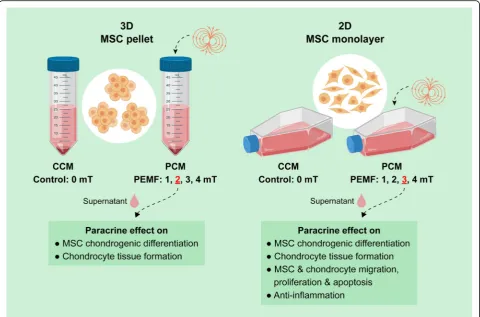
R E S E A R C H
Open Access
Pulsed electromagnetic fields potentiate
the paracrine function of mesenchymal
stem cells for cartilage regeneration
Dinesh Parate
1,2,3, Nurul Dinah Kadir
3, Cenk Celik
3, Eng Hin Lee
3,4, James H. P. Hui
3,4,
Alfredo Franco-Obregón
1,2,5*and Zheng Yang
3,4*Abstract
Background:The mesenchymal stem cell (MSC) secretome, via the combined actions of its plethora of biologically active factors, is capable of orchestrating the regenerative responses of numerous tissues by both eliciting and amplifying biological responses within recipient cells. MSCs are“environmentally responsive”to local micro-environmental cues and biophysical perturbations, influencing their differentiation as well as secretion of bioactive factors. We have previously shown that exposures of MSCs to pulsed electromagnetic fields (PEMFs) enhanced MSC chondrogenesis. Here, we investigate the influence of PEMF exposure over the paracrine activity of MSCs and its significance to cartilage regeneration.
Methods:Conditioned medium (CM) was generated from MSCs subjected to either 3D or 2D culturing platforms, with or without PEMF exposure. The paracrine effects of CM over chondrocytes and MSC chondrogenesis, migration and
proliferation, as well as the inflammatory status and induced apoptosis in chondrocytes and MSCs was assessed.
Results:We show that benefits of magnetic field stimulation over MSC-derived chondrogenesis can be partly ascribed to its ability to modulate the MSC secretome. MSCs cultured on either 2D or 3D platforms displayed distinct magnetic sensitivities, whereby MSCs grown in 2D or 3D platforms responded most favorably to PEMF exposure at 2 mT and 3 mT amplitudes, respectively. Ten minutes of PEMF exposure was sufficient to substantially augment the chondrogenic potential of MSC-derived CM generated from either platform. Furthermore, PEMF-induced CM was capable of enhancing the migration of chondrocytes and MSCs as well as mitigating cellular inflammation and apoptosis.
Conclusions:The findings reported here demonstrate that PEMF stimulation is capable of modulating the paracrine function of MSCs for the enhancement and re-establishment of cartilage regeneration in states of cellular stress. The PEMF-induced modulation of the MSC-derived paracrine function for directed biological responses in recipient cells or tissues has broad clinical and practical ramifications with high translational value across numerous clinical applications.
Keywords:Pulse electromagnetic fields, Mesenchymal stem cells, Cartilage, Paracrine
Background
Articular cartilage is an avascular tissue with limited intrin-sic capacity for regeneration. In combination with the limi-tations of existing treatment modalities, joint injuries often deteriorate with time into articular joint disease [1, 2].
Mesenchymal stem cells (MSCs), with their capacity for expansion and demonstrated multipotency for a variety of tissue lineages, have been championed as a promising cell source for the repair and regeneration of many degenera-tive, inflammatory, or autoimmune diseases [3].
Neverthe-less, despite their anticipated, but at times not
substantiated, potential, MSC-based strategies have often fallen short of initial expectations [4]. Although the poten-tial applicability of MSCs for cartilage repair was inipoten-tially postulated based on their ability to differentiate into chon-drocytes and to participate in the formation of tissue, it has
© The Author(s). 2020Open AccessThis article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
* Correspondence:[email protected];[email protected]
1Department of Surgery, National University of Singapore, Singapore 119228, Singapore
3Department of Orthopaedic Surgery, Yong Loo Lin School of Medicine, National University of Singapore, NUHS Tower Block, Level 11, 1E Kent Ridge Road, Singapore 119288, Singapore
become increasingly evident that part of the reparative value of MSCs is attributed to their paracrine manner of
developmental entrainment [5]. In non-contact co-culture
experiments between MSCs and primary chondrocytes, im-provements in chondrocyte proliferation, phenotype main-tenance, and enhanced matrix synthesis were shown to be credited to MSC-derived trophic factors [6,7]. A paracrine role of MSCs was further supported by preclinical studies showing that intra-articular injection of MSCs prevented the development of post-traumatic arthritis and promoted cartilage regeneration in damaged joints [8], as well as im-proved clinical outcomes and indices of cartilage repair in second-look arthroscopy [9].
The spectrum of trophic factors released by MSCs is col-lectively referred to as the secretome. Via the combined ac-tions of its plethora of biologically active factors, the MSC secretome is capable of establishing a regenerative micro-environment for the repair of injured tissues by both elicit-ing and amplifyelicit-ing biological responses within recipient cells. Compositional analyses of the secretome using mass spectrometry, next-generation sequencing, or lipid profiling have identified cytokines and growth factors with diverse biological properties that have been implicated in a vast array of cellular processes ranging from cell activation and proliferation to differentiation. Components of the secre-tome have also been ascribed key roles in critical aspects of tissue regeneration such as promoting angiogenesis, inhibit-ing apoptosis, immunomodulation, anti-inflammation, and stem cell homing to sites of injury [10–12]. In particular, MSC exosomes were demonstrated to possess therapeutic potential for cartilage repair in osteochondral defects [13,
14] as well as protection against cartilage and bone degrad-ation in in vivo models of osteoarthritis [15,16].
The function of the MSC secretome is activated by local micro-environmental cues that modulate MSC dif-ferentiation as well as that of subsidiary tissues [17,18].
Changes in growth factors/cytokines [19, 20], oxygen
tension [21, 22], or environmental mechanical cues
aris-ing from the extracellular matrix [23, 24] or substrate
stiffness [25, 26] have been shown to directly influence MSC paracrine activity. Scaffolds of distinct composition and cytoarchitecture influence how the cellular
mechan-otransduction machinery translates environmental
mechanical and chemical cues into transcriptional and paracrine responses. Moreover, growing MSCs as 3D spheroids encourages cell-cell interactions that upregu-late paracrine activities with anti-inflammatory and an-giogenic properties [27–29]. Furthermore, diverse forms of mechanical stimulation such as shear stress, tensile stress, and compression have been shown to promote
the production of reactive oxygen species (ROS) [30], as
well as alter the secretome profile of MSCs [31, 32].
Accordingly, activation of mitochondrial respiration is
known to activate the muscular secretome [33].
We have previously shown that pulsed electromagnetic fields (PEMFs) activate calcium-permeable transient re-ceptor potential (TRP) channels, promoting both in vitro chondrogenesis [34] and myogenesis [35] by activating a calcium-mitochondrial transcriptional and epigenetic
axes governing survival and development [35].
PEMF-induced MSC differentiation has been correlated with
increased expression of TGFβand BMP2 [36–38].
More-over, the expression and paracrine action of these growth factors was also demonstrated in electrically
driven MSC chondrogenesis [39]. PEMF exposure has
also been shown to exert anti-inflammatory effects by upregulating A2A and A3ARs, thereby mitigating the ex-pression of pro-inflammatory cytokines [40, 41]. This is supported by studies showing PEMF inhibition of the PGE2 and cycloxigenase-2 (COX-2) pathways, reducing the expression of pro-inflammatory cytokines (6, IL-8) while augmenting anti-inflammatory factors (cAMP, IL-10) in synovial fibroblasts from bovine and
osteoarth-ritic patients [42–44]. Here, we provide evidence that
the previously described pro-chondrogenic of PEMF stimulation [34] can be largely attributed to its
modula-tion of the MSC secretome (Fig. 1). PEMF stimulation
was shown to modulate the chondrogenic, chemotactic, anti-inflammatory, and antiapoptotic activities of the MSC secretome in the form of conditioned medium (CM) harvested from PEMF-exposed MSCs. PEMF stimulation, via its effects over MSC paracrine signaling, might hence represent a manner of promoting regener-ation in an inflammatory joint environment.
