RESEARCH ARTICLE
A dynamic regulatory network explains ParaHox gene control of
gut patterning in the sea urchin
Rossella Annunziata* and Maria Ina Arnone‡
ABSTRACT
The anteroposterior patterning of the embryonic gut represents one of the most intriguing biological processes in development. A dynamic control of gene transcription regulation and cell movement is perfectly orchestrated to shape a functional gut in distinct specialized parts. Two ParaHox genes,XloxandCdx, play key roles in vertebrate and sea urchin gut patterning through molecular mechanisms that are still mostly unclear. Here, we have combined functional analysis methodologies with high-resolution imaging and RNA-seq to investigateXloxandCdx regulation and function. We reveal part of the regulatory machinery responsible for the onset of Xloxand Cdxtranscription, uncover a Wnt10 signal that mediatesXloxrepression in the intestinal cells, and provide evidence ofXlox- andCdx-mediated control of stomach and intestine differentiation, respectively. Our findings offer a novel mechanistic explanation of how the control of transcription is linked to cell differentiation and morphogenesis for the development of a perfectly organized biological system such as the sea urchin larval gut.
KEY WORDS: Stomach, Intestine, Gene regulatory network
INTRODUCTION
Epithelial cell sheet formation has been a crucial step in the evolution of multicellularity. In particular, the development of an internalized gastrointestinal system contributed to the increase of animal morphological complexity conferring the capacity to perform extracellular digestion and releasing organisms from body size constrains, which very likely allowed the development of specialized structures. Accordingly, the molecular system orchestrating digestive tube development can arguably be considered to be one of the first evolved developmental patterning systems and may involve genes that were later co-opted for the development and patterning of new body parts (Nielsen, 2008; Roberts, 2000; Wolpert, 1994), thus providing a model with which to investigate widely employed developmental processes such as differentiation, morphogenesis and axis formation. The embryonic gut of most deuterostomes forms through common developmental processes during gastrulation: endodermal cells are internalized by invagination through the blastopore of sea urchin and frog embryos and, in a very similar way, ingress with combined involution and migration movements through the primitive streak of the avian and mammalian epiblast. In all cases, the first endodermal cells to exit either the blastopore or the primitive streak will contribute to the anterior definitive endoderm, whereas the later moving cells will contribute to the posterior endoderm. After
gastrulation, the primitive gut of most animals seems a tube with no apparent differentiation at the morphological level. However, several distinct or partially overlapping transcription factor and signaling molecule expression territories can be detected along the whole length of the archenteron (Jacobs et al., 2012; Lengyel and Iwaki, 2002; McGhee, 2007; Sherwood et al., 2011; van den Brink, 2007; Zorn and Wells, 2009). The interaction between molecular regulators leads, eventually, to the establishment of fine boundaries of gene expression subdividing the gut into distinct domains, precursors of the different digestive organs.
Two ParaHox transcription factors, Xlox and Cdx, have been proposed as central actors in the gut development in several animals (Brooke et al., 1998; Holland, 2013). Cdx is required for posterior gut development in many cases examined, and Xlox is crucial for more anterior gut domains and pancreas formation. In addition, a number of signaling molecules have been shown to regulate different aspects of vertebrate gut formation (Feng et al., 2012; Jacobs et al., 2012; Spence et al., 2011). However, characterizing the molecular interactions responsible for gut patterning has proved to be a difficult task in vertebrate models such as mouse or chick, in which the generation of transgenic lines is often required and tandem genome duplications occurred (Donoghue and Purnell, 2005).
Most sea urchins are indirect developers and go through a bilateral swimming (and feeding) pelagic larval stage (the pluteus), before metamorphosis leads to the formation of the adult form with its characteristic pentameric body plan. The pluteus has a functional tripartite gut composed of a muscular esophagus, a large spherical stomach and a small tubular intestine (Burke, 1981). A cardiac sphincter between the esophagus and the stomach, and a pyloric sphincter between the stomach and the intestine separate the gut compartments. The small number of cells and the detailed description of cell movements during gastrulation (McClay, 2011), the extensive molecular understanding of the early endoderm specification mechanisms (Croce and McClay, 2010; Davidson et al., 2002; Peter and Davidson, 2010) and the reduced number of gene duplications compared with vertebrates, all make the sea urchin embryo an excellent but so far largely unexploited model to investigate the molecular dynamics of gut development.
ParaHox gene involvement in the sea urchin gut patterning has been already demonstrated (Arnone et al., 2006; Cole et al., 2009). Although Xlox is required for the formation of the pyloric sphincter, Cdx is dedicated to setting up the posterior border of Xlox expression within the intestine.
In this study, we combine the use of functional analysis methodologies with high resolution imaging, RNAseq and qPCR to show that sea urchin larval gut patterning is achieved through the integration of autonomous and conditional mechanisms of cell fate specification. First, we demonstrate that the early active endodermal transcription factors Hox11/13b, FoxA, Bra and Blimp1a (Arenas-Mena et al., 2006; Livi and Davidson, 2006; Oliveri et al., 2006; Rast et al., 2002) synergistically regulate ParaHox gene expression. Then, Received 6 November 2013; Accepted 14 April 2014
Stazione Zoologica Anton Dohrn, Cellular and Developmental Biology, Villa Comunale, Napoli 80121, Italy.
*Present address: UMR 7238 CNRS-UPMC, Les Cordeliers 15, Rue de l’Ecole de Médecine, Paris 75006, France.
‡
Author for correspondence ([email protected])
DEVEL
O
we uncover a Wnt10 signaling event mediating Cdx-controlled repression ofXloxtranscription in posterior gut cells, and provide evidence of a positive auto-regulatory input of Cdx on its own transcription. Moreover, we show that Xlox is required for the activation of stomach terminal differentiation genes and also for the transcription of the muscle-specific myosin heavy chain gene in the pyloric sphincter cells. Finally, we identify a number of additional developmental processes potentially controlled by Xlox and we do that through a genome-wide comparative transcriptome analysis on Xlox morphant gastrulae and plutei. From all the above studies, Xlox clearly emerges as a crucial player of cell differentiation in the sea urchin embryo, whereas Cdx acts as major regulator of intestinal cell identity.
To our knowledge this is the first study, in a non-chordate deuterostome (with the experimental advantages that this implies), to have investigated the gut patterning process at the functional level and offers a great contribution to our general understanding of the molecular mechanisms involved in the development of a complex and highly organized structure such as the embryonic digestive apparatus.
RESULTS
Dynamic control of gene transcription in the developing sea urchin gut
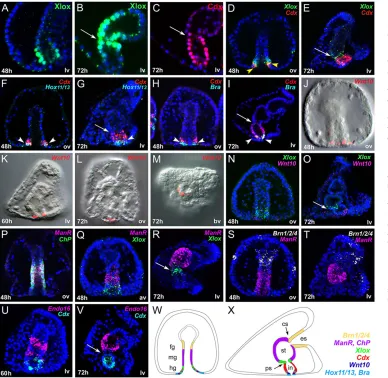
The first step in the study of the functional regulation of cell differentiation is the detailed analysis in time and space of the actively transcribed factors putatively involved in the process. Since late gastrula stage, the archenteron of the embryo can be subdivided in three regions, foregut, midgut and hindgut that will, respectively, form esophagus, stomach and intestine of the pluteus larva. We analyzed Xlox and Cdx protein localization and gut gene expression relative to Xlox and Cdx transcript localization, from the post gastrular embryo to the pluteus larval stages, including active transcription factors and terminal differentiation genes (Fig. 1).
The expression dynamics of SpLox and SpCdx proteins have been analyzed by immunolocalization, using specific antibodies developed in our laboratory, and the protein versus mRNA spatial localization in the embryos and larvae have been compared. Xlox protein localization at late gastrula stage matched the transcript pattern, being detected in around 30 cells of the posterior gut (Fig. 1A,D,N,Q). However, whereasXloxtranscripts are strictly confined to around 10 cells of the larval pyloric sphincter, we detected the protein in about 30 cells covering the pyloric sphincter and part of the stomach posterior hemisphere, mostly on the aboral side (Fig. 1B,E,O,R). We detected Xlox protein in the stomach domain until the 3-week larval stage. We propose that, late in gastrulation, the most anterior cells expressing XloxmRNA stop transcribing the gene but retain the protein and develop to form part of the larval stomach. Cdx protein localization exactly matched the corresponding intestinal mRNA expression at all stages (Fig. 1C-I,U,V).
