BARN OWLS (
185
0
0
Full text
(2)
(3)
(4)
(5)
(6)
(7)
(8)
(9)
(10)
(11)
(12)
(13)
(14)
(15)
(16)
(17)
(18)
(19)
(20)
(21)
(22)
(23)
(24)
(25)
(26)
(27)
(28)
(29)
(30)
(31)
(32)
(33)
(34)
(35)
(36)
(37)
(38)
(39)
(40)
(41)
(42)
(43)
(44)
(45)
Figure


+7



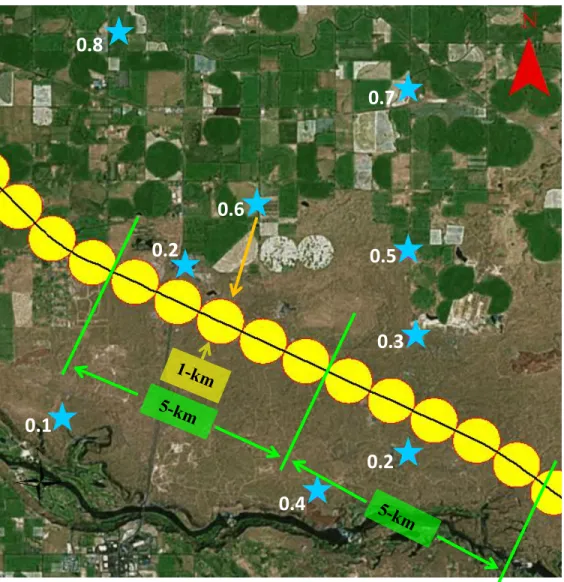
Related documents
Sectional specification: Fixed surface mount multilayer capacitors of ceramic dielectric, Class 2..
Recognized degree that includes completion of a teacher preparation program consisting of a minimum of 48 semester hour credits of education coursework and supervised student
As in the case of individuals with chronic conditions, Dutch respondents put a lower value on extra insurer services than the German subsample, and they have no positive WTP for a
We systematically reviewed quantitative and qualitative evidence on the associations between gender inequities and women’s and children’s nutri- tional status, their diets,
One way to keep up your intake of calcium-rich foods is to enjoy a variety of cultured dairy products including buttermilk, acidophilus milk, yogurt, frozen yogurt, and Swiss
Ekzistojne dy lloje te bashkimit te veprave: bashkimi ideal dhe bashkimi real.. BASHKIMI IDEAL – Ekziston kur personi me nje veprim kryen dy apo me shume vepra penale per te
Limitation of Liability - As Macleay Valley Travel acts only as agent for the Airlines, Hotels, Coach Companies etc, providing accommodation, transportation and other services,
faithful partner in marriage…expectations were mutually concordant and a self-enforcing convention was locked in.” 86 With regard to dating, the idea that, “I can’t start dating
