The Effects of Musical Training on Movement
Pre-Programming and Re-programming Abilities:
An Event-Related Potential Investigation
Melis Anatürk & Ines Jentzsch
School of Psychology & Neuroscience, University of St Andrews, UK
Author Note
Melis Anatürk, School of Psychology & Neuroscience, University of St Andrews; Ines Jentzsch, School of Psychology& Neuroscience, University of St Andrews.
Abstract
Two response precuing experiments were conducted to investigate effects of musical skill level on the ability to pre- and re-program simple movements. Participants successfully used advance information to prepare forthcoming responses and showed response slowing when precue information was invalid rather than valid. This slowing was, however, only observed for partially invalid but not fully invalid precues. Musicians were generally faster than non-musicians, but no group differences in the efficiency of movement pprogramming or re-programming were observed. Interestingly, only musicians exhibited a significant foreperiod Lateralized Readiness Potential (LRP) when response hand was pre-specified or full advance information was provided. These LRP findings suggest increased effector-specific motor preparation in musicians than non-musicians. However, here the levels of effector-specific preparation did not predict
preparatory advantages observed in behaviour. In sum, combining the response precuing and ERP paradigms serves a valuable tool to examine influences of musical training on movement pre- or re-programing processes.
The Effects of Musical Training on Movement Pre-programming and Re-programming Abilities: An Event-Related Potential Investigation
Instrumental music performance frequently involves the rapid execution of well-practiced and sequential uni- or bi-manual movements (Jerde et al., 2006; Palmer, 2006). For example, a top-level pianist is able to produce up to 1,800 notes per minute (Münte et al., 2002). In order to achieve such advanced-level musical ability, individuals need to engage in a significant amount of deliberate practice that is usually gained over a period of many years (often initiated in early childhood; Sloboda & Davidson, 1996; Williamon & Valentine, 2000). During these rehearsals, instrumentalists repeatedly perform finger movements required to produce each musical note, for example by pressing successive piano keys (Watson, 2006). This extensive training leads to the development of fine motor skill (Willingham, 1999; Jabusch, 2006) as demonstrated by the superior performance of musicians, compared to non-musicians, on finger tapping (e.g. Franěk et al., 1991; Repp & Doggett, 2007) and motor sequence learning tasks (e.g. Landau & D’Esposito, 2006).
A widely distributed brain network is recruited during instrument playing (Baumann et al., 2007) and the high motor and auditory demands placed on the developing brain by intensive musical practice seem to lead to structural and functional neural adaptations (Pascual-Leone et al., 2005). For example, children who received music lessons over a 15-month period exhibited enlargements in the right precentral gyrus, right primary auditory cortex and corpus callosum compared to controls (Hyde et al., 2009). Neuroanatomical and
musician and non-musicians (for a review, see Wan & Schlaug, 2010), particularly within the motor network (Herholz & Zatorre, 2012). Indeed, neuroimaging studies indicate that professional musicians exhibit larger grey matter volume in the primary motor cortex (Amunts et al., 1997; Gaser & Schlaug, 2003; Elbert et al., 1995) and cerebellum (Hutchinson et al., 2003), relative to non-musicians. Furthermore, structural enlargements in the primary motor cortex (M1) appear to correspond to the type of instrument that an individual specializes in (Elbert et al., 1995; Bangert & Schlaug, 2006). For example, Bangert and Schlaug (2006) reported that professional string-players exhibited pronounced cortical representations of their left hand (i.e. the hand predominantly used to play their instrument) in the right M1. These enlarged M1 representations may enable musicians to execute their movements more
efficiently (Jäncke et al., 2000; Jentzsch et al., 2014).
Movement Pre-programming
Voluntary movements are prepared before they are performed (Keele, 1968; Churchland et al., 2006) and the response precuing paradigm
(Rosenbaum, 1980; 1983) is frequently utilized to investigate these processes. In such tasks, a precue appears before target onset and conveys advance partial information about a forthcoming response (i.e. ‘foreperiod’). Precues can
indicate, for example, the hand (left or right), type of finger (i.e. index or middle) or direction of movement. Subsequently, a target provides complete information about the required response. If participants utilize precue information to prepare their responses in advance, their reaction times (RT) are significantly faster in informative precue trials compared to non-informative trials (for a review, see Leuthold et al., 2004). This suggests that even incomplete information about forthcoming movements can activate the necessary motor programs, which enable faster responses (Ulrich et al., 1998; Shojaei & VaezMousavi, 2007; but see Goodman & Kelso, 1980). However, response precuing tasks are restricted to overt measures of movement preparation, hence, are frequently combined with
the ERP method (for a review, see Leuthold et al., 2004). ERP studies examine the neural correlates of action preparation: the foreperiod Lateralized Readiness Potential (LRP; Gratton et al., 1988) and foreperiod Contingent Negative
Variation (CNV; Walter et al., 1964), extracted from electrophysiological recordings of neural activity with high temporal accuracy (Coles et al., 1995; Friedman, 2000).
arising from the frontoparietal and centroparietal regions, respectively. The earlier component indexes non-motoric processing such as sensory preparation,
anticipatory attention and orienting towards the warning stimulus (e.g., Brunia & van Boxtel, 2001; Gomez et al., 2004; Loveless & Sanford, 1974; Rohrbaugh, & Gaillard, 1983), whereas the late component corresponds to muscle-unspecific motor preparation (Rohrbaugh et al., 1976; for a review, see Leuthold et al., 2004). This study focuses on the late foreperiod CNV,which has been suggested to reflect the combined activity of lateral premotor and supplementary/cingulate motor areas (Leuthold & Jentzsch, 2001). In response precuing tasks, the late foreperiod CNV is detected during all trials, although its amplitude increases with the amount of advance information provided by the precue (Mackay & Bonnet, 1990; Leuthold et al., 2004). The foreperiod LRP, an index of selective motor activation of the specific effector representation and originates most likely from the primary motor cortex (Deecke et al., 1976; Leuthold & Jentzsch, 2001; Jentzsch & Leuthold, 2002). It can therefore only be detected when the specific effector (e.g. left or right hand) is known in advance. This ERP component is recorded over the C3’ and C4’ electrode sites and calculated by subtracting the
As mentioned earlier, musicians have been reported to show structural enlargements in motoric areas of the brain. Musicians may therefore be more efficient at preparing their actions compared to non-musicians (Jäncke et al., 2000). For example, professional instrumentalists have been shown to anticipate their forthcoming movements when performing musical compositions (Engel et al., 1997) and more musically experienced individuals exhibit more anticipatory behaviour (Palmer & Pfordresher, 2003; Palmer & Dalla Bella 2004). Indeed, RT latencies generally seem to decrease as musical experience increases (e.g., Jentzsch et al., 2014). Moreover, young adults with musical training respond faster on uni-manual and bi-manual RT tasks compared to musically-naïve individuals (Hughes & Franz, 2007). Overall, musical practice may accelerate motor preparation processes, thus reducing their contribution to RT. Thus, the first experiment reported here aimed to directly examine the influence of musical practice on selective movement preparation.