We then analyzedCdxmRNA spatial expression relative toXlox, Hox11/13bandBraat both late gastrula (48 h) and pluteus stages. Experiments at earlier gastrula stage (40-44 h), demonstrate that the first cells expressingCdxalso express Xlox,Hox11/13band Bra (data not shown), thus suggesting thatCdx,Xlox,Hox11/13band Bratogether define a specific cellular regulatory state, exclusive to this most posterior part of the late gastrula archenteron, and hinting to a possible regulatory interaction among them. Moreover, considering the fact thatXlox,Hox11/13b andBra expression in those cells starts beforeCdxtranscription activation (Arenas-Mena et al., 2006; Arnone et al., 2006; Rast et al., 2002), we contemplated and investigated a possible role forXlox,Hox11/13bandBrainCdx
regulation. At the late gastrula stage, the domain ofCdxexpression expands, overlapping anteriorly only withXloxand posteriorly only withHox11/13band Bra(Fig. 1D-I). The expression domains of Cdx, Xlox, Hox11/13b and Bra in the pluteus show a different relative pattern reflecting the significant changes occurred in the morphology of the gut during the transition from late gastrula to pluteus stages:Cdxis now expressed in the whole intestine,Xloxis transcribed only in the pyloric sphincter cells, andHox11/13band Brain the most posterior cells of the intestine. Although no sign of co-expression is observable at pluteus stage betweenXloxandCdx (Fig. 1E), a partial overlapping betweenCdxandHox11/13b, and betweenCdxandBrais still detectable in the posterior cells of the gut (see arrowheads in Fig. 1G,I).
A very remarkable discovery that we present in this work is the finding of a Wnt10 signal in the posterior gut cells of the embryo. We found thatWnt10transcription starts a few hours after the activation of Cdx, and is localized in the posterior cells of the gut at late gastrula (Fig. 1J), prism (Fig. 1K) and pluteus stages (Fig. 1L,M). The cells expressingWnt10apparently represent a subset of the posterior-most Cdxpositive cells. Interestingly, from the moment these cells start expressing Wnt10, Xlox transcripts become undetectable in them (Fig. 1N) and when the gut attains its full length, all the intestinal cells are depleted fromXloxtranscripts (Fig. 1O).
Finally, we found that the stomach-specificManrC1AandChP genes are already co-expressed at mid gastrula stage in the cells that will give rise to the larval stomach (Fig. 1P) and are never expressed inXlox- (Fig. 1Q,R),Cdx- (data not shown) orBrn1/2/4-positive cells (Fig. 1S,T). A third stomach marker,Endo-16, transcribed in both midgut and hindgut cells at late gastrula (Ransick et al., 1993), is progressively cleared from the intestinal cells in parallel with the activation of Cdx, and becomes confined to the stomach larval cells. No signs of co-expression ofEndo-16andCdxhave been detected either at prism or at pluteus stages (Fig. 1U,V).
XloxandCdxtranscription regulation in the developing gut After the high resolution gene expression analysis, we investigated the regulatory mechanisms of Xlox and Cdx transcription. We perturbed protein translation by injecting morpholino antisense oligonucleotides (MOs), then analyzedXloxandCdxmRNAs and, in some cases, proteins, in the 72 h morphant larvae.
The results of the gene perturbation experiments are depicted in Fig. 2, and supplementary material Figs S1 and S2. A description of the postulated gene regulatory dynamics is provided below. In control larvae,Cdxtranscripts are localized in the intestinal cells, whereasXloxtranscripts are detectable in the cells of the pyloric sphincter. Interestingly, Cdx transcripts were undetectable in the intestinal cells of Hox11/13b, Bra, FoxA, Cdx and Blimp-1a morphant larvae (Fig. 2A-F), suggesting that they are involved in Cdx regulation. Crucially, in Hox11/13b, Bra, FoxA and Cdx knockdown larvae (in whichCdxexpression is lost),Xloxtranscripts accumulated ectopically in the intestinal cells (Fig. 2G-L), providing additional support to Cdx repressive role onXloxtranscription (Cole et al., 2009). Moreover, Xlox transcripts were not detectable in Blimp-1a morphants (Fig. 2L), suggesting that Blimp-1a might be required forXloxtranscription, which also explained the absence of Cdxfrom Blimp-1a morphants, asCdxtranscription requires Xlox input (Cole et al., 2009). However,Blimp-1atranscription is active in a broader domain compared withXlox(Livi and Davidson, 2006), suggesting that other unknown factors are expressed in prospective Xlox-positive cells and recruited for its activation.
An intriguing finding has been the impairment of Cdx transcription when its own protein formation was blocked
DEVEL
O
(Fig. 2E), with the consequent accumulation ofXloxtranscripts in the posterior larval gut cells (Fig. 2K). We suggest that Hox11/13b, Bra and FoxA, together with Xlox, all cooperate for the activation of Cdx in the subset of gut cells destined to form the intestine. Subsequently, because in the larval intestine only some Cdx -positive cells still express the factors responsible for its initial activation (see doublein situhybridization in Fig. 1), Cdx becomes recruited for its own transcription,‘locking-in’the regulatory state of intestinal cells.
The injection of morpholinos blocking the translation of GataE and TgiF (Howard-Ashby et al., 2006; Lee and Davidson, 2004) did not produce any change in theXloxandCdxtranscript distribution (supplementary material Fig. S1), allowing us to exclude their involvement in Xlox and Cdx regulation. Most of the results presented in this section and the demonstration of morpholino functionality have been confirmed through the use of anti- Lox, Cdx and Bra antibodies (supplementary material Fig. S2).
Taking all the data into account, we propose a model that places Hox11/13b, Bra, FoxA and Xlox together as members of the regulatory machinery activatingCdxtranscription. As expression of the first three genes starts many hours before Cdx activation, we hypothesize that Xlox functions as a switch controlling the exact time at whichCdxtranscription is initiated. Finally, Blimp1a results as one of the activators ofXloxtranscription: its expression starts many hours before the beginning of gastrulation, thus suggesting the presence of additional factors involved inXloxtranscription activation.
A Wnt10 signaling mediates Cdx repressive action onXloxin intestinal cells
[image:3.612.50.438.58.436.2]As described above, we found thatCdxactivation requires Hox11/13b, FoxA, Bra and Xlox, and that after the translation of Cdx protein,Xlox transcription stops in the developing intestine. In order to explore the possible involvement of intercellular signaling in a Xlox-Cdx positive-negative feedback loop, we screened by qPCR (see Materials and methods for details) a variety of signaling molecules expressed during sea urchin embryonic development. The screening revealed that SpWnt10 transcript number was strongly reduced in Cdx knockdown larvae, persuading us of its involvement in Cdx-regulated processes. We then tested Wnt10 function through the embryonic injection of a morpholino designed to block its translation. Wnt10 knockdown larvae presented an almost normal gut characterized by a significant reduction of the pyloric sphincter constriction when compared with control larvae (for the phenotype of the morphants, see the full projection of confocal z-series in Fig. 3). Interestingly, Xlox and Cdx doublein situ hybridization revealed that the two genes were co-expressed in the intestinal cells of Wnt10 knockdown larvae. Immunolocalization experiments confirmed that Xlox protein ectopically accumulates in Wnt10 knockdown larval intestines (supplementary material Fig. S2K). These results significantly contributed to the understanding ofXlox and Cdx regulation, indicating that Cdx-negative control of Xlox transcription is indirect and occurs via a Wnt10 signaling event (Fig. 3A-D). Very importantly,Wnt10transcripts were not Fig. 1. Dynamics of gene expression in the developing sea urchin gut. (A-C) Xlox and Cdx protein accumulation visualized by immunostaining. (D-V) mRNA localization determined by two-color fluorescentin situhybridization. Nuclei are stained with DAPI and depicted in blue. The antibodies and probes used, the age and the orientation of the embryos are reported in each panel: h, hours post fertilization; ov, view from the oral side; lv, lateral view; bv, view from the blastopore. Overlapping domains of expression are highlighted by arrowheads. Arrows indicate the position of the pyloric sphincter in the pluteus larvae. Full projections of confocalz-series (E,N,O,Q-V) or single slices (A-D,F-M,P) are shown. (W,X) Schematic of a late gastrula in frontal view (W) and a pluteus larva in lateral view (X): endodermal domains are depicted with different colors according to gene expression. Overlapping domains of expression are represented with colored lines. ManrC1Aname has been shortened to ManRfor clarity. fg, foregut; mg, midgut; hg, hindgut; es, esophagus; st, stomach; in, intestine; ps, pyloric sphincter; cs, cardiac sphincter.
