections revealed that the brain appeared compressed within the cranial vault. This re- sulted in a narrowing of the ventricular passages, al- though there was no apparent obstruction. In addition, the medulla appeared to be repositioned upward to- ward the osteogenic membrane, thereby occluding the subarachnoid space (Figure 3, C and D). This occurred just below the roof of the fourth ventricle wherecerebro-spinal fluid passes out of the brain.
and ?6pub mutants also displayed an alter- ation in the organization of the dorsal region of the developing spinal cord that was first detectable at 12.5 dpc and was most prominent at 14.5 dpc (Figure 4). In the cervical region a population of cells completely eclipsed the white matter (funicular zone; arrowheads in Figure 4B). Throughout the length of the spinal cord the dorsal architecture was misshapen (Figure 4D), dis-
Analysis of piebald Deletion Mice
SISDttMblSlSDttMb
453
IF"
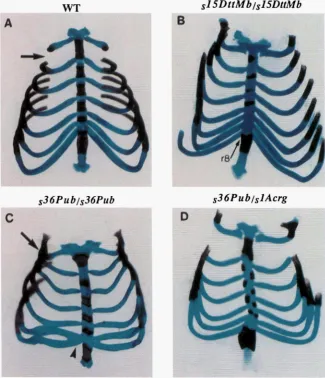
FIGURE 5.-Skeletal abnormalities af-
fectin the ribs and sternum in sZ5-
and
fm
homozygotes. (A) Ventral viewof the sternum region from a 18.5dpc control littermate, displaying the attach- ment of seven ribs and the normal forma- tion of (from top to bottom) the manu- brium sterni, four sternebrae and
xi hoid process. (B) Sternum from szLb mutant showing a T8 to T7 ante- rior transformation, illustrated by the at-
tachment of the eighth pair of ribs to
the sternum (r8). The manubrium sterni and first three sternebrae are reduced while the fourth sternebra is absent. (C) Sternum from mutant displaying the characteristic close juxtaposition of the first and second ribs (compare arrow in A with arrow in C) and the fusion of the sixth and seventh ribs (arrowhead in C)
.
(D) Sternum from ?-/sIAW mutant showing misalignment of the ribs leading to disorganization of the sternebrae.playing an expanded funicular zone as cells appeared to flatten along its ventral margin. These cells then extended dorsolaterally to displace the funicular zone. The formation of the dorsal funiculus appeared altered in the s~~~~~~ mutants (arrow in Figure 4D), possibly reflecting a developmental retardation in the ventral recession of the central canal. In addition, an increased number of cells were scattered throughout the dorsal funiculus. At 16.5 dpc a population of dark-staining smaller cells located in the dorsal horn of normal lit- termates were absent in the slZDnMb and
gfifib
mutants (Figure 4, E and F). Cells of comparable morphology were present in the dorsal horn of mutants by 18.5 dpc, however, they were not as widely dispersed throughout the region. Other cell types that arise from the dorsal spinal cord seemed unaffected. For example, the dorsal root ganglia that derive from neural crest cells appeared to be properly formed.Anterior transformations in the skeleton of s15DiDttMb
mutants The pronounced curvature of the body axis in all mutant animals, along with the occasional obser- vation of a concave depression of the thoracic basket, prompted us to analyze skeletons from s15D1iMb mutants.
In the sternum there were defects in 29% of the animals examined (Table
2
and Figure 5). The range of defects included a reduction of the manubrium sterni and re- maining sternebrae, with the occasional absence of the fourth sternebra. There was asymmetry in the attach- ment of the ribs, leading to a disorganized ossification pattern between the sternocostal junctions (Figure5D).
Defects in the formation of the xiphoid process were noted as a split in its ossification centers (Figure 6C).
454
W T
T. P. O'Brien et al.
SlSDttMbJslSDttMb
S I
SDttMblslSDttMb
A
B
, '
FIGURE 6.-Transformation of lumbar vertebrae in s15D1LMb homozygotes. (A) Ventral view of thoracic and lumbar region from 1 8 . 5 4 ~ ~ control littermate demonstrating the absence of ribs on the first lumbar vertebra (Ll) and the unilateral fusion (arrowhead in A) between the transverse processes of the sixth lumbar (L6) and the first sacral vertebra. This control skeleton provides an asymmetric example of a normal variation in vertebral pattern in which there are only five lumbar vertebrae (C7/ T13/L5). (B) A s15DtfMb mutant showing normal L1 and L6 vertebrae. (C) A slCDnM6 mutant showing the anterior transformation of L1, illustrated by an incomplete bilateral set of ribs on "Ll" with a normal "L6" resulting in an altered (C7/T14/L5) vertebral pattern. The ossification centers of the xiphoid process (x) are split (white arrow in C) in this mutant.
