OBERMIER, DALTON RICHARD. The Effect of Functional Teat Number on Reproductive Throughput (Under the direction of Dr. Mark Knauer).
Genetic selection for improving litter size without concurrently increasing the number of
functional teats in a linear fashion has resulted in a piglet nutrient access shortage causing
reduced piglet survival. Number of piglets born alive is often greater than the number of
functional teats in modern maternal lines. This increases competition between piglets and results
in nutrient intake shortages. Therefore, the objective of this study was to evaluate the effect of
functional teat number on reproductive throughput in swine. Teat traits recorded at farrowing
included total teat number (TT), functional teat number (FT), non-functional teat number (NFT),
and damaged teat number (DT); whereas functional mammary glands (FG) was measured at
weaning on 814 Landrace Large White F1 females. Sows and litters were evaluated on two levels: biological dam (their own progeny) and wean dam (all piglets nursing sow after
cross-fostering). Biological dam means for TT, FT, NFT, DT, and FG were 14.43, 13.96, 0.42, 0.03,
and 10.70, respectively. Biological dam production traits included litter size at weaning (LSW),
average piglet weaning weight (WW), litter weaning weight (LWW), and piglet survival
(LSW/TNB) (SURV). Variance components were estimated using gibbs sampler. Biological dam
genetic correlations between TT with FT, NFT, FG, LSW, WW, LWW, and SURV were 0.79,
0.49, 0.16, 0.11, 0.37, 0.38, and -0.19, respectively. Biological dam genetic correlations between
FT with NFT, FG, LSW, WW, LWW, and SURV were 0.09, 0.28, 0.39, 0.44, 0.49, and 0.35,
respectively. An addition of one TT changed LSW, LWW, and SURV by 0.13 piglets, 0.36 kg,
and 0.41%, respectively. An increase in one FT changed LSW, LWW, and SURV by 0.32
piglets, 1.28 kg, and 1.63%, respectively. Heritability estimates for TT and FT were 0.24 and
teat number. Results suggest that udder morphology, in particular functional teat number, plays a
significant role in piglet survival. Genetic selection to increase functional teat number should be
© Copyright 2020 by Dalton Richard Obermier
by
Dalton Richard Obermier
A thesis submitted to the Graduate Faculty of North Carolina State University
in partial fulfillment of the requirements for the degree of
Master of Science
Animal Science
Raleigh, North Carolina 2020
APPROVED BY:
_______________________________ _______________________________ Mark Knauer Todd See
Committee Chair
_______________________________ _______________________________ Kent Gray Jeremy Howard
ii DEDICATION
iii BIOGRAPHY
Dalton Richard Obermier was born and raised in York, Nebraska. He grew up showing
horses, cattle and pigs through 4H and FFA programs. Later in his show career, he developed a
strong passion for pigs and started raising and selling some of his own to other showman across
the state. Being raised in an avid Cornhusker household and wanting to further his show pig
business, he found himself at the University of Nebraska studying animal science during his
undergraduate education.
During his time as an undergraduate, he was a member of the Livestock Evaluation Team
and worked with Dr. Benny Mote as an undergraduate research assistant. Those experiences with
Dr. Mote opened his demographic to swine genetics and the many opportunities graduate school
has to offer. After graduating with a bachelor’s degree in animal science from UNL in 2018, he
completed an internship with Gestal where he conducted a research trial focusing on sow and
piglet performance on litters fed with their electronic lactation feeders; as well as looking at
piglet mortality while using hydraulic lift crates on a sow farm in Ontario, Canada.
In 2018, he decided to further his education in swine genetics at North Carolina State
University under the direction of Dr. Mark Knauer; as well as coach the NCSU Livestock
Judging Team. During the summer of 2019, he interned at Smithfield Premium Genetics and
iv ACKNOWLEDGMENTS
The author is much appreciative of Smithfield Premium Genetics for funding this project.
He also wants to give a special thanks to Dr. Kent Gray, Dr. Yijian Huang, Dr. Jan Marie
Bartholomew, and Dr. Jeremy Howard for sharing their expertise in research protocol and
statistical analysis. He wants to acknowledge farm manager Perry Sutton and his entire farm staff
v TABLE OF CONTENTS
LIST OF TABLES ... vi
LIST OF FIGURES ... vii
CHAPTER 1: Impact of nutrient access on piglet survival ... 1
Introduction ... 2
Section 1. Nutrient access ... 3
1.1. Piglet vitality ... 3
1.2. Piglet birthweight ... 4
1.3. Functional teat number ... 6
1.3.1. Udder morphology ... 6
1.3.2. Importance ... 7
1.3.3. Genetics... 8
1.4. Colostrum yield ... 11
1.5. Milk yield ... 14
1.6. Conclusion ... 16
Section 2. Genetic analysis ... 17
2.1. Genetic analysis ... 17
References ... 20
CHAPTER 2: The effect of functional teat number on reproductive throughput ... 27
Abstract ... 28
Introduction ... 30
Materials and methods ... 31
Animals ... 31
Data collection ... 31
Data editing ... 33
Statistical analysis ... 33
Results ... 34
Summary statistics ... 34
Variance Component Estimates ... 36
Regression Estimates ... 37
Discussion ... 38
Conclusion ... 45
References ... 48
vi LIST OF TABLES
Table 1 Descriptive statistics of teat traits at the time of farrowing and weaning,
production traits and piglet performance data ... 57
Table 2 LSMEANS for teat traits by parity ... 58
Table 3 LSMEANS of litter and piglet traits by parity ... 59
Table 4 Variance component estimates for teat traits and production traits. ... 60
Table 5 Phenotypic correlation (above the line) and genetic correlations (below the line) for teat traits with production traits ... 61
Table 6 Regression estimates for teat traits (biological dam) with litter size at weaning, weaning weight, litter weaning weight, and piglet survival ... 62
vii LIST OF FIGURES
Figure 1 Associations between birth weight with prenatal, pre-weaning, and nursery
survival (Fix et al. 2010) ... 25
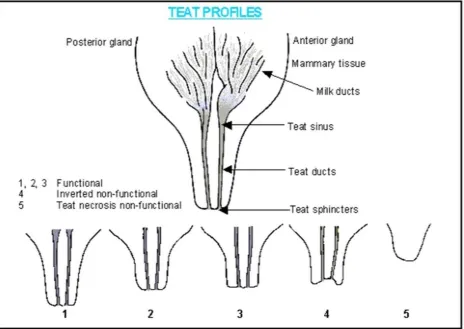
Figure 2 Muirhead and Alexander (1997) illustrate the anatomy of a functional teat and how to classify teats ... 26
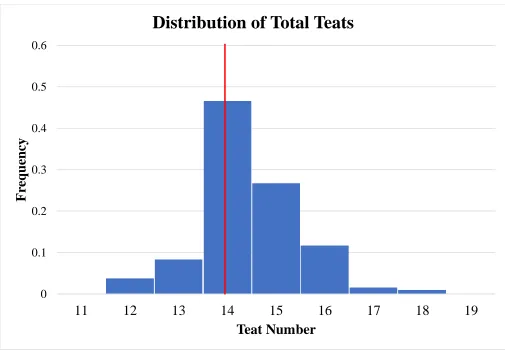
Figure 3 Histogram distribution of total teat number. Population mean represented by red line ... 50
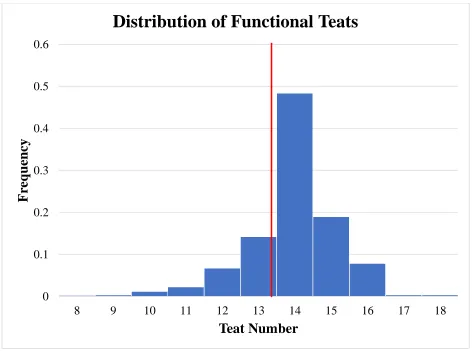
Figure 4 Histogram distribution of functional teat number. Population mean represented by red line ... 51
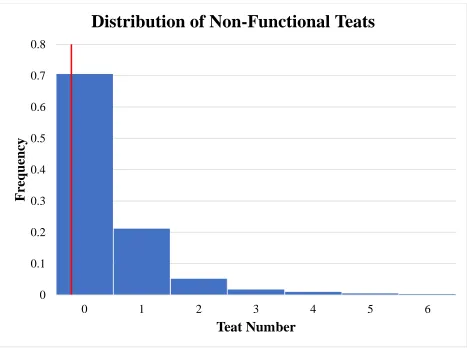
Figure 5 Histogram distribution of non-functional teat number. Population mean represented by red line... 52
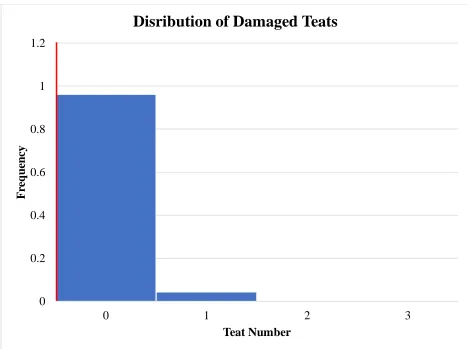
Figure 6 Histogram distribution of damaged teat number. Population mean represented by red line ... 53