DEVEL
O
detectable in Cdx knockdown larvae by in situ hybridization (Fig. 3E,F), confirming the role of Cdx in the activation of Wnt10 transcription. We propose that Cdx participates in Wnt10 transcription in the posterior cells of the intestine and that Wnt10 ligands diffuse towards the anterior side of the gut, clearing the intestinal cells ofXloxtranscripts through the activation of an effector molecule with repressive functions.
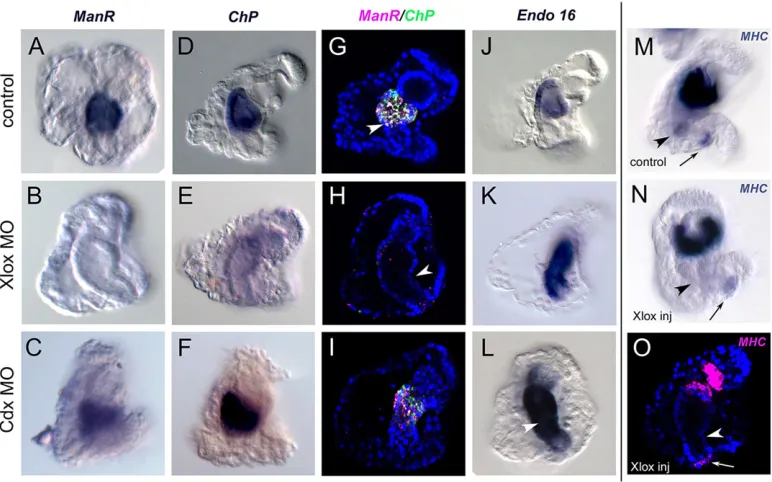
Xlox and Cdx control the AP patterning of the larval gut Digestive functions in the stomach are inhibited in Xlox knockdown larvae (Cole et al., 2009). We explored the expression of stomach terminal differentiation genes, ManrC1A, ChP and Endo-16, in Xlox and Cdx knockdown larvae. The expression of all three genes in Xlox morphants was severely affected (Fig. 4). In particular,ChP and ManrC1A transcription was dramatically reduced, to the point that no transcripts could be detected by either colorimetric (Fig. 4A,B,D,E) or fluorescentin situhybridization (Fig. 4G-H). Xlox is thus upstream of a cascade of regulatory events responsible for the differentiation of the larval stomach. However, the localization of XloxmRNA and protein relative toChPandManrC1Atranscripts during gut development (see Fig. 1) advocates the involvement of a signaling event in the induction of stomach cell differentiation. Although Cdx knockdown did not have any noticeable effect onChP
andManrC1Atranscription (Fig. 4C,F,I),Endo-16ectopic expression was observed in the intestine of both Xlox and Cdx morphants (Fig. 4J-L). We propose that Cdx is responsible for Endo-16 repression from the intestinal cells, acting either directly on its transcription or through the repression in the intestine of theEndo-16 midgut-hindgut activators, and that Xlox executes a repressive function onEndo16transcription (Cole et al., 2009) in the cells where Cdx is never active and that will eventually form the pyloric sphincter.
Xlox: a crucial player of cell differentiation in the sea urchin embryo
[image:4.612.67.546.55.221.2] [image:4.612.49.355.535.737.2]The most obvious morphological defect in Xlox knockdown larvae is the absence of the pyloric sphincter (e.g. compare confocal images in Fig. 4G,H), the cells of which express the muscle-specific terminal differentiation gene,Myosin heavy chain(SpMHC) (Andrikou et al., 2013; Venuti et al., 1993). In control larvae,MHCis expressed in the four muscular structures of the gut, the esophageal muscles and the cardiac, pyloric and anal sphincters. The esophageal muscles are of mesodermal origin whereas sphincter muscles are endodermal (Gustafson and Wolpert, 1967), suggesting the existence of different regulatory mechanisms involved in the acquisition of muscle cell identity in each type. We found thatMHCtranscripts were specifically absent from the cells forming the pyloric sphincter in Xlox morphants Fig. 2.XloxandCdxtranscriptional regulators.(A-L)XloxandCdxmRNA localization determined by chromogenicin situhybridization in control embryos (A,G) and in embryos injected with MOs directed against the translation of Hox11/13b (B,H), Bra (C,I), FoxA (D,J), Cdx (E,K) and Blimp1b (F,L) RNAs. All the embryos were fixed at 72 h. The orientation of the larvae is reported for each panel: ov, view from the oral side; av, view from the aboral side; lv, lateral view.
Fig. 3. A Wnt10 signaling event mediates Cdx repressive action onXloxin the presumptive intestine.(A-D) Two-color fluorescentin situhybridization coupled with nuclear staining (blue). In E and F, single-color fluorescentin situ hybridization (magenta) and larval cilia staining (green, acetylated tubulin immunostaining). In A-C, a single slice of a confocal stack showsXloxandCdxco-expression in the intestine of a Wnt10 knockdown larva.Xloxectopic expression in the intestinal cells is indicated by arrowheads. Absence ofWnt10transcripts in Cdx knockdown larvae is shown in F. (D-F) Full projections of confocalz-series. All larvae have been fixed at pluteus stage and are shown in lateral view, oral side right.
DEVEL
O
(Fig. 4M-O) and suggest that aMHC-specific cis-regulatory module active only in these cells is responsive, directly or indirectly, to an Xlox regulatory input. This regulatory mechanism drives the differentiation into muscles of a small population of endodermal gut cells, allowing the formation of the constriction between stomach and intestine.
A differential transcriptomic analysis for Xlox morphants In order to obtain a global vision of transcriptional responses to Xlox perturbation, we performed a genome-wide comparative transcriptome analysis (RNAseq) on Xlox knockdown versus control 48 and 72 h embryonic RNA (Fig. 5 and Table 1). The MA plots in Fig. 5 clearly show that the effect of Xlox perturbation on gene expression was much stronger at 72 h, thus potentially supporting our hypothesis of a signaling event depending on Xlox and directing stomach differentiation. In particular, we found 5767 differentially expressed transcripts, 23.2% of them upregulated and 28.8% downregulated (by at least 1.5 fold). In Table 1 we provide a list of genes whose expression was significantly affected in the differential RNAseq analysis, grouping them based on their functional category and providing the expression pattern in the sea urchin embryo, when available. Among these, there are genes, such as SpCdx, SpLox and Endo16, that have been previously identified as SpLox targets through qPCR andin situhybridization analysis by Cole et al. (2009). Among the most strongly newly identified downregulated genes are the enzymes involved in the metabolism of proteins, carbohydrates and lipids, possibly supporting a role for Xlox in the differentiation of a functional digestive system. In addition, we found a substantial decrease in the expression of Ache-13 (an acetylcholinesterase that functions in neuromuscular junctions) that we correlated with the absence of pyloric sphincter muscular fibers in Xlox morphant larvae (Fig. 4M-O). Likewise, three transcription factors of the Forkhead family (FoxY,FoxDandFoxP), mainly expressed in foregut and midgut territories, were significantly downregulated, thus suggesting their potential involvement in Xlox gut patterning control.NeuroD1, a
regulator of insulin production in vertebrates (Babu et al., 2008; Kaneto et al., 2009), was also strongly downregulated, indicating a possible evolutionary conserved cooperative function for Xlox and NeuroD1 in the sea urchin larva. Moreover, a GABA receptor and a cholinergic receptor were highly downregulated and we associated these results with the potential role of Xlox in the specification of some neural cells of the ciliary band from a group of ectodermal cells in which Xlox transcripts have been revealed (Cole and Arnone, 2009).
