_____________________________________________________________________________________________________ *Corresponding author: Email: [email protected];
ISSN: 2231-0614
SCIENCEDOMAIN international
www.sciencedomain.org
Role of Gibberellic and Indole Acetic Acid in
Altering Ocular Structure and Function of Mother
Rats and their Offspring
H. I. H. El-Sayyad
1*, M. M. Ramadan
1, M. H. Abou-Egla
1, Adel M. A. Asiri
2and M. E. El-Beeh
1,21
Department ofZoology, Faculty of Science, Mansoura University, Mansoura, Egypt. 2
Institiute of Scientific Research and Revival of Islamic Heritage, Um Al-Qura University, Makkah, Kingdom Saudia Arabia and Zoology Department, Faculty of Science, Mansoura University,
Mansoura, Egypt.
Authors’ contributions
This work was carried out in collaboration between all authors. All authors read and approved the final manuscript.
Article Information
DOI:10.9734/BJMMR/2015/19964 Editor(s): (1) Shashank Kumar, Department of Biochemistry, University of Allahabad, Allahabad, India. Reviewers: (1) Anonymous, Austria. (2)Atef Mahmoud Mahmoud Attia, Biophysical Laboratory, National Research Centre, Egypt. (3)Arturo Solis Herrera, Human Photosynthesis Research Center, Mexico. Complete Peer review History:http://sciencedomain.org/review-history/11406
Received 4th July 2015 Accepted 31st July 2015 Published 16th September 2015
ABSTRACT
Recently, plant growth hormones have been widely used in green houses and the consumptions of agriculture food products have markedly increased; however their toxicological aspects are still unclear. The aim of the present study is to illustrate the toxicities of plant growth hormones; gibberellic (GA3) and indole acetic acid (IAA) on ocular tissues of pregnant Wistar albino rats and their offsprings. Intra-gastric administration of either GA3 or IAA (100 mg/kg body weight) to mother rats were carried out for one month prior to conception as well as throughout gestation period (n=20). The control received saline-free organochemical compounds (n=10). At parturition, mother rats and their offsprings were sacrificed by light anesthesia, and their ocular regions were separated and investigated for light and transmission electron microscopy, immunochemical staining of Caspase-3, Bcl-2 and p53 and assessments of antioxidant enzymes and DNA fragmentation. Maternal administration of either GA3 or IAA was found to develop cataractous
lenses to about 6% (3/20) in GA3-treatment and 4% (2/20) in IAA-treatment. The applied organochemical compounds developed retinopathy in mother rats and their offsprings. These were characterized by apparent damage of ganglion and nuclear cells. In mothers, the photoreceptor’s outer segment showed spotty dissolution of their stacked membranes while in offsprings, there was still lack of differentiation. The retinal tissue showed discrete positive immunostaining foci with P53, Pcl2 and caspase-3, especially in ganglion and nuclear cells compared with control. The antioxidant enzymes' catalase (CAT), superoxide dismutase (SOD), glutathione reductase (GR) and peroxidase (GPx) showed apparent depletion and vice versa for malondialdhyde (MDA) and hydrogen peroxide (H2O2), the markers of cell death. DNA fragmentation was markedly detected.
Finally, it was concluded that maternal administration of GA3 and IAA led to the development of retinopathy and development of cataractous lenses parallel to the increase of immunostaining of caspase-3, Pcl2 and P53 and decrease of the endogenous antioxidant system and increase of DNA damage.
Keywords: Lens; retina; mother rats; offsprings; gibberellic and indole acetic acid.
1.INTRODUCTION
Vision is very important and there is little information about the epidemiology of ocular disease among the pre-school-aged children [1]. Recently, plant growth hormones have been widely used in green houses [2] and their amounts exceed the used insecticides in agriculture [3]. Although, the consumptions of agriculture food products have increased, the toxicological aspects of plant growth hormones are still unclear [4-7]. Plant growth hormones (PGHs) are divided into different groups of which GA3 belongs to gibberellins [8,9] and IAA, a member of auxins [10]. 0They are used to increase the growth and productivity of vegetables and fruits. Application of 1000 ppm GA3 to grapefruit trees was found to increase fruit size and decrease the concentration of the total acid and naringin concentration in the juice [11]. Indole acetic acid-treatment (10−4 and 10−2 M) helps to accelerate the ripening of fruits and also increase the levels of both ascorbic acid and glucose to a maximum level at the climacteric, but it decreases during ripening [12].
Concerning the toxicological aspects, gibberellic acid was found to induce hepatocellular carcinoma in Egyptian toad [4], hepatotoxicities in experimental animals [13-15] and damage of cerebellar neurons [15,16]. Also, microencephaly was reported in post-IAA-treatment [17]. Agri-cultural workers were found to be at high risk of ocular toxicities following exposure to pesticides [18]. Rat was proven to be a good model for the different pattern of ocular diseases [19-21].
The present study aimed to assess the ocular toxicities of GA3 and IAA in pregnant rats and their off springs.
2. MATERIALS AND METHODS
All experiments were conducted in accordance with the national laws for the use of animals in research and approved by the local ethical committee.
2.1 Application of Plant Growth
Hormones
The plant growth hormones; GA3 and IAA were purchased from Sigma Chemical Company (St. Louis, MO 6, USA). The applied dose of 100 ppm/kg body weight was orally administered, every other day for one month prior to conception as well as throughout gestation according to Troudi et al. [15].