(Figure 6C), resulting in a redistribution of vertebral pattern from C7/T13/L6 to C7/T14/L5 (Table
2).
There is a natural variation in mice for the presence of only 25 presacral vertebrae (C7/T13/L5) resulting from the fusion of the L6 and S1 transverse processes (GR~NEBERG 1952; KESSEL and GRUSS 1991;
LE
MOUEL
MC et al. 1992; S ~ ~ ~ A L L and POTTER 1993; SATOKATA etal. 1995). This was observed in the wild-type (24%) and
study (Table 2 and Figure 6A). However, s15D11Mb mutants never demonstrated a L6 to S1 transition (Figure 6, B and C). Consistent with the other skeletal transforma- tions, the absence of an L6 to S1 transition represents an anteriorization of vertebral identity, relative to con- trol littermates.
Skeletal defects in
d6Pub
mutants: A subset of the skel- etal malformations identified in the s~~~~~~~ mutants were also found in ?6'"b homozygous mutants (Table2). The ?6p"6 mutants displayed sternal defects and an
extra set of ribs on L1, although at a reduced frequency. However, two of the changes in vertebral identity associ- ated with sJSDtuM6 mutants, eight vertebrosternal ribs and invariant number of presacral vertebrae were not ob- served. Instead, a different set of skeletal defects not found in the s15Dtmb homozygotes was observed in the anterior axial and cranial skeleton in 86% of the mutants (Table 3).
The cranial skeletal defects included cleft palate. The secondary palate is composed of the maxillary and pala-
s15DttMb heterozygous littermates (23%) examined in this
tine bones that together form the palatal shelves. A ventral view of the ?6p"b mutant skulls revealed these
structures had not formed properly (Figure 7). By 14.5 dpc in normal embryos, the palatal shelves have rotated and elevated to fuse with each other and the nasal s e p tum. Coronal sections through the head of
s"""
mu- tants at 14.5 dpc demonstrated that the palatal shelves had neither rotated nor elevated (Figure7,
C and D). As late as 18.5 dpc, the shelves remained positioned vertically. A ventral view of the skull exposed additional defects, including a misshapen basioccipital bone with rounded lateral margins, an incompletely formed basis- phenoid and pterygoid bone, and a shortening and thickening of the tympanic ring, giving it a symmetric appearance (Figure7,
A and B).There were also homeotic transformations of the axial skeleton unique to the mutants. The most prominent defect involved a transformation of the axis (C2) toward the identity of the atlas (Cl). The neural arches of the axis were almost always broadened, re- sulting in a morphology more closely resembling that of the atlas. In extreme cases the neural arches of the axis were split, widened, and fused with the neural arches of the atlas (Figure 8B). An anterior arch of the atlas (aaa) characteristically present on C1 was also observed on C2, although it was small and fused to the aaa on C1 (Figure 8C). Other features of the axis, such as the dens, appeared unaffected. The ?6pub mu-
Analysis of piebald Deletion Mice 455
TABLE 3
Incidence of skeletal abnormalities associated with s?6fib homozygotes
Genotype
+/+"
27/27 36/+ 36/36 15/36 36/lAcrg 36/24 15/+ 15/15 15/lAcrg 15/24Total Cleft palate
Basisphenoid incomplete Basioccipital rounded Axis (C2)
Neural arches broadened and split
Atlas ( C l )
Neural arches fused with
Anterior arch of the ( m a ) Fused ribs
1 and 2 Unilateral Bilateral 6 and 7
Unilateral Bilateral
Cartilage malformations Hyoid
Larynx Trachea Neural arches Limbs
1 0
0 0 0
2 (4) 1 (4) 11 (31)
0 0 3 ( 8 )
0 0 10 (28)
0 0 2 (6)
0 0 0
37 17/23 (74) 16/25 (64) 23/26 (88)
19 (51)
10 (27) 9 (24)
6 (16)
4 2
3 4 7 (19)
36 (97)
36 (97) 35 (95) 36 (97) 36 (97)
21 0 2 (9.5) 0
3 (14)
0 0
0
0
9 (43) 3 (14) 5 (24) 1 (5) 0
13
0
2 (16) 11 (85)
3 (24)
4 (30)
6 (46)
1 ( 8 )
1 0 7 (54) 5 2
13 (100)
13 (100) 13 (100) 13 (100) 13 (100)
0 0 0
0 1 (2) 0
0 0 0
0 0 1 (6)
0 0 0
+,
s or Cast/Ei; 27, s'"~; 15, s15DttMb; 36, s36pu6; lAcr , sIAng; 24, .?4pu6. Values in parentheses are percentages."+/+,
Littermates from the s ~ ~ X ~ ?6pub/+, ~ ~ ~JDtfMb/+
~ / +and ?6w/+ intercrosses used to generate the mutant offspringfor the skeletal analysis and genotyped as Cast/Ei/Cast/Ei, Cast/Ei/s and s/s using the D14Mit8 molecular marker. These control littermates were pooled as no significant differences were observed in the analysis of individual populations.