The expression of several gut transcription factors, e.g.SpGataE (Lee and Davidson, 2004),SpHnf1andSpTgiF(Howard-Ashby et al., 2006), andSpPtf1a(Annunziata et al., 2013b), was not significantly affected in Xlox knockdown larvae. Finally, this analysis did not reveal any significantly affected signaling molecule-encoding gene, thus leaving unresolved the mechanism mediating Xlox function in the activation of stomach differentiation genes and introducing new possible hypotheses, such as Xlox requirement in the control of post-translational events, something not testable with the approaches followed so far.
Interestingly, the validation by qPCR of one of the most affected genes,SpPpglcp(a phosphoglycolate phosphatase), with a 3 h time resolution, from 44 h to 50 h (supplementary material Fig. S3), has revealed a strong decrease in its expression since the first analyzed stage (44 h), hinting a possible direct role for Xlox in the activation ofPpglcptranscription. However, information aboutPpglcpspatial expression would be needed for a more complete interpretation of the strong effect of Xlox knockdown on its transcription.
DISCUSSION
Xlox as a potential tissue fate organizer at the stomach-intestine boundary
[image:5.612.113.499.58.299.2]In the present study, we show that the Xlox transcription factor is required to induceChPandManrC1Atranscription in the stomach cells. This, together with the reduction of digestive function and the alteration of the stomach morphology in Xlox morphants (Cole et al., Fig. 4. Xlox controls the differentiation of stomach and pyloric muscles; Cdx repressesEndo-16in the intestinal cells.(A-F,J-N) mRNA localization determined by chromogenicin situhybridization, (G-I,O) full projections of confocalz-series of two-color (G-I) or single-color (O)in situhybridization coupled with nuclear staining (blue). All the larvae have been fixed at 72 h and are shown in lateral view, oral side right. The positions of the pyloric (arrowhead) and anal (arrow) sphincters are indicated. In O, the full projection of az-stack shows absence of pyloric sphincter.
DEVEL
O
2009), as well as the strong reduction in expression of digestive enzymes observed in our transcriptomic analysis, provide a strong support to Xlox key role in the development of a functional stomach. However, the detailed expression dynamics analyzed in this work imply the existence of a signaling event (S1 in Fig. 6) under the control of Xlox that induces the expression of the two stomach-specific genes. Moreover, the sharp boundary of gene expression between stomach and esophagus suggests that the stomach precursor cells are already committed to their future fate, but require an Xlox-induced signal in order to differentiate. Additionally, Xlox is upstream of the entire cascade that regulates intestine formation by acting as a decisive activator ofCdxtranscription in the intestine precursor cells (Cole et al., 2009). Xlox also drives the formation of the pyloric sphincter constriction, triggering the differentiation of muscles from a small subset of endodermal cells, via the activation ofMHCtranscription. Interestingly, the murine Xlox homolog Pdx-1 has a crucial role in pancreas formation and differentiation, and is also involved in pyloric sphincter morphogenesis (Offield et al., 1996). Tissue boundaries keep physically separated neighboring groups of cells with distinct differentiation fates and often act as organizing centers for compartment patterning (for a review, see Dahmann et al., 2011). We propose thatXlox-positive cells located at the stomach-intestine boundary may function as an organizer of gut domains in both the anterior and posterior sides. Thus, the pyloric sphincter represents a very powerful model for investigating the gene regulatory interactions acting at the level of morphological boundaries, and the sea urchin embryo is well suited to this kind of study. A powerful approach
we employed in this context was the genome-wide differential transcriptomic analysis on Xlox morphants. This type of study has provided a large amount of information and has high potential in resolving Xlox patterning functions. Among the most strongly affected genes were the ones encoding metabolic enzymes, muscle-associated proteins and neuronal factors, plus some transcription factors expressed in the gut, strongly suggesting that Xlox is upstream of a number of events leading to the gut AP differentiation.
Finally, we detected Xlox proteins in cells where the mRNA was untraceable. Analyses of gene expression through transcript detection are widely used in the construction of gene regulatory networks. Our study highlights the importance of taking protein in addition to transcript localization into account when defining cellular regulatory states.
Cdx conserved function and regulation in intestine differentiation
[image:6.612.48.401.56.428.2]Whereas the foregut and midgut domains appear definitively established at the late gastrula stage, undergoing mainly morphological changes during the subsequent stages of development, the hindgut domain remains a dynamic site of cell fate specification throughout the late gastrula and early prism stages (see Fig. 6). A crucial event taking place during hindgut development is the activation ofCdx transcription in the most posterior cells of the late gastrula gut; we found a cassette of regulatory factors, namely Xlox, Hox11/13b, FoxA and Bra, involved in Cdx activation. These four transcription factors are all necessary forCdxtranscription, although we do not
Fig. 5. Global effect of Xlox translation perturbation on gene transcription.
(A,B) MA plots showing logarithmic fold changes (on they-axis) as a function of the mean of normalized counts (on thex-axis), in A at 72 h, in B at 48 h. Genes with a significant response at adjustedPvalue <0.05 are in red. Genes mentioned in Table 1 that display 1.5<FC<−1.5 at 72 h are in green, with the exception ofXloxwhich is in blue and magenta in A and B, respectively.
DEVEL
O
know whether they function directly onCdxactivation or recruit intermediate factors. One possibility, as they are all expressed in the cells whereCdxtranscription starts, is that they might function in an
‘AND’ logic (Davidson, 2010), probably together with other transcription factors and/or co-factors, to drive the specification of intestinal cells. Cdx, however, has an‘exclusion effect’(Oliveri and
[image:7.612.51.561.70.616.2]Davidson, 2007), repressing a potential alternative cell regulatory state available to the intestinal cells (its effect onXloxandEndo-16). We discovered that the role of Cdx in repressingXloxin the intestine is mediated by a Wnt10 ligand. In addition, we showed thatWnt10 ligands are actively transcribed in the most posterior cells of the gut at the late gastrula-prism stage and propose that they diffuse towards Table 1. Main effects ofXloxtranslation perturbation on gene transcription
Functional
category Gene name Gene ref*
FC at 48 h
Adjusted Pvalue
at 48 h FC at 72 h
Adjusted Pvalue at 72 h
Gene category
Main domain of expression
at 48 h Reference
Gene regulation and signal transduction
Sp-Lox SPU_020637 1.20 0.02 12.73 0.00E+00 TF HG Arnone et al., 2006 Sp-FoxY SPU_010403 2.17 0.00 10.27 7.02E−30 TF SM Ransick et al., 2002 Sp-IrxB SPU_011246 1.87 0.00 4.61 4.72E−55 TF na
Sp-Nkx3.2 SPU_017837 1.84 0.00 4.43 2.75E−32 TF Endo+AO M.I.A., unpublished Sp-Otx SPU_010424 −1.06 0.79 4.21 1.46E−21 TF Endo+Ecto Li et al., 1997 Sp-Tbr SPU_025584 1.98 0.00 3.88 3.79E−22 TF PMC Oliveri et al., 2002 Sp-Ets4 SPU_008528 1.52 0.00 3.65 0.00E+00 TF Endo+NSM M.I.A., unpublished Sp-Notchl5_1 SPU_016016 1.86 0.00 3.61 2.44E−147 Notch ligand na
