Embryonic stem cell-derived extracellular vesicles enhance the therapeutic effect of mesenchymal stem cells
Full text
Figure

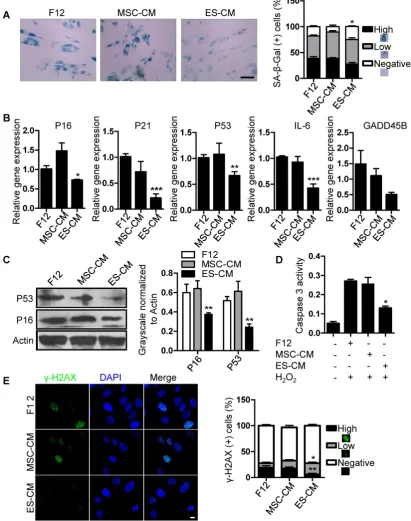
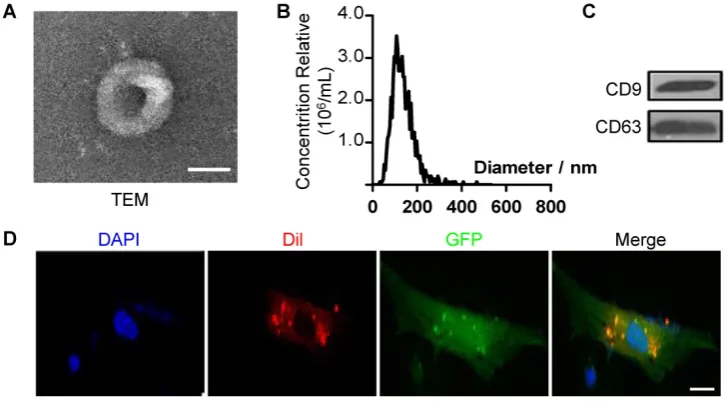
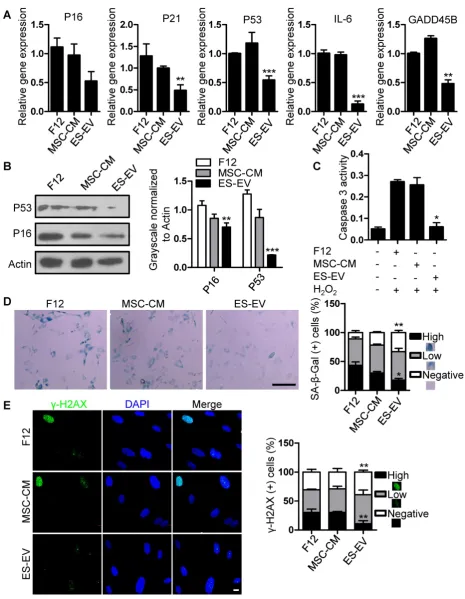
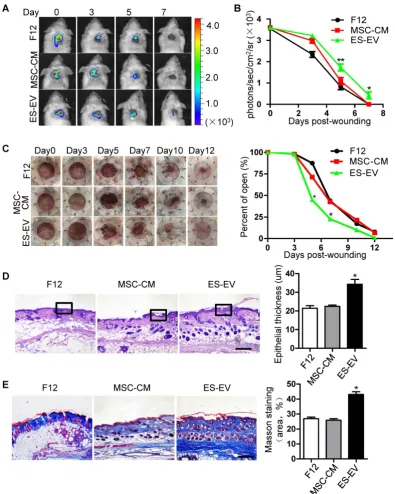
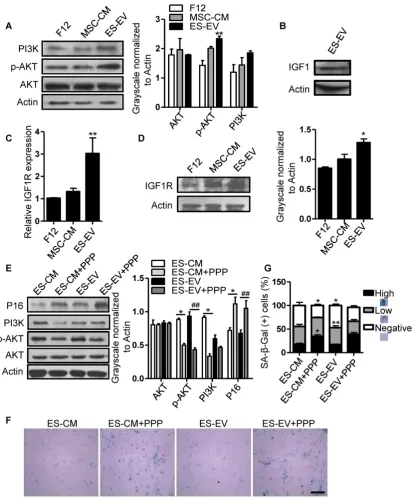
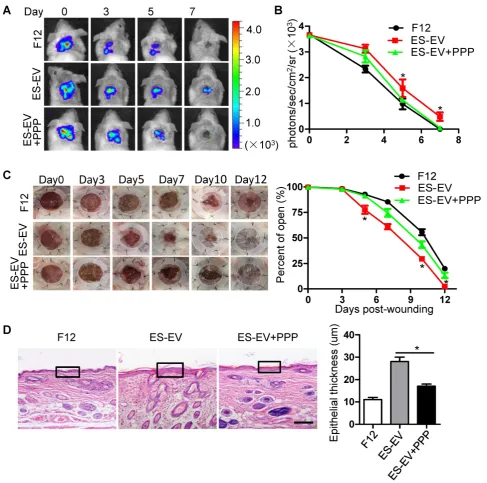
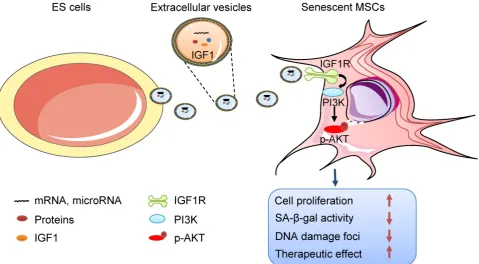
Related documents
Communication needs equal competence as mirrored in the following stories; the equality is shown by the same element (source of creation) of the soul and the angel. So,
MPS Data Store enables on-demand sensor data transmission by separating the search function and storage function between the MPS Cloud and the mobile devices..
Group 2 (Fanta fed rats) and Group 6(Coca-cola fed rats) showed high level of glucose (p<0.05) compared to normal control, while the groups administered with Sprite, Schweppes
Primary gastrointestinal lymphomas in Turkey: a retrospective analysis of clinical features and results of treatment. Gastrointestinal lymphoma in southern
influence of Service Recovery and Loyalty on perceived Service Quality”. A study of Hotel Customers in Spain”. 68) Morrison Coulthard, Lisa .J., “Measuring Service Quality. A
Ideally, as a partnership progresses different practices and attitudes are required at both partnership and organizational level if the partnership is to continue to sustain
Our results shows that the regional economic growth has a strong space correlation; After considering the spatial factors, the 67 cities have the convergence trend