the first and second ribs (65%) that in more severe instances (16%) resulted in their fusion. This possibly reflects an anterior transformation of T2 toward the identity of T1. There was also a fusion between the sixth and seventh ribs just before their attachment to the sternum (Figure 5C).
Cartilage malformations in mutants: The most
penetrant phenotype displayed by the ?6pub mutants was
defects in the cartilaginous skeleton. These defects were inherited as a semi-dominant trait, as the malformations were also observed in heterozygotes, although they were less severe and reduced in penetrance (Table 3 ) . Abnormalities were seen in the segmentally arranged C-shaped rings of the tracheal cartilage, which often displayed lateral fusions and were incompletely formed (Figure 9, A and B). The cartilages of the larynx were also incompletely formed and fused with adjacent struc- tures. The lesser horns of the hyoid bone were markedly thickened and fused to the greater horns (Figures 8B and 9B).
Cartilage defects were also present in the axial and appendicular skeleton. ?6pub mutants displayed lateral fusions between the neural arches of the sacral, lumbar and thoracic vertebrae (Figure 9, C and D). The neural arches consistently remained open along the dorsal midline, a condition commonly described as spina bi- fida occulta. Comparable defects in both the fore- and hindlimbs were restricted to the distal cartilaginous ele-
ments. In the forelimbs the carpal bones were poorly defined and the synovial joints appeared indistinct ow- ing to fusions between the thickened cartilages of the metacarpals and phalanges (Figure 9, E and F).
Ednrb does not account for the sIfrDttMb and
36hb
mu- tant phenotypes: Mice homozygous for the spontane- ous piebaldnull mutation, s', and the targeted disruption of the gene survive to weaning, well past the time whenloss of Ednrb cannot explain the neonatal lethality. How- ever, neither s' nor the targeted mutation mice have been specifically examined for CNS or skeletal defects, leaving open the possibility that these defects may result from the loss of Ednrb gene function. To test this, mice carrying the s27pu6-induced allele, which contains a point mutation in a highly conserved amino acid residue be- lieved critical for G protein-coupled receptor activity (M. K SHIN and S. M . TILGHMAN, unpublished data), were utilized. These mice display the null phenotype for piebald but have the advantage over s' mice that no additional deletion of DNA has been detected to date. When s~~~~~ homozygotes were examined, no defects in the architecture of the dorsal spinal cord were observed and there was no incidence of congenital hydrocepha- lus. Their skeletons did not exhibit malformations or changes in vertebral identity comparable to those seen and mutants (Tables 2 and 3). They did show a higher incidence of the (C7/T13/L5) vertebral
S15DtlMb and s~~~~~ homozygotes die, suggesting that the
456 T. P. O'Brien et al.
A
A
WT
' " 9 4
Iru
FIGURE 7.-Cranial skeletal defects in the ?6PzLb homozygotes. (A) Ventral view of the skull from a 18.5-dpc control littermate
and a ?6fi* mutant (B). The lateral margins of the basioccipital bone (bo) are rounded in the mutant, the basisphenoid bone
(bs) is incompletely formed showing a persistence of the hypophyseal foramen, the pterygoid process (p) is poorly formed (white asterisk in B), the palatine (pl) and maxillary (m) shelves are absent or poorly formed (white asterisks in B) and the dorsal distal tip of the tympanic ring (t) is shortened and thickened. (C) Frontal section through the head of a 14.5dpc control littermate. (D) In the mutants the palatal shelves (ps) have failed to rotate or elevate and remain lateral to the tongue
(tg) that appears to be abnormally positioned beneath the nasal septum.