Movement Re-programming
Prepared movements must occasionally be modified before they are executed, such as when an unforeseen change occurs in the environment. This process of response re-programming enables humans to flexibly engage with their surroundings (Larish & Frekany 1985; Stelmach et al., 1988). Response precuing tasks can also be used to investigate this form of movement preparation (Leuthold & Jentzsch, 2002b), by including invalid precues that encourage
additional, inhibitory as well as motoric re-programming processes involved in invalid trials (Larish & Frekany, 1985; Lépine et al., 1989). Again, to our
knowledge no previous study has addressed the influence of musical practice on the ability to re-program an incorrectly pre-specified motor programme. A recent study by Jentzsch et al. (2014) suggested that musicians even at an amateur level might have a better ability to detect conflicts and errors compared to non-musicians. Thus, from this evidence one could predict musicians to be more efficient at re-programming their movements. This would also fit with brain
imaging data suggesting an involvement of the ACC not only in error and conflict detection in general, but also specifically detecting conflict between existing and newly activated motor plans (Leuthold & Jentzsch, 2002b).Thus, Experiment 2 investigates whether amateur musicians show a better ability for motor re-programming compared to non-musical control participants.
Experiment 1
Based on the hypothesis that musical practice leads to more efficient responses (Jäncke et al., 2000; Jentzsch et al., 2014), musicians are expected to generally respond faster to targets compared to non-musicians. Moreover, if musicians do engage in more efficient motor preparation, they should respond faster in trials with finger and hand precues compared to non-informative and ambiguous conditions. As previous studies consistently report structural enlargement in the M1 and associated motor areas (e.g. Elbert et al., 1995), musicians are predicted to exhibit larger foreperiod late CNV (for all precue conditions) and foreperiod LRP amplitudes (only when hand is pre-specified, as no foreperiod LPR is predicted for the finger precue condition), relative to non-musicians.
Method Participants
Thirty-one participantscompleted a single session approximately 2-hours in length. Sixteen of these individuals were classified as musicians (>1000 hours of accumulated practice time) and fifteen as non-musicians (< 1000 hours of accumulated practice time). Data from one additional participant were excluded, as that participant was substantially older (56 years old) than all other
participants (range: 18 to 34 years old).
All participants provided written informed consent, had normal or
corrected-to-normal vision and were reimbursed for their participation with £10. The study was conducted after receiving ethical approval from the Psychology and Neuroscience School Ethics Committee.
PANAS Mood Questionnaire: The trait version of the Positive and
Negative Affect Schedule (PANAS; Watson et al., 1988) was used to control for
the effects of general positive and negative affect on movement preparation. In this self-report questionnaire, respondents rated each of the 20 words (describing different feelings and emotions) using a 5-point scale (1=very slightly or not at all;
5=extremely), on the extent to which they generally experience each emotion.
Scores are generated separately for negative and positive affect and can range from 10-50. The trait version of the PANAS has good internal consistency and test-retest reliability (Watson et al., 1998).
Music Experience Questionnaire: The Music Experience Questionnaire
(Jentzsch et al., 2014) was used to collect demographic (e.g. age, gender)
information and details of each respondent’s musical experience and knowledge. Respondents completed open-ended questions about the musical instruments that they currently play or have previously played. They were required to provide the number of years spent actively engaging with an instrument, the start age and where applicable, the end age, any formal examinations taken and
accumulated practice time of each instrument. Four 5-point scales (1=None or Not Able; 5 =Extensive or Very Able) also measure respondents’ ability to read
music, their knowledge of music history/ theory and overall musical ability. A general musical knowledge score is then calculated for each individual (score range: 0-20). The final two questions focus on the average number of hours per week spent listening to music and number of hours per month spent engaging in non-musical activities (Jentzsch et al., 2014).
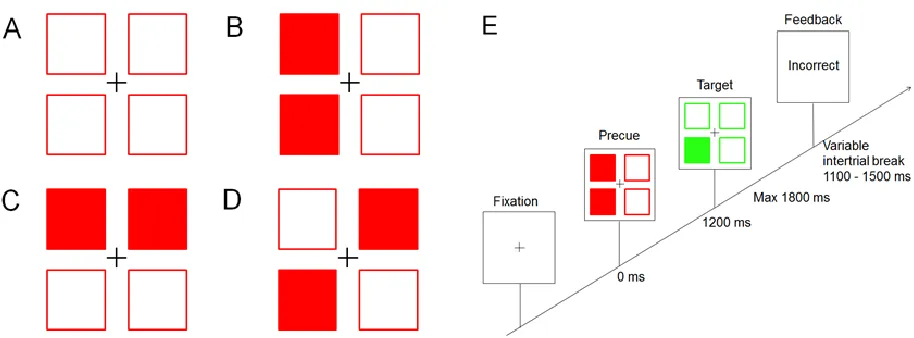
information was presented within four empty squares (0.4°x 0.4° visual angle) that were displayed on a white background and aligned with one another to form
a large square (1° x 1°). In this task, precues provided advance partial
information about response hand, fingers, ambiguous parameters or no
information. Precues provided valid information in 100% of trials. The possible response alternatives were conveyed by two filled red squares, see Figure 1.
During trials where response hand was identified, the left-top/ left-bottom or
right-top/right-bottom squares were filled in red, which indicated the left or right hand,
respectively. When the precue displayed advance information about fingers, the
top-left/top-right squares (i.e. left and right middle fingers) or the bottom two
squares became filled in red (i.e. left and right index fingers). Ambiguous precues
(i.e. neither finger nor hand parameters pre-specified; Jentzsch et al., 2004)
appeared as either top-left/bottom-right or top-right/bottom-left red squares.
When no advance information was provided, all squares remained unfilled. A
formation of four green squares (3 unfilled; 1 filled) that were of identical size and
location to the red squares served as target and appeared immediately after
precue offset indicating the correct response finger.
displayed for 1000 ms. After the offset of trial feedback or when correct
[image:12.595.94.551.196.365.2]responses were made, a blank screen was displayed until the start of the next trial, which randomly varied between 1100-1500 ms. Figure 1 (panel E) provides an illustration of a trial timeline.
Figure 1. Left: Examples of each precue condition of Experiment 1. A. No information.
B. Response hand information (indicates left hand in example). C. Response finger
information (indicates left and right middle fingers in example). D. Ambiguous information
(indicates left index finger and right middle finger in example). Right: E. Schematic
diagram of the order of events within a single trial, over time.