2.2 Experimental Work
groups (n=10); control, GA3 & IAA treatments. At parturition, the pregnant female
control and experimentally-treated mothers were sacrificed by light anesthesia, and their occular regions were removed. Seven of the mothers were dissected, and their occular tissues were separated and kept in a refrigerator for biochemical and DNA investigation. For light and transmission electron microscopy, three of the mothers undergone anesthesia and were intraventrically injected with either 2.5% glutaraldhyde or 10% phosphate buffered formalin to allow primary fixation before incision. At the same time, thirty offsprings were separated from both control and experimental groups. Unilateral incisions of the right eye were carried out for biochemical investigations (n=20) and DNA damage (n=10), meanwhile the other ones were arranged for light (n=25) and transmission electron microscopy (n=5). Following light microscopic investigations; the incidence of cataractous lenses was recorded.
2.3 Antioxidant Enzymes Assays
At parturition, fasted pregnant rats
offsprings of control and experimental groups were sacrificed by light chloroform anesthesia and dissected. The right eye regions of 10 mothers and 20 offsprings were separated and categorize into five replicates per each group. The retinal tissues were homogenized and centrifuged at 5000 rpm for 5 min. The supernatants were separated and kept frozen at -20ºC for determination of CAT and
reductase (GR) and peroxidase (GP) and SOD. Glutathione reductase (GSH reductase) was determined according to Prins and Loose [22] depending on the precipitation of protein using tungestate-sulfuric acid solution and formation of yellow color after reaction with 5,5` dithiobis- 25-nitrobenzoic acid (DNTB) and read at a wavelength of 412 nm.
On the other hand, glutathione peroxidase (GPx) (EC 1.11.1.9) was determined according to Paglia and Valetine [23] depending on the production of oxidized glutathione after reducing of organic peroxide by GPx, the oxidation of NADPH to NADP+ is accompanied by a decrease in absorbance at 340 nm.
Regarding CAT (EC 1.11.1.6), determination was carried out according to Bock et al.
assay is based on the disappearance of H the presence of the enzyme activity. The reaction mixture containing 0.05 M phosphate buffer (pH 7.0), 1.2 mM H2O2 and 0.2 ml of
Sayyad et al.; BJMMR, 10(12): 1-14, 2015; Article no.
treatments. At rats of both treated mothers were sacrificed by light anesthesia, and their occular regions were removed. Seven of the mothers were dissected, and their occular tissues were ed and kept in a refrigerator for biochemical and DNA investigation. For light and transmission electron microscopy, three of the mothers undergone anesthesia and were intraventrically injected with either 2.5% glutaraldhyde or 10% phosphate buffered lin to allow primary fixation before incision. At the same time, thirty offsprings were separated from both control and experimental groups. Unilateral incisions of the right eye were carried out for biochemical investigations (n=20) meanwhile the other ones were arranged for light (n=25) and transmission electron microscopy (n=5). Following light microscopic investigations; the incidence of cataractous lenses was recorded.
2.3 Antioxidant Enzymes Assays
pregnant rats and their offsprings of control and experimental groups were sacrificed by light chloroform anesthesia and dissected. The right eye regions of 10 mothers and 20 offsprings were separated and categorize into five replicates per each group. retinal tissues were homogenized and centrifuged at 5000 rpm for 5 min. The eparated and kept frozen at C for determination of CAT and glutathione GP) and SOD. Glutathione reductase (GSH reductase) was determined according to Prins and Loose [22] depending on the precipitation of protein using sulfuric acid solution and the formation of yellow color after reaction with 5,5` nitrobenzoic acid (DNTB) and read
On the other hand, glutathione peroxidase (GPx) (EC 1.11.1.9) was determined according to Paglia and Valetine [23] depending on the production of oxidized glutathione after reducing of organic peroxide by GPx, the oxidation of ccompanied by a
Regarding CAT (EC 1.11.1.6), determination et al.[24]. The assay is based on the disappearance of H2O2 in
the presence of the enzyme activity. The reaction mixture containing 0.05 M phosphate and 0.2 ml of
homogenate were allowed to stand for 30 min. The absorbance of the sample was read against distilled water at zero time (A1) and after 30 s (A2) at 240 nm.
On the other hand, the SOD (EC1.15.1.1) was determined according to Niskikimi
method depends on the formation of reduced nitro blue tetrazolium and absorbance was read at 560 nm.
2.4 Determination of Lipid Peroxides and Free Radicals
The level of malondialdehyde (lipid peroxidation end product) was determined according to Ohkawa et al. [26] after estimating the reddish pink color developed from incubation of the retinal tissue homogenate (0.2 ml) with sodium dodecyl sulfate (100μl), acetic acid (750 20%), thiobarbituric acid (50μl of 0.6%) and distilled water (300μl). The colour is estimated at 532 nm and the extent of lipid peroxidation is expressed as nmol/mg protein.
Also, hydrogen peroxide content in retinal tissues was determined after incubating the retinal tissues with 3, 5-dichloro
benzensulfonic acid and 4-aminophenazone and absorbance was determined at 510 nm
2.5 Light Microscopic Investigations
One-day old retina from maternal and offspring of both control and experimentally treated groups were immediately fixed in 10% phosphate buffered formalin (pH 7.4), dehydrated in ascending grades of ethyl alcohol, cleared in xylene and mounted in molten paraplast at 58-62ºC. Serial 5 µm sagital histological sections were cut and stained with hematoxylin and eosin and examined under bright-field light microscopy. The incidence of cataractous lenses was determined in offsprings after investigating twenty offspring eyes. In case of maternal retina, the thickness of whole retina as well as inner and outer nuclear (INL and ONL) and inner and outer plexiform (IPL and OPL) were determined using linear ocular micrometer.