Methods
PEMF exposure system
The PEMF device used in this study has been previously described [34, 35]. Briefly, the device produces spatially homogeneous, time-varying magnetic fields, consisting
of barrages of 20 × 150μs on and off pulses for 6 ms
re-peated at a frequency of 15 Hz. The magnetic flux
dens-ity rose to predetermined maximal level within ~ 50μs
(~ 17 T/s) when driving field amplitudes between 0.5 and 4 mT. Unless explicitly noted, all samples were ex-posed once for 10 min. All PEMF-treated samples were compared to time-matched control samples (0 mT) that were manipulated in exactly the same manner as experi-mental samples, including placement into the PEMF-generating apparatus for the designated time, except that the apparatus was not set to generate a magnetic field.
Human bone marrow MSC culture
5% CO2atmosphere. The expanded MSCs were used
be-tween passages 5 and 6.
Conditioned media (CM) from MSC in 3D and 2D culture platforms
Conditioned media (CM) was generated from 3D culture
pellets as previously described [34]. Briefly, MSCs were
expanded in expansion media until 70–80% confluency be-fore being subjected to pellet formation (2.5 × 105cells per pellet) and left in expansion media overnight. The follow-ing day the expansion media was replaced with 0.5 ml of low-glucose DMEM (Life Technologies) media without FBS. PEMF treatment was applied for a duration of 10 min at 1, 2, 3, and 4 mT, or 2 mT for 30 min. CMs from pellet culture were collected 24 h post-PEMF exposure and pooled for later use.
To generate CM from 2D culture platforms, MSCs were cultured in T75 flasks at a seeding density of 1.5–2 × 105
in expansion media. At 50–60% confluency, the expansion media was replaced with 10 ml of low-glucose DMEM media without FBS. PEMF exposure was applied at 1, 2, 3, and 4 mT for 10 min, or 2 mT for 30 min, and the CM generated (PCM) was collected at 24 h post-PEMF expos-ure, pooled, and used for further application and analysis.
Control CM (CCM) from 2D and 3D culture platform was generated from similarly cultured MSC without PEMF treatment (0 mT).
In either culturing platform, CM was collected and subsequently concentrated 10× by high centrifugation force using a protein concentrator with a molecular weight cut-off of 3 kDa (Thermo Fisher Scientific, USA). In subsequent experiments, the concentrated CM was diluted 1:10 with low-glucose DMEM media without FBS prior to application to the experimental culture to achieve a final working strength of 1× CM.
MSC chondrogenesis
Chondrogenic differentiation of MSCs was induced in 3D pellet cultures as previously described [45, 46]. Briefly,
2.5 × 105 cells were centrifuged to form pellets and
cul-tured in a chondrogenic differentiation medium containing high-glucose DMEM supplemented with 4 mM proline,
50μg/mL ascorbic acid, 1% ITS-Premix
(Becton-Dickin-son, San Jose, CA), 1 mM sodium pyruvate, and 10−7M
dexamethasone (Sigma Aldrich, St Louis, MO), for up to 7 or 21 days in the presence of 10 ng/mL of transforming
growth factor-β3 (TGFβ3; R&D Systems, Minneapolis,
chondrogenic differentiation media to avoid interference with TRPC channel gating [34,35]. To study the chondro-genic potential of the MSC secretome, the chondrochondro-genic
media, in the absence or presence of TGFβ3, was
supple-mented with CM.
Chondrocyte redifferentiation
Chondrocytes were isolated from pig (animal not directly involved in this study) articular cartilage following enzym-atic digestion as previously described [47]. Isolated chon-drocytes were expanded in low-glucose DMEM with 10% FBS without antibiotics and used at passage 1 for all the experiments. Chondrogenic redifferentiation was induced in 3D pellet cultures in the presence of 10 ng/mL of TGFβ3. Briefly, 2 × 105chondrocyte cells were centrifuged to form pellets and kept overnight in expansion media at
37 °C in 5% CO2atmosphere. To study the chondrogenic
potential of the MSC secretome for chondrocyte rediffer-entiation, the expansion media was replaced with chon-drogenic media supplemented with the CM and cultured up to 7 or 21 days with medium change every 2–3 days.
Cell migration
The migration of MSCs and chondrocytes in response to CM was assessed using a 24-well Transwell culture
(8μm pore size, Millipore, Germany). Briefly, MSCs (3 ×
104) or chondrocytes (5 × 104) were suspended in 300μl of low-serum culture medium (DMEM supplemented with 0.5% FBS (Life Technologies)) and placed into the upper chamber, and CM was added to the lower cham-bers of the Transwell culture, containing DMEM with 0.5% FBS. After 16 h, the upper surface of the Transwell filters was swabbed to remove cells. Cells on the under-side of the filter, representing the migrated cells, were then fixed in 4% (v/v) paraformaldehyde and stained with hematoxylin and eosin (Sigma Aldrich). The cells in five randomly selected fields at 40× magnification were counted to indicate migrated cells.
Inflammatory induction of MSCs and chondrocytes
MSCs or chondrocytes were plated at 1.5 × 104or 3 × 104
cells/well, respectively, in a 24-well plate in DMEM
con-taining 10% FBS. IL-1β(5 ng/ml; RnD systems) was added
to the expansion media at 24 h after cell seeding to simu-late inflammatory conditions. To investigate the inflamma-tion modulatory effect of MSC secretome, CM was added to the culture 24 h after the induction of inflammation.
MSCs or chondrocytes, induced with IL-1β, without
sub-sequent CM treatment served as inflammation (positive) controls, whereas non-inflammation (negative) controls consisted of MSCs or chondrocytes without the addition
of IL-1β and CM treatments. Cells and media were
harvested 24 and 48 h post-supplementation with CM for
RNA and nitric oxide synthase (NOS) analysis. NOS
activity in the media was analyzed with a NOS assay kit (Abcam, USA). Real-time PCR analysis was performed on the harvested cells to assess the inflammation modulation as a result of the CM.
For post-chondrogenic inflammation induction, MSC
pellets were administered IL-1β(5 ng/ml) for 24 h before
being supplemented with CM. The inflammation modu-latory effect of the CM with reference to MSC-derived chondrogenesis was investigated by real-time PCR ana-lysis at day 7 of differentiation.
Cell proliferation and apoptosis
To assess cell proliferation DNA was analyzed using
Quant-iT™ PicoGreen™ dsDNA Assay Kit (Life
Tech-nologies) over a period of 3 days. For determination of antiapoptotic capacity of CM, MSCs or chondrocytes were seeded at 1.5 × 104or 3 × 104cells/well in a 24-well plate and treated with Staurosporin (200 nM, Sigma Al-drich) for 2 h in the presence of CM. The extent of apoptosis was indicated by Caspase 3/7 activity using a Caspase 3/7 assay kit (Promega, Singapore).
Real-time PCR analysis
Total RNA was extracted using the RNeasy® Mini Kit (Qiagen, Germany). Reverse transcription was performed
with 100 ng total RNA using iScript™cDNA synthesis kit
(Bio-Rad, USA). Real-time PCR was conducted using the SYBR® green assay on ABI Step One Plus Real-Time PCR System (Applied Biosystems, Life Technologies, USA). Real-time PCR program was set at 95 °C for 10 min, followed by 40 cycles of amplifications, consisting of a 15 s denaturation at 95 °C and a 1 min extension step at 60 °C. The human and porcine primer sequences
used in this study are listed in Additional file 1: Table
S1. The level of expression of the target gene,
normal-ized to GAPDH, was then calculated using the 2−ΔΔCt
formula with reference to the undifferentiated MSC. Re-sults were averaged from triplicate samples of two inde-pendent experiments.
ECM and DNA quantification
using a Col 2 standard. Values for sGAG and Col 2 content obtained were normalized to the total DNA content of respective samples, measured using Pico-green dsDNA assay (Molecular Probes, OR, USA). Qua-druplicates of each group were analyzed from two independent experiments.
Secretome analysis
A RayBio fluorescent antibody array (Genomax Tech-nologies, SG) was customized for analyzing the secre-tome of MSC. The CM of 2D cultured MSCs without (0 mT) and with PEMF exposure at 3 mT for 10 min were concentrated 10× using a protein concentrator with a molecular weight cut-off of 3 kDa (Thermo Fisher Scientific, USA). The staining of the arrays was per-formed according to the manufacturer’s protocol. Images were acquired using a GenePix 4000B microarray scan-ner and analyzed with GenePix Pro software (Molecular Devices, USA) for the relative fluorescent intensities of the customized protein targets.
Statistical analysis
All experiments were performed in biological replicates
(n= 3) and results reported as mean ± standard deviation
(SD). Statistical analysis was carried out by Student’s t
test for comparison between two groups using the Microsoft Excel software. The level of significance was set atp< 0.05. All quantitative data reported were aver-aged from at least two independent experiments.