Participants sat in a dark room, approximately 80 cm from a 17-inch CRT
computer monitor on which the response precuing task was presented. They
were first instructed to position their middle and index fingers on the four response keys mounted on an ERTS keypad. The left and right middle fingers remained on the two outer response keys and the index fingers on the inner response keys throughout the experiment. Participants were informed that they would be presented with a configuration of four squares, each related to a
Participants performed 12 practice trials to become familiar with the
stimulus-response mapping. The experimenter then explained the meaning of the precues and emphasized the importance of utilizing this information to prepare responses, because it would facilitate both quick and accurate task performance. Participants performed 6 experimental blocks (2 practice and 48 experimental trials each). Presentation of the different trial types was randomized within blocks. After each experimental block, participants received feedback on their performance, including their mean RT as well as number and percentage of response errors. Subsequently, participants performed a second response precuing task (described in Experiment 2) before completing the trait version of
the PANAS (Watson et al., 1988) and Musical Experience Questionnaire
(Jentzsch et al., 2014).
Electrophysiological Recordings and Analysis
A BIOSEMI Active-Two amplified system with 72 Ag/AgCl electrodes was used to record EEG signals. Four electrodes positioned were near each eye measured horizontal and vertical eye movements. The Common Mode Sense
(CMS) and Driven Right Leg (DRL) electrodes were utilized as reference and
ground electrodes, respectively. Moreover, EEG and electro-ocular activity was
excluded from the analysis. Epochs were then averaged for each of the 8
experimental conditions (4 precues x 2 hands), a low-pass filter of 20 Hz applied, and average-referenced.
Trials with EEG artifacts, response latencies shorter than 150ms or longer than 1000ms post target-onset were excluded from RT and EEG analyses. The foreperiod LRP was calculated for left and right hand responses separately, where M1 activity from the recording site (i.e. C3/C4) ipsilateral to each response hand was subtracted from the activity in the contralateral site. The resulting value was averaged across all left and right hand responses to remove ERP activity not
associated with the motoric preparation of hand movements. Furthermore, the late foreperiod CNV was analysed on the Cz electrode site, where the late CNV showed showed maximal activity in the present data. A 200 ms analysis window was chosen for the mean foreperiod CNV and foreperiod LRP amplitudes, which started 200 ms before target-onset. In addition, we also analysed the response target LRP amplitude using automatic peak detection software and applying a search window 200 to 600 ms after target onset. The mean foreperiod and target-related ERP components were calculated separately for musicians and non-musicians, for each precue category and a baseline of 200 ms prior to precue onset was used throughout.
Results Demographic Information
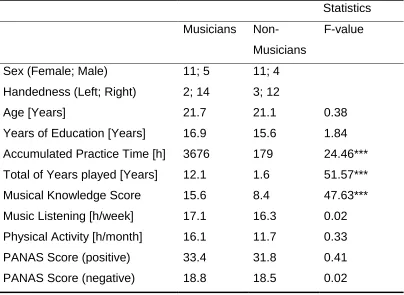
Table 1 provides the mean, standard deviation and F-statistic of the Analysis of Variances (ANOVAs) with factor group conducted for each
and PANAS scores for general negative affect and positive affect between musicians and non-musicians. Musicians had significantly more musical experience (i.e. accumulated practice time, larger total of years played and higher general musical knowledge) compared to non-musicians.
Table 1: Demographic participant information
Statistics Musicians
Non-Musicians
F-value
Sex (Female; Male) 11; 5 11; 4
Handedness (Left; Right) 2; 14 3; 12
Age [Years] 21.7 21.1 0.38
Years of Education [Years] 16.9 15.6 1.84 Accumulated Practice Time [h] 3676 179 24.46*** Total of Years played [Years] 12.1 1.6 51.57***
Musical Knowledge Score 15.6 8.4 47.63***
Music Listening [h/week] 17.1 16.3 0.02 Physical Activity [h/month] 16.1 11.7 0.33
PANAS Score (positive) 33.4 31.8 0.41
PANAS Score (negative) 18.8 18.5 0.02
***: p < 0.001.
Main musical instrument played Musical Practice level
Musicians 6 Piano, 4 Violin, 2 Flute, 3 Guitar, 1 Clarinet Non-Musicians n.a.
Behavioural Data
hand, ambiguous precues) and between-subjects factor group (musicians vs. non-musicians) were performed on RTs for correct responses and on arcsine-transformed error rates.
Reaction Times: Musicians generally responded faster (M = 349ms) than non-musicians (M = 383ms), F(1, 29) = 6.13, p = .019, ηp2 = 0.18. There was a significant effect of precue category, F(3, 87) = 78.87, p < .001, ηp2 = 0.73. Bonferroni corrected post-hoc analyses revealed that participants were slowest for the non-informative precue condition (391 ms) compared to all three partial precue conditions, all Fs > 61.07, all ps < .001. Participants responded
significantly slower when the precue provided hand information (366 ms) compared to both the finger precue ( 352 ms) and the ambiguous precue
condition (354 ms), all Fs > 25.18, all ps < .001, with the two latter conditions not significantly differing from each other, F < 1.0. There was no significant
Precue Information
No Finger Hand Ambiguous
Error Rate
[%]
0 1 2 3
Non-Musicians Musicians
Mea
n Re
acti
on
T
ime
[
ms]
[image:17.595.86.415.81.476.2]0 250 300 350 400
Figure 2: Mean RT (top) and percentage of error rates (bottom) for musicians and
non-musicians, for each of the four precue conditions in Experiment 1.
Electrophysiological Data
Foreperiod CNV Amplitude:A 2 (group) x 4 (precue category) mixed
the three partial precue conditions not significantly differing from each other (all ps > .05). There was no significant effect of musical practice on the foreperiod CNV amplitude (musicians: M = -6.5 μV; non-musicians: M = -4.9 μV), F(1, 29)= 1.18, p = .29, nor was there any significant interaction between precue category and group, F(3, 87)= 1.67, p = .18.
0 500 1000 1500
C N V A mpl itud
e at C
z [ V] -8 -6 -4 -2 0 2 Precue Target Musicians ms 0 500 1000 1500
[image:18.595.88.515.224.389.2]No Precue Finger Precue Hand Precue Ambiguous Precue Precue Target Non-Musicians
Figure 3. The mean foreperiod CNV amplitudes on Cz during trials with no or partial
advance information (finger, hand, or ambiguous) in Experiment 1, for musicians and
non-musicians. The topographic spline map on the right depicts the spatial distribution of
the average activity in a time 200 window immediately preceding target onset.
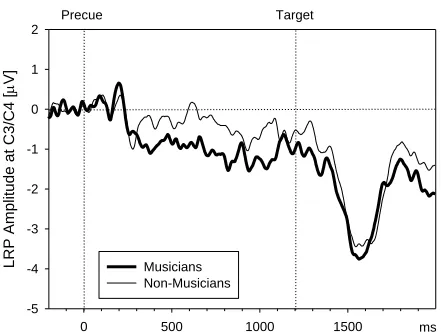
Foreperiod LRP Amplitude: Planned comparisons using one-tailed t-tests were performed on LPR amplitudes, testing whether the LRP deviates significantly from zero in the negative direction for musicians and non-musicians. We only expect the LRP to differ from zero in the hand precue condition, which is depicted in Figure 4. Musicians displayed a significant foreperiod LRP amplitude in the hand precue condition (-1.1 μV), t(15) = -1.93, p= .037. The foreperiod LRP amplitude in the hand precue condition did not differ significantly from zero for the
non-musicians (-0.5 μV), t(14) = -1.21, p= .13. The LRP amplitude did not significantly differ from zero in any other precue conditions (all ps > 0.10).