2.6 Transmission Electron Microscopic (TEM) Investigations
TEM was carried out for three specimens of both maternal and offspring's retina. Retina speci mens of both control and experimental groups were separated and fixed in cold 2.5% glutaral
; Article no.BJMMR.19964
homogenate were allowed to stand for 30 min. of the sample was read against ) and after 30 s
On the other hand, the SOD (EC1.15.1.1) was determined according to Niskikimi et al. [25]. The method depends on the formation of reduced nitro blue tetrazolium and absorbance was read
Lipid Peroxides and
(lipid peroxidation determined according to . [26] after estimating the reddish pink color developed from incubation of the retinal tissue homogenate (0.2 ml) with sodium l), acetic acid (750μl of l of 0.6%) and ur is estimated at 532 nm and the extent of lipid peroxidation is
Also, hydrogen peroxide content in retinal tissues was determined after incubating the dichloro-2-hydroxy-minophenazone and absorbance was determined at 510 nm[27].
2.5 Light Microscopic Investigations
day old retina from maternal and offspring of both control and experimentally treated groups were immediately fixed in 10% phosphate buffered formalin (pH 7.4), dehydrated in ascending grades of ethyl alcohol, cleared in xylene and mounted in molten C. Serial 5 µm sagital histological sections were cut and stained with hematoxylin and eosin and examined under field light microscopy. The incidence of cataractous lenses was determined in offsprings pring eyes. In case of maternal retina, the thickness of whole retina as well as inner and outer nuclear (INL and ONL) and inner and outer plexiform (IPL and OPL) were determined using linear ocular
Transmission Electron Microscopic
glutaral-dhyde in cacodylate buffer (pH 7.4) for about five hours followed by washing in phosphate buffer (pH 7.4) and post-fixation in buffered 1% osmium tetraoxide. The specimens were washed thoroughly in buffer and dehydrated in ascending series of cold ethyl alcohol, cleared in propylene oxide and mounted in epoxy resin. Ultrathin sections were cut with a LKB Ultratome IV (LKB Instruments, Bromma, Sweden), mounted on grids, stained with uranyl acetate and lead citrate and examined at a joel 100CXl Transmission electron microscope (Musashino 3-chome Akishima Tokyo 196-8558, Japan).
2.7 Caspase-3, Bcl-2 and p53 Immuno-histochemical Staining
Five micrometer (5 μm) histological sections of formalin-fixed, paraffin-embedded retinal tissues were cut and placed on polylysine-coated glass slides. After overnight packing at 65ºC, tissue sections were deparaffinized in xylene and rehydrated in descending grades of alcohol. Endogenous peroxidase activity was exhausted by incubation of tissue sections in 3% H2O2 for
10 min at room temperature. Histological pre-pared slides were selected for the primary monoclonal mouse anti-human antibody of caspase-3 (DAKO, clone MIB5, 1:50, mouse), Bcl-2 (124 Dako, Carpinteria, Calif.) and p53 (DO7, Dako, Carpinteria, Calif.) and were incubated overnight at 4ºC. After washing with buffer, slides were incubated with a secondary biotin linked anti-mouse antibody for 50 min at room temperature; and with the streptoavidin-peroxidase complex for 50 min. Buffer washing was carried out between each step. Sections were then washed with distilled water, incubated with developing solution (diaminobenzidine-hydrogen peroxide; DAKO), and counterstained with hematoxylin. Immunoreactivity was identi-fied as brown nuclear or cytoplasmic labelling in retinal and lens counterstained with hematoxylin. Negative controls were performed for all cases by omitting the primary antibody, which in all instances resulted in negative immunoreactivity. Finally, the sections were examined under bright field light olympus microscope with a digital canon camera.
2.8 Single Cell Gel Electrophoresis
(Comet Assay) and DNA
Fragmentation:
In case of Comet assay, five fresh retinal specimens of both control and experimental groups were homogenized in chilled
homo-genizer buffer, pH 7.4, containing 75 mM NaCl and 24 mM Na2 ethylene-diamine-tetra-acetic
acid (EDTA), pH 13, to obtain a 10% tissue solution. The homogenates were kept on ice during and after homogenization. Six micro liters (6 µl) of the homogenate was suspended on 0.5% low melting agarose and sandwiched between a layer of 0.6% normal-melting agarose and a top layer of 0.5% low melting agarose on fully frosted slides. The slides were kept on ice during the polymerization of each gel layer. After the solidification of the 0.6% agarose layer, the slides were immersed into a lysis solution (1% sodium surcosinate, 2.5 M NaCl, 100 mM Na2EDTA, 10 mm Tris-HCl, 1% Triton X-100,
and 10% DMSO) at 4ºC. After 1 h, the slides were placed into the electrophoresis buffer (0.3 M NaOH, 1 mM Na2EDTA, pH 13) for 10 min at
37ºC to allow DNA to unwind. Electrophoresis was performed for 10 min at 300 mA and 1 V/cm. The slides were neutralized with Tris-HCl buffer, pH 7.5, and stained with 20 mg/ml ethidium bromide. Each slide was analyzed using a Leitz Orthoplan (Wetzlar, Germany) epifluorescence microscope. One hundred cells were analyzed on each slide using the Comet assay II automatic digital analysis system. Both tail length and tail intensity were measured automatically by image analysis software [28]. DNA fragmentation was carried out according to Duke and Sellins [29]. Tissue samples of both control and experimental groups were mixed thoroughly with 100 ml of lyses buffer (10 mM Tris HCl/10 mM EDTA/0.5% Triton X-100, pH 8.0), and centrifugation for 20 min at 4ºC (14,000 x g). The spernatant was transferred to another tube. DNA concentrations were determined. DNA electrophoresis was carried out on 1.5% agarose gel and compared with a ladder marker.
2.9 Statistical Analysis
The obtained data were subjected to statistical analysis using Student’s t-test. All values were expressed as means±S.E.M. and the level of significance for all experiments was considered at p value < 0.05.
3. RESULTS
3.1 Antioxidant Enzymes and Malon-dialdhyde
Sayyad et al.; BJMMR, 10(12): 1-14, 2015; Article no.BJMMR.19964
SOD, CAT, GR and GP showed a considerable reduction after treatment with both hormones. There were marked increase of malondialdehyde (lipid peroxidation product) and reactive oxygen species, especially of those subjected to IAA- treatment.