Results
Chondrogenic potential of PEMF-conditioned media generated from MSCs in 3D culture
We have previously shown that MSCs in pellet culture (3D) exhibited an enhancement in chondrogenic induc-tion when exposed to PEMFs at a discrete efficacy window
of 2 mT applied once for 10 min [34]. Here we show that
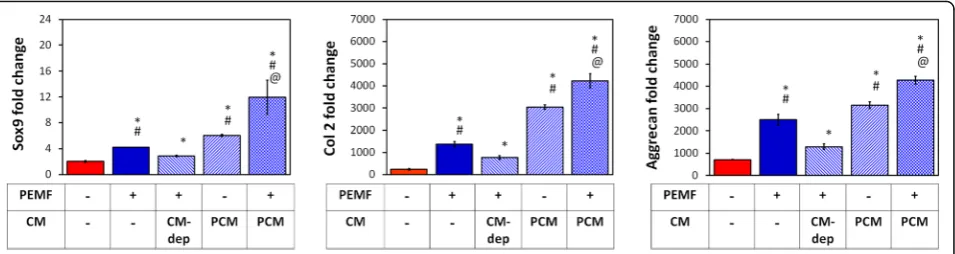
the MSC secretome contributes to the chondrogenic po-tential of PEMF exposure. The characteristic PEMF-induced upregulations of Col 2, Aggrecan, and Sox9 were attenuated by the removal of PCM and its replacement with age-matched media harvested from naïve (CM-de-prived; CM-dep) unexposed sister cultures 24 h following PEMF exposure (Fig.2), serving as a manner to selectively deprive cells of PEMF-mediated paracrine stimulation, while leaving other collateral PEMF-dependent responses intact. On the other hand, PEMF-induced chondrogenic induction could be transferred to naïve MSCs with the transfer of CM harvested from PEMF-treated MSCs (PCM) 24 h after exposure. Moreover, PCM acted syner-gistically with the effects of direct PEMF exposure. The re-sults indicate that the chondrogenic attributes of PEMF exposure is partially mediated through the paracrine activ-ity of the MSC secretome.
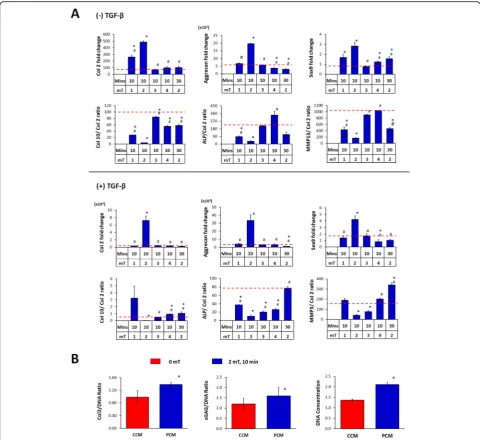
MSC pellet cultures were exposed to PEMFs of varying amplitudes and durations and the generated PCMs were transferred to naïve (unexposed) MSC pellet cultures undergoing chondrogenic induction. PCM generated from MSC pellets exposed at 2 mT for 10 min produced the greatest induction of chondrogenic markers (Col 2,
aggre-can, Sox9) in naïve MSC pellets (Fig. 3a), matching our
previously described EMF efficacy window [34].
Con-versely, the expression ratios of type X collagen (Col 10), alkaline phosphatase (ALP) and matrix metallopeptidase 13 (MMP13) to Col 2, indices of chondrogenic hyper-trophy, were most strongly suppressed by PCM generated at the peak of the efficacy window (2 mT for 10 min).
TGFβ3 is commonly used to facilitate chondrogenic
in-duction [45,46]. Notably, PCM generated at 2 mT for 10 min was capable of promoting chondrogenesis in the ab-sence of TGFβ3 by four to fivefold. In the presence of 10
ng/ml TGFβ3, chondrogenic induction was further
en-hanced by PCM exhibiting increases of > 10-fold for both Col 2 and aggrecan. Sox9 expression levels were more modestly enhanced with PCM generated at 2 mT for 10 min in the absence (~ 3-fold) or presence (~ 4-fold) of
TGFβ. By contrast, PCM harvested from 3D MSC cultures
that were exposed to PEMFs of greater amplitudes or lon-ger durations did not exhibit any significant effects relative to pellets administered control CM (CCM, 0 mT). The in-hibition of chondrogenic hypertrophy by PCM harvested at peak PEMF amplitude (2 mT) was also preserved in the presence of TGFβ3 (Fig.3a).
The chondrogenic enhancement afforded by PCM ad-ministration at the transcriptional level was further cor-roborated at the protein level, whereby the PCM obtained from MSC 3D pellet cultures following exposure to 2 mT PEMFs produced significant increases in both Col 2 and sGAG proteins with an associated increase in cellular
DNA content relative to CCM (Fig.3b).
Chondrogenic potential of PCM generated from MSCs in 2D culture
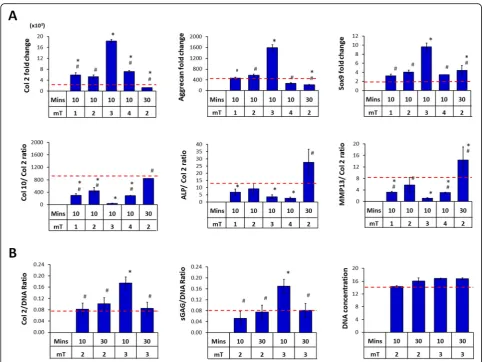
It is well established that the availability and nature of cellular-substrate interactions influence the paracrine activ-ity of MSCs [18, 23, 48]. We next assessed the chondro-genic potential of PCM harvested from MSCs grown in 2D cultures. MSCs cultured on the surface of tissue culture dishes were exposed to PEMF amplitudes ranging from 0 to 4 mT for single 10 min exposures and the generated CMs were then transferred to naïve (unexposed) MSC pel-let cultures undergoing chondrogenic induction in the
presence of TGFβ. PCM harvested from MSCs exposed to
enhancement relative to MSC pellets treated with control
CM (CCM, 0 mT) (Fig. 4a). As with 3D MSC-generated
PCM, downregulations in the expression of the hyper-trophic markers Col 10, ALP, and MMP13 were most
ap-parent with peak PCM (3 mT) compared to CCM (Fig.4a).
The chondrogenic enhancement apparent at the transcrip-tional level with PCM generated from 2D MSCs exposed to 3 mT for 10 min was also corroborated at the protein level, with a twofold increase in Col 2 and sGAG deposition
compared to non-PEMFed CCM (Fig.4b). Accordingly, no
increase in ECM deposition was detected with PCM gener-ated at off peak conditions.
Effects of PCM on chondrocytes
We next investigated the chondrogenic effects of the MSC-PCM on chondrocyte redifferentiation.
Chondro-cytes as pellet cultures in the presence of TGFβ were
treated with PCM generated from MSCs cultured in ei-ther 2D or 3D platforms exposed at their respective peak amplitudes of 3 mT and 2 mT. Both PCMs better sus-tained the chondrogenic phenotype in redifferentiating chondrocytes, relative to their respective CCM. In-creased expression of Col 2 and aggrecan (1.5 to 2-fold) was detected, whereas decreases in the expression of Col 1 and Col 10 were observed, relative to the respective
CCM (Fig.5a). The relative efficacy of either PCM at its
associated peak amplitude and platform on primary chondrocytes was further corroborated at the protein level for both Col 2 (> 3-fold increase) and sGAG (~
2-fold increase) production (Fig. 5b). The results suggest
that PCMs harvested from either 2D or 3D MSC cul-tures at their respective peak amplitudes and exposure durations were capable of offsetting chondrocyte de-differentiation and promote the expression of hyaline cartilage markers.
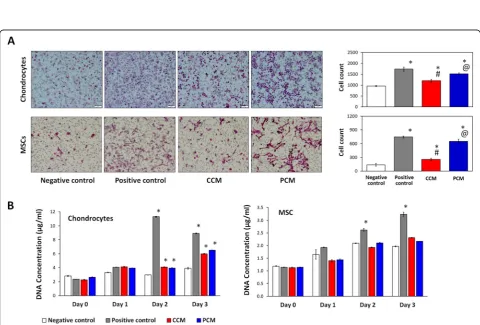
Effect of PCM on chondrocyte and MSC migration and proliferation
We assessed if 2D MSC-generated PCM could affect the migration and proliferation of chondrocytes and MSCs, the two cell types localized in the articular joint environ-ment that participate in cartilage regeneration [8, 49]. CCM and PCM enhanced cell migration of both chon-drocytes and MSCs relative to respective negative
con-trols (Fig. 6a). PCM, however, produced a further
twofold and fourfold increase in migratory capacity for chondrocytes and MSCs, respectively, relative to the CCM, approaching the migration level observed for the positive controls (10% FBS). Chondrocyte proliferation was enhanced to similar magnitudes with either CCM or PCM, whereas MSC proliferation was largely unaffected by either CM under the presented culturing conditions
(Fig.6b). PCM thus appears to hold potential for
enhan-cing MSC-derived paracrine-dependent chemotaxis of MSCs and chondrocytes.