ANOVA revealed no significant effects, all Fs < 1. Importantly, there was no significant effect of musical practice on the target-related LRP amplitude (musicians: M = -4.3 μV; non-musicians: M = -4.3 μV), F(1, 29) < 1. Thus, the absence of a foreperiod LRP in the non-musician group cannot be attributed to generally reduced lateralized motor activity for this group.
ms
0 500 1000 1500
LRP
Ampli
tude at C3/C4 [
V]
-5 -4 -3 -2 -1 0 1 2
Musicians Non-Musicians
[image:19.595.88.309.244.410.2]Precue Target
Figure 4. The mean LRP amplitudes during trials with partial advance information about
response hand in Experiment 1. Data for musicians and non-musicians are
superimposed.
Discussion
The results of the first experiment suggest that musicians, although
significant, so the lack of an observed group difference cannot be attributed to participants’ general lack to engage with the precue information. Some evidence for differences in brain activity between groups was observed. First, the CNV amplitude seemed generally slightly enlarged for musicians compared to non-musicians, although this effect was not significant. Also, only non-musicians, but not non-musicians, exhibited a significant foreperiod LRP when response hand was pre-specified. Importantly, lateralized activity during the response phase (target-related LRP) did not differ between groups. Together, these LRP findings are surprising, especially given that no behavioural differences in the size of the precue effect were found between groups. Given that the target-related LRP was of similar size for both groups, differences in the foreperiod LRP amplitude cannot have resulted from structural differences between groups. In order to further explore the link between behavioural results and the foreperiod LRP, we run a correlation between the size of the foreperiod LRP precue effect (no information precue minus hand information precue) and the hand precue effect in RTs (no information precue minus hand information precue) across all participants, discarding factor group. The correlation was not significant, r = 0.02. We will come back to discussing this finding in more detail in the general discussion.
In sum, despite the evidence that our group of musicians responded faster in general and showed differences in lateralized preparatory activity in motoric areas of the brain, they were not selectively advantaged in the ability to use advance information to prepare for an upcoming motor response.
Experiment 2 examined the effects of musical skill level on movement re-programming skills. This experiment used a response precuing task with precues that provided either no advance information, or full information that could be valid or alternatively, providing invalid information about hand, finger, both parameters. Trials with full valid precue information can be compared to non-informative precues to potentially replicate and further extend the findings reported in experiment 1. More specifically, participants should respond faster and exhibit larger CNV amplitudes on trials with full precues relative to non-informative trials (Leuthold et al., 2004). Also, a foreperiod LRP should arise for full advance precues, irrespective of the validity of the precue (which is only determined at response target onset). Thus, group differences in the precue effect and the
foreperiod LRP amplitude can again be explored. Experiment 2 therefore aimed at replicating the rather surprising foreperiod LRP results from experiment 1, namely that only musicians but not the non-musicians group, showed significant foreperiod LRP amplitudes, but similar target-related LRP, by including a full valid precue condition that allows participants to pre-specify both finger and response hand in advance.
In addition, behavioural responses to invalid precues can be used to index motor re-programming abilities as participants must modify their prepared response before executing the required movement (Leuthold & Jentzsch, 2002b). More specifically, participants are expected to respond slower on invalid precue trials compared to non-informative trials, if they engage in movement re-programming. If musicians are more efficient in these processes, re-programming costs should be smaller in musicians compared to non-musicians.
Participants
All participants from Experiment 1 took part on Experiment 2 after completion of experiment 1.
Materials and Procedures
Stimuli and apparatus were the same as in experiment apart from the changes mentioned in the following.
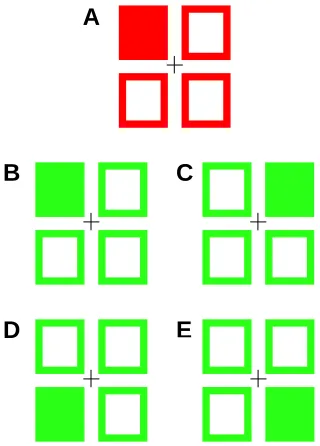
Response Precuing Task:Precues consisted of a filled red square and three empty squares, thus indicating a single finger. There were a total of five precue categories: valid full information (VFP; 50% of trials), no information (NP; 25%) and invalid information (25%) about hand (IHP), finger (IFP) or both parameters (IBP). As shown in Figure 5, the invalid conditions indicated a response alternative that was different from the finger signalled by the target, whereas valid precues
indicated the correct response finger. A higher proportion of valid trials was chosen to encourage participants to utilize precue information and prepare their responses. Participants were informed about these probabilities before the start of the
experiment. The presentation order of precue categories was randomized within each of the 6 experimental blocks (2 practice and 48 experimental trials each).
Figure 5. A) Example of a full information precue (precuing the left middle finger). B)
Valid target: Target appears in the same location as the precue. C) Invalid hand target:
Target indicates same finger (i.e. middle) but on the other hand. D) Invalid finger target:
target indicates same hand but different response finger. E) Invalid both: target indicates
a different hand and finger response.
Electrophysiological Recordings and Data Analysis
The electrophysiological recordings and data analysis were identical to Experiment 1, apart from the following changes. The mean foreperiod CNV, foreperiod LRP, and target-related LRP during trials with valid full precues and no information were submitted to mixed ANOVAs. Trials with invalid precues were not included in the ERP analyses because there were too few trials to split the data by response hand.
Results Behavioral Data – Analysis 1
The RTs and percentage of errors for experiment 2 are displayed in Figure 6. Mixed ANOVAs with within-subjects factor precue category (NP, VFP, IHP,
A
B ...
C .
D .
IFP, IBP) and between-subjects factor group (musicians vs. non-musicians) were performed on RTs for correct responses and on arcsine-transformed error rates.
Precue Information
NP VFP IFP IHP IBP
Error
Rate [%
]
0 4 8 12
Non-Musicians Musicians
Mean Reaction T
ime
[ms
]
[image:24.595.87.373.143.479.2]0 250 300 350 400 450
Figure 6: Mean RT (top) and percentage of error rates (bottom) for musicians and
non-musicians, for each of the five precue conditions in Experiment 2. NP: No Precue
Information; VFP: Valid Full Precue; IFP: Invalid Finger Precue; IHP: Invalid Hand
Precue; IBP: Invalid Both Finger and Hand Precue.