3.2 Morphometry of Maternal Retinal Thickness
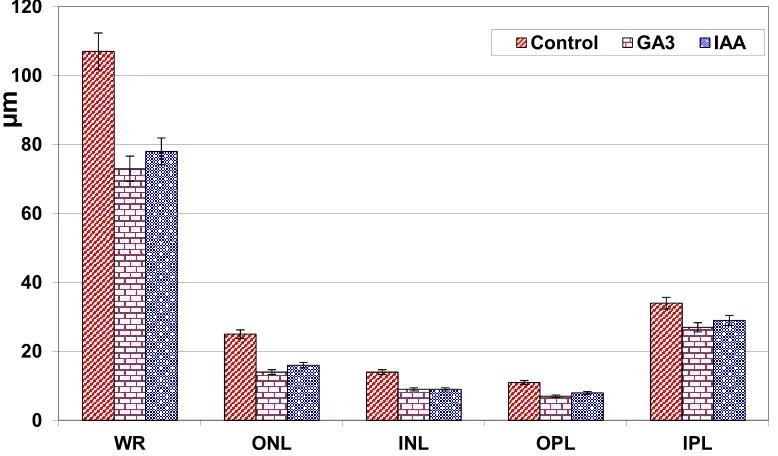
Measurements of whole retina, INL and ONL and IPL and OPL thickness were performed on the retinal sections in the control and GA3 and IAA-treated mothers. The thickness of the assayed whole retina revealed a significant reduction after GA3 and IAA treatment, meanwhile the retinal layers showed non-significant change (Fig. 1).
3.3 Light and Transmission Electron Microscopy of Maternal Retina
At the light microscopic level, the control retina is composed of eight regularly arranged cell layers; the retinal nerve fiber and ganglion layer, inner plexiform, inner nuclear, outer plexiform, outer nuclear, photoreceptors and pigmented layer. The ganglion cells appear at one-two cell
layered thick. The photoreceptors are clearly differentiated into rod and cone cells. A single-cell layer of the pigmented epithelium with a prominent basal lamina underlined by Bruch's membrane that separates from the chorio-capillaries is shown in Fig. 2A.
In experimental gibberellic and indole acetic-treated mother, the nerve fiber layer showed widespread vacuoles infiltrated in between the ganglion cells which become less dense. There was a comparative reduction of inner and outer nuclear cell densities. The photoreceptor outer segments appeared fragile. Few numbers of the pigmented epithelium were observed (Fig. 2 A1 and A2).
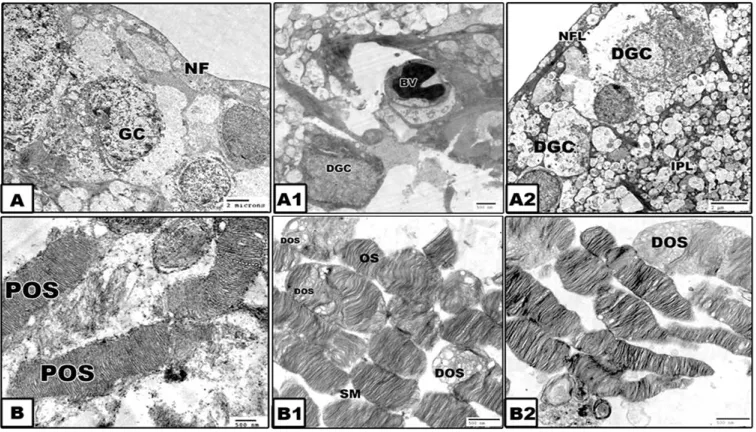
At the ultrastructural level, the control retina possessed widespread ganglion cell in-between myelinated nerve fibers. The inner & outer nuclear cells are regularly oriented. The outer segment of rods and cones appeared as a tubular structure arranged perpendicular to the pigmented epithelium and interdigitated with their microvillus system. The inner compart-ments of the outer segcompart-ments are composed of regularly oriented stacked membranes (Fig. 3A- B).
Fig. 1. Maternal retinal cell layers thickness (m) of GA3 and IAA-treatment comparing with control
Each result represents the mean±SD of 10 replicates. Significant at p ˂ 0.05 for only whole retina of experimental groups. WR, Whole retina; ONL, Outer nuclear layer; INL, Inner nuclear layer; OPL, Outer plexiform layer;
IPL, inner plexiform 0
20 40 60 80 100 120
WR ONL INL OPL IPL
µ
m
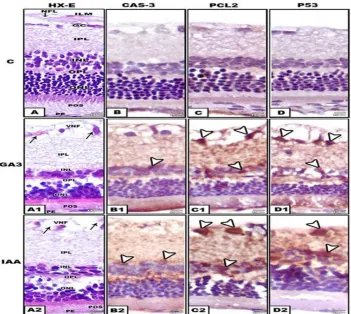
Fig. 2. Formalin-fixed histological and immunochemical staining of section of maternal retina. A-A2.HX-E. A. Control showing normal structure. A1. GA3-treatment. A2. IAA-treatment. Both
treatment comparative reduction of nuclear layers and degeneration of ganglion and pigmented cells. B-B2. Immunochemical staining with caspase -3 B. Negatively stained control. B1 and B2. GA3 and IAA . Arrow head showing immunochemical reaction in nuclear
layers. C-C2. Immunochemical staining with Pcl2 C. Negatively stained control. C1 and C2. GA3 and IAA . Arrow head showing immunochemical reaction in gaglion and nuclear layer.