pattern, which may reflect the influence of the genetic background. Consistent with this explanation, 15/29 (52%) of the s'7pu6/s heterozygous littermates also dis- played this variation. In any case, this pattern represents a posteriorization of vertebral identity and is therefore in the opposite direction of the changes observed in
Complementation analysis of s " ~ : The analysis of
s'7p"b mice argued that the CNS and skeletal defects of
cannot be attributed to the absence of the Ednrb gene product. To localize the genes responsible for these defects, s ~ ~ " ~ " " / s ~ ~ ' ~ compound heterozygotes were generated. The sIAT deletion allele is especially useful for complementation studies because its proxi- mal breakpoint is outside the s151'11M6 and s3'IPu6 proximal breakpoints, while its distal breakpoint lies within both deletions (see Figure 1B). Thus, it should complement
s15DllMb
mutants.
s15DtlMb
the defects associated with genes in the distal portion of these deletions. slAq homozygotes die between 10.5 and 11.5 dpc, suggesting that a gene(s) required for embryonic development lies in the most proximal por- tion of the slAT deletion (T. P. O'BRIEN and S. M. TILGHMAN, unpublished data).
at the expected 25% frequency, died as neonates, weighed less than their littermates and displayed the characteristic s15D1iMb and s'6p"6 mutant external mor-
phology. The s15D1iMb/s1Aq mutants had hydrocephalus
as shown by a moderate expansion of the lateral and third ventricles, and displayed alterations in the dorsal spinal cord at 14.5 dpc. Twenty-five percent of s15D"M6/
slAT skeletons displayed sternal malformations as seen
in the s15D6iMb skeletons. Furthermore 31% of the ani- mals exhibited eighth ribs that were lengthened to a p
Analysis of piebald Deletion Mice 457
c
*
c
n
r
u
L?-
i
F
-a
1
a
4
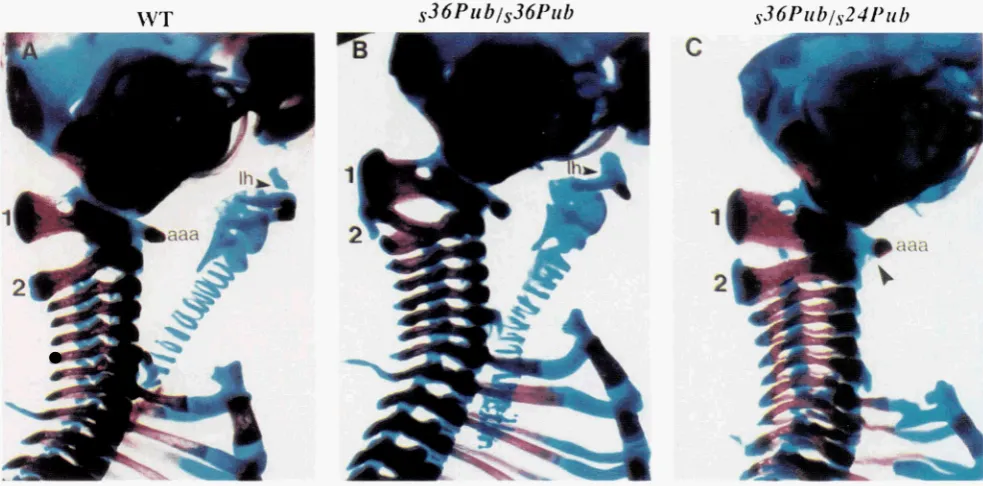
1FIGURE S.-Skeletal defects in the cervical region of s'6pub homozygotes. (A) Lateral view of control littermate at 18.5 dpc showing normal morphology of the first two cervical vertebrae, the atlas (1) and axis (2). The atlas ( C l ) displays broad neural arches and a characteristic anterior arch of the atlas (aaa). (B) In the s36pu" mutant the neural arches of the axis are broadened, split and fused with neural arches of the atlas. Also note the lesser horn (lh) of the hyoid is thickened and fused with the greater horn. (C) The $6pub/?4pub mutant displays a more complete transformation of the axis, shown by the broadening of the neural arches and the formation of a small aaa that is fused (arrowhead in C) with the aaa typically present on the atlas.
Complementation analysis of The axial and cranial skeletal defects and cartilage malformations unique to ?6hb were also subjected to genetic analysis.
As
expected, the s'~"''''~ allele was able to fully comple- ment all of the ?6"ub-specific axial and cranial skeletal abnormalities (Table 3), placing the gene(s) responsi- ble outside the deleted region held in common be- and ?6Pzrb. The cartilage malformationswere present in the s ~ ~ " ~ ~ ~ ~ / s ~ ~ ~ ~ " ' ~ animals in the same structures and with the same frequency as in the P h 6
heterozygotes (Table 3).