Reaction Times: Musicians tended to be faster (375 ms) than
non-musicians (402 ms), F(1, 29) = 3.92, p = .057, = .11. There was a significant effect of precue category, F(4, 116) = 42.84, p < 0.001, ηp2 = .60. Bonferoni-corrected posthoc- tests confirmed the following ordering: VFP (352 ms) < IBP (378 ms) = NP (389 ms) < IHP (408 ms) = IFP (415 ms). There was no
.97.
Error Rates: Error rates did not significantly differ between musicians (6.1%) and non-musicians (3.6%), F(1, 29) = 1.89, p = .18. There was a significant effect of precue category, F(4, 116) = 16.07, p < .001, ηp2 = .36. Bonferoni-corrected posthoc- tests confirmed the following ordering: VFP (1.2%) = NP (2.0%) = IBP (3.9%) < IHP (8.5%) = IFP (9.1%). There was no significant interaction between group and precue category, F(4, 116) = 1.74, p = .15.
Behavioural Data – Analysis 2
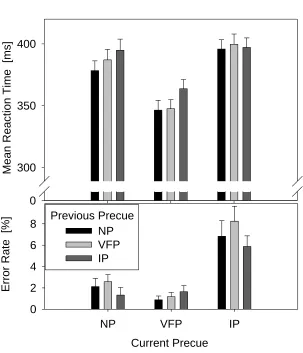
In order to investigate whether musicians and non-musicians differ in making behavioural adjustments after the experience of an invalid trial, we also performed a sequential analysis, taking into account both previous and current trial validity (e.g., Arjona, Escudero, & Gomez, 2014). For this analysis only trials with correct responses in both trials N-1 and trial N were included and the
different invalid precue condition were collapsed into one Invalid Precue (IP) condition, in order to achieve sufficient numbers of trials. The mean RTs and error rates for this analysis are displayed in Figure 7. Mixed ANOVAs with within-subjects factor previous precue category (NP, VFP, IP), current precue category (NP, VFP, IP), and between-subjects factor group (musicians vs.
Me
an R
eact
ion T
ime
[m
s]
0 300 350 400
Current Precue
NP VFP IP
Er
ror
Rat
e
[%
]
0 2 4 6 8
[image:26.595.88.391.93.446.2]NP VFP IP Previous Precue
Figure 7: Mean RTs and Error Rates for current (trial N) No Information Precues (NP);
Valid Full Precue (VFP), and Invalid Precues (IP) as a function of previous (trial N-1)
precue category.
when the previous precue had been invalid, compared to previous NP and VFP conditions. Again, no interactions with factor group were observed, all Fs <1.
Error Rates: There was a significant effect of current precue category, F(2, 58) = 31.46, p < 0.001, ηp2 = .52. There was also a significant effect of previous precue category, F(2, 58) = 4.18, p < 0.020, ηp2 = .13. No other effects reached significant levels, all ps > .10.
Electrophysiological Data
Foreperiod CNV Amplitude: A 2 (group) x 2 (precue category) mixed
ANOVA revealed a significant precue effect, F(1, 29) = 16.52, p < .001, ηp2 = 0.36, with larger (i.e. more negative) foreperiod CNV amplitude for trials that provided full advance information (-5.7 μV) compared to non-informative precues (-4.0 μV). There was no significant effect of musical practice on the foreperiod CNV amplitude (musicians: M = -5.6 μV; non-musicians: M= -4.2 μV), F(1, 29)= 0.79, p = .38, nor was there any significant interaction between precue category and group, F < .1.0.
0 500 1000 1500
CN V A mpl itud e a
t Cz [
V] -8 -6 -4 -2 0 2 Precue Target Musicians ms 0 500 1000 1500
No Precue Full Precue
Precue Target
[image:27.595.88.516.504.662.2]Non-Musicians
Figure 8. The mean foreperiod CNV amplitudes during trials with full valid and no precue
information in Experiment 2, for musicians and non-musicians. The topographic spline
map on the right depicts the spatial distribution of the average activity in a time 200
Foreperiod LRP Amplitude: Planned comparisons using one-tailed t-tests were performed on LPR amplitudes, testing whether the LRP deviates
significantly from zero in the negative direction for musicians and non-musicians. We expect the LRP to differ from zero in the full precue condition but not in the no precue condition. Musicians displayed a significant foreperiod LRP amplitude in the full precue condition (-0.74 μV), t(15) = -2.74, p= .008. The foreperiod LRP amplitude in the full hand precue condition did not differ significantly from zero for the non-musicians (-0.48 μV), t(14) = -1.05, p= .16. The LRP amplitude did not significantly differ from zero in the non-informative precue condition for either groups (both ps > 0.10).
Target-related LRP amplitude: A 2 (group) x 2 (precue category) mixed ANOVA revealed no significant effects, all Fs < 1.74, all ps > .19. Importantly, there was no significant effect of musical practice on the target-related LRP amplitude (musicians: M = 4.1 μV; non-musicians: M = -3.7 μV), F < 1. Again, this finding suggests no general group difference in lateralized motor activity.
ms
0 500 1000 1500
LRP
Ampli
tude [
V]
-4 -3 -2 -1 0 1
Musicians Non-Musicians
[image:28.595.89.322.518.696.2]Precue Target
Figure 9. The mean LRP amplitudes during trials with full valid advance information
are superimposed.
Discussion
The aim of the second experiment was two-fold. We wanted to replicate the findings of Experiment 1 using only non-information and full information precues and also test participants’ ability to re-program their response when a precue provided invalid advance information.
First, the results of Experiment 2 largely replicated the findings of the first experiment. Again, musicians tended to respond faster in general. Musicians also did not differ from non-musicians in their ability to selectively prepare a
forthcoming movement using pre-specified movement information. More specifically, whereas the precue effect itself (i.e. significantly shorter RTs and larger foreperiod CNV amplitudes on fully informative compared to
non-informative precues) was highly significant, no group effect in these measures was observed. Again, we found only musicians, but not non-musicians, to exhibit a significant foreperiod LRP when response hand was pre-specified. Importantly, lateralized activity during the response phase (target-related LRP) did not differ between groups. In order to further explore the functional significance of the foreperiod LRP amplitude, we run a correlation between the size of the precue effect (no information precue minus full information precue) in foreperiod LRP and in RTs across all participants, discarding factor group. Replicating the findings from experiment 1, the correlation was not significant, r = -0.02.
precues). However, when both parameters were invalid, no reaction time
disadvantage was observed compared to no-information precues. Interestingly, this result might suggest that it is easier to completely discard a pre-specified motor program when all movement parameters are incorrect compared to when the pre-planned program contains partially correct information and only a subset of parameters needs to be inhibited and re-programmed.
Importantly, despite the presence of clear re-programming costs, again no group differences were observed. In other words, we did not find any differences between musicians and non-musicians in the ability to re-program an invalidly pre-programmed motor response.