D-D2. Immunochemical staining with P53. D. Negatively stained control. D1 and D-D2. GA3 and IAA. Arrow head showing immunochemical reaction in ganglion and nuclear layer Abbreviations; CC, Choricapillaries; GC, Ganglion cell; ILM, Inner limiting membrane; INL, inner nuclear layer;
IPL, Inner plexiform layer; NFL, Nerve fiber layer; ONL, Outer nuclear layer; OPL, Outer plexiform layer; PE, Pigmented epithelium; POS, Photoreceptor outer segment ; VNF, Vacuolated nerve fiber
In contrast, the experimental treated groups exhibited widespread degenerated ganglion cells and vacuolation of the nerve fiber layer. Numerous spots of necrotic stacked membranes of photoreceptor outer segments were detected (Figs. 3 A1-B1 and A2-B2).
3.4 Light and Transmission Electron Microscopy of Lens and Retina of Off Springs
3.4.1 Lens
Examining 20 histological eye specimens from the offsprings of each experimental group
Sayyad et al.; BJMMR, 10(12): 1-14, 2015; Article no.BJMMR.19964
of foamy cells invading the lens forming characteristic pattern of lenticular cataract. A case of post-capsular cataract was detected (Fig. 4 B-B3).
3.4.2 Retina
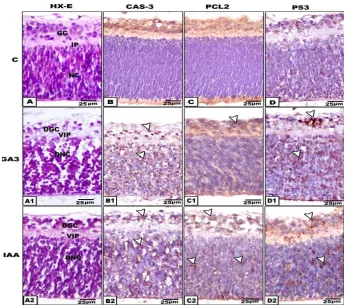
Light microscopic observations revealed that the control was composed of ganglion and nerve fiber, inner plexiform, undifferentiated nuclear layer and pigmented epithelium (Fig. 5A). However, in offsprings maternally-treated with the used plant hormones, the retinal cell layers lacked normal integrity and possessed comparative reduction of their cell density. There was apparent degeneration of the nuclear cells especially that of GA3-treated group. The ganglion, nerve fiber and inner plexiform layers appeared vacuolated (Fig. 5A1 and A2).
At the ultrastructural level, the control offsprings showed normal retinal cells, including ganglion, nuclear and photoreceptor's inner segments (Fig. 6A-A2). However, offsprings maternally treated with either GA3 or IAA showed deformation of photoreceptor inner segments. Many of the nuclear cells were degenerated and took abnormal fusiform structures with convolution of their nuclear envelope and electron-dense deposits of their nuclear chromatin (Pyknosis). The ganglion cells showed apparent necrosis and vacuolation of their surrounding nerve fibers. Offsprings maternally treated with IAA showed the highest degree of pathological alterations in comparison to the control (Fig. 6 B-B2 and C-C2).
3.5 Immunochemistry of Caspase-3, Bcl-2 and p53 of the Retina of Mother Rats and their off springs
Concerning mother rats, the retinal cells were partially stained with P53 for nuclei and Pcl2 and caspase-3 for cytoplasm especially in ganglion and nuclear cells of GA3 and IAA – treatment (Fig. 2 B1-B2, C1-C2,D1-D2) compared with negative staining in control (Fig.2 B-D). In offsprings, abundant immunoreactive P53, Pcl2 and caspase-3 were detected throughout ganglion and nuclear cells (Fig. 7 B1-B2, C1-C2, D1-D2) compared with negative staining of control (Fig. 5 B-D).
3.6 Genomic DNA Damage
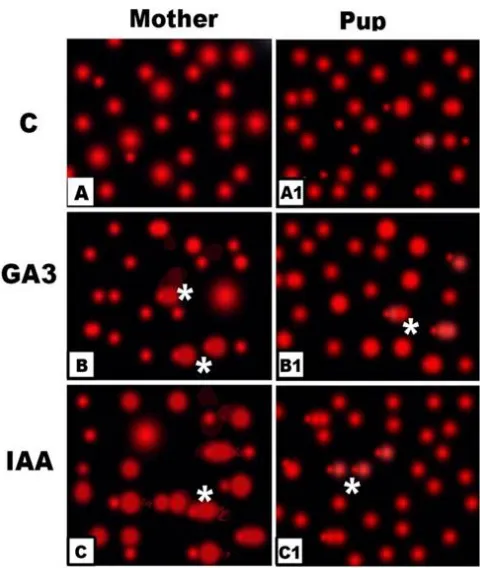
From Fig. 8, DNA fragmentation was markedly increased in retina of treated mothers and their offsprings. Also, the retinal cells of treated individuals revealed apparent detachment with marked increase of tail manifesting DNA (Fig.7).
4. DISCUSSION
From the present findings, maternal–treatments with the applied plant growth hormones led to the development of cataractous lenses in their offsprings with an average ratio of 6% in GA3 and 4% in IAA, being of the post-capsular and nuclear cataracts. These findings were first recorded and may lead to impairment of the visual function.
Table 1. Effect of gibberellic and indole acetic acid on antioxidants, lipid peroxidation and free radicals of eye of mothers and their offsprings
Mother Pups
C GA3 IAA C GA3 IAA
CAT (U/g tissue)
384.74±2.45 372.38±3.83* 368.22±3.21* 178.32±12.03 173.61±11.8** 159.32±11.9**
SOD (U/g tissue)
14.77±1.07 15.01±1.47** 9.83±0.64* 21.92±0.96 19.45±5.09** 13.53±3.26*
GR
(mg/g tissue)
1.11±0.68 0.46±0.02* 0.421±0.011* 4.17±1.50 2.48±0.20** 1.98±0.34*
GPx (U/g tissue)
39.49±1.19 21.54±1.07* 20.38±0.97* 25.49±4.38 15.44±2.37* 11.86±1.27*
MDA (n mol/g tissue)
9.89±0.74 14.8±0.96* 14.9±1.16* 6.62±0.84 15.01±3.5* 16.87±1.48
H2O2(m mol/g tissue)
124.28±2.31 231.72±2.1* 244.49±2.24 152.93±10.7 270.84±22.9* 283.5±21.96*
Fig. 3. Transmission electron micrographs of retina of mothers. A and B. Control showing normal retinal ganglion and photoreceptor outer segments with stacked membranes. A1 and
B1. GA3-treatment. A2 and B2. IAA-treatment. Both treatments showing degeneration and vacuolation of ganglion cells and degenerated patched of photoreceptor outer segments.
