Genomic integration of Wnt/β catenin and BMP/Smad1 signaling coordinates foregut and hindgut transcriptional programs
Full text
Figure
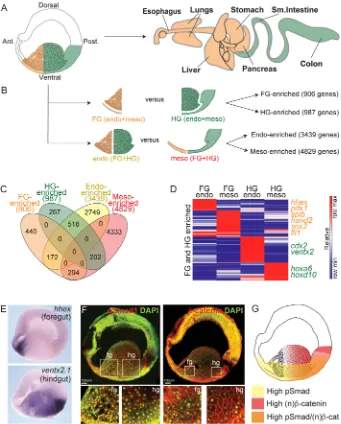
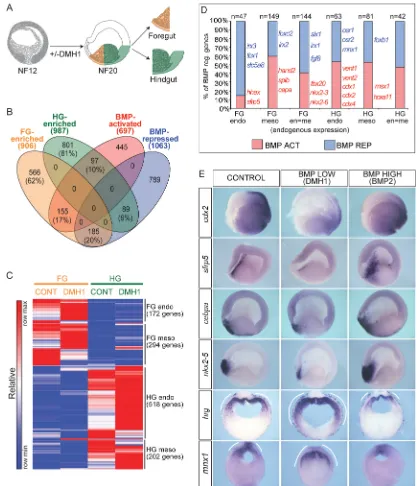
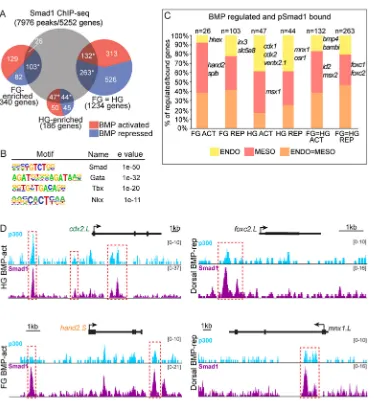
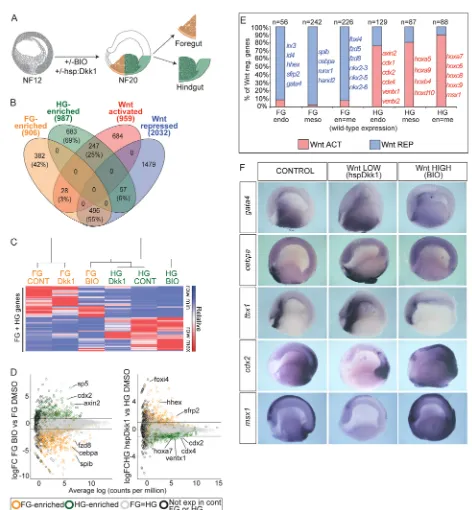
Related documents
The interaction between steel tube and concrete core is the discussing issue for understanding the behavior of concrete- filled steel tube columns (CFST).The
C lassical galactosemia (McKusick 230400) is an autosomal recessive disorder of galactose me- tabolism caused by a deficiency of galactose- 1-phosphate uridyltransferase (GALT;
Dynamics of total soils contamination by metals AGC sanitary protection zone, 1997−2012: Zc(MnCrVNiCoCuAgZnPbSnMo). Since then, this kind of diagnostics with detailed
According to Table 9 is possible to load all the required parameters for the key binding use case is 283.14 ms (one issuer public key, a CL tuple and the system parameters needed
Under the horizontal smoothing number of 6, 3 and 1, the correlation coefficients between the BLH derived by graphics distribution method (GDM) algorithm and ground-based lidar
[Resistance Pattern of Enterococci Isolated from Nosocomial Infections in the Hospitals Located in Gonbad and Gorgan Cities, Iran]. 2014; 8(Suppl
when the menB , hemB , or cydB qoxB mutant strains were grown in similar conditions, indicating that these strains are significantly impaired for the ability to generate a
At 35 dpf, only three assemblages exhibited significantly different relative abundances in HF versus LF environmental microbiotas from tanks with fish, and all were enriched in the