General Discussion
This study aimed to investigate the effects of musical ability on the behavioural and neural correlates of movement pre- and re-programming. The combined results of Experiment 1 and 2 indicate that our sample of young amateur musicians was no more efficient at preparing or re-programming their movements than non-musicians.
Participants successfully used the advance information to prepare for the forthcoming response as indicated by faster response times, lower error rates and increased CNV amplitudes, when precues provided partial (Experiment 1) or full valid (Experiment 2) compared to no advance information. Furthermore, when precues incorrectly pre-specified one movement parameter in advance,
participants showed substantial reaction time and accuracy costs compared to the no advance information condition. Interestingly, this effect was not present when all movement parameters had to be re-programmed.
finding fits with a number of previous reports suggesting a link between musical training and response speed (e.g. expert musicians: Amer et al., 2013; older adults; Khemthong et al., 2012; Amer et al., 2013; Metzler et al., 2013; amateur musicians: Hughes & Franz, 2007; Jentzsch et al., 2014). Importantly, despite these general group differences, musicians did not differ from non-musicians in their ability to selectively prepare a forthcoming movement using pre-specified movement information or to re-program in incorrectly pre-specified motor plan.
In both experiments, we found only musically trained participants, but not non-musicians, to exhibit a significant foreperiod LRP amplitude when response hand was pre-specified, despite the fact that the subsequent response target elicited an LRP of similar size in both groups. The latter finding does rule out the possibility that the lack of foreperiod LRP in the non-musician group is due to structural differences or a lower signal-to noise ratio compared to the musicians group. Interestingly, the size of the foreperiod LRP amplitude did not predict the precue effects observed in behaviour as in both experiments, no correlation between the two measures was observed.
the motor system. We could speculate more generally that effector-specific preparation as indicated by lateralized foreperiod motor activity might only have minimal behavioural consequences. However, we need to point out that these suggestions are highly speculative and need to be specifically tested in future experiments. Also, although both our sample size and our trial number is
comparable to that used in previous studies reporting foreperiod LRP effects, the power might not have been sufficient to detect significant foreperiod LRPs in the non-musicians group.
There are other limitations of the present study that suggest some caution in the interpretations of our results. The present study uses a cross-sectional design. Although we matched participant groups for age, sex, education, and mood, the group differences found in overall speed and foreperiod LRP
amplitudes may be explained by innate differences between groups rather than being the result of the musical training itself. More specifically, individuals with, for example, enlarged motor areas and other innate differences (e.g. highly intelligent individuals; Prat & Just, 2008) may be more inclined towards musical practice (Schellenberg, 2011). In addition, musically trained participants might also show a different level of engagement in non-musical activities that improve fine motor skill, such as playing video games (Drew & Waters, 1986) or sports (Krampe, 2002). In order to address this issue, we required participants to provide information about their hobbies (“List any other non-music-related hobbies you have” and “How many hours per month you spend on each of
groups). However, participants were not explicitly asked about their involvement in physical and video gaming activities.
The lack of evidence for motor planning or re-programming differences between groups in our study does not necessarily contradict the hypothesis that musically trained individuals are more at efficient movement preparation
compared to non-musicians (Jäncke et al., 2000), due to several other
methodological shortcomings. For example, a few of the individuals classified as ‘non-musicians’ possessed some musical experience (<1000 hours), which may have reduced the likelihood of detecting significant group differences. Consistent with this, in an earlier study (Jentzsch et al., 2014) we found that even individuals with low levels of musical practice (i.e. 200-2000 hours) display shorter RT
latencies compared to individuals with no musical experience. Also, our musically trained participant group might not have a sufficiently high number of practice hours to yield a significant effect of musical training on movement
within a 10-year period; Gruhn et al., 2003), nor do they to older adults. Nevertheless, given the general group differences in response speed and
lateralized foreperiod LRP activity observed in the present study, it is unlikely that the lack of group difference in the specific pre- and re-programming measures can be attributed to these shortcomings.
References
Altenmüller, E., Marco‐Pallares, J., Münte, T.F., & Schneider, S. (2009). Neural Reorganization Underlies Improvement in Stroke‐induced Motor Dysfunction by Music‐supported Therapy. Annals of the New York Academy of Sciences, 1169, 395-405. DOI: 10.1111/j.1749-6632.2009.04580.x
Amer, T., Kalender, B., Hasher, L., Trehub, S E., & Wong, Y. (2013).
Do older professional musicians have cognitive advantages?. PloS one, 8, e71630. DOI: 10.1371/journal.pone.0071630
Amunts, K., Schlaug, G., Jäncke, L., Steinmetz, H., Schleicher, A., Dabringhaus, A., & Zilles, K. (1997). Motor cortex and hand motor skills: structural
compliance in the human brain. Human Brain Mapping, 5, 206-215. DOI: 10.1002/(SICI)1097-0193(1997)5:3<206::AID-HBM5>3.0.CO;2-7
Arjona, A., Escudero, M., & Gomez, C.M. (2014). Updating of attentional and premotor allocation resources as function of previous trial outcome. Scientific Reports, 4, 4526. DOI: 10.1038/srep04526
Bangert, M., & Schlaug, G. (2006). Specialization of the specialized in features of external human brain morphology. European Journal of Neuroscience, 24, 1832-1834. DOI: 10.1111/j.1460-9568.2006.05031.x
Baumann, S., Koeneke, S., Schmidt, C.F., Meyer, M., Lutz, K., and Jancke, L. (2007). A network for audio–motor coordination in skilled pianists and non-musicians. Brain Research, 1161, 65-78. DOI:
10.1016/j.brainres.2007.05.045
Brunia, C.H.M., & van Boxtel, G.J.M. (2001). Wait and see. International Journal of Psychophysiology, 43(1), 59-75. DOI: 10.1016/S0167-8760(01)00179-9
Carrillo-de-la-Peña, M.T., Galdo-Alvarez, S., & Lastra-Barreira, C. (2008). Equivalent is not equal: Primary motor cortex (MI) activation during motor imagery and execution of sequential movements. Brain Research, 1226, 134-143. DOI: 10.1016/j.brainres.2008.05.089
Churchland, M.M., Yu, B.M., Ryu, S. I., Santhanam, G., & Shenoy, K.V. (2006). Neural variability in premotor cortex provides a signature of motor
preparation. The Journal of Neuroscience, 26, 3697-3712. DOI: 10.1523/JNEUROSCI.3762-05.2006
Coles, M.G.H., Smid, H.G.O.M., Scheffers, M.K., & Otten, L.J. (1995). Mental chronometry and the study of human information processing. In: M.D. Rugg & M.G.H. Coles (Eds.), Electrophysiology of Mind: Event-related Brain
Potentials and Cognition (pp. 86-131). Oxford: Oxford University Press.
movements in man: Cerebral potentials and theory. Biological Cybernetics, 23, 99-119. DOI: 10.1007/BF00336013
Drew, B., & Waters, J. (1986). Video games: Utilization of a novel strategy to improve perceptual motor skills and cognitive functioning in the non-institutionalized elderly. Cognitive Rehabilitation, 4, 26-31.