Sp-FoxA SPU_006676 1.12 0.00 1.97 1.34E−269 TF MG+HG+NSM Oliveri et al., 2006 Sp-Gatae SPU_010635 1.11 0.00 1.69 1.10E-56 TF MG+HG+NSM Lee and Davidson,
2004 Sp-Tgif SPU_018126 −1.07 0.08 1.40 3.89E−25 TF Endo Howard-Ashby
et al., 2006 Sp-Ptf1a SPU_002677 1.20 0.06 −1.18 5.21E−02 TF MG+SCBC Annunziata et al.,
2013b
Sp-Cdx SPU_024715 −1.18 0.29 −1.53 1.78E−07 TF HG Arnone et al., 2006 Sp-Brn1-2-4 SPU_016443 −1.12 0.02 −1.58 1.86E−29 TF FG+SCBC Cole and Arnone,
2009
Sp-FoxB SPU_004551 1.07 0.94 −1.71 6.86E−02 TF Endo David et al., 1999 Sp-FoxM SPU_025590 −1.01 1.00 −1.90 1.57E−18 TF Ubiq Tu et al., 2006 Sp-Rx SPU_014289 −1.15 0.57 −1.94 1.75E−06 TF Endo+AO Burke et al., 2006 Sp-SoxE SPU_016881 1.06 0.73 −2.15 3.47E−83 TF NSM Luo and Su, 2013 Sp-FoxD SPU_014418 −1.34 0.00 −2.26 6.31E−33 TF HG+AO Tu et al., 2006 Sp-Tbx6 SPU_020346 −1.49 0.14 −2.74 4.05E−12 TF PMC+NSM Gene expression
database‡ Sp-PaxC SPU_000276 −1.00 1.00 −2.93 8.44E−62 TF AO+CB Gene expression
database‡ Sp-Egfrp SPU_004845 −1.95 0.40 −3.67 5.28E−106 Notch or Notch ligand na
Sp-NeuroD1 SPU_024918 na na −3.73 2.06E−16 TF na Sp-Tgfbr3 SPU_027380 1.25 0.92 −6.11 1.57E−03 TGFβreceptor na
Sp-Phox2 SPU_013464 na na −6.57 1.13E−03 TF na
Sp-FoxP SPU_009876 1.21 0.00 −7.12 5.04E−18 TF FG Tu et al., 2006 Cytoskeleton
and membrane
Sp-Endo16 SPU_011038 1.43 0.00 8.24 0.00E+00 Ca2+-binding protein MG+HG Ransick et al., 1993 Sp-Gelsolin SPU_003985 −1.74 0.65 −3.71 4.19E−35 Actin-binding protein na
Sp-Actb SPU_006661 1.73 0.00 −6.50 1.68E−62 Cytoskeleton actin na Sp-Ache-13 SPU_005168 na na −8.36 5.54E−24 Acetylcholinesterase na Metabolism
and respiration
Sp-Cyp3L12 SPU_027796. 5.76 0.00 15.89 6.56E−91 Cytochrome p450 na Sp-MeprinAaL SPU_030013 3.13 0.00 4.75 1.69E−85 Protein metabolism na Sp-Alpi SPU_022639 −1.31 0.03 −2.85 2.72E−63 Intestinal AP
precursor
na
Sp-Mgl SPU_010842 na na −3.94 8.64E−04 Methionineγ-lyase na
Sp-Cpa3L_2 SPU_017120 na na −5.44 5.61E−72 Carboxypeptidase MG Annunziata et al., 2013b Sp-Cub/
TolloidL
WHL22.2178 1.00 1.00 −7.79 8.64E−04 Cubilin, intestinal factor
na
Sp-PlbL SPU_015099 na na −9.91 6.11E−15 Phospholipase MG Annunziata et al., 2013b SPU_007135 SPU_007135 na 0.77 −10.70 1.06E−59 Regucalcin na
Sp-Aadac_1 SPU_014449 na na −10.74 3.56E−42 Lipase na Sp-SpdecL SPU_002513 na na −10.78 4.05E−22 Sphingomyelinase C na SPU_013685 SPU_013685 na na −16.02 4.01E−42 Lipase na Sp-Ppglcp SPU_017527 −5.45 0.00 −20.15 0.00E+00 Phosphoglycolate na
Phosphatase
*Gene reference according to spbase (http://sugp.caltech.edu/SpBase/). ‡http://goblet.molgen.mpg.de/cgi-bin/seaurchin-database.cgi
AO, apical organ; AP, alkaline phosphatase; CB, ciliary band; Ecto, ectoderm; Endo, endoderm; FG, fold change; FG, foregut; HG, hindgut; MG, midgut; na, not available; NSM, non skeletogenic mesoderm; PMC, primary mesenchyme cell; SCBC, scattered ciliary band cells; SM, small micromere lineage; TF, transcription factor; Ubiq, ubiquitous.
DEVEL
O
the anterior side of the gut, progressively clearing the intestinal cells fromXlox(and probably other) transcripts. We do not know whether the newly identified Wnt signal works throughβ-catenin or activates a non-canonical Wnt pathway. The identity of the receptor mediating this Wnt10 signaling event is also unknown. Four Frizzled receptor genes have been identified so far in the Strongylocentrotus purpuratus genome but no information is available about the expression dynamics of these genes during embryogenesis. The expression of these receptors has been deeply investigated in another sea urchin species, Paracentrotus lividus, indicating Fzd9/10 as possible candidate for mediating Wnt10 signaling in the hindgut (Croce et al., 2006; Lhomond et al., 2012; Robert et al., 2014). It would be interesting to study the dynamics of expression of the four Frizzled receptor genes in S. purpuratus in order to identify a possible candidate for the transduction of the Wnt10 signal to the intestinal cells. Interestingly, mechanisms of function for Cdx transcription factors involving activation of Wnt signaling have been previously observed in mouse, precisely in the uro-rectal mesoderm development (van de Ven et al., 2011). Moreover, Cdxappears, together withBra,FoxAandWnt, as part of a conserved cassette of factors regulating posterior gut development. These genes are all expressed in the blastopore or blastopore equivalent of frog, zebrafish, Drosophila and mouse (Lengyel and Iwaki, 2002). Furthermore, Cdx genes, Wnt signaling, Bra and posterior Hox genes act together to control posterior morphogenesis in the different
murine embryonic germ layers (van de Ven et al., 2011). We found Cdx,Wnt10,FoxA,BraandHox11/13bactively involved in the sea urchin posterior gut development, suggesting conservation of a gene network functioning for the differentiation of posterior structures. The strong reduction ofBraexpression in Cdx morphant embryos observed by qPCR (supplementary material Fig. S4), together with the very similar expression pattern observed forBraandWnt10from late gastrula stage until pluteus, allow us to propose two possible scenarios: (1)Brareceives a fundamental regulatory input from Cdx and participates, together with Cdx, inWnt10activation in the very posterior cells of the post gastrular embryo; (2) Cdx activatesWnt10 expression indirectly, by activating Bra. Both scenarios would provide an explanation for the fact thatWnt10is expressed only in a subset ofCdx-expressing cells, although we have no data so far to support a role for Bra inWnt10activation. This would not be the first case in which Bra activates Wnt signals: in chordates, Bra recruits canonical Wnt signaling to sustain the posterior mesodermal progenitors during the outgrowth of the body and to regulate the mechanism of somitogenesis (Martin and Kimelman, 2009).
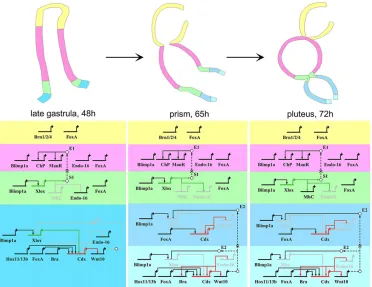
[image:8.612.122.494.57.344.2]We also showed thatEndo-16transcripts accumulate ectopically in the intestine of Cdx morphant larvae and that Hox11/13b is required for activatingCdxtranscription, thus providing a candidate for the factor hypothesized to mediate the Hox11/13b knockdown phenotype in the posterior gut (Arenas-Mena et al., 2006). Interestingly, Cdx also functions as a repressor in the intestine of vertebrates: Cdx2 Fig. 6. Regulatory states and the view from the nuclei of interactions in each compartment of the sea urchin developing gut.In the upper panels, three stages of the sea urchin embryonic gut are depicted in lateral view. Different colors are used for compartments showing exclusive regulatory states. In the lower part, the regulatory states and the interactions found within this study are summarized, using the same color code as the schematics of guts above. The wiring between the genes is shown with solid lines for representation issues but none of them has been demonstrated to be direct. In the diagram, arrows represent positive regulation, bars represent repression. The white bullets, together with the black dashed lines, indicate signaling events: S1 represents the unknown signaling molecule under the control of Xlox and directing stomach genes activation through the action of an unknown effector gene (E1); E2 represents the effector gene responsible for Xlox repression in the intestinal cells.XloxandCdxgenes, together with their inputs (when active), have been colored green and red, respectively. Most of the results obtained throughin situhybridization have been validated by qPCR (see supplementary material Fig. S4). The input ofCdx onBratranscription has been discovered by qPCR (supplementary material Fig. S4). The expression dynamics ofGataE,Blimp1aandFoxAhave been extrapolated from published works (Lee and Davidson, 2004; Livi and Davidson, 2006; Oliveri et al., 2006).ManrC1Ahas been shortened toManRfor clarity.