To determine whether the gene(s) responsible mapped to the proximal or distal region of the
Zhm
deletion, ? 6 p ' r b / s 1 A ~ animals were analyzed. s l A q res-cued the $6n'b-specific anterior axial and cranial skeletal defects. The cartilage malformations were comparable to those seen in the ?6"ub heterozygotes (Table
3).
Thisresult suggests that the gene(s) responsible maps to the deleted region of
?"'*
distal to the s15D16M6 breakpoint.To ensure that the absence of the g6'*-specific de- fects in the ?6pub/s'Affg mutants reflected true comple-
mentation and not genetic background differences, ?6hb/s24hb compound heterozygotes were analyzed. In- deed, the cartilage malformations were observed in 100% of the animals (Table 3). In addition, ?6hb/s24h6 mutant skeletons clearly displayed malformations com- parable to those of the ?6pub homozygotes, although
some differences were evident. The penetrance in- creased for the transformation of ventral aspects of the axis, based on a comparison between the
s36h6
andtween
S15DtlMbproach or contact the sternum, although a strict ante- rior transformation of T8
was
not evident as none were fused with the sternum. This maps the gene(s) responsi- ble for neonatal lethality, hydrocephalus, spinal cord malformation and skeletal defects to the proximal re- gion of thes'5D1"6
deletion (Figure 1C)./s animals did not display the full ex- tent of the axial skeletal transformations observed in the s~~~~~~ homozygotes (Table
2).
For example, the frequency of an L6 to S1 transition was intermediate when compared with s15DnMb mutant and control popula- tions, and an additional pair of ribs on L1 was not observed at a frequency greater than that seen in the heterozygotes. We attribute these differences to variability in the genetic backgrounds of d S D t t M b and s I A qmice, based on the analysis of two larger deletions,
d
6
'
*
and sZ4"*, that completely encompass s'5DnM6 (Figure 1B). For example, s15D11M6
/?'-
animals did not displaya higher incidence of eight vertebrosternal ribs, but an extra rib on
L1
was evident in43% of the animals,
identical in frequency to s15D11M6 mutants. On the other animals strict anterior transforma- tions of T8 and L l were not observed (Table2).
There- fore, two deletions,s'6P"'
and each spanning thedeletion, do not confer the same set of skeletal defects when present over the s15DtUM6 chromosome (Ta- ble
2). Based on these observations, it appears likely
that the expressivity of the s15DtUM6 skeletal defects arehighly sensitive to the differences in the genetic back- grounds of the deletion allele stocks.
The s15D11M6 I A q
S15DttM6
hand, in s15DllMb/s241.'ub
458 T. P. O'Brien et al.
?6p'L6/~24Pub mutants for a duplicated aaa on C2 fused to
the aaa of C1 (Figure 8C). There were fusions between the first and second ribs and sixth and seventh ribs, although the frequencies differed from those seen in the ?6p"6 mutants. A notable difference occurred with
the most anterior defects, where there was a reduced incidence of the incomplete formation of the basisphe- noid bone and an absence of clefting of the secondary palate (Table 3 ). This may reflect the background dif- ference of the s ~deletion allele stock, although a ~ ~ ~ " strict complementation of the palatal defect cannot be excluded.
DISCUSSION
The identification of genes important in mammalian development has relied on a relatively small number of experimental approaches. The isolation of murine homologues for genes discovered in other species, espe- cially Drosophila melanogaster, has proven to be a particu- larly successful strategy. Recently, genes whose expres- sion patterns are revealed by promoter/enhancer trap methodologies represent an alternative approach
(FRIEDRICH and SORIANO 1991). The analysis of mice carrying spontaneous mutations or large chromosomal deletions represents a third approach, one that has the potential to uncover novel genes that may be missed by the other methods (NISWANDER et al. 1989). This study establishes that genes contained in the piebald deletion complex affect embryogenesis, patterning of the skele- ton, and CNS development. Based on complementation analysis, the genes have been separated genetically and localized to different chromosomal regions.
The
d5-
deletion: The s15D1"' allele was the small- est deletion examined. In addition to Ednrb, slfDtiMb was deleted for the flanking markers D14Mit93 and D14Mit94. D14Mit93 maps 0.5 cM proximal to Ednrb, whereas D14Mit94 could not be separated from Ednrb (Figure 1A) and is contained on a 670-kb Ednrb YAC (H. CHEN, D. L. METALLINOS and S. M. TILGHMAN, un- published results).mutants were smaller than their lit- termates and exhibited a pronounced curvature of the body axis. While these phenotypes were fully penetrant, there was some variability between littermates in the extent of their expression. The s15DL1M6 homozygotes in- variably died immediately following delivery from a p parent respiratory failure.