Elbert, T., Pantev, C., Wienbruch, C., Rockstroh, B., & Taub, E. (1995). Increased cortical representation of the fingers of the left hand in string players. Science, 270, 305-307. DOI: 10.1126/science.270.5234.305
Engel, K.C., Flanders, M., & Soechting, J.F. (1997). Anticipatory and sequential motor control in piano playing. Experimental Brain Research, 113, 189-199.
DOI: 10.1007/BF02450317
Franěk, M., Mates, J., Radil, T., Beck, K., & Pöppel, E. (1991). Finger tapping in musicians and nonmusicians. International Journal of Psychophysiology, 11, 277-279. DOI: 10.1016/0167-8760(91)90022-P
Friedman, D. (2000). Event-related brain potential investigations of memory and aging. Biological Psychology, 54, 175-206. DOI:
10.1016/S0301-0511(00)00056-9
Gaser, C., & Schlaug, G. (2003). Gray matter differences between musicians and nonmusicians. Annals of the New York Academy of Sciences, 999, 514-517.
DOI: 10.1196/annals.1284.062
Gomez, C.A., Flores, A., & Ledesma, A. (2007). Fronto-parietal networks activation during the contingent negative variation period. Brain Research Bulletin, 73(1-3), 40-47. DOI: 10.1016/j.brainresbull.2007.01.015
Gomez, C. M., Fernandez, A., Maestu, F., Amo, C., Gonzalez-Rosa, J.J.,
Vaquero, E., & Ortiz, T. (2004). Task-specific sensory and motor preparatory activation revealed by contingent magnetic variation. Cognitive Brain
Research, 21(1), 59-68. 10.1016/j.cogbrainres.2004.05.005
Goodman, D., & Kelso, J.S. (1980). Are movements prepared in parts? Not under compatible (naturalized) conditions. Journal of Experimental Psychology: General, 109, 475-495. DOI: 10.1037//0096-3445.109.4.475
Gratton, G., Coles, M.G.H., Sirevaag, E. J., Eriksen, C.W., & Donchin, E. (1988). Pre-and poststimulus activation of response channels: a
psychophysiological analysis. Journal of Experimental Psychology: Human
Perception and Performance, 14, 331-344.DOI:
10.1037/0096-1523.14.3.331
Hanna-Pladdy, B., & MacKay, A. (2011). The relation between instrumental musical activity and cognitive aging. Neuropsychology, 25, 378 -386. DOI: 10.1037/a0021895
Herholz, S.C., & Zatorre, R.J. (2012). Musical training as a framework for brain plasticity: behavior, function, and structure. Neuron, 76, 486-502. DOI: 10.1016/j.neuron.2012.10.011
Hutchinson, S., Lee, L.H., Gaab, N., & Schlaug, G. (2003). Cerebellar volume of musicians. Cerebral Cortex 13, 943–949. DOI: 10.1093/cercor/13.9.943
Hughes, C.M., & Franz, E.A. (2007). Experience-dependent effects in unimanual and bimanual reaction time tasks in musicians. Journal of Motor Behavior, 39, 3-8. DOI: 10.3200/JMBR.39.1.3-8
Hyde, K.L., Lerch, J., Norton, A., Forgeard, M., Winner, E., Evans, A.C., & Schlaug, G. (2009). The effects of musical training on structural brain development. Annals of the New York Academy of Sciences, 1169, 182-186. DOI: 10.1111/j.1749-6632.2009.04852.x
Jabusch, H.C. (2006). Movement analysis in pianists. In: E. Altenmüller, M. Wiesendanger and J. Kesselring (Eds.) Music, Motor Control and the Brain
(pp. 91-108). New York: Oxford University Press.
Jäncke, L., Shah, N. J., & Peters, M. (2000). Cortical activations in primary and secondary motor areas for complex bimanual movements in professional pianists. Cognitive Brain Research, 10, 177-183. DOI: 10.1016/S0926-6410(00)00028-8
Jentzsch, I., & Leuthold, H. (2002). Advance movement preparation of eye, foot, and hand: a comparative study using movement-related brain potentials.
Cognitive Brain Research, 14, 201-217. DOI: 10.1016/S0926-6410(02)00107-6
Jentzsch, I., Leuthold, H., & Ridderinkhof, R. (2004). Beneficial effects of ambiguous precues: Parallel motor preparation or reduced premotoric processing time?. Psychophysiology, 41, 231-244. DOI: 10.1111/j.1469-8986.2004.00155.x
Jentzsch, I., Mkrtchian, A., & Kansal, N. (2014). Improved effectiveness of performance monitoring in amateur instrumental musicians.
Neuropsychologia, 52, 117-124. DOI: 10.1016/j.neuropsychologia.2013.09.025
Jerde, T.E, Santello, M., Flanders, M., & Soechting, J.F. (2006). Hand
Keele, S.W. (1968). Movement control in skilled motor performance.
Psychological Bulletin, 70, 387-403. DOI: 10.1037/h0026739
Khemthong, S., Pejarasangharn, U., Uptampohtiwat, T., & Khamya, A. (2012). Effect of Musical Training on Reaction Time A Randomized Control Trial in Thai Elderly Individuals. Music and Medicine, 4, 16-21.
Krampe, R.T. (2002). Aging, expertise and fine motor movement. Neuroscience & Biobehavioral Reviews, 26, 769-776. DOI:
10.1016/S0149-7634(02)00064-7
Landau, S.M., & D'Esposito, M. (2006). Sequence learning in pianists and nonpianists: an fMRI study of motor expertise. Cognitive, Affective & Behavioral Neuroscience, 6, 246-259. DOI: 10.3758/CABN.6.3.246 Larish, D.D., & Frekany, G.A. (1985). Planning and preparing expected and
unexpected movements: reexamining the relationships of arm, direction, and extent of movement. Journal of Motor Behavior, 17, 168-189.
Lépine, D., Glencross, D., & Requin, J. (1989). Some experimental evidence for and against a parametric conception of movement programming. Journal of Experimental Psychology: Human Perception and Performance, 15,
347-362. DOI: 10.1037//0096-1523.15.2.347
Leuthold, H. (2003). Programming of expected and unexpected movements: Effects on the onset of the lateralized readiness potential. Acta
Psychologica, 114, 83-100. DOI: 10.1016/S0001-6918(03)00051-9
Leuthold, H., & Jentzsch, I. (2001). Neural correlates of advance movement preparation: a dipole source analysis approach. Cognitive Brain Research, 12, 207-224. DOI: 10.1016/S0926-6410(01)00052-0
Leuthold, H., & Jentzsch, I. (2002a). Distinguishing neural sources of movement preparation and execution: An electrophysiological analysis. Biological Psychology, 60, 173-198. DOI: 10.1016/S0301-0511(02)00032-7
Leuthold, H., & Jentzsch, I. (2002b). Spatiotemporal source localisation reveals involvement of medial premotor areas in movement reprogramming.