DEVEL
O
conditional ablation from early murine endoderm results in a posterior-to-anterior gut transformation, through the replacement of the posterior intestinal epithelium with esophageal epithelia (Gao et al., 2009). Furthermore, ectopic expression ofPdx1(the ortholog of the sea urchinXloxgene) has been demonstrated in Cdx2 mouse mutant intestine (Grainger et al., 2010). As already suggested by Grainger et al. (2010), this would hint at conservation of the cross-regulatory loop described in sea urchin between the two genes. However, two phases of Cdx activation have been observed in chordates (Chawengsaksophak et al., 2004; Osborne et al., 2009; Reece-Hoyes et al., 2002), the first in the blastopore-primitive streak regions, where it is required for axial elongation, at least in mouse (Chawengsaksophak et al., 2004), and the second in the posterior gut, where it is involved in intestinal patterning. In the sea urchin embryo, Cdxtranscription starts towards the end of gastrulation. Thus, in the sea urchin, Cdx function in the early phases of gastrulation has been lost and only its function in posterior gut patterning has been retained. This condition is more likely to be an echinoid peculiarity rather than a feature common to all ambulacrarians or to all echinoderms, as Cdx ‘biphasic’ transcription has been described in sea stars and hemichordates (Annunziata et al., 2013a; Ikuta et al., 2013). Very importantly, the fact that Cdx is not required in the early steps of sea urchin gastrulation offers a great advantage for the study of its role in the intestine differentiation, facilitating the interpretation of functional studies.
Conclusions and perspectives
The purpose of this work was to investigate the gut patterning process in its entirety, exploring the dynamics of gene expression and regulation in space and time, a feasible objective using the sea urchin embryo as model system. The data we present here clearly show that while the initial patterning of the gut tube relies mainly on the A-P distribution of transcription factor territories of expression, the subsequent compartmentalization depends, at least partially, on signaling events under the control of the two ParaHox genes,Xlox andCdx(see Fig. 6). It is important to note that none of the interactions found in this study has been demonstrated to be direct and that the wiring diagram presented in Fig. 6 remains far from being complete, missing many other levels of regulation such as the involvement of small non coding RNAs, post translational modifications, protein-protein interactions and chromatin remodeling.
Our findings suggest that Xlox ancestral function in the endoderm was probably to pattern the digestive tube directing stomach cell differentiation and pyloric sphincter formation, as supported by conservation of its gut domain of expression in several protostomes and non-vertebrate deuterostomes. The vertebrate homolog Pdx-1 has been possibly coopted for the development of the pancreas, maintaining its role in the formation of the stomach-intestine constriction. Similarly, the crossregulatory loop between Xlox and Cdxfirstly described in sea urchin (Cole et al., 2009) appears as a conserved gut patterning mechanism among deuterostomes (Grainger et al., 2010). We predict that many of the hereby demonstrated regulatory interactions shaping the sea urchin gut, and the many others we will find by the ongoing ChIPSeq and RNASeq analyses downstream of SpLox and SpCdx, will be instrumental for the discovery of yet unknown patterning mechanisms of the vertebrate gut.
MATERIALS AND METHODS Animals, embryo cultures
AdultStrongylocentrotus purpuratus were obtained from Patrick Leahy
(Kerchoff Marine Laboratory, California Institute of Technology, Pasadena, USA) and housed in circulating sea water aquaria in the Stazione Zoologica
Anton Dohrn of Naples, Italy. Gametes were obtained by standard methods and embryos were cultured at 15°C in filtered sea water diluted 9:1 with de-ionized water.
RNAin situhybridization
For single gene expression, we followed the protocol outlined previously
(Minokawa et al., 2004). Fluorescent double WMin situhybridization were
performed as described previously (Cole et al., 2009). RNA probe
sequences forSpLox, SpCdx, SpBrn1/2/4, SpBra and SpEndo16 are as
previously published:SpLoxandSpCdx(Arnone et al., 2006);SpBrn1/2/4
(Cole and Arnone, 2009);SpBra(Rast et al., 2002); andSpEndo16(Ransick
et al., 1993).SpHox11/13b,SpChP,SpManrC1A,SpMHCbacterial clones
were picked from theS. purpuratuscDNA library available in the laboratory
(http://goblet.molgen.mpg.de/cgi-bin/seaurchin-database.cgi).SpWnt10cDNA
was obtained by RT-PCR from 65 h embryo total RNA (using the following
primers: SpWnt10-F: 50-AGACGATGGAATTGCTCCAG-30; SpWnt10-R:
50-GGTTAACCCATTGCGAGCTA-30) and then cloned into the Topo-TA
cloning vector (Invitrogen).
Immunohistochemistry
For acetylated tubulin staining combined with in situ hybridization, a
1:250 dilution of the mouse monoclonal anti-acetylated tubulin antibody (T7451; Sigma-Aldrich) was added to the blocking solution containing the peroxidase-conjugated antibody and incubated overnight at 4°C. The next
day the embryos were processed forin situhybridization, then incubated
for 1 h in blocking solution with a dilution 1:1000 of the AlexaFluor 488 goat anti-mouse immunoglobulin G (IgG) (A21202; Molecular Probes), washed in MOPS (3-morpholinopropane-1-sulfonic acid) buffer and mounted for imaging. For Xlox and Cdx immunostaining, larvae were fixed in 2% PFA in phosphate-buffered saline (PBS) for 10 min at room temperature, washed multiple times in PBS with 0.1% Tween-20 (PBST), blocked in 5% goat serum in PBST and incubated overnight at 4°C with the custom primary antibodies diluted at working concentration (1:500; PRIMM) in 5% goat serum in PBST. For Bra immunostaining, embryos and larvae were fixed in 2% PFA in PBS for 10 min, then for 1 min in methanol, blocked in 5% goat serum in PBST and incubated overnight with the custom antibody at working concentration (1:200; A11008; PRIMM). Following primary antibody incubation, embryos and larvae were washed several times in PBST, incubated for 1 h at room temperature with the secondary antibody Alexa Fluor 488 goat anti-rabbit IgG (Molecular Probes, Invitrogen) diluted 1:1000 in 5% goat serum in PBST, washed in PBST and mounted for imaging with a confocal microscope (Zeiss 510Meta).
MO microinjection
For each experiment and for each morpholino oligonucleotide (MO), around 400 embryos were injected with approximately 2-4 pl of oligonucleotide solution at 0.1 mM and each experiment was repeated three times. In all experiments, as a negative control, embryos were injected with 0.1 mM of the standard control morpholino and compared side by side with uninjected and knockdown embryos. The injection of the standard control morpholino (GeneTools) did not have any effect on the development of embryos. MOs against SpLox, SpHox11/13b, SpFoxA, SpBlimp1a, SpGataE, SpTgiF and SpBra protein translation and the morpholino targeting the donor splice site
between the first and secondSpCdxexons, were already available (http://sugp.
caltech.edu/endomes/) (Arenas-Mena et al., 2006; Cole et al., 2009; Livi and Davidson, 2006; Oliveri et al., 2006; Rast et al., 2002). MOs against SpWnt10 and SpCdx translation were newly designed and acquired from Gene
Tools (Corvallis) (SpWnt10, 50-AACTGCATCTGCTTACGATTCATAC-30;
SpCdxM, 50-TGGGTGCAGATACTCTAGCGTCATC-30).
Screening of signaling molecules
In order to identify the signaling molecule responsible for the midgut differentiation process, 15 signaling ligand encoding genes have been analyzed. The genes included in the analysis were selected from the
Strongylocentrotus purpuratusSignaling Ligand Page (available at http://
140.109.48.251/ICOBUserfile/SuYuLab/Su_and_Yu_Lab/Home.html), a
DEVEL
O
database created by Yi-Hsien Su’s laboratory (Institute of Cellular and Organismic Biology, Academia Sinica, Taiwan), in which the temporal expression pattern of all signaling ligands expressed during sea urchin embryonic development is provided. The signaling ligand database was screened using as selective criteria the windows of activation of the genes. In particular, the ligands whose expression started or significantly increased a
few hours after 44 hpf (the developmental time when Cdx expression
begins) were considered to be eligible signaling molecules and screened by qPCR. The differential quantitative expression of the selected genes in control and Cdx knockdown embryos (data not shown) was analyzed. The
15 signaling molecules included in the analysis were:SpWnt8, SpIGF2,
SpHGF,SpAgrin3,SpVegF,SpBMP3,SpBMP5/8,SpNotch-lik2, SpNotch-lig3,SpNotch-lid5,SpEphrin,SpSHH,SpWnt10,SpWnt4andSpFGF9/16/
20. For the qPCR, the primers were designed based on the sequences
available in the signaling ligand database.