Lead citrate and uranyl acetate
Abbreviations; GC, Ganglion cell; DOS, Degenerated outer segment; NF, Nerve fiber; OS, Outer segment; POS, Photoreceptor outer segment
Fig. 4. Histological section of lens of offsprings. A. control lens showing normal arrangement of lens fibers. A1-A3. GA3-treatment. A1. Arrow head showing post-capsular cataract. A3. Arrow head showing nuclear swollen and degeneration of lens fibers. B-B3. IAA-treatment.
B-B1. Showing degeneration of lens fibers around the nuclear zone. B2. Showing marginal foam-like cells invading lens forming lenticular cataract. B3. arrow head pointing
post-capsular cataract
Abbreviations; FC, Foam cell; LF, Lens fiber; DLF, degenerated lens fiber; LC, Lenticular cataract; NC, Nuclear cataract. Hx-E.
Similar findings were reported in Long-Evans rats fetuses maternally treated with mirex (organochlorine insecticide) at doses of 6
Sayyad et al.; BJMMR, 10(12): 1-14, 2015; Article no.BJMMR.19964
IAA was found in mammalian tissues, including blood [31] and in several organs such as lung, kidney, liver and brain [32] as a result of consuming diet rich in vegetable stems or synthesized from tryptophan [33]. The accumulation of indole acetic acid was responsible for marked ultrastructural changes and death of cultured neutrophils [5] either by necrosis or apoptosis [34,35]. The hazardous effects may result from the production of reactive oxygen species [36]. Consequently, there was a close association between the increased tryptophan level and cataractous lens [37], indicating that the carcinogenic tryptophan pyrolysis products were present in human cataractous lenses. Kynurenic acid (tryptophan
metabolite) levels were found to be elevated in senile nuclear human cataracts and in cataractous lenses of rats with experimentally induced diabetes[38].
Also, both maternal administrations of either GA3 or IAA exerted interesting findings of retinal damage in the form of apparent cell death of ganglion and nuclear cells as well as, partial lysis of their inner stacked membranes of photoreceptor outer segment. Maternal retinal damage was reflected by the similar pattern of retinopathy of their offsprings. However, the photoreceptors of the offsprings were still premature and mainly formed inner segments which were disorganized.
Fig. 5. Formalin-fixed histological and immunochemical staining of section of offspring retina. A-A2.HX-E. A-A2. Hx-E. A. Control retina. C1&C2. Maternally treated with GA3 and IAA showing comparative reduction of ganglionic and nuclear cells. B-B2. Immunochemical staining with caspase -3 B. Negatively stained control. B1 and B2. GA3 and IAA. Arrow head showing immunochemical reaction. C-C2. Immunochemical staining with Pcl2 C. Negatively stained control. C1 and C2. GA3 and IAA . Arrow head showing immunochemical reaction in gaglion and nuclear layer. D-D2. Immunochemical staining with P53. D. Negatively stained control. D1 and D2. GA3 and IAA. Arrow head showing immunochemical reaction in ganglion
and nuclear layer
Similar findings were also reported among farmers applying pesticides in the Agricultural Health Study from Iowa and North Carolina post fungicides use (maneb or mancozeb and ziram) [39].
The observed histopathological and ultrastructural findings were confirmed by the applied immunochemical staining of P53, Pcl2 and caspase-3, which revealed abundant immunoreactive staining in ganglion and nuclear cells manifesting the presence of apoptic cell death. Apoptosis can mediate pro-apoptotic or anti-apoptotic members to release Cytochrome c
into the cytosol which, activates caspase-3, leading to apoptosis. Bcl-2 is essential in the process of apoptosis because it suppresses the initiation of the cell-death process by inhibiting mitochondrial permeability [40].
The observed retinopathy in treated offsprings may be attributed to the transplacental passage of the used plant hormones or their metabolites and interference with histogenesis and differentiation of retinal cells. Troudi et al.[16] detected neuronal cell damage in rat offsprings maternally given 200 ppm gibberellic acid in drinking water daily from the 14th day of pregnancy until Day 14 post partum. According to Furukawa et al. [4] and Yilmaz and Celik [41], the central nervous system is more susceptible to oxidative damage of PGRs due to its high oxygen consumption rate and relatively low levels of defense mechanisms against toxicity. High levels of polyunsaturated fatty acids were found to facilitate oxidative damage[42].
Fig. 6. Transmission electron micrographs of retina of offsprings. A-A2. Control showing normal ganglion, nuclear cells and photoreceptor inner segments. B-B2. Gibberellic
acid-treatment. C-C2. Indole acetic acid. A-C. Ganglion cells. A1-C1. Nuclear cells. A2-C2. Photoreceptor inner segments. Both offsprings maternally treated showing pyknotic cell death of ganglionic and nuclear cells degeneration of ganglion cells and degenerated patched
Sayyad et al.; BJMMR, 10(12): 1-14, 2015; Article no.BJMMR.19964
Fig. 7. Comet photomicrographs of retinal cells of GA3 and IAA- treated mothers and their offsprings showing increased increased stretched cells manifested DNA damage (*). A. Control mother. B. GA3-treated mother. C. IAA-treated mother. A1.Control offspring. B1.