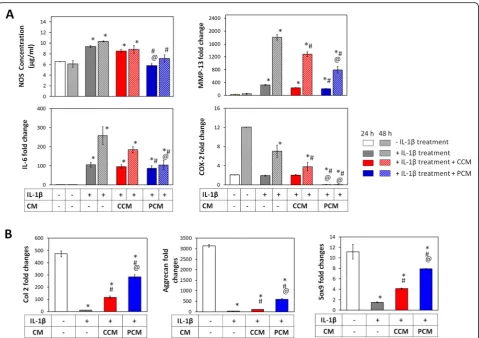
Anti-inflammatory effects of PCM on chondrocytes and MSCs
The MSC secretome has been ascribed
anti-inflammatory properties [14, 29]. We investigated the
potential immunomodulatory attributes of 2D MSC-generated PCM over inflammation-induced
chondro-cytes. Primary chondrocytes treated with IL-1βexhibited
attenuated expression of the ECM markers, Col 2 and aggrecan, coincident with an increased expression of the pro-inflammatory markers, IL-6, MMP13, and COX-2,
as well as enhanced NOS activity (Fig. 7). Although
NOS activity was suppressed by either PCM or CCM
relative to the inflammatory control (IL-1β treatment),
PCM exerted an additional ~ 2–3-fold greater suppres-sion of NOS compared to CCM that was sustained throughout the 48-h examination period. Suppressions Fig. 2Brief PEMF exposure stimulates MSC-derived paracrine activity to promote chondrogenesis. MSCs in 3D pellet cultures undergoing
in the levels of IL-6 (> 1.5- and 2.5-fold), MMP-13 (10-and 13-fold), (10-and COX-2 (> 2.5- (10-and 2-fold) were also ob-served commencing at 24 h after administration of either CCM or PCM, respectively. Notably, chondrocytes exhib-ited elevated basal levels of IL-6 and COX-2 in the non-inflammation controls (no IL-1βtreatment), indicating sig-nificant levels of resting inflammation in chondrocytes. Re-instated expression of chondrogenic markers (Col 2 and aggrecan) was observed following administration of the
PCM at 24 and 48 h relative to the inflammatory control and CCM (Fig.7) that, although were mitigated relative to the non-inflammatory controls, were showing a trend to-wards recovery at 48 h.
Similar to chondrocytes, IL-1βtreatment elicited a state of inflammation in MSCs that was evidenced by upregula-tions in IL-6, MMP-13, COX-2, and nitric oxide synthase
(NOS) (Fig. 8a). By contrast, MSCs were overall more
resistant to developing IL-1β-induced inflammation as
Fig. 3Effect of PEMF-induced conditioned media (CM) from 3D cultured MSCs on chondrogenic differentiation of MSCs. MSCs in 3D culture were subjected to PEMF exposure at different intensities and durations. The generated CM was collected 24 h post-PEMF exposure and tested for chondrogenic effect over naive (unexposed) MSC pellet cultures undergoing chondrogenic differentiation in the absence or presence of TGFβ.a
Real-time PCR analysis of cartilaginous and hypertrophic marker expression after 7 days of differentiation normalized to GAPDH and presented as fold-changes relative to levels in undifferentiated MSCs. The expression of hypertrophic markers was presented as the ratio to Col 2 expression.b
appreciable levels of inflammatory markers were generally only achieved after 48 h. Reductions in MMP-13 and COX-2 were observed with both CCM and PCM adminis-tration at 24 and 48 h post-adminisadminis-tration with overall greater suppression observed with PCM. The levels of COX-2 showed significant reductions at both 24 and 48 h upon administration of PCM, particularly with reference
to non-inflammatory controls (no IL-1βtreatment). IL-6
levels were suppressed most strongly 48 h after adminis-tration of PCM relative to the inflammatory control
(IL-1βtreatment). PCM administration also produced
signifi-cant suppressions of IL-1β-induced NOS activity at both
24 and 48 h, whereas CCM showed no effect at either time point. Notably, the level of NOS activity observed in re-sponse to PCM administration at 48 h was identical to
that in the non-inflammatory control, indicating complete normalization of basal NOS activity.
The effects of PCM over MSC chondrogenesis were also examined under conditions of induced inflammation.
IL-1β treatment suppressed the expression of the
chondro-genic markers, Col 2, aggrecan, and Sox9 (Fig.8b). Partial rescue of Col 2 (> 9-fold and 20-fold), aggrecan (> 3-fold and 15-fold) and Sox9 (> 2.5-fold and 5.5-fold) was achieved in samples supplemented with either CCM or PCM, respectively, compared to the inflammatory controls
(IL-1βtreatment). As noted, PCM consistently produced
greater levels of protection than CCM. PEMF treatment hence was capable of enhancing MSC-derived paracrine factors capable of attenuating cellular inflammation, par-tially reinstating MSC chondrogenesis.
Fig. 5Effects of PEMF-induced conditioned medium (CM) on chondrocyte redifferentiation. CM was generated from MSCs cultured on 2D and 3D platforms with and without PEMF exposure at 3 mT and 2 mT, respectively, or unexposed. CM collected 24 h post-PEMF exposure was administered to chondrocyte pellet cultures.aReal-time PCR analysis of cartilaginous marker expression after 7 days of redifferentiation was normalized to GAPDH and presented as a fold-change relative to the level expressed in day 0 chondrocytes. Col 1 and Col 10 expressions are shown as ratios relative to Col 2 expression.bQuantification of cartilaginous extracellular matrix macromolecules generated by chondrocytes after 21 days of redifferentiation. Data shown represent means ± SD,n= 6 from 2 independent experiments. * denotes significant increases compare to respective non-PEMFed CM (CCM, 0 mT) (red solid bars)
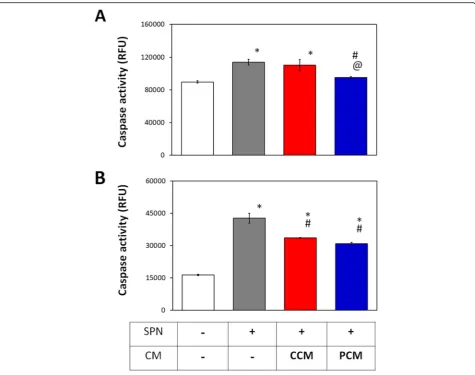
Effect of PCM on chondrocyte and MSC apoptosis
We have previously shown that brief (10 min) exposure to PEMFs was capable of attenuating basal apoptosis in
myoblasts during early myogenesis [35]. We evaluated
the potential of 2D MSC-generated CM to modulate apoptosis in chondrocytes and MSCs subjected to Staur-osporin (200 nM). Basal caspase activity was elevated in control chondrocytes reflecting the elevated resting
in-flammation demonstrated previously (Fig. 7) and could
be further augmented with exposure to Staurosporin. PCM significantly suppressed caspase activity in chon-drocytes after Staurosporin treatment, effectively nor-malizing basal apoptosis levels (Fig. 9a). By contrast, no antiapoptotic effect was observed in chondrocytes ad-ministered CCM relative to the Staurosporin-treated control. MSCs in which apoptosis was induced with Staurosporin and supplemented with either CCM or PCM exhibited a comparable and significant decrease in caspase activity relative to the Staurosporin-treated
control (Fig.9b). PEMF treatment hence potentiates the
paracrine-mediated attenuation of apoptosis of MSC CM in chondrocytes that could ultimately translate to improved regenerative responses.
Expression and secretion of paracrine factors modulating differentiation, proliferation, migration, and inflammation from PEMF-treated MSCs
The ability of PEMFs to influence the production and re-lease of factors associated to chondrogenic differentiation, proliferation, migration, and inflammatory modulation was examined from 2D cultures of MSCs. Factors exam-ined included bone morphogenic protein (BMP), trans-forming growth factor (TGF), thrombospondin (TSP),
insulin-like growth factor (IGF), COX-2, IL-10, and inter-leukin 1 receptor antagonist (IL-1ra). The expression of BMP2, BMP4, TSP-2, IL-1ra, and IL-10 were upregulated by ~ 2-fold, or greater, 24 h after PEMF treatment, relative
to non-exposed MSCs. TGFβ1, TGFβ3, IGF-2, and
COX-2 exhibited less significant increases, whereas TSP-1 showed no change in expression (Fig.10a).