Experimental Brain Research, 144, 178-188. DOI: 10.1007/s00221-002-1043-7
Leuthold, H., Sommer, W., & Ulrich, R. (2004). Preparing for action: Inferences from CNV and LRP. Journal of Psychophysiology, 18, 77-88. doi:
10.1027/0269-880318.2-3.77
Loveless, N.E, & Sanford, A.J. (1974). Slow potential correlates of preparatory set. Biological Psychology, 1(4),303-14. DOI:10.1016/0301-0511(74)90005-2
correlates of preparation for movement direction and force.
Electroencephalography and Clinical Neurophysiology, 76, 47-62. DOI: 10.1016/0013-4694(90)90057-Q
Metzler, M.J., Saucier, D.M., & Metz, G.A. (2013). Enriched childhood
experiences moderate age-related motor and cognitive decline. Frontiers in Behavioral Neuroscience, 7, 1-8. DOI: 10.3389/fnbeh.2013.00001
Münte, T.F., Altenmüller, E., & Jäncke, L. (2002). The musician's brain as a model of neuroplasticity. Nature Reviews Neuroscience, 3, 473-478. DOI: 10.1038/nrn843
Nagai, Y., Critchley, H.D., Featherstone, E., Fenwick, P.B.C., Trimble, M.R., & Dolan, R.J. (2004). Brain activity relating to the contingent negative
variation: an fMRI investigation. Neuroimage, 21, 1232-1241. DOI: 10.1016/j.neuroimage.2003.10.036
Osman, A., Moore, C.M., & Ulrich, R. (2003). Temporal organization of covert motor processes during response selection and preparation. Biological Psychology, 64, 47-75. DOI: 10.1016/S0301-0511(03)00102-9
Palmer, C. (2006). The nature of memory for music performance skills. In: E. Altenmüller, M. Wiesendanger & J. Kesselring (Eds.) Music, Motor Control
and the Brain (pp. 39-53). New York: Oxford University Press.
Palmer, C., & Dalla Bella, S. (2004). Movement amplitude and tempo change in piano performance. Journal of the Acoustical Society of America, 116, 1842-1842.
Palmer, C., & Pfordresher, P.Q. (2003). Incremental planning in sequence production. Psychological Review, 110, 683-712. DOI: 10.1037/0033-295X.110.4.683
Pascual-Leone, A., Amedi, A., Fregni, F., & Merabet, L.B. (2005). The plastic human brain cortex. Annual Review of Neuroscience, 28, 377-401. DOI: 10.1146/annurev.neuro.27.070203.144216
Prat, C.S., & Just, M.A. (2008). Brain bases of individual differences in cognition.
Psychological Science Agenda, 22, 1-6.
Repp, B.H., & Doggett, R. (2007). Tapping to a very slow beat: a comparison of musicians and nonmusicians. Music Perception: An Interdisciplinary Journal,
24, 367-376.DOI: 10.1525/MP.2007.24.4.367
Rohrbaugh, J.W., & Gaillard, A.W. (1983). Sensory and Motor Aspects of the Contingent Negative Variation. Advances in Psychology, 10, 269-310. Rohrbaugh, J.W., Syndulko, K., & Lindsley, D.B. (1976). Brain wave components
of the contingent negative variation in humans. Science, 191, 1055-1057.
Rosenbaum, D.A. (1980). Human movement initiation: specification of arm, direction, and extent. Journal of Experimental Psychology: General, 109, 444-474. DOI: 10.1037/0096-3445.109.4.444
Rosenbaum, D.A. (1983). The movement precuing technique: Assumptions, applications, and extensions. Advances in Psychology, 12, 231-274. Schellenberg, E.G. (2011). Examining the association between music lessons
and intelligence. British Journal of Psychology, 102, 283-302. DOI: 10.1111/j.2044-8295.2010.02000.x
Schneider, S., Schönle, P. W., Altenmüller, E., & Münte, T.F. (2007). Using musical instruments to improve motor skill recovery following a stroke.
Journal of Neurology, 254, 1339-1346. DOI: 10.1007/s00415-006-0523-2
Seidler, R.D., Bernard, J.A., Burutolu, T.B., Fling, B.W., Gordon, M.T., Gwin, J.T., Kwak, Y., & Lipps, D.B. (2010). Motor control and aging: links to age-related brain structural, functional, and biochemical effects. Neuroscience &
Biobehavioral Reviews, 34, 721-733. DOI: 10.1016/j.neubiorev.2009.10.005
Shojaei, M., & VaezMousavi, S.M. (2007). The Effect of the Nature of Precued Parameters on Reaction Time of a Force Production Task. International
Journal of Applied Sports Sciences, 19, 12-24.
Sloboda, J. & Davidson, J. (1996). The young performing musician. In: I. Deliège and J. A. Sloboda (Eds.) Musical Beginnings: Origins and Development of
Musical Competence (pp. 171-190). Oxford: Oxford University Press.
Stelmach, G.E., Goggin, N.L., & Amrhein, P.C. (1988). Aging and the
restructuring of precued movements. Psychology and Aging, 3, 151-157.
DOI: 10.1037//0882-7974.3.2.151
Ulrich, R., Leuthold, H., & Sommer, W. (1998). Motor programming of response force and movement direction. Psychophysiology, 35, 721-728. DOI: 10.1111/1469-8986.3560721
Walter, W., Cooper, R., Aldridge, V.J., McCallum, W.C., & Winter, A.L. (1964). Contingent negative variation: an electric sign of sensori-motor association and expectancy in the human brain. Nature, 203, 380-384. DOI:
10.1038/203380a0
Wan, C.Y., & Schlaug, G. (2010). Music making as a tool for promoting brain plasticity across the life span. The Neuroscientist, 16, 566-577. DOI: 10.1177/1073858410377805
Ward, N.S., & Frackowiak, R.S.J. (2003). Age‐related changes in the neural correlates of motor performance. Brain, 126, 873-888. DOI:
10.1093/brain/awg071
brief measures of positive and negative affect: The PANAS scales. Journal of Personality and Social Psychology, 54, 1063-1070. DOI: 10.1037/0022-3514.54.6.1063
Watson, A.H.D. (2006). What can studying musicians tell us about motor control of the hand?. Journal of Anatomy, 208, 527-542. DOI: 10.1111/j.1469-7580.2006.00545.x
Wild-Wall, N., Sangals, J., Sommer, W., & Leuthold, H. (2003). Are fingers special? Evidence about movement preparation from event–related brain potentials. Psychophysiology, 40, 7-16. DOI: 10.1111/1469-8986.00002
Williamon, A., & Valentine, E. (2000). Quantity and quality of musical practice as predictors of performance quality. British Journal of Psychology, 91, 353-376. DOI: 10.1348/000712600161871