Offsprings maternally treated with GA3. C2. Offspring maternally treated with IAA
Fig. 8. Genomic DNA fragmentation of retina of gibberellic and indoleacetic acid -treated mothers and their offsprings showing increased DNA band fragmentation
The observed retinopathy was confirmed by the detected DNA fragmentation and marked increase of malondialdehydes and free radicals. Similar findings were reported in the retina of mice after treatment with organophosphorus chloropyrifos [43].
In addition, there was a detected depletion of the retinal antioxidant enzyme's SOD, CAT and GR and GPx in experimentally treated mothers and their offsprings.
the antioxidant enzymes system[48] which led to the impairment of homeostasis and reduce cellular defense system against toxicity originated by the liberation of active oxygen forms [49].
5. CONCLUSION
Finally, it can be concluded that the plant growth hormones decreased the endogenous antioxidant system and increased the malonadialdhydes and hydrogen peroxides which were assessed by cytological alterations of retinal cell layers and DNA damage.
ACKNOWLEDGEMENT
The authors would like to thank the Institiute of Scientific Research and Revival of Islamic Heritage, Um Al-Qura University, Makkah, kingdom Saudia Arabia for their financial support.
COMPETING INTERESTS
Authors have declared that no competing interests exist.
REFERENCES
1. Friedman DS, Repka MX, Katz J, Giordano L, Ibironke J, Hawes P, Burkom D, James M, Tielsch JM. Prevalence of decreased visual acuity among preschool-aged children in an American Urban population. Ophthalmology. 2008;115:1786–1795. 2. Serrani JC, Fos M, Atarés A,
García-Martínez JL. Effect of gibberellin and auxin on parthenocarpic fruit growth induction in the cv Micro-Tom of tomato. J. Plant Growth Regul. 2007;26:211-221.
3. Mickel LG. ‘‘Plant Growth Regulators’’ Controlling biological behavior with chemicals. Chem. Eng. News. 1978;56:18. 4. El-Mofty MM, Sakr SA. Induction of
neoplasms in the Egyptian Toad by gibberellin A3. Oncology. 1983;45:61– 64. 5. De Melo MP, De Lima TM, Pithon-Curi TC,
Curi R. The mechanism of indole acetic acid cytotoxicity. Toxicol. 2004;148:103-111.
6. Furukawa S, Abe M, Usuda K, Ogawa I. Indole-3-acetic acid induces microence-phaly in rat fetuses. Toxicol Pathol. 2004; 326:659-67.
7. Hsiao G, Shen MY, Lin KH, Chou CY, Tzu NH, Lin CH, Chou DS, Chen TF, Sheu JR.
Inhibitory activity of kinetin on free radical formation of activated platelets in vitro and on thrombus formation in vivo. Eur J Pharmacol. 2003;465:281-87.
8. Arous S, Boussaid M, Marrakchi M. Plant regeneration from zygotic embryo hypocotyls of Tunisian chilli Capsicum annuum L. J Appl. Hortic. 2001;3:17–22. 9. Chaari-Rkhis A, Maalej M. Ouled
Messaoud S, Drira N. Invitro vegetative growth and flowering of olive tree in response to GA3 treatment. Afr J Biotechnol. 2006;5:2097–2302.
10. Celik I, Ozbe KH, Tuluce Y. Effects of sub chronic treatment of some plant growth regulators on serum enzyme levels in rats. Trends J. Biol. 2002;26:73–76.
11. Berhow MA, Vandercook CE. The reduction of naringin content of grapefruit by applications of gibberellic acid. Plant Grow Reg. 1992;11:75-80.
12. Broughton WJ, Guat T. Storage conditions and ripening of the custard apple Annona squamosa L. Scientia Horticulturae. 1979; 10:73-82.
13. Ustun H, Tecimer T, Ozmen M, Topcuoglu SF, Bozcuk NA. Effects of gibberellic acid and benzoprenin on mice. Histopatalogic review, Ank. Patol. Bult. 1992;9:36–40. 14. Celik, I. and Kara, M. The effects of some
plant growth regulators on activity of eight serum enzymes in vitro. J Environ Sci. Health A. 1997;32:1755–1761.
15. Troudi A, Samet MA, Zeghal N. Hepatotoxicity induced by gibberellic acid in adult rats and their progeny. Exp. Toxicol.Pathol. 2010;62:637-642.
16. Troudi A, Bouaziz H, Soudani N, Ben Amara I, Boudawara T, Touzani H, Lyoussi B, Zeghal N. Neurotoxicity and oxidative stress induced by gibberellic acid in rats during late pregnancy and early postnatal periods: Biochemical and histological changes. Exp Toxicol Pathol. 2012;64: 583-90.
17. Furukawa S, Usuda K, Abe M, Hayashi S, Ogawa I. Indole-3-acetic acid induces microencephaly in mouse fetuses. Exp Toxicol Pathol. 2007;59:43-52.
18. Jaga K, Dharmani C. Ocular toxicity from pesticide exposure: A recent review. Environ Health Prev Med. 2006;11:102-7. 19. Good PA, Claxson A, Morris CJ, Blake DR.
Sayyad et al.; BJMMR, 10(12): 1-14, 2015; Article no.BJMMR.19964
20. Morrison JC, Moore CG, Deppmeier LM, Gold BG, Meshul CK, Johnson EC. A rat model of chronic pressure-induced optic nerve damage. Exp Eye Res. 1997;64:85-96.
21. Gatzioufas Z, Charalambous P, Seitz B, Evers S, Jonescu-Cuypers C, Krause M, Thanos S. Cholinergic inhibition by botulinum toxin in a rat model of congenital glaucoma raises intraocular pressure. Br J Ophthalmol. 2008;92:826-831.