The s~~"'"' mutants exhibited a malformation of the developing spinal cord that was restricted to the dorsal region along the entire rostrocaudal axis. Both the dif- ferentiation and the distribution of cell types arising in the dorsal horn appeared to be altered. The birth and differentiation of specific neuronal classes approxi- mately follows a ventro-to-dorsal temporal gradient. The first to originate are the large motor neurons of the ventral horn, followed by the more widely distrib-
The S15D11Mh
FIGURE 9.-Cartilage malformations in d6"'" mutants. (A) Ventral view of whole mount of the trachea and hyoid bone from a 1 8 . 5 1 1 ~ ~ control littermate and a s~~'"'' mutant (B)
showing the fusions between the cartilages of the lesser horn (lh) and greater horn (gh) of the hyoid bone (top arrow in
B), the greater horn (gh) of the hyoid and the rostral horn of the thyroid cartilage (tc) (second arrow in B), and the cricoid cartilage (rc) and trachea (tr) (third arrow in B). The cartilaginous rings of the trachea (tr) are abnormal displaying lateral fusions and openings along their ventral surface (bot- tom arrow in B). (C) Lateral view of the posterior axial skele- ton from a control littermate and a
d6P7'b
mutant (D) demon- strating fusions between the neural arches (na) of the lumbar and sacral vertebrae (arrowhead in D). (E) Dorsal view of the forelimb from a newborn control littermate and a newborn?6W16 mutant (F). The bones in the carpal region (cr) includ-
Analysis of piebald Deletion Mice 459
uted relay neurons of the intermediate gray. Among the last to form are the interneurons of the dorsal horn and the dorsomedial gray (ALTMAN and BAYER 1984). The defects in the s’5DtfMb mutant spinal cord, arising between 12.5 and 14.5 dpc, were spatially and tempo- rally coincident with the formation of this latest class of cell types.
Some features of the sljDttMb mutant spinal cord were consistent with a developmental delay, such as the for- mation of the dorsal funiculus and the time of appear- ance of differentiated cells in the dorsal horn. However, the morphology of the s ’ spinal cord does not sim- ~ ~ ~ ~ ~ ~
ply reflect an earlier gestational stage. Further studies are needed to address how the altered morphology re- lates to developmental timing and the specification, growth and differentiation of both neuronal and glial cell types in the spinal cord.
Another external feature of the s’5DtfMb fetuses was a swelled dome-shaped head, attributable to a moderate expansion of the lateral and third ventricles. A struc- tural malformation affected the subarachnoid spacejust below the roof of the fourth ventricle. It is possible that this defect, combined with a narrowing of the ventricu- lar passages, interferes with the transit of cerebro-spinal fluid leading to the relatively mild hydrocephalus seen
The s ~homozygotes displayed ~ ~ ’ ~ a number ~ ~ of ~skele- tal defects, the most consistent ofwhich were malforma- tions in the sternum. The sternum is formed by the fusion of paired ventromedially located sternal rudi- ments. The ribs contact the sternal rudiments just be- fore closure, forming the sternocostal junctions. The intercostal space undergoes ossification to become the segmentally arranged sternabrae while the points of ar- ticulation between the sternum and ribs persist as carti- lage. In this way the attachment of the ribs influences the proper formation of sternal pattern (CHEN 1952, 1953). The sternums from the s‘5DtfMb mutants exhibited disorganized and compromised formation of the ster- nabrae, directly correlating with the misalignment or crowded attachment of the ribs.
homozygotes also displayed anterior transfor- mations of vertebrae in the midthoracic and lumbar region of the skeleton (Table 2). Aspects of this pheno- type were remarkably similar to those characteristic of loss-of-function mutations in members of the Hox gene family. This included their restriction to a specific axial domain and their variable penetrance and expressivity. The defects were comparable to mutations in Hoxa-5, Hoxb-4 and Hoxd-3, which affect sternal development (CONDIE and CAPECCHI 1993; JEANNOTTE et al. 1993; RAMIREZ-SOLIS et al. 1993), those in Hoxc-8, Hoxa-5 and Hoxa-10 affecting thoracic and lumbar vertebae (LE MOUELLIC et al. 1992; JEANNOTTE et al. 1993; SATOKATA et al. 1995) and the transformations of lumbar and sa- cral vertebrae in Hoxa-IO, Hoxa-l l and Hoxd-l l muta- tions (SILIALL and POTTER 1993; DAVIS and CAPECCHI
in the s/51>11Mb
mutants.