Antibody arrays were used to characterize the secretion profile of the MSC CM. PEMF exposure at 3 mT pro-moted the secretion of BMP-2, BMP-4, TSP-2, and IL-1ra
(Fig. 10b), consistent with observed increases in gene
expression (Fig.10a). By contrast, the protein secretion of
TGFβ1 and IL10 was dampened by PEMF treatment,
while TGFβ3 and TSP-1 secretion was not affected,
reflecting transcriptional to translation temporal disparity.
Discussion
MSCs secrete a plethora of bioactive factors, which through paracrine means, synchronize the regenerative responses of neighboring cellular communities. For clinical exploitation, strategies to selectively modulate the secretory function of MSCs have been advanced and include subjecting MSCs to a variety of micro-environmental biochemical and mechan-ical cues, as well as biophysmechan-ical perturbations [24,25, 50]. We previously showed that MSC chondrogenic differenti-ation in 3D pellet cultures could be enhanced by brief exposure to low amplitude and frequency PEMFs
adminis-tered for 10 min at an optimal amplitude of ~ 2 mT [34].
The involvement of transient receptor potential cation
(TRP) channels and downstream Ca2+ signaling were
myogenesis and underlying mitochondrial adaptation to oxidative stress via a process of magnetic mitohormesis [35]. Accordingly, activation of mitochondrial respiration is known to activate the muscular secretome [33].
In this study, we examined the effects of brief PEMF ex-posure on the MSC secretome and its ability to promote MSC-induced chondrogenesis and chondrocyte cartilage tissue formation. The results demonstrated that the cap-acity of PEMFs to induce MSC chondrogenesis is contrib-uted to by PEMF modulation of MSC secretome. As such, exchange of the bathing medium 24 h post-PEMF expos-ure compromised chondrogenesis, clearly indicating the necessity of released paracrine factors (Fig. 2). Harvested CM from PEMF-treated MSCs (PCM) was moreover cap-able of conferring chondrogenic enhancement onto naïve MSCs and, furthermore, demonstrated synergy when
combined with direct PEMF exposure of MSCs. Notably, the PEMF parameters best suited for the production of chondrogenic MSC-PCM from 3D pellets was 2 mT for 10 min (Fig.3), identical to the exposure parameters previ-ously determined most efficacious at promoting 3D MSC
chondrogenesis in response to direct exposure [34]. PCM
generated under this exposure paradigm rendered signifi-cant chondrogenic enhancement compared to CCM obtained from non-exposed MSCs. Furthermore, the ob-served PCM-mediated increase in the expression of cartil-aginous matrix was accompanied by a downregulation of hypertrophic markers, Col 10, ALP, and MMP13. The pro-chondrogenic effect of PCM was also evident in the con-text of chondrocyte redifferentiation, promoting cartilage
formation with superior hyaline phenotype (Fig.5). Taken
together, our data demonstrates that MSCs subjected to Fig. 8Effect of PEMF-induced conditioned medium (CM) on inflamed MSCs pre- (a) and post- (b) chondrogenic induction.aInflammation was induced in MSCs with 5 ng/ml IL-1βfor 24 h before administration of CCM or PCM generated from 2D MSC cultures. Expression of inflammatory genes and NOS activity was assayed at 24 (plain bars) and 48 h (hatched bars) after CM treatment.bMSCs undergoing chondrogenesis in 3D pellet cultures were treated with 5 ng/ml IL-1β24 h prior to the administration of CCM or PCM. Expression cartilaginous markers was analyzed after 7 days of differentiation, normalized to GAPDH and presented as fold-changes relative to levels in undifferentiated MSCs. Data shown represent means ± SD,n= 6 from 2 independent experiments. * denotes significant differences compared to the non-inflamed controls (no IL-1β treatment), # denotes significant differences compared to the inflammation controls (IL-1βalone treatment), and @ denotes significant differences compared to respective CCM. Col 2 = type II collagen; COX-2 = cycloxigenase-2; IL-1β= interleukin-1β; IL-6 = interleukin-6; MMP-13 =
chondrogenic induction under conditions of 3D culture re-spond to PEMF stimulation with secretome modulation that could provide both autocrine and paracrine anabolic effects with ultimate relevance to the in vivo articular scenario.
Components of the MSC secretome, in the form of CM or as isolated extracellular vesicles (EVs), have dem-onstrated therapeutic potential in cases of osteochondral lesions [13, 14] or in osteoarthritis animal models [15,
16]. The therapeutic effect of the MSC secretome has
been largely attributed to the multifaceted stimulation of chondrocyte proliferation, migration, cartilaginous ECM generation, and the attenuation of the inflammatory and apoptotic microenvironment associated with injury or joint degeneration [14–16, 51, 52]. The demonstration that the delivery of MSC secretory products was effica-cious in cartilage regeneration revealed the possibility of
using MSC-derived secretory products as a cell-free therapeutic for joint injury and osteoarthritis. Notably, CM harvested from MSCs in conventional 2D tissue cul-ture had similar chondrogenic potency as 3D PCM in terms of cartilage ECM formation and the capacity to re-duce hypertrophic and fibrocartilage development for both MSCs and chondrocytes, although responsive to a distinct, yet higher (3 mT for 10 min), electromagnetic
efficacy window (Figs. 4 and 5). Chondrocyte migration
had been previously reported to be stimulated by
MSC-derived EVs [14, 51]. Here we show that migration was
also enhanced for both chondrocytes and MSCs by PCM
(Fig.6), suggesting the potential for PCM to
chemotacti-cally attract chondrocytes or MSCs to the vicinity of an articular injury to promote regeneration.
(e.g., matrix metalloproteinases) are upregulated, perpetu-ating inflammation, cartilage matrix degradation, and
chondrocyte apoptosis [53]. MSC-derived EVs have been
previously demonstrated to inhibit the adverse effects of inflammatory cytokines on cartilage homeostasis [14–16,
51, 52]. Here we show that the MSC-derived CMs are
generally protective against cellular inflammation reflected by suppressions of chondrocyte NOS activity, inflamma-tory markers (IL-6 and COX-2), and catabolic proteinases
(MMP13) (Fig. 7). Significantly, the protective impact of
PCM generated from PEMF-exposed MSCs was even greater as revealed by stronger suppressions of NOS activ-ity, preservation of chondrocyte anabolic activities, indi-cated by upregulating severely suppressed cartilage ECM
genes expression (Fig. 7), and rescuing the
Staurosporin-induced chondrocytes apoptosis (Fig.9), an effect not ob-served with non-PEMFed control CM (CCM).
The protection conferred by PCM may hence poten-tially be extended to an inflamed scenario within the ar-ticular environment. Resident MSCs in the joint cavity, arising from either the synovium or bone marrow, in the case of microdrilling/microfracture, have been shown to participate in cartilage regeneration, either by engraft-ment or as endogenous providers of secreted trophic
factors [54, 55]. The regenerative efficacy of resident
MSCs, however, would be influenced by the inflamma-tory environment within the joint. Under conditions of
induced inflammation, MSCs experienced increased NOS activity, elevated expressions of IL-6, Cox-2, and MMP13, and attenuated chondrogenic differentiation, degenerative conditions that were effectively suppressed
with PCM (Fig. 8) and could potentially be translated to
the in vivo scenario. Our results indicate that the brief exposure of MSCs to low-amplitude PEMFs heightens the anti-inflammatory potential of their secretome, al-luding to an enhanced therapeutic application to attenu-ate cartilage damage and restore MSC regenerative capacity in an inflamed articular environment.
MSCs secrete trophic factors including IGF-1, PDGF,
FGF, VEGF, and members of the TGFβ superfamily in
response to environmental cues or stimuli [18, 19].
Ac-cordingly, PEMF-based enhancement of MSC differenti-ation has been previously linked with the expression and
secretion of the TGF and BMP families [36–38].