RNA extraction, RNA-seq and data analysis
Three biological replicas of 500 injected withSpLoxMO and control
(KCl-injected) embryos were collected at 48 and 72 h. About 1μg of RNA was
extracted from each sample using the RNAqueous-Micro Kit (Ambion, Life Technologies-Invitrogen) and used for RNA-seq. cDNA libraries were prepared with 1 µg of starting total RNA and using the Illumina TruSeq RNA Sample Preparation Kit (Illumina), according to TruSeq protocol. Library size and integrity were determined using the Agilent Bioanalyzer 2100. Each library was diluted to 2 nM and denaturated, 8 pM of each library was loaded onto cBot (Illumina) for cluster generation with cBot Paired End Cluster Generation Kit (Illumina) and sequenced using the Illumina HiSeq 1500 with 100 bp paired-end reads in triplicate, obtaining
∼31-38 million reads for replicate. The sequencing service was provided by
the Laboratory of Molecular Medicine and Genomics (http://www. labmedmolge.unisa.it) at the University of Salerno, Italy. Sequence read quality was controlled using FastQC program (http://www.bioinformatics. bbsrc.ac.uk/projects/fastqc/). Filtered reads were mapped to the Sea Urchin genome Spur_3_1.LinearScaffold downloaded from SpBase (www. spbase.org) using TopHat v2.0.8b (Trapnell et al., 2009), that runs on Bowtie 2 version 2.1.0 (Langmead et al., 2009). The reads were counted using the HTSeq-package (Anders and Huber, 2010). Normalization of read numbers between samples and differential expression analysis was performed using DEseq (Anders and Huber, 2010). The transcripts with an
adjusted P<0.05 were considered to be differentially expressed.
Differential expression is represented with MA-plots generated with the DESeq software.
Acknowledgements
We acknowledge Patrick Leahy and the Southern California Sea Urchin Company for animal supply; Davide Caramiello for taking care of the sea urchins; the SZN Molecular Biology Service for technical assistance; the Laboratorio di Medicina Molecolare e Genomica, Universitàdegli Studi di Salerno, Italy, for the RNA-seq experiment; Giorgio Giurato and Francesca Rizzo for the MA-plots; Eric Davidson for Bra, FoxA, GataE and TgiF MOs; Yi-Hsien Su for the signaling molecule database; Evelyn Houliston and Pedro Martinez for revising our manuscript; and Claudia Cuomo for the kind experimental help.
Competing interests
The authors declare no competing financial interests.
Author contributions
R.A. performed all the experimental work with the exception of the microinjections ( performed by M.I.A.), contributed to the interpretation of the results and wrote the manuscript. M.I.A. conceived and supervised the project, contributed to the analysis of the data and prepared the figures.
Funding
This work was supported by the‘Evonet’( project 215781) EU-Marie Curie Early Training Network, and (a fellowship to R.A.) by POR Campania FSE 2007-2013 Project MODO, Model Organism.
Supplementary material
Supplementary material available online at
http://dev.biologists.org/lookup/suppl/doi:10.1242/dev.105775/-/DC1
References
Anders, S. and Huber, W.(2010). Differential expression analysis for sequence count data.Genome Biol.11, R106.
Andrikou, C., Iovene, E., Rizzo, F., Oliveri, P. and Arnone, M. I. (2013). Myogenesis in the sea urchin embryo: the molecular fingerprint of the myoblast precursors.EvoDevo4, 33.
Annunziata, R., Martinez, P. and Arnone, M. I. (2013a). Intact cluster and chordate-like expression of ParaHox genes in a sea star.BMC Biol.11, 68.
Annunziata, R., Perillo, M., Andrikou, C., Cole, A. G., Martinez, P. and Arnone, M. I.
(2013b). Pattern and process during sea urchin gut morphogenesis: the regulatory landscape.Genesis52, 251-268.
Arenas-Mena, C., Cameron, R. A. and Davidson, E. H. (2006). Hindgut specification and cell-adhesion functions of Sphox11/13b in the endoderm of the sea urchin embryo.Dev. Growth Differ.48, 463-472.
Arnone, M. I., Rizzo, F., Annunciata, R., Cameron, R. A., Peterson, K. J. and Martı́nez, P. (2006). Genetic organization and embryonic expression of the ParaHox genes in the sea urchin S. purpuratus: insights into the relationship between clustering and colinearity.Dev. Biol.300, 63-73.
Babu, D. A., Chakrabarti, S. K., Garmey, J. C. and Mirmira, R. G.(2008). Pdx1 and BETA2/NeuroD1 participate in a transcriptional complex that mediates short-range DNA looping at the insulin gene.J. Biol. Chem.283, 8164-8172.
Brooke, N. M., Garcia-Fernàndez, J. and Holland, P. W. H.(1998). The ParaHox gene cluster is an evolutionary sister of the Hox gene cluster.Nature392, 920-922.
Burke, R. D.(1981). Structure of the digestive tract of the pluteus larva of dendraster excentricus (echinodermata: echinoida).Zoomorphology98, 209-225.
Burke, R. D., Angerer, L. M., Elphick, M. R., Humphrey, G. W., Yaguchi, S., Kiyama, T., Liang, S., Mu, X., Agca, C., Klein, W. H. et al.(2006). A genomic view of the sea urchin nervous system.Dev. Biol.300, 434-460.
Chawengsaksophak, K., de Graaff, W., Rossant, J., Deschamps, J. and Beck, F.
(2004). Cdx2 is essential for axial elongation in mouse development.Proc. Natl. Acad. Sci. U.S.A.101, 7641-7645.
Cole, A. G. and Arnone, M. I.(2009). Fluorescentin situhybridization reveals multiple expression domains for SpBrn1/2/4 and identifies a unique ectodermal cell type that co-expresses the ParaHox gene SpLox.Gene Expr. Patterns9, 324-328.
Cole, A. G., Rizzo, F., Martinez, P., Fernandez-Serra, M. and Arnone, M. I.(2009). Two ParaHox genes, SpLox and SpCdx, interact to partition the posterior endoderm in the formation of a functional gut.Development136, 541-549.
Croce, J. C. and McClay, D. R.(2010). Dynamics of Delta/Notch signaling on endomesoderm segregation in the sea urchin embryo. Development 137, 83-91.
Croce, J., Duloquin, L., Lhomond, G., McClay, D. R. and Gache, C.(2006). Frizzled5/8 is required in secondary mesenchyme cells to initiate archenteron invagination during sea urchin development.Development133, 547-557.
Dahmann, C., Oates, A. C. and Brand, M.(2011). Boundary formation and maintenance in tissue development.Nat. Rev. Genet.12, 43-55.
David, E.-S., Luke, N. H. and Livingston, B. T.(1999). Characterization of a gene encoding a developmentally regulated winged helix transcription factor of the sea urchin Strongylocentrotus purpuratus.Gene236, 97-105.
Davidson, E. H.(2010). Emerging properties of animal gene regulatory networks.
Nature468, 911-920.
Davidson, E. H., Rast, J. P., Oliveri, P., Ransick, A., Calestani, C., Yuh, C.-H., Minokawa, T., Amore, G., Hinman, V., Arenas-Mena, C. et al.(2002). A genomic regulatory network for development.Science295, 1669-1678.
Donoghue, P. C. J. and Purnell, M. A.(2005). Genome duplication, extinction and vertebrate evolution.Trends Ecol. Evol.20, 312-319.
Feng, R., Xiao, C. and Zavros, Y.(2012). The role of Sonic Hedgehog as a regulator of gastric function and differentiation.Vitam. Horm.88, 473-489.