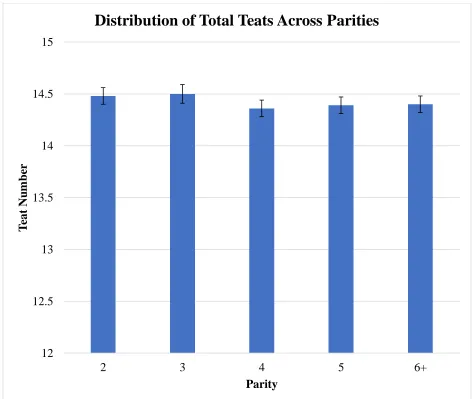
Figure 7 LSMEANS for total teat number across parities ... 54
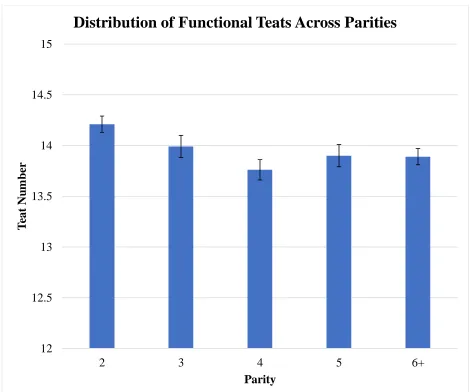
Figure 8 LSMEANS for functional teat number across parities ... 55
1 CHAPTER 1
2 Introduction
Piglet survival has been a point of interest for producers and researchers across the globe
as it pertains to both producer profitability and animal well-being. Traditionally, traits of interest
for genetic improvement have focused on increasing litter size, growth rate, and backfat. Yet
selection for greater litter size at birth has arguably come at the cost of steadily increasing
pre-weaning mortality. From 2005 to 2017, number born alive increased by over two pigs per litter
(11.82 to 13.90 piglets, respectively) and number of piglets weaned increased from 9.30 to 10.30,
respectively (Knauer and Hostetler, 2013; Stalder, 2018). Unfortunately, total number born
increased nearly twice as much as number weaned increased, indicating piglet survival rates have
decreased. From the year 2005 to 2017, piglet survival decreased by 4.5% (78.6 to 74.1%,
respectively) (Knauer and Hostetler, 2013; Stalder, 2018). Pre-weaning mortality is the result of
a complex interaction between the sow, piglet, and the environment (Mainau et al., 2015).
Common causes of piglet mortality include crushing, low viability piglets, and starvation
(Feldpausch et al., 2019). Proper nutrient access will mitigate causes of mortality. Piglet nutrient
access is the driving factor in piglet survival and can perhaps be explained by piglet birth weight,
piglet vitality, sow functional teat number, sow colostrum yield and sow milk yield. Increasing
the number of functional teats in the sow herd will aid in enhancing milk production from the
sow and nutrient accessibility for the piglet (Chalkias et al., 2013).
The number of live born piglets can be greater than the number of functional teats
accessible in some hyper-prolific genetic lines (Rohrer and Nonneman, 2017). This results in a
higher level of competition between piglets and a lower level of early life-weight gain across the
litter (Quesnel et al., 2012). An increase in competition reduces the probability every piglet is
3 functional teat number would have a positive impact on the piglet’s ability to access nutrients,
perhaps increasing colostrum intake. Higher piglet colostrum intake leads to advantages in
growth and development, passive immunity protection between the sow and piglet, and enhances
piglet survival rates (Quesnel et al. 2012). Taken together, previous research suggests
understanding the economic importance and genetics of functional teat number will help
maximize both producer profitability and piglet survival.
Section 1. Nutrient access 1.1. Piglet vitality
Piglet vitality is defined as the strength and vigor of the piglet, with higher vitality
resulting in greater survival rates (Schodl et al., 2019). Piglet vitality varies between individuals
in a litter and is perhaps the most important factor explaining early pre-weaning mortality at the
piglet level (Mainau et al., 2015). Neonatal asphyxia (hypoxia) is the result of severe acidosis
and is the major factor that impacts piglet vitality. Piglets born later in the birth order and those
that come from a prolonged birth period are more likely to have an elevated level of hypoxia
(Mainau et al., 2015). Other negative impacts from hypoxia include poor thermoregulation,
failure of passive transfer of immunity from sow to piglet and greater susceptibility to infections;
which all factor into piglet mortality (Mainau et al., 2015). Piglets with higher vitality will be
more competitive in gaining and maintaining teat access to support their body temperature, thus
giving them an advantage to survive (Decaluwe, 2018).
Piglet vitality can be measured by evaluating the interval between parturition and first
contact with a teat, latency to first suckle, body temperature at 24 hours of age, and early life
weight gain (Sacy et al., 2011). One of the early scoring systems introduced to measure the
4 (Schodl et al., 2019). The APGAR scoring system is perhaps not currently practical for
producers to use due to its complexity and time constraints, as a result more practical methods of
quantifying piglet vitality like latency to first suckle are used to categorize piglet vitality at birth.
Schodl et al. (2019) studied the relationship between piglet vitality and piglet mortality
using 3,171 litters across 23 farms to study the relationship between piglet vitality and piglet
mortality. Using a simplistic scoring system (1 = low vitality to 4 = high vitality), they found as
piglet vitality increased, piglet mortality decreased. The authors reported piglet mortality for
vitality scores 1, 2, 3, and 4 were 29, 23, 16, and 8%, respectively. In the same study authors
reported a heritability of 0.11 for vitality score, suggesting the trait may be somewhat difficult to
improve. Piglet vitality and piglet birth weight are often thought to be related, with low birth
weight piglets more likely to have low vitality. However, Trujullo-Ortega (2011) found that the
heaviest birth weight piglets were also associated with lower vitality scores. Objectively
quantifying piglet vitality through machine learning is being investigated and perhaps will allow
for better understanding of the biology involved and more accurate selection for piglet vigor.
1.2. Birth weight
Selection for larger litters has resulted in less consistency among piglet birth weights and
a higher prevalence of underweight piglets at birth that may lack adequate nutrient intake early in
life (Schodl et al., 2019). An increase in one piglet per litter has shown to decrease piglet
birthweight by 30 to 50 g (Opschoor et al. 2010). Kennedy (2017) reported genetic correlations
between total number born and birth weight for populations of Large White and Landrace sow of
0.71 and 0.34, respectively. These unfavorable associations can be explained by an increase in
intrauterine crowding, with intrauterine growth restriction (IUGR) having a negative effect
5 birth weight, body mass index, and/or crown-to-rump length (Feldpausch et al., 2019). Piglets
with lower birth weights are less likely to receive adequate amounts of colostrum when
compared to larger piglets (Quesnel et al., 2012), resulting in decreased survival rates, lower
2-hour weight gain and lower 24-2-hour weight gain (Vasdal and Anderson, 2012). Low birth weight
piglets tend to have decreased locomotor skills, vitality scores, and aggression to suckle
(Feldpausch et al., 2019), which results in a prolonged latency to first suckle when compared to
larger piglets (Quesnel et al., 2012). Latency to first suckle is related to both survival and growth,
as discussed in the previous section.