s15DftMb
1994; SATOKATA et al. 1995). Thus, the skeletal alter- ations in the s’5Dt1Mb mutants resembled mutations in the Hox genes more centrally located within the clusters (group 3 through group 11)
,
rather than those with more anterior or posterior expression boundaries.The deletion: Mice homozygous for the ?6Pub
deletion exhibited characteristics of s15DftMb homozy- gotes, including respiratory failure at birth, small size, axis curvature, hydrocephalus and malformation of the spinal cord.
The cartilage malformations identified in ?6Pub homo-
zygotes were highly penetrant and were evident at re- duced frequency in
s’@*
heterozygotes (Table 3). This raised the possibility that the phenotype was due to a gain-of-function rather than a loss-of-function muta- tion, for example, due to the expression of a novel gene created at the deletion breakpoint of ?6pub. Thisexplanation is difficult to reconcile with the observation
/s mutant mice displayed cartilage mal- formations analogous to those seen in the
s?“~”
hetero- zygotes. Molecular mapping indicated that the de- letion encompasses most, if not all, of the region missing in the chromosome. It is unlikely that this allele would harbor the same breakpoint-specific mutation capable of producing a dominant cartilage phenotype. Furthermore, ?6fib/s24pub mutant mice dis- played fully penetrant cartilage defects, indistinguish- able from ?6pub homozygotes. Taken together, these re- sults suggest that the cartilage defects observed in the?6pub heterozygotes represent haplc-insufficiency of a
gene(s) deleted in both the
g6‘*
and s~~~~ alleles. The ?6fib homozygotes exhibited the sternal malfor-mations seen in the s ” homozygotes. In addition, ~ ~ ~ ~ ~
they displayed skeletal defects distinct from those associ- ated with the s’5DttMb mutants. These ?6pub-specific de- fects were comparable to those for the targeted disrup- tion of the Hox genes with more anterior expression boundaries. For example, C2 to C1 transformations were observed with Hoxd-3 and Hoxb4 mutations (CON-
DIE and CAPECCHI 1993; RAM~REZ-SOLIS et al. 1993) and in Hoxa-4, Hoxb4 and Hoxd-4 compound mutants (HORAN et al. 1995). Cleft palate was seen in a Hoxa-2 disruption (GENDRON-MAGUIRE et al. 1993; RIJLI et al.
1993), and the altered morphology of the basioccipital bone was displayed in mutations for the Hoxa-1 and Hoxa-2 genes (LUFKIN et al. 1991; GENDRON-MAGUIRE et
al. 1993; RIJLI et al. 1993). There were also similarities to “induced skeletal malformations (KESSEL and GRUSS 1991; KESSEL 1992) and those seen with loss-of-function mutations in the RARs (LOHNES et al. 1993, 1994; MEN-
DELSOHN et al. 1994). The comparison between the mutant and the RAB null phenotypes is particularly relevant because of the shared defects in cartilaginous structures and the axial and cranial skeleton.
460 T. P. O'Brien et al.
the embryo (reviewed in KRUMLAUF 1994). Changes in vertebral identity have also been observed in retinoic acid (RA)-induced skeletal alterations (KESSEL and GRUSS 1991; KESSEL 1992) and disruptions in the gene for the RARy (LOHNES et al. 1993). Hox gene expression is influenced by RA through the action of its cellular receptors (LANGSTON and GUDAS 1992; POPPERL and FEATHERSTONE 1993; MARSHALL et ul. 1994) Several stud- ies have established that the RARs are involved in axial patterning and are probably acting within the Hox path- way (reviewed in CONLON 1995). The phenotype o h served in the s15Dt1Mb and ?6Pub mutants suggest that the
genes affected might also act within the Hox pathway. The most prominent of the cranial skeletal defects was cleft palate. A number of different genes have been shown to affect palatal development, such as Msxl (SA-
TOKATA and MAAS 1994), Gubru3 (CULIAT et UL. 1993; NAKATSU et al. 1993) and Hoxu-2 (GENDRON-MAGUIRE et al. 1993; RIJLI et al. 1993), reflecting the complex coordination of morphogenetic movements required for the proper formation of the secondary palate (FER-
GUSON 1988). Any perturbation in the coordinated out- growth, rotation and elevation, or cellular adhesion leading to fusion could disrupt palatal development. The palate defect in the ?6pub mutants appeared to
af-
fect palatal shelf rotation and elevation.