More-over, the capacity of PCM to significantly augment car-tilaginous ECM production, despite the conventional
presence of a relatively high dose of exogenous TGFβ3
(10 ng/ml), alludes to the presence of additional factor(s) necessary for chondrogenic induction. Accordingly, we did not detect significant differences in the expression of
TGFβ between PEMFed and non-PEMFed MSCs
(Fig.10) and moreover, TGFβ1 secretion was instead
re-duced in the PCM. By contrast, PEMF exposure of
MSCs produced ≥2-fold increases in the expressions of
Fig. 10PEMF exposure modulates the secretion of MSC-derived paracrine factors.a2D cultures of MSCs without (0 mT) and with PEMF exposure at 3 mT for 10 min were subjected to real-time PCR analysis after 24 h. Data shown represents means ± SD,n= 6 from 3 independent
BMP2, BMP4, TSP2, and IL1ra that were further corrob-orated at the protein level using antibody arrays. BMP2 [56], BMP4 [57], and TSP2 [58–60] have been previously implicated in chondrogenic induction. Indeed, autocrine action of TSP-2 from umbilical cord MSCs has been shown to suppress hypertrophic phenotype development [60] and could possibly account for our observed suppres-sion of MSC and chondrocyte hypertrophy in response to administration of PCM. BMP2 and BMP4, on the other
hand, are implicated as chemotactic agents for MSCs [61]
and could be responsible for the enhanced migratory effect we observed exerted by PCM on chondrocytes and MSCs. Finally, IL-1ra is the biological inhibitor of IL-1β, associated with the severity of diseased cartilage degeneration [62] and has been demonstrated to reduce cartilage catabolism [63]. Nonetheless, the screening and confirmation of candidate factors participating in the chondrogenic enhancement ob-served with PCM is far from complete. In addition, the pos-sible role of exosomal CD73-mediated adenosine activation of AKT, ERK, and AMPK signaling [14,64] and the
partici-pation of exosomal microRNAs [65, 66] that have been
previously implicated in regulating chondrocyte anabolic ac-tivity and cartilage degradation have yet to be examined. Further work will be required to fully decipher the paracrine mechanisms responsible for the enhanced anabolic and pro-tective effect observed here in response to PCM administra-tion, including the examination of the contribution of EVs to our reported PEMF-mediated secretome responses.
PEMF stimulation has been previously shown by others to exert anti-inflammatory effects by activating adenosine receptors [41,44], while modulating intracellular calcium
and activating the mechanotransduction FAK/Rho
GTPases signaling pathways to induce MSC migration
[67]. PEMF exposure was also shown to activate a
calcium-mitochondrial axis stimulating mitochondrial respiration and promoting both mitochondriogenesis and myogenesis as well as reducing basal apoptosis and increasing telomere length via a process of adaptive
mag-netic mitohormesis [35]. Uniting these seemingly
dispar-ate responses is the finding that mechanical stimulation
enhances mitochondrial ROS formation [30]. Accordingly,
the generation of mitochondrial ROS has been shown to stimulate the secretome activity and is required for the de-velopment of the cellular and organismal adaptations
against cellular inflammation [33]. It thus appears that
mechanotransduction and mitochondrial respiration act upstream of secretome-mediated anti-inflammatory re-sponses and appear to be modulated by magnetic fields as reported here. Subsequent studies will examine the mito-chondrial contributions to the MSC secretome modula-tion induced by PEMF exposure.
We have previously demonstrated cell type-specific elec-tromagnetic efficacy windows for PEMF-induced cytotox-icity of breast cancer cells [68], myogenic induction [35], or
MSC-based chondrogenic induction [34]. Here, we extend
the biophysical criteria defining the electromagnetic efficacy window by showing that the culturing microenvironment could modulate magnetic sensitivity of MSCs and down-stream secretome activity. 2D and 3D MSC culture plat-forms exhibited distinct sensitivities to PEMF stimulation; a predominance of cell-cell interactions in the 3D configur-ation gave rise to higher magnetic sensitivity with lower ac-tivation window. The optimum PEMF amplitude to produce PCM with enhanced biological efficacy from 2D and 3D MSC cultures was 3 mT and 2 mT, respectively. MSCs grown on 2D TCP adopted fibroblastic morpholo-gies and exhibited a predominance of cell-substrate interactions, whereas MSCs in 3D exhibited rounded morphologies and were largely dominated by cell-cell con-tacts. These distinct mechano-environmental scenarios likely elicited different mechanotransduction responses, which in turn converged with, or differently conditioned, the cellular response to magnetic field stimulation. For in-stance, cellular aggregation has been shown to trigger cadherin-related cell-cell interactions, which in itself aug-ments MSC-dependent wound healing, myogenic, anti-inflammatory, and angiogenic responses [27–29, 48]. It is thus possible that altering the cellular mechanical environ-ment would influence MSC secretome composition and function as well as response and sensitivity to magnetic field exposure. Despite differences in PEMF sensitivities, similar levels of chondrogenic outcome and cartilage for-mation resulted from either 2D or 3D PCM administration (Figs. 3, 4, and 5). It remains to be resolved, however, whether any differences exist in the exact secretome profile of the PCM (or CCM) harvested from either platform.
Conclusions
We provide evidence that brief exposure to low ampli-tude PEMFs enhanced the ability of MSCs to produce and secrete paracrine factors capable of promoting car-tilage regeneration as well as protecting against adverse inflammatory conditions. Furthermore, this report high-lights the importance of optimizing PEMF exposure pa-rameters for MSCs subjected to different culturing conditions. Collectively, our results indicate that PEMF stimulation could augment the production and release of the MSC paracrine repertoire for the ultimate enhance-ment of cartilage regeneration.
Additional file
Additional file 1:Table S1.A) Human PCR primer sequences. B) Porcine primer sequence (DOCX 13 kb)
Abbreviations
2: Type II Collagen; Col 1: Type I Collagen; Col 10: Type X Collagen; COX-2: Cycloxigenase-2; ECM: Extracellular matrix; EVs: Extracellular vesicles; IGF: Insulin-like growth factor; IL-1β: Interleukin-1beta; 6: Interleukin 6; IL-10: Interleukin 10; IL-1ra: Interleukin 1 receptor antagonist;
MSC: Mesenchymal stem cell; MMP13: Metalloproteinase 13; NOS:Nitric oxide synthase; PCM: PEMFed generated CM; PEMF: Pulsed electromagnetic field; PGE2: Prostaglandin E2; PCR: Polymerase chain reaction; sGAG: Sulfated glycosaminoglycan; Sox9: SRY-Box 9; TGF: Transforming growth factor; TRP: Transient receptor potential; TSP: Thrombospondin
Acknowledgements
The authors acknowledge Zac Goh (iHealthtech, National University of Singapore) for the graphical abstract shown in Fig.1.
Funding
The study was supported by National Medical Research Council of Singapore (NMRC/CIRG /1403/2014) and the Lee Foundation of Singapore. Dinesh Parate was supported by NUS Research scholarship and NUS Department of Surgery Funds.
Availability of data and materials
The authors declare that the data supporting the findings of this study are available within the article and its supplementary information files.
Authors’contributions
DP conducted the bulk experimentation and analyzed the data as well as contributed to the writing of the manuscript. NDK and CC executed some of the experiments, participated in data acquisition and analysis. AF-O and EHL acquired funding and edited the manuscript. JHPH edited the manuscript. AF-O provided the PEMF device and expertise, conceptualized and designed the study, and participated in manuscript writing. ZY designed the study, interpreted the data, and wrote the manuscript. All authors read and ap-proved the final manuscript.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
AF-O is an inventor on patent WO 2019/17863 A1, System and Method for Applying Pulsed Electromagnetic Fields, and contributes to QuantumTx Pte. Ltd., which elaborates on the use of similar magnetic fields for human use. All other authors declare that they have no competing interests.
Author details
1Department of Surgery, National University of Singapore, Singapore 119228, Singapore.2Biolonic Currents Electromagnetic Pulsing Systems Laboratory, BICEPS, National University of Singapore, Singapore, Singapore.3Department of Orthopaedic Surgery, Yong Loo Lin School of Medicine, National University of Singapore, NUHS Tower Block, Level 11, 1E Kent Ridge Road, Singapore 119288, Singapore.4Tissue Engineering Program, Life Sciences Institute, National University of Singapore, DSO (Kent Ridge) Building, #04-01, 27 Medical Drive, Singapore 117510, Singapore.5Institute for Health Innovation & Technology, iHealthtech, National University of Singapore, Singapore, Singapore.
Received: 19 November 2019 Revised: 15 January 2020 Accepted: 20 January 2020
References
1. Marcacci M, Filardo G, Kon E. Treatment of cartilage lesions: what works and why? Injury. 2013;44(Suppl 1):S11–5.
2. Loeser RF, Goldring SR, Scanzello CR, Goldring MB. Osteoarthritis: a disease of the joint as an organ. Arthritis Rheum. 2012;64(6):1697–707.
3. Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, et al. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284(5411):143–7.
4. Toh WS, Brittberg M, Farr J, Foldager CB, Gomoll AH, Hui JH, et al. Cellular senescence in aging and osteoarthritis. Acta Orthop. 2016;87(sup363):6–14.
5. Somoza RA, Welter JF, Correa D, Caplan AI. Chondrogenic differentiation of mesenchymal stem cells: challenges and unfulfilled expectations. Tissue Eng Part B Rev. 2014;20(6):596–608.
6. Wu L, Leijten JC, Georgi N, Post JN, van Blitterswijk CA, Karperien M. Trophic effects of mesenchymal stem cells increase chondrocyte proliferation and matrix formation. Tissue Eng Part A. 2011;17(9–10):1425–36.
7. Levorson EJ, Santoro M, Kasper FK, Mikos AG. Direct and indirect co-culture of chondrocytes and mesenchymal stem cells for the generation of polymer/ extracellular matrix hybrid constructs. Acta Biomater. 2014;10(5):1824–35. 8. Maumus M, Jorgensen C, Noel D. Mesenchymal stem cells in regenerative
medicine applied to rheumatic diseases: role of secretome and exosomes. Biochimie. 2013;95(12):2229–34.
9. Ha CW, Park YB, Kim SH, Lee HJ. Intra-articular mesenchymal stem cells in osteoarthritis of the knee: a systematic review of clinical outcomes and evidence of cartilage repair. Arthroscopy. 2019;35(1):277–88. e2 10. Kupcova SH. Proteomic techniques for characterisation of mesenchymal
stem cell secretome. Biochimie. 2013;95(12):2196–211.
11. Van Linthout S, Stamm C, Schultheiss HP, Tschope C. Mesenchymal stem cells and inflammatory cardiomyopathy: cardiac homing and beyond. Cardiol Res Pract. 2011;2011:757154.
12. Oh JY, Lee RH, Yu JM, Ko JH, Lee HJ, Ko AY, et al. Intravenous mesenchymal stem cells prevented rejection of allogeneic corneal transplants by aborting the early inflammatory response. Mol Ther. 2012;20(11):2143–52.
13. Zhang S, Chu WC, Lai RC, Lim SK, Hui JH, Toh WS. Exosomes derived from human embryonic mesenchymal stem cells promote osteochondral regeneration. Osteoarthr Cartil. 2016;24(12):2135–40.
14. Zhang S, Chuah SJ, Lai RC, Hui JHP, Lim SK, Toh WS. MSC exosomes mediate cartilage repair by enhancing proliferation, attenuating apoptosis and modulating immune reactivity. Biomaterials. 2018;156:16–27. 15. Wang Y, Yu D, Liu Z, Zhou F, Dai J, Wu B, et al. Exosomes from embryonic
mesenchymal stem cells alleviate osteoarthritis through balancing synthesis and degradation of cartilage extracellular matrix. Stem Cell Res Ther. 2017;8(1):189. 16. Cosenza S, Ruiz M, Toupet K, Jorgensen C, Noel D. Mesenchymal stem cells
derived exosomes and microparticles protect cartilage and bone from degradation in osteoarthritis. Sci Rep. 2017;7(1):16214.
17. Murphy MB, Moncivais K, Caplan AI. Mesenchymal stem cells: environmentally responsive therapeutics for regenerative medicine. Exp Mol Med. 2013;45:e54. 18. Kusuma GD, Carthew J, Lim R, Frith JE. Effect of the microenvironment on
mesenchymal stem cell paracrine signaling: opportunities to engineer the therapeutic effect. Stem Cells Dev. 2017;26(9):617–31.
19. Rodriguez TM, Saldias A, Irigo M, Zamora JV, Perone MJ, Dewey RA. Effect of TGF-beta1 stimulation on the secretome of human adipose-derived Mesenchymal stromal cells. Stem Cells Transl Med. 2015;4(8):894–8. 20. Li C, Li G, Liu M, Zhou T, Zhou H. Paracrine effect of inflammatory
cytokine-activated bone marrow mesenchymal stem cells and its role in osteoblast function. J Biosci Bioeng. 2016;121(2):213–9.
21. Teixeira FG, Panchalingam KM, Anjo SI, Manadas B, Pereira R, Sousa N, et al. Do hypoxia/normoxia culturing conditions change the neuroregulatory profile of Wharton jelly mesenchymal stem cell secretome? Stem Cell Res Ther. 2015;6:133.
22. Paquet J, Deschepper M, Moya A, Logeart-Avramoglou D, Boisson-Vidal C, Petite H. Oxygen tension regulates human mesenchymal stem cell paracrine functions. Stem Cells Transl Med. 2015;4(7):809–21. 23. Sullivan KE, Quinn KP, Tang KM, Georgakoudi I, Black LD 3rd. Extracellular
matrix remodeling following myocardial infarction influences the therapeutic potential of mesenchymal stem cells. Stem Cell Res Ther. 2014;5(1):14. 24. Jose S, Hughbanks ML, Binder BY, Ingavle GC, Leach JK. Enhanced trophic
factor secretion by mesenchymal stem/stromal cells with glycine-histidine-lysine (GHK)-modified alginate hydrogels. Acta Biomater. 2014;10(5):1955–64. 25. Seib FP, Prewitz M, Werner C, Bornhauser M. Matrix elasticity regulates the
secretory profile of human bone marrow-derived multipotent mesenchymal stromal cells (MSCs). Biochem Biophys Res Commun. 2009;389(4):663–7. 26. Yang H, Nguyen KT, Leong DT, Tan NS, Tay CY. Soft material approach to
induce oxidative stress in mesenchymal stem cells for functional tissue repair. ACS Appl Mater Interfaces. 2016;8(40):26591–9.
27. Santos JM, Camoes SP, Filipe E, Cipriano M, Barcia RN, Filipe M, et al. Three-dimensional spheroid cell culture of umbilical cord tissue-derived mesenchymal stromal cells leads to enhanced paracrine induction of wound healing. Stem Cell Res Ther. 2015;6:90.
stem cells derived from human umbilical cord blood for myocardial infarction. Mol Ther. 2012;20(7):1424–33.
29. Bartosh TJ, Ylostalo JH, Mohammadipoor A, Bazhanov N, Coble K, Claypool K, et al. Aggregation of human mesenchymal stromal cells (MSCs) into 3D spheroids enhances their antiinflammatory properties. Proc Natl Acad Sci U S A. 2010;107(31):13724–9.
30. Chatterjee S, Fisher AB. Mechanotransduction: forces, sensors, and redox signaling. Antioxid Redox Signal. 2014;20(6):868–71.
31. Kasper G, Dankert N, Tuischer J, Hoeft M, Gaber T, Glaeser JD, et al. Mesenchymal stem cells regulate angiogenesis according to their mechanical environment. Stem Cells. 2007;25(4):903–10. 32. Gardner OF, Fahy N, Alini M, Stoddart MJ. Differences in human
mesenchymal stem cell secretomes during chondrogenic induction. Eur Cell Mater. 2016;31:221–35.
33. Scheele C, Nielsen S, Pedersen BK. ROS and myokines promote muscle adaptation to exercise. Trends Endocrinol Metab. 2009;20(3):95–9. 34. Parate D, Franco-Obregon A, Frohlich J, Beyer C, Abbas AA, Kamarul T, et al.
Enhancement of mesenchymal stem cell chondrogenesis with short-term low intensity pulsed electromagnetic fields. Sci Rep. 2017;7(1):9421. 35. Yap JL, Tai YK, Fröhlich J, Fong CHH, Yin JN, Foo ZL, Ramanan S, Beyer C,
Toh SJ, Casarosa M, Bharathy N, Kala MP, Egli M, Taneja, Lee CN, Franco-Obregón A. Ambient and supplemental magnetic fields promote myogenesis via a TRPC1-mitochondrial axis: evidence of a magnetic mitohormetic mechanism. FASEB J. 2019;33(11):12853–12872.https://doi. org/10.1096/fj.201900057R.
36. Aaron RK, Wang S, Ciombor DM. Upregulation of basal TGFbeta1 levels by EMF coincident with chondrogenesis--implications for skeletal repair and tissue engineering. J Orthop Res. 2002;20(2):233–40.
37. Jansen JH, van der Jagt OP, Punt BJ, Verhaar JA, van Leeuwen JP, Weinans H, et al. Stimulation of osteogenic differentiation in human osteoprogenitor cells by pulsed electromagnetic fields: an in vitro study. BMC Musculoskelet Disord. 2010;11:188.