Several studies have investigated the relationship between piglet birth weight and piglet
survival, and piglet growth to create thresholds to identify undesirable birth weights. Feldpausch
et al. (2019) evaluated over 4,000 piglets to identify birth weight thresholds to maximize piglet
survival. They found 1.11 kg to be an appropriate threshold, with 15.2% and 84.8% of the piglet
population weighing less and more than 1.11 kg, respectively. Pre-weaning mortalities for low
birth weight and normal birth weight piglets were 34.4% and 8.2%, respectively; indicating
piglets weighing less than 1.11 kg were 5.9 times more likely to die. Similarly, Main et al. (2002)
evaluated 2,204 piglets to identify the relationship between piglet birth weight with pre-weaning
growth and survival. The author reported a threshold of 1.05 kg to significantly improved a
piglet’s chance of survival. Results showed piglets weighing ≤ 1.05 kg at birth compared to those
weighing ≥ 1.99 kg had lower weaning weights (5.2 vs. 8.8 kg), lower lactation average daily
gain (0.17 vs. 0.27 kg per day), and higher pre-weaning mortality (24.2 vs. 4.6%, respectively).
In agreement, Cabrera et al. (2012) examined the effect of piglet birth weight on pre-weaning
survival using 745 pigs. The authors found that piglets weighing ≤ 0.9 kg had a reduced chance
6 Fix et al., (2010) estimated the relationship between piglet birth weight and survival across
different life stages in over 6,000 piglets. The authors reported that lower piglet birth weights
were associated with reduced piglet quality and lower prenatal, pre-weaning, and nursery
survival rates; see associations for piglets’ survival in Figure 1.
1.3. Functional teats 1.3.1. Udder morphology
Understanding udder morphology in sows allows for accurately categorizing teats and
thus selection for functional teats as opposed to non-functional teats is feasible. Suckling is
required to initiate milk production in pigs because sows lack a mammary cistern, meaning that
one functional teat is require per piglet (Lundeheim et al., 2013). Teats are typically categorized
into two different groups, functional and non-functional. Functional teats must have a
well-developed sphincter and rear a sufficient amount of milk (Towers, 2016). Non-functional teats
either have no milking capabilities or don’t produce enough milk to adequately supply enough
nutrients to the piglet. Inverted teats are non-functional because they shaped like an inverse
crater and typically lack a sphincter (Towers, 2016). Inverted teats are also more prone to
mastitis, an infection of the mammary glands. Supernumerary teats (smaller and shorter in size
compared to normal teats) may produce limited amounts of milk but are still considered
non-functional (Towers, 2016). Teats that are environmentally damaged (aggressive piglet sucking or
injury) are also considered to be non-functional due to their inability to rear milk. Larger litters
are associated with a greater occurrence of wounded teats on the sow (Norring et al., 2016).
Environmentally damaged teats are less likely to occur when compared to supernumerary or
7 Previous studies have concluded that the mean for total number of teats is approximately
14 across swine populations (Clayton et al., 1981; Borchers et al., 2002; Lundeheim et al., 2013;
Krupa et al., 2016). Lundeheim et al. (2013) analyzed a population of 57,000 Yorkshires and
reported a population mean of 14.2 with 67% of animals having all functional teats. Willham and
Whatley (1963) reported that the number of teats on each side is not always symmetrical.
As sows grow in age, they also grow in size; which can negatively affect udder
morphology and result in poorer teat access. The upper teat row of the sow may be too high, and
the lower teat row perhaps be too close to the floor for piglets to access in larger sows (Vasdal
and Anderson, 2012). Teat diameter and inter-teat distance can also be negatively affected,
resulting in a lower chance for a piglet to latch on at first suckle. Balzani et al. (2016b) studied
the latency to first suckle across sow parity and found a tendency for piglets born in larger litters
and from multiparous sows to take longer to make first contact with the teats. Latency to first
suckle was not impacted by the vitality score or birth weight of the piglet but was shorter for
those born later in the litter (Balzani, 2016b). Teats posterior from the naval of the sow were
chosen first by 41% of piglets, 33% chose the teats anterior to the naval, and only 27% of piglets
chose the middle teats surrounding the naval (Balzani, 2016b).
1.3.2. Importance
The number of functional teats and accessibility to those teats are two fundamental
limiting factors affecting how many piglets a sow can raise. Evaluating the impact of teat
characteristics on piglet throughput is not a recent discovery; Enfield and Rempel (1961)
reported that an increase in one functional teat improved the number weaned by 0.27 and
weaning weight by 11.7 pounds per litter. More recently, Wiegert (2019) reported similar results
8 (2019) reported an additional 0.27 piglets at weaning with each additional functional teat at
farrowing. Collectively, research suggests that increasing the number of functional teats will
enhance piglet throughput and functional teat number should be estimated or quantified prior to
farrowing to improve cross-fostering strategies.
At the commercial level, phenotypic traits that need to be evaluated when selecting replacement gilts are udder morphology, teat quality, and structural confirmation (Stalder et al., 2010). Phenotypic selection for replacement gilts takes place at different times depending on the trait of interest. Selection at the GDU typically has a minimum threshold needed in order to enter the herd, however this is often negated when demand for gilts is high (Knauer et al., 2018). Total teat number is used in most cases to estimate maternal capabilities and occurs either early on in life (nucleus) or in the gilt development unit (GDU). Typically, seven functional teats on each side (14 total) with proper teat spacing and size is desired upon entry into the herd. Total teat number can be obtained early in life (2 to3 days of age) yet accurate differentiation between functional and non-function teats cannot be determined at this time (Towers, 2016). Chalkias et al. (2013) explained this as 8% of their female Yorkshire population had at least one non-functional teat at 3 weeks of age and 18% at 100 kg live weight, with the total number of teats staying consistent. According to Muirhead and Alexander (1997), producers do not count inverted teats towards the total, even though some inverted teats do become functional through mammary gland development. Gilts with superior genetic potential may be culled due for not meeting a functional teat threshold (Chalkias et al., 2013).
1.3.3. Genetics
Research suggests that there are differences among breeds for teat number (Alston-Mills
9 2019). Total teat number for Duroc, Landrace, Yorkshire and Meishan were 12.5, 14.9, 13.7, and
17.0, respectively (Hirooka et al., 2001). Similarly, Son et al. (2019) analyzed data from three
commercial pig breeds: Large White (n=20,366), Landrace (n=23,398), and Duroc (n=10,044).
Mean values for total teat number were higher in Large White and Landrace (16) when
compared to Duroc’s (13). Earnhardt (2019) reported averages of total teats, functional teats, and
non-functional teats on a population of Landrace x Large White F1 females were 14.93, 13.90
and 1.03, respectively. Hence, why maternal lines are composed of Landrace and Large White
breeds and Duroc genetics are integrated to improve growth and carcass characteristics.
Previous studies suggest selection for teat traits is possible. Heritability estimates for total
teat number range between 0.07 and 0.42, indicating variation among results (McKay and
Rahnefeld, 1990 and Lundheim et al. 2013). This variation may reflect differences in collection
time and/or if males were included in the population(s). Earnhardt (2019) analyzed data from
3,701 sows and reported heritability estimates for total teat number, functional teat number and
non-functional teat number of 0.26, 0.22, and 0.12, respectively. In contrast, Lundeheim et al.
(2013) reported somewhat greater heritability estimates on a population of Yorkshire sows for
total teat number, functional teat number, and non-functional teat number of 0.39, 0.31, and 0.09,
respectively. Chalkias et al. (2013) used a population of Yorkshires to compare heritability
estimates for functional teats and non-functional teats at 3 weeks of age and 100 kg live weight.
In this study, heritability estimates for functional teats decreased from 3 weeks of age to 100 kg
live weight (0.36 to 0.42 vs. 0.29, respectively) and changed slightly for non-functional teats
from 0.02 to 0.03, respectively (Chalkias et al, 2013). Some variation may be explained by gilts
being different ages and having different body conditioning at 100 kg live weight (Chalkias et
10 weeks and recording the occurrence of non-functional teats at 100 kg live weight to use as a
threshold for later management. Son et al. (2019) reported heritability estimates for total teat
number for Large White, Landrace and Duroc of 0.41, 0.39, and 0.28, respectively. Taken
together, these results indicate there’s variation of heritability estimates for teat traits, but genetic
selection is possible.
Previous studies suggest that teat traits are genetically correlated with production traits of
interest. Lundeheim et al. (2013) reported non-functional teats were positively genetically
correlated with litter size (0.20) and number of stillborns (0.34), suggesting that genes impacting
non-functional teats may also have a negative impact on reproductive throughput. Lundeheim et
al. (2013) showed total teats had a positive correlation with functional teats and non-functional
teats, but functional teats and non-functional teats had a negative correlation. Recently,
Earnhardt (2019) reported genetic correlations for total teat number with number weaned and
functional glands at weaning of 0.25 and 0.17, respectively. In the same study, functional teat
number showed stronger genetic correlations for functional teat number with number weaned
and functional glands at 0.50 and 0.72, respectively. Earnhardt (2019) showed non-functional
teats having a negative genetic correlation with number weaned and functional glands at weaning
at -0.38, and -0.57, respectively. Improving on the total teat number will most likely result in
aiding milk production, assuming that all teats produce the same amount of milk (Earnhardt,
2019). Previous research has shown that there are differences in the heritability of teat traits
across breeds.
Researchers have investigated the genomics of functional teat number and discovered
genetic markers that control teat number in swine. Quantitative trait locus (QTL) analysis is a
11 marker. Genetic markers may allow for faster response to selection through exploitation of loci
that independently control teat number (Rohrer and Nonneman, 2017). Rohrer and Nonneman
(2017) reported QTL on SSC1 and 7 explained substantial variation in teat number. The SSC1
QTL is the also known for vertebra number and carcass length, giving indications that these traits
are controlled by common genes to teat number (Rohrer and Nonneman, 2017). Increasing sow
body length may have a negative impact on sow retention and crate sizes will likely need
modification. Researchers concluded that genetic variation in teat number in commercial swine
populations is associated with the VRTN gene on the SSC7 chromosome and vertebra number in
Meishan x occidental cross populations (Rohrer and Nonneman, 2017). The same authors studied
the effect of VRTN on teat number and found other potential QTL markers that impact teat
number including SSC4:24 and SSC10:52 Mb. Each copy of the mutant VRTN allele increased
the total teat number by 0.35 and this region accounted for 30% of genomic variation associated
with total teat number. Rohrer and Nonneman (2017) removed random effect of litter to improve
heritability from 0.41 to 0.49, respectively.
1.4. Colostrum yield
The majority of pre-weaning mortality, within the first day of life, is due to inadequate
intake of colostrum (Quesnel et al., 2012). When colostrum intake exceeds 200 g, mortality rates
were as low as 7.10% and when intake was under 200 g, pre-weaning mortality rates increased to
43.4%, Devillers et al. (2011) reported. Piglets that consumed greater than 290 g of colostrum
tended to be ~ 2 kg heavier at six weeks of age when compared to those who consuming less
than 290 g (Alexopoulos et al., 2018). Colostrum production is the first secretion of the
mammary gland and is the result of lactogenesis II, which starts shortly before parturition and
12 controls milk let down and is produced in the hypothalamus and then released from the posterior
pituitary gland (Wiegert, 2018). Colostrum can be identified from its high concertation of
proteins and is readily available for ~24 hours after parturition and then a cyclical sucking patter
evolves. (Quesnel et al., 2012). Immunoglobin (Ig) G is the major protein is the major protein in
colostrum and is one of five isotypes (Ig A, D, E, G, and M) that differentiate in location and
function (LeBien and Tedder, 2008). The immunoglobulin composition of the colostrum changes
over the first 24 hours after farrowing, changing from a higher protein density to a higher fat
content as time goes on (Quesnel, 2019). Colostrum intake is key to enhancing the piglet’s
passive immunity, thermoregulatory ability, weight gain and subsequent survival (Vasdal and
Anderson, 2012). Intake of colostrum also aids in the development of mucosal brush border
enzymes which enhance the metabolism of lactose and maltose (Alexopoulos et al., 2018).
Colostrum contains hormones and growth factors that promote piglet growth and development
both before and after lactation (Wiegert, 2018). Researchers have focused on improving the
composition of colostrum, but due to its difficulty to measure, less work has been done on
improving colostrum yield (Quesnel, 2019).
Sow colostrum quantity and quality can be impacted multiple ways including hormonally
or environmentally (Quesnel, 2019). Colostrum yield is highly variable among sows, and unlike
milk yield, it is not impacted as much by litter size and suckling intensity (Quesnel, 2019).
Devillers et al. (2007) found colostrum yield per sow was 3.67 kg with individual piglet
colostrum intake at 300 g. Wiegert (2018) reported both a higher sow colostrum yield and
individual colostrum intake of 5.5 kg and 468 g, respectively. Colostrum synthesis is driven by
hormones with prolactin having a positive influence and progesterone having a negative
13 junctions in the mammary epithelium and IgG’s transferring into lacteal secretions (Quesnel,
2019). An environmental effect that could impact colostrum yield includes inducing parturition,
a common practice to allow producers to have tighten farrowing groups and cluster farrowing
times when employees are present. Oxytocin is often administered to induce colostrum let down
during farrowing and/or expedite the farrowing process. Devillers et al., 2007 showed a
significant decrease in colostrum yield for sows induced with PGF2 alpha at day 114 of gestation
compared to non-induced sows (3.37 vs. 4.16 kg). Perhaps more work needs to be done to better
understand the biology behind colostrum production and strategies to increase colostrum yield.
Sow diet, particularly late gestation diet, has been shown to impact colostrum
composition and consumption. Immunoglobulin G can also be increased by introducing late
gestation diets with certain ingredients such as fish oils and probiotics (Quesnel et al., 2019). The
amount of lipids in the sow’s diet has shown to directly impact the fatty acid make-up of
colostrum, as well as piglet colostrum intake. Wiegert (2019) reported for every additional gram
of total added fat intake for the sow in late gestation, piglet colostrum intake and sow colostrum
yield increased by 0.03 and 0.30 g, respectively.
Sow parity has shown to have an effect on the IgG concentration in the colostrum,
explaining 6% of the variation in piglet serum IgG concentration (Cabrera et al., 2012). Gilt
colostrum IgG concentrations have been shown to be lower and decrease more rapidly in the 24
hours after parturition that what is found in multiparous sows; resulting in a lower amount of
colostrum intake for gilt litters. Declerck et al. (2016) reported that colostrum fat levels
decreased by 11% with each increase in parity. This study also showed factors such as breed and
live born piglets also can influence colostrum composition, accounting for 26 and 14%,
14 1.5. Milk yield
Milk yield has shown to be influenced by many factors, particularly nutritionally, and is
heavily related with sow consumption and piglet growth. Metabolizable energy requirements for
the sow are high during lactation because they have to meet maintenance requirements, as well
as convert energy into milk production. Restricting feed intake during lactation, similar to
gestation diets, has shown to decrease overall milk production and litter performance, therefore
ad-libitum (full-feed) feeding is commonly seen in lactation diets (Pluske et al., 2009). Daily
changes in nutrient requirements take place throughout lactation to compensate for changes in
consumption, milk composition, and sow weight loss (Thekkoot et al., 2016). The first two
weeks of lactation see the greatest loss of body reserves to help support milk production, thus
needing an energy dense diet. Peak milk production is around 16 to 18 days of lactation, in
which 65-70% of the sow’s energy consumption is converted into milk production. Strathe et al.
(2017) showed sow milk yield and piglet average daily gain to have a correlation of 0.89, with
average daily feed intake of the sow being less related, but still positive (r = 0.45). Without
proper milk production from the sow, piglet survival and growth rates will be severely
compromised.
Amino acids, particularly lysine, have increased in concentration for lactation diets
recently to compensate for the additional milk production needed to adequately support larger
litters. Lysine is the first-limiting amino acid for lactating sows fed soybean meal-based diets
(Danielsen, 1992). Recommended lysine composition in the diet for lactating sows varies across
previous research from 4.9, 8.1, and 10.5 g per kg (NRC, 2012; Hojgaard et al., 2019; Gourley et
al., 2017). Hojgaard et al. (2019) found their requirement by tracking litter average daily gain
15 per kg having no impact on litter gain. Converting dietary lysine into milk lysine has shown to be
67 to 88% efficient (NRC, 2012; Hojgaard et al. 2019), with 7% of milk’s crude protein
composition being lysine. During peak lactation, over 95% of the sow’s daily Lysine
requirement is related to milk production (Feyera and Theil, 2017); hence the importance of
understanding its impact on the sow and piglets.
Various sow and piglet characteristics also play a key role in sow milk yield. Parity is a
major factor in nutrient requirements, with gilts needing the most energy because they are still
growing and maturing (Thekkoot et al., 2016). Parity has also been reported to influence sow
average daily feed intake and milk production, as well as piglet average daily gain. Strathe et al.
(2017) showed that parity 1 sows have a lower average daily feed intake than parity 4 sows (5.4
vs. 6.4 kg per day) and lower average milk yield (10.7 vs. 11.5 kg per day). Due to the strong
relationship between sow consumption and piglet weight gain, piglets reared from parity 1 sows
had lower average daily gain when compared to piglets reared from parity 4 sows (188 vs. 229 g
per day). Sow backfat (common way to measure body reserves) is measured prior to farrowing in
some countries and feeding levels is adjusted during lactation, with heavier conditioned sows
needing less nutrients and thin sows needing additional nutrients (Thekkoot et al., 2016). Body
weight loss in sows during lactation is correlated with piglet average daily gain (r = 0.42) and
milk yield (r = 0.43) (Strathe et al., 2017). This goes back to the maternal biology of
mammalians, in which sows will sacrifice their own weight to produce nutrients for piglets. First
parity sows have shown to lose 26% of their body fat during lactation, compared 16% from
parity 3 and 4 sows, due to a large portion of their body weight being mobilized for milk
16 primiparous sows compared to multiparous sows because of their lower average daily feed intake
and milk production (Hojgaard et al., 2019).
The on-going selection for increasing litter size has produced sows that are
hyper-prolific, resulting in higher demands for milk production and increase nutrient requirements for
the sows Unlike colostrum yield, milk yield is heavily impacted by litter size and piglet weight
as both factors enhance piglet suckling behavior which stimultates milk production (Quesnel,
2019). In agreement, Thekkoot et al. (2016) reported litter size and mammary gland tissue are
directly correlated resulting in higher milk production in larger litters. Total number born seems
to have little to no effect on piglet average daily gain or body weight loss, but number weaned
has shown to have a strong positive correlation to piglet average daily gain and body weight loss
(0.50 and 0.33, respectively. (Strathe et al., 2017).
Environmental factors, particularly room temperature, have also shown to have a strong
impact on milk yield. The thermal comfort range for lactating sows is 15°C to 25°C, and sows
that are kept in rooms over this range show a decreased average daily feed intake and milk yield
(Vilas et al., 2018). Sows that are uncomfortably warm are less likely to get up to eat and drink,
leading to a domino effect that results in less milk production. Many key elements play a role in
how much milk is produced for each individual sow and are often manipulated in different ways.
Conclusion
It’s obvious that nutrient access, particularly functional teat number, is a limiting factor in
piglet survival when number born alive exceeds the number of functional teats accessible.
Pre-weaning mortality rates continue to be unacceptably high across the industry, with over 50% of
pre-weaning deaths occurring in the first 3 days of lactation (Cabrera et al., 2012) The majority
17 include several factors, including inadequate nutrient intake leading to low vigor. Light weight,
lower vitality piglets are at the greatest risk of mortality, couple that with not enough accessible
functional teats and piglet survival decreases. Considering incidences where number weaned
exceeds the number of teats is rare, functional teat number serves as a vital role in the maternal
capabilities of the sow (McKay and Rahnefeld, 1990).
Production systems should aim to improve the number of functional teats in a linear
relationship to litter size to adequately meet the nutritional needs of the piglets and improve
survival (Chalkias et al., 2013). Functional teat number is a moderately heritable trait ranging
from 0.22 to 0.31 (Lundeheim et al., 2013; Krupa et al., 2016; Earnhardt et al., 2019) and thus
response to selection is expected. Continuing selection for litter size with no regard to increasing
the number of functional teats available will increase the number of pre-weaning mortalities and
will continue to be a both an economic and animal well-being concern for producers. Better
understanding of the genetics, impacts, and relationships between teat traits and production traits
of interest will serve as template for improving the maternal capabilities of the sow.
Section 2. Genetic analysis 2.1. Genetic analysis
Understanding descriptive statistics is the first step in analyzing a data set. Descriptive
statistics are either univariate (involves one variable), bivariate (relationship between two
variables), and multivariate (relationship between multiple variables). Descriptive statistics
quantifies frequencies, means, proportions, ranges, and distributions and are most commonly
displayed in a plot or graph format. Descriptive statistics are key step in statistical analysis
because they depict the raw data in an interpretable way and can give insight on outliers and
18 Estimation of variance components are used to better understand genetic variation in
traits and the genetic relationship between measures. Genetic parameters can then be used to
predict breeding values that can be translated into indexes for selection. Currently, there are
many methods used in estimating variance components, but analysis of variance (ANOVA) and
different forms of restricted maximum likelihood (REML) are commonly used. The standard
procedure for the estimation of genetic parameters and accurate selection in both plant and
animal species is through the use of REML/BLUP (Best Linear Unbiased Prediction) (de
Resende, 2016). The program BLUP increased selection accuracy for additive and dominant
genetic effects (de Resende, 2016). This procedure is known as mixed model methodology and is
optimal for genotypic evaluation (de Resende, 2016). Through REML/BLUP programs,
comparisons among individuals or groups over time and location, simultaneously corrects for
environmental effects, estimation of variance components and prediction of genetic values are
feasible (de Resende, 2016). The GLM (General Liner Model) procedure of SAS (SAS Institute,
Inc., Cary, NC) is a popular program for analysis of variance within and across populations.
Regression, correlation, and variance components estimates require special
accommodations to account for certain interactions and/or relationships in each unique data set,
with certain programs more ideal in cases than others. For instance, Zhang et al. (2011) used the
BLUP method to estimate the heritability and REML to estimate breeding values, variance, and
covariances. Univariate and bivariate models in REML were carried out by Earnhardt (2019)
used to estimate variance components for total teats, functional teats, and non-functional teats.
Variance and covariance, heritability and both genotypic and phenotypic correlations can also be
estimated by using paternal half-sib and full-sib analysis. In this study, fixed effects included line
19 combined an analyzed. Wiegert (2018) analyzed data from four experiments by themselves and
as a meta-analysis across experiments. For meta-analysis, one of the categorical effects will be
the experiment, accounting for variation explained for observations being in that experiment.
Devillers (2007) used the GLM procedure of SAS to analyze individual piglet colostrum intake
across litters. Litter effect is accounted for by the GLM procedure and adjusts the piglets in that
litter accordingly, within differences among the individuals as least square means. There are
conflicting views in which program is best to process animal data, but all are similar in which
20 References
Alexopoulos, J. G., Lines, D. S., Hallett, S., and Plush, K. J. (2018). A review of success factors for piglet fostering in lactation. Animals, 8(3):38.
Alston-Mills, B., Iverson, S.J., and Thompson, M.P. (2000). A comparison of the composition of milk from Meishan and crossbred pigs. Livestock Science, 63: 85-91.
Balzani, A., Cordell, H. J., and Edwards, S. A. (2016a). Development of a methodology to describe udder conformation in sows. Animal: An International Journal of Animal Bioscience, 10(3):432-439.
Balzani, A., Cordell, H. J., and Edwards, S. A. (2016b). Relationship of sow udder morphology with piglet suckling behavior and teat access. Theriogenology, 86(8):1913-1920. Balzani, A., Cordell, H. J., Sutcliffe, E., and Edwards, S. A. (2016c). Sources of variation in
udder morphology of sows. Journal of Animal Science, 94(1):394-400. Balzani, A., Cordell, H. J., Sutcliffe, E., and Edwards, S. A. (2016d). Heritability of udder
morphology and colostrum quality traits in swine. Journal of Animal Science, 94(9):3636-3644.
Borchers, N., Reinsch, N., and Kalm, E. (2002). Teat number, hairiness and set of ears in Pietrain cross: variation and effects on performance traits. Archives Animal Breeding, 45:465-480.
Cabrera, R. A., Lin, X., Campbell, J. M., Moeser, A. J., and Odle, J. (2012). Influence of birth order, birth weight, colostrum and serum immunoglobulin G on neonatal piglet survival. Journal of Animal Science and Biotechnology, 3(1):42.
Chalkias, H., Rydhmer, L., and Lundeheim, N. (2013). Genetic analysis of functional and non-functional teats in a population of Yorkshire pigs. Livestock Science, 152(2-3):127-134. Clayton, G. A., Powel, J. C., and Hiley, P. G. (1981). Inheritance of teat number and teat
inversion in pigs. Animal Science, 33(3): 299-304.
Danielsen, V. (1992). Lysine for lactating sows. SEGES Danish Pig Research Centre, Copenhagen, Denmark. Report:817.
Decaluwe, R. (2018). 4 piglet parameters for lifetime performance. Piglet Progress, December 21, 2018.
21 de Resende, M. D. V. (2016). Software Selegen-REML/BLUP: a useful tool for plant breeding.
Crop Breeding and Applied Biotechnology, 16(40):330-339.
Devillers, N., C. Farmer, J. Le Dividich, and A. Prunier. 2007. Variability of colostrum yield and colostrum intake in pigs. Animal, 1(7):1033-1041.
Devillers, N., Le Devidisch, J., and Prunier, A. (2011). Influence of colostrum intake on piglet survival and immunity. Animal, 5(10): 1605-1612.
Enfield, F.D., and Rempel, W.E. (1961). Inheritance of teat number and relationship of teat number to various maternal traits in swine. Journal of Animal Science, 20(4): 876-879 Knauer, M.T., Wiegert J.G., and Earnhardt, A. (2019). Genetics of swine functional teats.
National Hog Farmer Daily Updates. August 1, 2019.
Earnhardt, A.L. (2019) The genetics of functional teats in swine. North Carolina State University, M.S. Thesis.
Feldpausch, J.A., Jourquin J., Bergstrom J.R., Bargen J.L., Cokenkroger C.D., Davis D.L., Gonzalez J.M., Nelssen J.L., Puls C.L., Trout W.E., and Ritter M.J. (2019). Birth weight threshold for identifying piglets at risk for preweaning mortality. Translational Animal Science, 3(2): 633-64.
Feyera, T., and Theil, P.K. (2017). Energy and lysine requirements and balances of during transition and lactation: a factorial approach. Livestock Science, 201:50-57.
Fix, J.S., Cassady, J.P., Holl, J.W., Herring, W.O., Culbertson, M.S., and See, M.T. (2010). Effect of piglet birth weight on survival and quality of commercial market swine. Livestock Science, 132(1-3): 98-106.
Gourley, K., Nichols, G., Sonderman, J., Spencer, Z., Woodworth, J., Tokach, J., DeRouchey, J., Dritz, S., Goodband, R., and Kitt, S. (2017). Determining the impact of increasing
standardized ileal digestible lysine for primiparous and multiparous sows during lactation. Translational Animal Science, 1:426-436.
Hirooka, H., de Koning, D. J., Harlizius, B., van Arendonk, J. A. M., Rattink, A. P., and Groenen, M. A. (2001). A whole-genome scan for quantitative trait loci affecting teat number in pigs. Journal of Animal Science, 79(9):2320-2326.
Hojgaard, C.K., Bruun, T.S., and Theil, P.K. (2019). Optimal lysine in diets for high-yielding lactating sows. Journal of Animal Science, 97(10):4289-4281.
22 King, R.H. (2000). Factors that influence milk production in well-fed sows. Journal of Animal
Science, 78(3):212-218.
Knauer, M.T. (2013). US swine industry productivity analysis, 2005 to 20010. Journal Swine Health Productivity, 12(5):248-252.
Kurpa, E., Žáková, E., Krupová, Z., and Michaličková, M. Estimation of genetic parameters for teat number and reproduction and production traits from different data sources for Czech dam breeds. Livestock Science, 191:97-102.
LeBien, T.W. and T.F. Tedder. 2008. B lymphocytes: how they develop and function. Blood. 112:1570-1580.
Lundeheim, N., Chalkias, H., and Rydhmer, L. (2013). Genetic analysis of teat number and litter traits in pigs. Acta Agriculturae Scandinavica, 63(3):121-125.
McKay, R. M. and Rahnefeld, G. W. (1990). Heritability of teat number in swine. Canadian Journal of Animal Science, 70:425-430.
Main, R.G., Tokach, M.D., Goodband, R.D., Nelssen, J.L., and Dritz, S.S. (2009). Effects of increasing lysine: calorie ratio in pigs grown in a commercial finishing environment. Kansas Agriculture Experiment Station Research Reports, 0:10.
Mainau, E., Temple, D., and Manteca. X. (2015). Pre-weaning mortality in piglets. FAWEC, November, 2015.
Muirhead, M. R. and Alexander, T. J. L. (1997). Introduction to the anatomy and physiology of the pig. In: Managing pig health and the treatment of disease. 2nd Edition. 5M Publishing, Sheffield, UK.
Norring, M., Valros, A., Munksgaard, L., Puumal, M., Kaustell, K., and Saloniemi, H. (2007). The development of skin, claw and teat lesions in sows and piglets in farrowing crates with two concrete flooring materials. Acta Agri. Scandinavica, 56(3-4):148-154. NRC. 2012. Nutrient requirements of swine. 11th rev. ed. The National Academies Press,
Washington, DC.
Opschoor, C.T., Bloemhof, S., Knauer, M., and Knol, E. (2010). Management influences on birth weight, phase 2. Topigs Technical Report.
Pluske, J.R., Turpin, D.L., and Kim, J.C. (2018). Gastrointestinal tract (gut) health in the young pig. Science Direct, 4(2):187-196.
23 Quesnel, H., Farmer, C. (2019). Review: nutritional and endocrine control of colostrogenesis in
swine. Animal, 13(1):26-34.
Rekiel. A., Wiecek, J., Batorska, M., and Kulisiewicz, J. (2014). Effect of sow prolificacy and nutrition on pre- and postnatal growth of progeny. Annals of Animal Science, 14 (1):3-15.
Rohrer, G.A., and Nonneman, D.J. (2017). Genetic analysis of teat number in pigs reveals some developmental pathways independent of vertebra number and several loci which only affect a specific side. Genetic Selection Evolution, 49:4.
Sacy, A. (2011). Measuring piglet vitality. Pig Progress, June 22, 2011.
Schodl, K., Revermann, R., Winckler, C., Fuerst-Waltl, B., Leeb, C., Willam, A., Knapp, P., and Pfeiffer, C. (2019). Assessment of piglet vitality by farmers-validation of a scoring scheme and estimation of associated genetic parameters. Animal, 9(6):317.
Son, M.V., Lopes, M.S., Martell, H.J., Derks, M.F., Gangsei, L.E., Kongsro, J., Wass,
M.N.Grindflek E. H., and Harlizius, B. (2019). A QTL for number of teats shows breed specific effects on number of vertebrae in pigs: bridging the gap between molecular and quantitative genetics. Frontiers in Genetics, March 26, 2019.
Stalder, K. (2010). Screening replacement gilts for skeletal and reproductive soundness. National Hog Farmer Daily Update. October 19, 2010.
Stalder, K. (2018). 2018 Pork Industry Productivity Analysis. Pork Checkoff, September 11, 2018.
Strathe, A.V., Bruun, T.S., and Hansen, C.F. (2017). Sows with high milk production had both a high feed intake and high body mobilization. Animal. 11(11):1913-1921.
Thekkoot, D., Plastow, G., Kemp, B., Boddicker, N., Dekkers, J., and Rothschild, M. (2016). Blueprint: improving sow lactation performance. National Hog Farmer Daily Update. April 29, 2016.
Towers, L. (2016). Genetics of Teat Number in Swine. The Pig Site. February 12, 2016. Trujillo-Ortega, M.E. (2011). Porcine neonates failing vitality score: physio-metabolic profile
and latency to the first teat contact. Czech Journal Animal Science, 56(11):499-508. Vasdal, G. and Andersen, I. L. (2012). A note on teat accessibility and sow parity consequences
24 Vilas, B.P., Lanferdini, E., Palencia, J.P, Lemes, M.G., Teixeira de Abreu, M.L., de Souza
Cantarelli, V., Ferreira, R.A. (2018). Heat negatively affects lactating swine: a meta-analysis. Journal of Thermal Biology, 74:325-330
Wiegert, J.G. (2019). Effects of practically increasing amino acids and energy in late gestation on colostrum intake, colostrum composition and sow performance. North Carolina State University, Phd dissertation.
Wiegert, J. and Knauer, M. (2018). 98 Sow functional teat number impacts colostrum intake and piglet throughput. Journal of Animal Science, 96:51-52.
Willham, R. L. and Whatley Jr., J. A. (2010). Genetic variation in nipple number in swine. Journal of Animal Breeding and Genetics, 78(1-4):350-363.
26
27 CHAPTER 2
28 Abstract
Genetic selection for improving litter size without increasing the number of functional
teats in a linear fashion has resulted in a piglet nutrient access shortage causing reduced piglet
survival. Number of piglets is often greater than the number of functional teats in modern
maternal lines. This increases competition between piglets and results in nutrient intake
shortages. Therefore, the objective of this study was to evaluate the effect of functional teat
number on reproductive throughput in swine. Teat traits recorded at farrowing included total teat
number (TT), functional teat number (FT), non-functional teat number (NFT), and damaged teat
number (DT); whereas functional mammary glands (FG) was measured at weaning on 814
Landrace Large White F1 females. Sows and litters were evaluated on two levels: biological dam (their own progeny) and wean dam (all piglets nursing sow after cross-fostering). Biological
dam means for TT, FT, NFT, DT, and FG were 14.43, 13.96, 0.42, 0.03, and 10.70, respectively.
Biological dam production traits included litter size at weaning (LSW), average piglet weaning
weight (WW), litter weaning weight (LWW), and piglet survival (LSW/TNB) (SURV). Variance
components were estimated using GibbsF90. Biological dam genetic correlations between TT
with FT, NFT, FG, LSW, WW, LWW, and SURV were 0.79, 0.49, 0.16, 0.11, 0.37, 0.38, and
-0.19, respectively. Biological dam genetic correlations between FT with NFT, FG, LSW, WW,
LWW, and SURV were 0.09, 0.28, 0.39, 0.44, 0.49, and 0.35, respectively. An addition of one
TT changed LSW, LWW, and SURV by 0.13 piglets, 0.36 kg, and 0.41%, respectively. An
increase in one FT changed LSW, LWW, and SURV by 0.32 piglets, 1.28 kg, and 1.63%,
respectively. Heritability estimates for TT and FT were 0.24 and 0.21, respectively, indicating
that response to selection is feasible. These results suggest that functional teat number is more
29 udder morphology, in particular functional teat number, plays a significant role in piglet survival.
Genetic selection to increase functional teat number should be emphasized to improve
30 Introduction
Piglet survival has been a point of interest for producers and researchers across the globe
as it pertains to both producer profitability and animal well-being. Traditionally, traits of interest
for genetic improvement have focused on increasing litter size, growth rate, and backfat. Yet
selection for greater litter size at birth has arguably come at the cost of steadily increasing
pre-weaning mortality. Pre-pre-weaning mortality is the result of a complex interaction between the sow,
piglet, and the environment. Common causes of piglet mortality include crushing, low viability,
and starvation; with crushing being the most common cause early on in lactation (Feldpausch et
al., 2019). Piglets get crushed due to several factors, with inadequate colostrum intake being the
underlying cause of most of them. Perhaps nutrient access for all piglets will mitigate causes of
mortality. Increasing the number of functional teats in the sow herd is one way to enhance milk
production from the sow and nutrient access for the piglet to improve piglet survival (Chalkias et
al., 2016).
The number of live born piglets can be greater than the number of functional teats
accessible in some hyper-prolific genetic lines (Rohrer and Nonneman, 2017). This results in a
higher level of competition between piglets and a lower level of early life-weight gain across the
litter (Quesnel et al., 2012). In previous years, researchers have focused on the impact total teat
number has on performance traits such as number weaned or litter weaning weight and the
heritability’s across teat traits (Enfield and Rempel, 1961; Lundeheim et al., 2013; Wiegert,
2018; Earnhardt, 2019), however little data exists relating the number of functional teats to
performance traits such as weaning weight, litter weaning weight, and survivability. Better
understanding the relationship between functional teat number and production traits of interest
31 understanding the economic importance and genetics of functional teat number will help
maximize both producer profitability and piglet survival. Therefore, the objective of this study
was to evaluate the impact of functional teat number on reproductive throughput in swine.
Materials and methods Animals
Data from 814 Landrace Large White F1 females (Smithfield Premium Genetics, Rose Hill, NC), within a 2,400 head commercial sow farm in eastern North Carolina were used. Parity
distribution for the study population ranged from parity 2 to parity 6+ and had similar
observations per parity group (P2=164, P3=121, P4=147, P5=125 and P6+=178). Sow herd was
PRRS stable. Sows were housed in stalls and group pen from day 0 to 42 and day 42 to 112,
respectively,of gestation.They were then moved to farrowing with either 24 or 48 individual
stalls per room. Sows had ad-libitum feed and water access throughout lactation. All piglets were
individually tagged and weighed 24 hours post parturition. A Nomad Trimble scanner (Trimble
Incorporated, Sunnyvale, CA) equipped with Chad 1.3 software (Smithfield Premium Genetics,
Rose Hill, NC) was attached to a Digi-Star scale (Topcon Agriculture, Livermore, CA) and used
for the collection of both birth and weaning weights. Both individual and litter weights were
captured at birth and weaning.
Data collection
Sampling occurred daily from May 2019 to August 2019. Teat traits were observed on all
sows within 24 hours post-farrowing. Underline traits included total teat number (TT), functional
teat number (FT), and non-functional teat number (NFT). Teats were classified according to a
classification system (Figure 1) developed by Muirhead and Alexander (1997). Functional teats
Non-32 functional teats were defined as inverted (turned inward at the tip) or supernumerary (smaller in
length and diameter) (Muirhead and Alexander, 1997). Environmentally damaged teats (DT)
were also categorized as NFT, these being the result of injury. Sows were observed while
standing, unless a clear visual of the sow’s underline was obtainable while laying down.
Farrowing included birth weight (BW), number born alive (NBA), stillborn piglets (SB), total
number born (TNB), mummified fetuses, crate location, contemporary group and parity.
Cross-fostering occurred post tagging and weighing of the piglets (~24 hours of age).
Piglets placed (PP) was the total number of piglets in a sow’s crate post cross-fostering and
included mortalities that occurred between cross-fostering and piglets placed analysis. Piglets
placed ranged from 12 to 14 piglets depending on the number born alive for litters farrowed that
day. Piglets were weighed again at 19 days of age. Individual weaning (WW) and litter weaning
(LWW) were recorded. Number of functional mammary glands (FG) was recorded during
collection of weaning weights. A teat was considered a functional mammary gland if a sow had a
full mammary gland producing milk. Damaged teats that resulted from catching on the flooring
or aggressive sucking behavior, were still classified as if they had a functional mammary gland if
they were still producing milk. All sows were observed standing up to improve the consistency
of scoring.
Sows were evaluated at two different levels, biological dam and wean dam, with each
sow being placed into both groups. Biological dam evaluated each sow’s biological piglets
through weaning, whether they were cross-fostered or not. Wean dam evaluated each sow’s
weaned pigs, whether they were her biological piglets or not. For the biological dam, litter size
at weaning (LSW) was computed from the collection of individual weaning weights, whereas
33 (SURV) was calculated as litter size at weaning divided by total number born. For the wean dam,
pre-weaning mortality (PWM) was computed by dividing wean number by piglets placed. A
small number of sows were moved to different rooms post cross-fostering to group litters by
birth date.
Data editing
RStudio v. 1.1.456 and Mac Terminal v. 2.8.3 were used to edit collected data. Sows that
died, moved to nurse a different litter, had missing observations, and/or had total litter mortality
post cross-fostering were removed from the data set. Litters with missing birth or weaning
weights were also not included in the data set. After editing, 735 sows and 7,760 piglets were
available for analysis.
Statistical Analysis
Wean dam was used for part of the linear regression analysis, yet the biological dam was
used for all variance components and summary statistics. Variance component estimates for each
trait were performed using univariate and bivariate models for all traits: total teat number,
functional teat number, non-functional teat number, functional mammary gland number, litter
size at weaning, weaning weight, litter weaning weight, and piglet survival. Sow pedigrees were
traced back 6 generations and include the granddam, grandsire, dam and sire. Misztal et al.
(2016) developed a family of programs (BLUPF90) for mixed-model statistical analysis with a
particular focus in animal breeding applications. One of the programs, GIBBS3F90, implements
a blocking factor for a Gibbs Sampler (sequencing algorithm that approximates distributions of
variables). Sequencing results from GIBBS3F90 are used for POSTGIBBSF90 to estimate
variance components. In the current study, two-trait analysis was used. The first 10,000 iterations
34 used to compile parameter estimates, with associated standard errors. Heritability estimates were
calculated by using the following equation:
where h2 is the heritability, a is the additive genetic variance, and e is the residual
variance. Genetic and phenotypic correlations were estimated between traits through binary
analysis and displayed in POSTGIBBSF90 output.
Linear regressions were calculated using the linear model function in RStudio v. 1.1.456,
which fits linear models and computes regressions estimates and associated standard errors.
Sows and litters were analyzed at two levels: biological dam (sow’s biological litter) and wean
dam (litter after cross-fostering). Linear regressions were determined for total teat number,
functional teat number, and functional mammary gland number on litter size at weaning, wean
number, weaning weight, litter weaning weight, piglet survival and pre-weaning mortality for
both biological dam and wean dam when appropriate. Categorical fixed effects included
contemporary group (farrowing room and month) and parity. For biological dam, total number
born was a covariate in regression models for litter size at weaning, litter weaning weight, and
piglet survival. Litter size at weaning was a covariate in the weaning weight model. For wean
dam, piglets placed was a covariate in regression models for wean number, pre-weaning
mortality, and litter weaning weight, and weaning weight included wean number as a covariate.
All models included sow as a random effect.
35 Population distributions for total teat number, functional teat number, non-functional teat
number, and damaged teat number are displayed in Figures 3, 4, 5, and 6 respectively. All
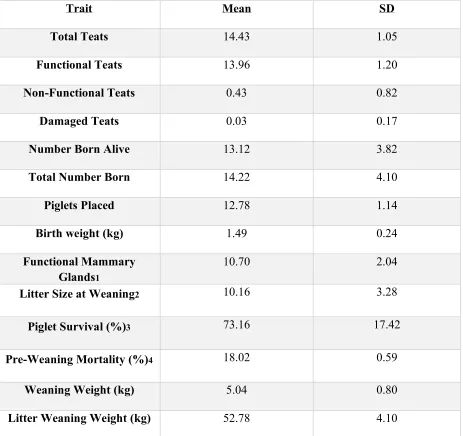
summary statistics apply to biological dam unless stated otherwise. Population averages for total
teat number, functional teat number, non-functional teat number, and damaged teat number were
14.43, 13.96, 0.42, and 0.03, respectively. Normal distributions (P>0.05) were seen for total teat
number and functional teat number. Population averages for reproductive traits such as
individual birth weight, total number born, and number born alive were 1.49 kg, 14.22, and
13.12, respectively. The average for piglets placed was 12.78 across the population. Piglet
survival averaged 73% across the population, hence 27% of the total number born piglets died
prior to weaning. These mortalities include still borns. For wean dam, pre-weaning mortality was
18%. For traits observed at weaning, weaning weight, litter size at weaning, litter weaning
weight, and functional mammary glands at weaning, population averages were 5.04 kg 10.45,
52.78 kg and 10.70, respectively. Summary statistics for teat traits at farrowing and weaning,
reproductive performance and piglet performance data are shown in Table 1.
Parity distributions for teat traits can be found in Table 2. For total teat number, no
significance difference (P>0.10) was seen across parities. Functional teat number was greater
(P<0.01) in parity 2 compared to parity 6+ sows (14.21 vs. 13.89). Parity 6+ sows had twice
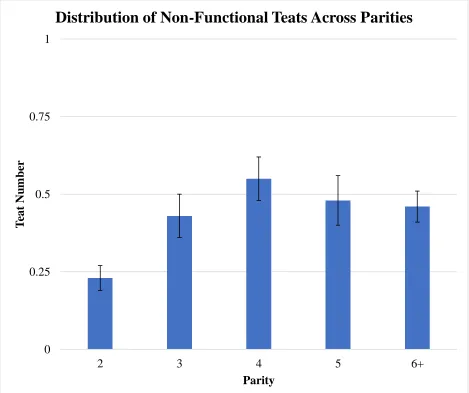
(P<0.01) the number of non-functional teats compared to parity 2 sows (0.46 vs. 0.23)
Distributions for total teat number, functional teat number, and non-functional teat number
across parities can be found in Figures 7, 8 and 9, respectively. No differences (P>0.10) were
observed in damaged teat number across parities. Parity distributions for piglets placed,
functional mammary gland number, litter size at weaning, weaning weight, litter weaning
36 placed on average than parity 6+ sows (13.00 vs. 12.57), greater (P<0.01) functional mammary
gland number (11.21 vs. 10.21), greater (P<0.01) litter size at weaning (11.00 vs. 9.90), greater
(P<0.01) litter weaning weight (55.33 vs. 49.84 kg), and greater (P<0.01) piglet survival (78 vs.
68%). No difference ((P>0.10) was observed for weaning weight across parities.
Variance component estimates
Estimates of variance components for teat and performance traits are reported in Table 4.
The heritability estimate for total teat number was numerically greater than the estimate for
functional teat number (0.24 vs. 0.21). The highest observed heritability for teat traits at
farrowing was non-functional teats (h2 = 0.46). The heritability estimate for functional mammary
gland number was lower than teat traits at farrowing at 0.15. Litter size at weaning’s heritability
estimate was 0.15. The heritability estimate for weaning weight was higher than litter weaning
weight at 0.23 and 0.21, respectively. The heritability estimate for piglet survival was 0.29.
Genetic and phenotypic correlation estimates between all teat and performance traits are
reported in Table 5. Genetic correlations between total teat number with functional teat number,
non-functional teat number and functional mammary gland number are 0.79, 0.49, and 0.16,
respectively. The genetic correlations between functional teat number with non-functional teat
number and functional mammary gland number are 0.09 and 0.28, respectively. Functional teat
had a higher correlation with litter size at weaning (0.39) compared to total teat number (0.11)
and non-functional teat number (-0.40). The genetic correlation between total teat number and
weaning weight was slightly lower when compared to functional teat number and weaning
weight at 0.37 and 0.44, respectively. Non-functional teat number and weaning weight show a
genetic correlation of 0.14. Similarly, functional teat number had a greatest correlation with litter
37 the teat traits observed at farrowing, functional teat number had the only positive correlation with
piglet survival at 0.35, whereas total teat number and non-functional teat number expressed
negative correlations with piglet survival at -0.19 and -0.82, respectively. At weaning, functional
mammary gland had positive correlations with litter size at weaning, weaning weight, and litter
weaning weight at 0.84 and 0.68 and 0.87, respectively, but had a negative correlation with piglet
survival at -0.56. The genetic correlations between litter size at weaning and weaning weight,
litter weaning weight, and piglet survival were 0.64, 0.83, and 0.69, respectively. The strongest
correlation between all traits was between weaning weight and litter weaning weight at 0.92. The
genetic correlation between litter weaning weight and piglet survival was 0.54.
Regression Estimates
Linear regression results evaluating the impact of an additional total teat, functional teat,
and functional mammary gland on litter size at weaning, weaning weight, litter weaning weight,
and piglet survival were carried out for the biological dam. For the wean dam, regressions were
estimated on the impact of one additional total teat, functional teat, and functional mammary
gland on wean number, weaning weight, litter weaning weight, and pre-weaning mortality. For
the biological dam, the addition of one total teat, one functional teat, and one functional
mammary gland all had a positive regression on litter size at weaning at 0.13, 0.32, and 0.52
piglets, respectively. Note that a near 1.0 regression estimate is not seen for functional mammary
gland number on litter size at weaning because the biological litter and weaned litter differ. The
addition of one total teat and functional teat had a negative impact on weaning weight at -0.02
and -0.03 kg, respectively, whereas an additional functional mammary gland resulted in adding
0.02 kg on weaning weight. For every total teat, functional teat, and non-functional teat added,