The functional map of the piebald deletion com- plex: As a first step in the functional analysis of the
piebald deletion complex, we set out to exclude the pos- sibility that the phenotypes observed in s~~~~~~~ and mutants could be due to the loss-of-function of piebald.
To do this, s~~~~~ homozygotes, which carry a point muta- tion in the gene, were analyzed for srsDttMb- and ?6pub-
specific alterations. None were observed, arguing that these phenotypes are due to the loss of genes in addi- tion to piebald.
To construct a functional map of the region (Figure lC), mice carrying different combinations of deletion alleles were analyzed. The neonatal lethality, CNS de- fects, vertebral transformations and the malformation of the sternum fell within the region bounded by the deletion. As the sIAcrg deletion fails to the rescue the neonatal lethality, hydrocephalus, and spinal cord malformation, they can be localized to the region ex- tending from the proximal breakpoint of s~~~~~~~~
through the distal breakpoint of s ' ~ ~ .
skeletal defects observed in the s15DttMb homozygotes. Although this could reflect a partial complementation of the skeletal phenotype, we do not favor this interpre- tation. This is based on the observation that the range of skeletal defects were either reduced or eliminated in mice carrying the s151't'Mb deletion in combination with the ?6Pub or alleles (Table 2). As these deletions
completely encompass the s15D11M6 deletion, it seems likely that the phenotypic variability in these allelic com- binations is the result of their heterogeneous genetic s15111tM6
The S15D1tMb/SIAng mice displayed only a subset of the
backgrounds, rather than true complementation of the specific defects. The s~~~~~~~ and ?6pub mice have
DNA derived from the original mutagenized stock, an (101 X C3H)FI male, but they have since been out- crossed to SSL/Le, Cast/Ei and the Oak Ridge T stock. s~~~~~ and sZAqmice have only been maintained opposite the Oak Ridge T stock and are thus more genetically homogeneous. Although the homeotic transformations of specific vertebrae appeared to depend on the back- grounds of the deletion allele stocks, the sternal defects were present in all the mutants. Given the likelihood that the sternal defects and skeletal transformations have the same underlying cause, we propose that the skeletal defects map to the proximal portion of the deletion along with the other s15u'"b-associated abnor- malities.
The modifylng effects of genetic background have been invoked to explain some of the variability in the skeletal defects reported for loss-of-function mutations in a number of Hox genes (LE MOUELLIC et al. 1992; CONDIEAND CAPECCHI 1993;JEANNO'rrE et al. 1993), the
RARs (LOHNES et ul. 1993; MENDELSOHN et al. 1994), and the bmi-1 gene (VAN DER LUGT et al. 1994). This point has been clearly demonstrated in the skeletal phe- notypes reported for mice carrying a mutation in the
Hoxb-4 gene (RAM~REZ-SOLIS et ul. 1993). Furthermore, the ability of teratogens to influence the patterning of the skeleton is dependent upon genetic background (RUSSELL 1979).
and slAq are able to rescue the
g6'"'*-
specific defects. This localizes the cartilage malforma- tions and anterior axial and cranial skeletal defects to the distal end of the deletion between the s15"t1M" and breakpoints. The s ~ deletion was unable to ~ ~ ~ "rescue the ?jPuh skeletal phenotype. One striking excep-
tion was the complete absence of a clefting of the sec- ondary palate in the ?6Pu6/~24Puub compound heterozy-
gotes. Although genetic background differences may once again be the explanation, in this instance it is also possible that the cleft palate is separable from the other abnormalities and that the s~~~~~ allele is able to rescue this defect. The relative position of the distal breakpoints of these two alleles has not been estab- lished, therefore, it is possible that the deletion in ?6Pub
extends further distal than s'"'~~.
Finally, the prenatal lethality of the SIAcrg homozygotes was rescued by both s15Dt1Mb' and s ~
. This maps the
~ ~ ' ~ ~region of lethality to the proximal-most portion of sIAq, between the slAmg and s~~~~~ proximal breakpoints. This region has been cloned on two YACs, both of which span the breakpoints, and is estimated to be <500 kb (L. J. KURIHARA, D. L. METALLINOS, T. P. O'BRIEN and S. M. TILGHMAN, unpublished observations).
In conclusion, this work establishes new mouse mu- tants that will be useful as models for studying hydro- cephalus, organization of the developing spinal cord and skeletal and cartilage morphogenesis. The goal of this
I5DltMb-