38. Amin HD, Brady MA, St-Pierre JP, Stevens MM, Overby DR, Ethier CR. Stimulation of chondrogenic differentiation of adult human bone marrow-derived stromal cells by a moderate-strength static magnetic field. Tissue Eng Part A. 2014;20(11–12):1612–20.
39. Kwon HJ, Lee GS, Chun H. Electrical stimulation drives chondrogenesis of mesenchymal stem cells in the absence of exogenous growth factors. Sci Rep. 2016;6:39302.
40. Varani K, Padovan M, Vincenzi F, Targa M, Trotta F, Govoni M, et al. A2A and A3 adenosine receptor expression in rheumatoid arthritis: upregulation, inverse correlation with disease activity score and suppression of inflammatory cytokine and metalloproteinase release. Arthritis Res Ther. 2011;13(6):R197. 41. Varani K, Vincenzi F, Ravani A, Pasquini S, Merighi S, Gessi S, et al. Adenosine
receptors as a biological pathway for the anti-inflammatory and beneficial effects of low frequency low energy pulsed electromagnetic fields. Mediat Inflamm. 2017;2017:2740963.
42. De Mattei M, Varani K, Masieri FF, Pellati A, Ongaro A, Fini M, et al. Adenosine analogs and electromagnetic fields inhibit prostaglandin E2 release in bovine synovial fibroblasts. Osteoarthr Cartil. 2009;17(2):252–62. 43. Ongaro A, Pellati A, Setti S, Masieri FF, Aquila G, Fini M, et al. Electromagnetic
fields counteract IL-1beta activity during chondrogenesis of bovine mesenchymal stem cells. J Tissue Eng Regen Med. 2015;9(12):E229–38. 44. Ongaro A, Varani K, Masieri FF, Pellati A, Massari L, Cadossi R, et al.
Electromagnetic fields (EMFs) and adenosine receptors modulate prostaglandin E (2) and cytokine release in human osteoarthritic synovial fibroblasts. J Cell Physiol. 2012;227(6):2461–9.
45. Raghothaman D, Leong MF, Lim TC, Toh JK, Wan AC, Yang Z, et al. Engineering cell matrix interactions in assembled polyelectrolyte fiber hydrogels for mesenchymal stem cell chondrogenesis. Biomaterials. 2014; 35(9):2607–16.
46. Yang Z, Wu Y, Li C, Zhang T, Zou Y, Hui JH, et al. Improved mesenchymal stem cells attachment and in vitro cartilage tissue formation on chitosan-modified poly(L-lactide-co-epsilon-caprolactone) scaffold. Tissue Eng Part A. 2012;18(3–4):242–51.
47. Yin L, Wu Y, Yang Z, Denslin V, Ren X, Tee CA, et al. Characterization and application of size-sorted zonal chondrocytes for articular cartilage regeneration. Biomaterials. 2018;165:66–78.
48. Qazi TH, Mooney DJ, Duda GN, Geissler S. Biomaterials that promote cell-cell interactions enhance the paracrine function of MSCs. Biomaterials. 2017; 140:103–14.
49. Goldring MB, Tsuchimochi K, Ijiri K. The control of chondrogenesis. J Cell Biochem. 2006;97(1):33–44.
50. Li Z, Kupcsik L, Yao SJ, Alini M, Stoddart MJ. Mechanical load modulates chondrogenesis of human mesenchymal stem cells through the TGF-beta pathway. J Cell Mol Med. 2010;14(6A):1338–46.
51. Zhu Y, Wang Y, Zhao B, Niu X, Hu B, Li Q, et al. Comparison of exosomes secreted by induced pluripotent stem cell-derived mesenchymal stem cells and synovial membrane-derived mesenchymal stem cells for the treatment of osteoarthritis. Stem Cell Res Ther. 2017;8(1):64.
52. Vonk LA, van Dooremalen SFJ, Liv N, Klumperman J, Coffer PJ, Saris DBF, et al. Mesenchymal stromal/stem cell-derived extracellular vesicles promote human cartilage regeneration in vitro. Theranostics. 2018;8(4):906–20. 53. Glyn-Jones S, Palmer AJ, Agricola R, Price AJ, Vincent TL, Weinans H, et al.
Osteoarthritis. Lancet. 2015;386(9991):376–87.
54. Hunziker EB, Rosenberg LC. Repair of partial-thickness defects in articular cartilage: cell recruitment from the synovial membrane. J Bone Joint Surg Am. 1996;78(5):721–33.
55. Zhang W, Chen J, Tao J, Jiang Y, Hu C, Huang L, et al. The use of type 1 collagen scaffold containing stromal cell-derived factor-1 to create a matrix environment conducive to partial-thickness cartilage defects repair. Biomaterials. 2013;34(3):713–23.
56. Schmitt B, Ringe J, Haupl T, Notter M, Manz R, Burmester GR, et al. BMP2 initiates chondrogenic lineage development of adult human mesenchymal stem cells in high-density culture. Differentiation. 2003;71(9–10):567–77. 57. Jiang Y, Chen LK, Zhu DC, Zhang GR, Guo C, Qi YY, et al. The inductive
effect of bone morphogenetic protein-4 on chondral-lineage differentiation and in situ cartilage repair. Tissue Eng Part A. 2010;16(5):1621–32. 58. Taylor DK, Meganck JA, Terkhorn S, Rajani R, Naik A, O'Keefe RJ, et al.
Thrombospondin-2 influences the proportion of cartilage and bone during fracture healing. J Bone Miner Res. 2009;24(6):1043–54.
59. Jeong SY, Kim DH, Ha J, Jin HJ, Kwon SJ, Chang JW, et al. Thrombospondin-2 secreted by human umbilical cord blood-derived mesenchymal stem cells promotes chondrogenic differentiation. Stem Cells. 2013;31(10):2136–48. 60. Jeong SY, Ha J, Lee M, Jin HJ, Kim DH, Choi SJ, et al. Autocrine action of thrombospondin-2 determines the chondrogenic differentiation potential and suppresses hypertrophic maturation of human umbilical cord blood-derived mesenchymal stem cells. Stem Cells. 2015;33(11):3291–303. 61. Fiedler J, Roderer G, Gunther KP, Brenner RE. BMP-2, BMP-4, and PDGF-bb
stimulate chemotactic migration of primary human mesenchymal progenitor cells. J Cell Biochem. 2002;87(3):305–12.
62. Lamacchia C, Rodriguez E, Palmer G, Vigne S, Martin P, Talabot-Ayer D, et al. Articular inflammation is controlled by myeloid cell-derived interleukin 1 receptor antagonist during the acute phase of arthritis in mice. Ann Rheum Dis. 2012;71(2):281–7.
63. Elsaid KA, Zhang L, Shaman Z, Patel C, Schmidt TA, Jay GD. The impact of early intra-articular administration of interleukin-1 receptor antagonist on lubricin metabolism and cartilage degeneration in an anterior cruciate ligament transection model. Osteoarthr Cartil. 2015;23(1):114–21. 64. Zhang S, Teo KYW, Chuah SJ, Lai RC, Lim SK, Toh WS. MSC exosomes alleviate
temporomandibular joint osteoarthritis by attenuating inflammation and restoring matrix homeostasis. Biomaterials. 2019;200:35–47.
65. Wu J, Kuang L, Chen C, Yang J, Zeng WN, Li T, et al. miR-100-5p-abundant exosomes derived from infrapatellar fat pad MSCs protect articular cartilage and ameliorate gait abnormalities via inhibition of mTOR in osteoarthritis. Biomaterials. 2019;206:87–100.
66. Mao G, Hu S, Zhang Z, Wu P, Zhao X, Lin R, et al. Exosomal miR-95-5p regulates chondrogenesis and cartilage degradation via histone deacetylase 2/8. J Cell Mol Med. 2018;22(11):5354–66.
67. Zhang Y, Yan J, Xu H, Yang Y, Li W, Wu H, et al. Extremely low frequency electromagnetic fields promote mesenchymal stem cell migration by increasing intracellular Ca (2+) and activating the FAK/Rho GTPases signaling pathways in vitro. Stem Cell Res Ther. 2018;9(1):143. 68. Crocetti S, Beyer C, Schade G, Egli M, Frohlich J, Franco-Obregon A. Low
intensity and frequency pulsed electromagnetic fields selectively impair breast cancer cell viability. PLoS One. 2013;8(9):e72944.
Publisher’s Note