Gao, N., White, P. and Kaestner, K. H.(2009). Establishment of intestinal identity and epithelial-mesenchymal signaling by Cdx2.Dev. Cell16, 588-599.
Grainger, S., Savory, J. G. A. and Lohnes, D.(2010). Cdx2 regulates patterning of the intestinal epithelium.Dev. Biol.339, 155-165.
Gustafson, T. and Wolpert, L.(1967). Cellular movement and contact in sea urchin morphogenesis.Biol. Rev.42, 442-498.
Holland, P. W. H.(2013). Evolution of homeobox genes.Wiley Interdiscip. Rev. Dev. Biol.2, 31-45.
Howard-Ashby, M., Materna, S. C., Brown, C. T., Chen, L., Cameron, R. A. and Davidson, E. H. (2006). Identification and characterization of homeobox transcription factor genes in Strongylocentrotus purpuratus, and their expression in embryonic development.Dev. Biol.300, 74-89.
Ikuta, T., Chen, Y.-C., Annunziata, R., Ting, H.-C., Tung, C.-H., Koyanagi, R., Tagawa, K., Humphreys, T., Fujiyama, A., Saiga, H. et al.(2013). Identification of an intact ParaHox cluster with temporal colinearity but altered spatial colinearity in the hemichordate Ptychodera flava.BMC Evol. Biol.13, 129.
Jacobs, I. J., Ku, W.-Y. and Que, J.(2012). Genetic and cellular mechanisms regulating anterior foregut and esophageal development.Dev. Biol.369, 54-64.
Kaneto, H., Matsuoka, T.-a., Katakami, N. and Matsuhisa, M. (2009). Combination of MafA, PDX-1 and NeuroD is a useful tool to efficiently induce insulin-producing surrogate beta-cells.Curr. Med. Chem.16, 3144-3151.
DEVEL
O
Langmead, B., Trapnell, C., Pop, M. and Salzberg, S. L.(2009). Ultrafast and memory-efficient alignment of short DNA sequences to the human genome.
Genome Biol.10, R25.
Lee, P. Y. and Davidson, E. H.(2004). Expression of Spgatae, the Strongylocentrotus purpuratus ortholog of vertebrate GATA4/5/6 factors. Gene Expr. Patterns5, 161-165.
Lengyel, J. A. and Iwaki, D. D.(2002). It takes guts: the Drosophila hindgut as a model system for organogenesis.Dev. Biol.243, 1-19.
Lhomond, G., McClay, D. R., Gache, C. and Croce, J. C.(2012). Frizzled1/2/7 signaling directs beta-catenin nuclearisation and initiates endoderm specification in macromeres during sea urchin embryogenesis.Development139, 816-825.
Li, X., Chuang, C.-K., Mao, C.-A., Angerer, L. M. and Klein, W. H.(1997). Two Otx proteins generated from multiple transcripts of a single gene in Strongylocentrotus purpuratus.Dev. Biol.187, 253-266.
Livi, C. B. and Davidson, E. H.(2006). Expression and function of blimp1/krox, an alternatively transcribed regulatory gene of the sea urchin endomesoderm network.Dev. Biol.293, 513-525.
Luo, Y.-J. and Su, Y.-H.(2012). Opposing nodal and BMP signals regulate left-right asymmetry in the sea urchin larva.PLoS Biol.10, e1001402.
Martin, B. L. and Kimelman, D.(2009). Wnt signaling and the evolution of embryonic posterior development.Curr. Biol.19, R215-R219.
McClay, D. R.(2011). Evolutionary crossroads in developmental biology: sea urchins.Development138, 2639-2648.
McGhee, J. D.(2007). The C. elegans intestine.WormBook, 1-36.
Minokawa, T., Rast, J. P., Arenas-Mena, C., Franco, C. B. and Davidson, E. H.
(2004). Expression patterns of four different regulatory genes that function during sea urchin development.Gene Expr. Patterns4, 449-456.
Nielsen, C.(2008). Six major steps in animal evolution: are we derived sponge larvae?Evol. Dev.10, 241-257.
Offield, M. F., Jetton, T. L., Labosky, P. A., Ray, M., Stein, R. W., Magnuson, M. A., Hogan, B. L. and Wright, C. V.(1996). PDX-1 is required for pancreatic outgrowth and differentiation of the rostral duodenum.Development122, 983-995.
Oliveri, P. and Davidson, E. H.(2007). Development: built to run, not fail.Science
315, 1510-1511.
Oliveri, P., Carrick, D. M. and Davidson, E. H.(2002). A regulatory gene network that directs micromere specification in the sea urchin embryo.Dev. Biol.246, 209-228.
Oliveri, P., Walton, K. D., Davidson, E. H. and McClay, D. R.(2006). Repression of mesodermal fate by foxa, a key endoderm regulator of the sea urchin embryo.
Development133, 4173-4181.
Osborne, P. W., Benoit, G., Laudet, V., Schubert, M. and Ferrier, D. E. K.(2009). Differential regulation of ParaHox genes by retinoic acid in the invertebrate chordate amphioxus (Branchiostoma floridae).Dev. Biol.327, 252-262.
Peter, I. S. and Davidson, E. H.(2010). The endoderm gene regulatory network in sea urchin embryos up to mid-blastula stage.Dev. Biol.340, 188-199.
Ransick, A., Ernst, S., Britten, R. J. and Davidson, E. H.(1993). Whole mount
in situhybridization shows Endo 16 to be a marker for the vegetal plate territory in sea urchin embryos.Mech. Dev.42, 117-124.
Ransick, A., Rast, J. P., Minokawa, T., Calestani, C. and Davidson, E. H.(2002). New early zygotic regulators expressed in endomesoderm of sea urchin embryos discovered by differential array hybridization.Dev. Biol.246, 132-147.
Rast, J. P., Cameron, R. A., Poustka, A. J. and Davidson, E. H.(2002). brachyury Target genes in the early sea urchin embryo isolated by differential macroarray screening.Dev. Biol.246, 191-208.
Reece-Hoyes, J. S., Keenan, I. D. and Isaacs, H. V. (2002). Cloning and expression of the Cdx family from the frog Xenopus tropicalis.Dev. Dyn.223, 134-140.
Robert, N., Lhomond, G., Schubert, M. and Croce, J. C.(2014). A comprehensive survey of wnt and frizzled expression in the sea urchin Paracentrotus lividus.
Genesis52, 235-250.
Roberts, D. J.(2000). Molecular mechanisms of development of the gastrointestinal tract.Dev. Dyn.219, 109-120.
Sherwood, R. I., Maehr, R., Mazzoni, E. O. and Melton, D. A.(2011). Wnt signaling specifies and patterns intestinal endoderm.Mech. Dev.128, 387-400.
Spence, J. R., Lauf, R. and Shroyer, N. F.(2011). Vertebrate intestinal endoderm development.Dev. Dyn.240, 501-520.
Trapnell, C., Pachter, L. and Salzberg, S. L.(2009). TopHat: discovering splice junctions with RNA-Seq.Bioinformatics25, 1105-1111.
Tu, Q., Brown, C. T., Davidson, E. H. and Oliveri, P.(2006). Sea urchin Forkhead gene family: phylogeny and embryonic expression.Dev. Biol.300, 49-62.
van de Ven, C., Bialecka, M., Neijts, R., Young, T., Rowland, J. E., Stringer, E. J., Van Rooijen, C., Meijlink, F., Novoa, A., Freund, J.-N. et al.(2011). Concerted involvement of Cdx/Hox genes and Wnt signaling in morphogenesis of the caudal neural tube and cloacal derivatives from the posterior growth zone.Development
138, 3451-3462.
van den Brink, G. R.(2007). Hedgehog signaling in development and homeostasis of the gastrointestinal tract.Physiol. Rev.87, 1343-1375.
Venuti, J. M., Gan, L., Kozlowski, M. T. and Klein, W. H.(1993). Developmental potential of muscle cell progenitors and the myogenic factor SUM-1 in the sea urchin embryo.Mech. Dev.41, 3-14.
Wolpert, L.(1994). The evolutionary origin of development: cycles, patterning, privilege and continuity.Dev. Suppl.79-84.
Zorn, A. M. and Wells, J. M.(2009). Vertebrate endoderm development and organ formation.Annu. Rev. Cell Dev. Biol.25, 221-251.