22. Prins HK, Loose JA. Glutathione “chapter 4” in biochemical methods in red cell genetics. Edited by J.J. Yunis. Academic press, N.Y.D. London. 1969;126-129. 23. Paglia DE, Valetine WN. Determination of
glutathione peroxidase activity. J. Lab. Clin. Med. 1967;158–169.
24. Bock PP, Kramer R, Pavelka M. Peroxisomes and related particles. In: Cell Biology Monographs 7, Springer, Berlin. 1980;44-74.
25. Niskikimi M, Rao NA, Yagii K. The occurrence of superoxide anion in the reaction of reduced phenazine metho-sulfate and molecular oxygen. Biochem. Biophys. Res. Comm. 1972;46:849–854. 26. Ohkawa H, Ohishi N, Yagi K. Assay for
lipid peroxides in animal tissues by thiobar-bituric acid reaction. Anal. Biochem. 1979; 95:351–358.
27. Fossati P, Prencipe L, Berti G. Use of 3,5-dichloro-2-hydroxybenzenesulfonic acid/4-aminophenazone chromogenic system in direct enzymic assay of uric acid in serum and urine. Clin. Chem. 1980;227-231. 28. Sasaki YF, Nishidate E, Izumiyama F,
Matsusaka N, Tsuda S. Simple detection of chemical mutagens by the alkaline single-cell gel electrophoresis Comet assay in multiple mouse organs. Mutat. Res. 1997;391:215-231.
29. Duke RC, Sellins CB. In: Kaplan JG, ed. Cellular Basis of Immune Modulation. New York: Liss. 1989;311–314.
30. Rogers JM, Grabowski CT. Mirex-induced fetal cataracts: lens growth, histology and cation balance, and relationship to edema. Teratology. 1983;273:343-9.
31. Hu T, Dryhurst G. Electrochemical and peroxidase O2-mediated oxidation of
indole-3-acetic acid at physiological pH. Electroanal Chem. 1997;432:7–18.
32. Martínez E, Artigas F, Suñol C, Tusell JM, Gelpí E. Liquid-chromatographic deter-mination of indole-3-acetic acid and 5-
hydroxyindole-3-acetic acid in human plasma. Clin. Chem. 1983;29:1354–57. 33. Tusell JM, Artigas F, Sunol C, Martinez E ,
Gelpi E. Comparison of high-performance liquid chromatography and gas chromate-graphy-mass spectrometry for the analysis of indole-3-acetic acid in brain tissue. J Chromatogr. 1984;306:338-4.
34. Mills MH, Finlay DC, Haddad PR. Determination of melatonin and monoa-mines in rat pineal using reversed-phase ion-interaction chromatography with fluore-scence detection. J. Chromatogr. Biomed. 1991;564:93–102.
35. De Melo MP, Pithon-Curi TC, Miyasaka CK, Palanch AC, Curi R. Effect of indole acetic acid on oxygen metabolism in cultured rat neutrophils. Gen. Pharmacol. 1998;31:573–578.
36. Kroemer G. The mitochondrial permeability transition pore complex as a pharma-cological target. An introduction. Curr. Med. Chem. 2003;10:1469–1472.
37. Proskuryakov SY, Konoplyannikov AG, Gabi VL. Necrosis: A specific form of programmed cell death?. Exp. Cell. Res. 2003;283:1–16.
38. Manabe S, Wada O. Carcinogenic tryptophan pyrolysis products in human lens. Exp Eye Res. 1989;48:351-63. 39. Zarnowski T, Rejdak R, Rummelt C,
Zrenner E, Grieb P, Okuno E, Schlötzer-Schrehardt U, Naumann GO, Kruse F , Jünemann AG. Immunohistochemical identification of kynurenine aminotrans-ferases in corpora amylacea in the human retina and optic nerve. Folia. Neuropathol. 2007;452:66-71.
40. Kirrane EF, Hoppin JA, Kamel F, David M, Umbach Boyes WK, DeRoos AJ, Alavanja M, Sandler DP. Retinal degeneration and other eye disorders in wives of farmer pesticide applicators enrolled in the agricultural health study. Am. J. Epidemiol. 2005;161:1020–1029.
41. Yilmaz Z Celik I. Neurotoxic and immuno toxic effects of indole-3-butyricacid on rats at subacute and subchronic exposure. Neurotoxicology. 2009;30:382–5.
42. Samson FE, Nelson SR. The aging brain, metals and oxygen free radicals. Cell. Mol. Biol. 2000;46:699–707.
combination of vitamins C and E Exp. Toxicol. Pathol. 2008;596:415-423. 44. Celik I, Tuluce Y. Effects of indole acetic
acid and kinetin on lipid peroxidation and antioxidant defense in various tissues of rats. Pest Biochem. Physiol. 2006;84:49– 54.
45. Tuluce Y, Celik I. Effects of indoleacetic acid and kinetin on lipid peroxidation and antioxidant defense in various tissues of rats. Pesticide Bioch. Physiol. 2006;84, 49–54.
46. Celik I, Turker M, Tuluce Y. Absisic acid and gibberellic acid cause increased lipid peroxidation and fluctuated antioxidant defense systems of various tissues in rats. J. Hazard Mater. 2007;148:623-629.
47. Regoli F, Principato G. Glutathione, glutathione dependent and antioxidant enzymes in mussel, Mytilus gallopro-vincialis, exposed to metals under field and laboratory conditions: Implications for the use of biochemical biomarkers. Aquat. Toxicol. 1995;31:143–164.
48. Halliwell B, Gutteridge JMC. Free radicals in biology and medicine, second reprint. Clarendon Press, Oxford. 1989;543. 49. Oruc-Ozcan E, Uner N. Marker enzyme
assessment in the liver of Cyprinus carpio L. exposed to 2, 4-D and azinphosmethyl. J. Biochem. Mol. Toxicol. 2002;16: 183-188.
© 2015 Sayyad et al.; This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Peer-review history: