A study of the developing pituitary gland in zebrafish
Paul Chandwani
Submitted in partial fulfilment of the requirements for the degree of Doctor of Philosophy
May 2002
Department of Anatomy & Developmental Biology, University College London,
ProQuest Number: 10014859
All rights reserved
INFORMATION TO ALL USERS
The quality of this reproduction is dependent upon the quality of the copy submitted. In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uest.
ProQuest 10014859
Published by ProQuest LLC(2016). Copyright of the Dissertation is held by the Author. All rights reserved.
This work is protected against unauthorized copying under Title 17, United States Code. Microform Edition © ProQuest LLC.
ProQuest LLC
789 East Eisenhower Parkway P.O. Box 1346
Acknowledgements.
I would firstly like to thank Steve for all his support, advice, patience and friendship over the duration o f this work. I also appreciate the decision o f Nigel Holder to
introduce me to Steve, and regret that I never got to know him.
The fascination and excitement of developmental biology was introduced to me by Ian Hope at the University of Leeds, and I thank him for his inspirational enthusiasm and for starting me off in the lab.
My family and friends have all been great and I owe much to my parents and to Rina, Anna, and Ravi for their support. Finally I thank everyone in the fish group, past and
ABSTRACT:
The pituitary gland is an endocrine organ with roles in growth, maturation and homeostasis. It is derived from two distinct cell populations in the embryo, which give rise to a mature gland with six distinct hormone secreting cell types. This organ, combined with the attributes o f the zebrafish, provides an ideal model system in which to study cell differentiation and intercellular communication, fimdamentals of developmental biology.
Initially a characterisation of gene expression within the anterior pituitary gland over the first four days of development was carried out using in situ hybridisation and antibody labelling. This provides a description o f normal development. This information was used to identify mutant zebrafish lines in which anterior pituitary development was in some way disrupted. Two such lines investigated in this work carry novel alleles.
A novel zebrafish gene, p tx l/3 , with expression in the developing anterior pituitary gland was studied, firstly to determine the identity of the gene in relation to a number o f possible homologues from other species and then to assess its role in pituitary development. An anti-sense approach was used to address function by preventing translation o f the protein within the zebrafish embryo.
Large-scale genetic screening is an invaluable method o f isolating novel mutant lines. I contributed to two such screens were conducted during the course of my studies; the first was at UCL and the second in Tuebingen, Germany, co-ordinated by Artemis pharmaceuticals. Both focused on isolating mutants with defects in early neurogenesis and axon pathway patterning in the embryonic forebrain. Embryos were screened by morphology and anti-tubulin antibody labelling, with more than 1 0 0
mutant lines being identified.
Novel mutant lines are useful tools in the study of development. One such line studied here, named u320, was genetically mapped to identify the location o f the mutant locus. A phenotypic characterisation of the line was conducted including gene expression analysis and transplantation experiments. This work provides information as to the likely activity of the mutated locus in this novel zebrafish line and the role of the affected gene in the context of development, with a focus on the anterior pituitary.
TABLE OF CONTENTS
Title 1
Acknowledgements 2
Abstract 3
Table o f contents 4
List of figures 11
List of tables 13
1.0 Introduction 14
1.1 The study of developmental biology 14
1.2 Model organisms 14
1.3 Zebrafish as a model organism 16
1.4 Zebrafish forebrain development 20
1.4.1 Specification of neurectoderm 20
1.4.2 Patterning the forebrain 23
1.4.3 Neurogenesis and axon guidance 25
1.5 Molecular pathways involved in forebrain development 31
1.5.1 The Hedgehog pathway 32
1.5.1.1 Zebrafish hedgehog genes 32
1.5.1.2 Hedgehog signal transduction 34
1.5.2 The Nodal pathway 37
1.5.2.1 Zebrafish nodal genes 37
1.5.2.2 The Nodal signalling pathway 38
1.5.2.3 Nodal activity and forebrain development 43
1. 6 The endocrine system 44
1.7 Organs of the endocrine system 46
1.7.1 The hypothalamus 46
1.7.2 The epiphysis 49
1.7.3 The thyroid gland 50
1.7.4 The adrenal gland 51
1.7.5 The pancreas 52
1.7.6 The gonads 52
1.8 The pituitary gland 55 1.8.1 Structure and function of the adenohypophysis 55
1.8.2 Pituitary disorders 58
1.8.3 Development of the pituitary gland 61
2.0 Methods and materials 63
2.1 Maintenance of zebrafish 63
2.2 Observation of embryos 63
2.2.1 Observation of live embryos 63
2.2.2 Observation of stained embryos 63
2.3 Molecular biology techniques 64
2.4 Whole mount in situ hybridisation to detect mRNA transcripts 64
2.4.1 Synthesis of antisense RNA probes 64
2.4.2 Single whole mount in situ hybridisation 65 2.4.3 Double whole mount in situ hybridisation 6 6
2.5 Whole mount antibody labelling 67
2.6 Acetylcholinesterase staining 6 8
2.7 Detection of cell death using acridine orange 6 8
2.8 Plastic embedding sectioning 6 8
2.9 Analysis of cell morphology using toluene blue 69
2.10 Injection mRNA into early stage embryos 69
2.10.1 Preparation of DNA for transcription 69
2.10.2 In vitro transcription of mRNA for injection 69
2.10.3 Injection of mRNA 70
2.11 Transplantation of cells 70
2.12 Biotin detection 71
2.13 Inj ection of morpholinos 71
2.14 PCR 72
2.14.1 RtPCR 72
2.14.2 Genomic PCR 72
2.15 Genetic screens 73
2.15.1 ENU mutagenesis 73
2.15.2 Tuebingen screen 73
3.0 A study of the developing anterior pituitary in zebrafish 75
3.1 Introduction 75
3.1.1 The pituitary gland 75
3.1.2 The tissues of the pituitary gland 75
3.1.3 Induction of the anterior pituitary gland 7 8
3.1.4 Patterning the anterior pituitary gland 87
3.1.5 Summary of anterior pituitary organogenesis 92
3.2 Results 93
3.2.1 A histological study of the developing anterior pituitary gland 93 3.2.2 AChE expression reveals bilateral induction of the
anterior pituitary 98
3.2.3 Zebrafish anterior pituitary induction 101
3.2.4 Patterning within the developing anterior pituitary gland 105 3.2.5 Other genes expressed within the anterior pituitary 111
3.2.6 Activation of hormone gene expression 112
3.3 Discussion 115
3.3.1 Summary of data 115
3.3.2 A comparison with the mouse model of anterior
pituitary development 116
3.3.3 Anterior pituitary development in zebrafish 118 4.0 A novel zebrafish ptx gene with a role in anterior
pituitary development 120
4.1 Introduction 120
4.1.1 The Ptx family of transcription factors 120 4.1.2 Expression of Ptxl in mouse and Xenopus 121 4.1.3 Expression of Ptx3 in Mouse and Xenopus 122 4.1.4 Functions of Ptx proteins in the anterior pituitary 123 4.1.5 Targeted g en e‘knock-down’ in zebrafish 124
4.1.6 A study of zebrafish Ptx 1 /3 in pituitary development 128
4.2 Results 128
4.2.1 p txl/3 gene identity and expression 128
4.2.1.1 Identity of a novel ptx gene at a nucleotide level 128
4.2.1.3 ptxl/3 expression in the diencephalon 134 4.2.2 Non-specific effects of ptxl/3 morpholino injection 134 4.2.3 Normal patterning in the forebrain in p txl/3 morphants 137 4.2.4 Eye development is effected following morpholino injection 137 4.2.5 Anterior pituitary development is initiated in morpholino injected
embryos 142
4.2.6 f f l b expression suggests the gonadotrope cell population is reduced 142 4.2.7 Expression of ptx target genes is reduced or lost in the anterior
pituitary 147
4.3 Discussion 150
4.3.1 Identity of zebrafish ptxl/3 150
4.3.2 Investigation of ptxl/3 function 151
4.3.3 Considerations of morpholino use 153
4.3.4 Summary 154
5.0 Genetic screening in zebrafish 156
5.1 Introduction 156
5.1.1 Two large scale genetic screens in zebrafish 156
5.1.2 Genetic screening as a tool 156
5.1.3 Mutagenic agents 160
5.1.4 Screening in zebrafish 162
5.1.4.1 Haploid screening 162
5.1.4.2 Diploid screening 163
5.1.5 The embryonic zebrafish brain 164
5.1.5.1 Screening the forebrain 164
5.1.5.2 Patterning the forebrain 167
5.1.5.3 The early axon scaffold 167
5.1.5.4 The pineal complex 173
5.1.6 Data from two large scale genetic screens in zebrafish 174
5.2 Results 175
5.2.1 The UCL genetic screen 175
5.2.4 Novel mutant line ul09 exhibits disrupted eye and forebrain
development 179
5.2.5 Novel mutant line ul48.2 exhibits a curly-down tail phenotype 182 5.2.6 Novel mutant line redneck {reJd"^^^) has blood circulation abnormalities
in the brain 185
5.2.7 The Tuebingen 2000 genetic screen 185
5.2.8 Scale of the genetic screen 188
5.2.9 Classes of mutant lines identified 188
5.2.10 A novel zebrafish mutant exhibiting defects in forebrain commissure
formation 189
5.2.11 A novel zebrafish mutation that effects forebrain patterning and eye
development 189
5.2.12 Novel zebrafish lines carrying mutations effecting development of the
brain 194
5.2.13 Mutant lines with non-brain phenotypes 194
5.3 Discussion 200
5.3.1 Large scale genetic screening in zebrafish 200
5.3.2 Assessment of the screening procedure 200
5.3.3 Isolation of lines displaying varied phenotypic defects 201
5.3.4 Unique genes and multiple alleles 201
5.3.5 Limitations of genetic screens 202
5.3.6 Potentials of genetic screens 203
6.0 A study of the pituitary gland in the silberblick/wntl 1 mutant 204
6.1 Introduction 204
6.1.1 The silberblick locus encodes the Wntl 1 protein 204
6.1.2 The gene 204
6.1.3 Wntl 1 function 205
6.1.3.1 The Wnt pathway 205
6.1.3.2 The canonical Wnt pathway 208
6.1.3.3 The non-canonical Wnt pathway 214
6.1.4 Zebrafish gastrulation 217
6.1.5 Outline of this study 219
6.2.1 Isolation of a novel silberblick mutant allele 220 6.2.2 wntl la mRNA transcripts have an insertion in slb^^"^^ embryos 220
6.2.3 The s mutation is lethal 228
6.2.4 The morphology of the silberblick mutant 228 6.2.5 The optic primordia are fused in silberblick embryos 232 6.2.6 Defects are evident in the forebrain of slb'^' embryos 235
6.2.7 The pituitary 241
6.2.8 Over expression of shh restores ff lb and pome expression in the
anterior pituitary and hypothalamus 249
6.2.9 Shh over expression in the mesendoderm affects pome expression in
the pituitary 255
6.3 Discussion 261
6.3.1 Novel zebrafish line ul48 carries a mutation in the wntl la gene 261 6.3.2 slb“^^^ carries a mutation in a splice site within the wntl 1 gene 261 6.3.3 Development of the anterior pituitary in slb'^' embryos 262 6.3.4 Over expression of shh facilitates anterior pituitary development
in slb-^- 262
6.3.5 Pituitary induction in slb'^' embryos 264
6.3.6 Summary 266
7.0 Characterisation of a novel mutant zebrafish line, u320 267
7.1 Introduction 267
7.1.1 Isolation of zebrafish line u320 267
7.1.2 A second allele isolated in the UCL genetic screen probably also carries a mutation in the same locus as u320 267
7.1.3 Characterisation of u320 268
7.1.4 Mutation mapping 268
7.1.5 Candidate gene approach 269
7.2 Results 272
7.2.1 Characterisation of line u320 272
7.2.2 Phenotypic characterisation of line u320 212
7.2.3 Cell death in u320''' embryos 272
7.2.5 In situ hybridisation analysis of gene expression
in u320'^' embryos 277
7.2.6 Gene expression in the u320'^' developing anterior pituitary 284 7.2.7 Transplantation experiments to rescue aspects of
the u320'^' phenotype 287
7.2.7.1 Transplantation experiments can restore Um3 expression
in the u320'^' anterior pituitary 288
1 2 .1 2 Transplantation experiments can restore pigmentation in u320'^’
embryos 291
7.2.8 Genomic characterisation of line u320 291
7.2.9 Candidate genes in the region of Z7871 291
1.2.9.1 ten m4 294
1.2.92 hsp70 294
1.2.9.3 tcf3b 295
1.2.9.4 lim l 296
1.2.9.5 rp2 299
7.2.9.6 spt5 299
7.3 Discussion 300
7.3.1 Characterisation of line u320 300
7.3.2 Functional autonomy of the u320 locus 301
7.3.3 Candidate genes at the u320 locus 303
7.3.4 Summary 305
8.0 General discussion and future directions 306
8.1 Anterior pituitary development in the zebrafish 306 8.2 Using the zebrafish anterior pituitary as a developmental research system 307
8.3 Future work 308
8.4 Summary 310
LIST OF FIGURES
1.1 Stages of zebrafish embryonic development 19
1.2 Genetic patterning and axon tract formation 28
1.3 Mechanisms o f axon guidance 30
1.4 Major components of the Hedgehog signalling pathway 36
1.5 The progression of the Nodal pathway 41
1. 6 The endocrine system of Human and Fish 48
3.1 The structure of the mature pituitary gland 77
3.2 The tissues of the developing pituitary gland 80
3.3 Molecular events in early anterior pituitary development in mouse 83 3.4 Cell lineage specification within the developing anterior pituitary 89 3.5 Histological analysis of the developing anterior pituitary (i) 95
(ii) 96 3.6 Acetylcholinesterase staining of early anterior pituitary cells 100
3.7 Gene expression during pituitary induction 103
3.8 Gene expression within the developing anterior pituitary (i) 107 (ii) 1 1 0
3.9 Expression of hormone-encoding genes 114
4.1 A phylogeny tree of selected ptx genes from various species 130 4.2 Expression o f/?ù:7/5 during zebrafish development 133 4.3 Double in situ hybridisation revealing ptxl/3 and TH expression 136 4.4 Morphology of live embryos injected with p txl/3 morpholino 139 4.5 Acetylated tubulin antibody labeling of morpholino injected embryos 141 4.6 Acetylcholinesterase staining in morpholino injected embryos 144 4.7 An in situ hybridisation for f f l b expression reveals reduced staining
in the anterior pituitary 146
4.8 Stainings for Um3 and pome mRNA transcripts in morpholino
inj ected embryos 149
5.1 Breeding scheme for genetic screening 166
5.2 Acetylated tubulin and opsin antibody labeling at 32 hours post
5.3 Acetylated tubulin and opsin antibody labeling at 60 hours post
fertilization 172
5.4 embryos show defects in heart and neuronal laterality 178 5.5 Defects in development of the eye and optic nerve in mutant
embryos from line ul09 181
5.6 ul48.2''' embryos have a curved body structure 184 5.7 embryos show an accumulation of red blood cells in
the dorsal brain 187
5.8 HC006'^' embryos exhibit abnormal forebrain development 191
5.9 Defective eye and forebrain development in IF084’^' embryos 193
5.10 Mutants exhibiting general brain phenotypes 196
5.11 Other mutants isolated from the Tuebingen screen 199
6.1 Expression of wntl la at tailbud stage 207
6.2 A summary of the Wnt pathway in the inactive and active states 211
6.3 w n tll sequence and inserted nucleotides 223
6.4 The genomic organisation of w n tll in the vicinity of the insertion site 225 6.5 The splice site at the exon 3/intron 3 boundary is mutated
in w n tll gene 227
6 . 6 Morphology of the silberblick mutant embryo (i) 230
(ii) 231 6.7 Organisation of the forebrain at early stages of zebrafish development 234
6 . 8 Expression of shh RNA in the developing silberblick brain 237
6.9 Gene expression in the silberblick brain 240
6.10 isll expression in the silberblick brain 243
6.11 f fl b expression is absent from the silberblick pituitary 246 6.12 pro-opiomelanocortin (pome) expression in silberblick embryos 248 6.13 Over expression of shh mRNA in silberblick embryos 252 6.14 Over expression of shh can induce f fl b expression in slb'^' embryos 254 6.15 The effect of shh mRNA injection on pome expression
in silberblick embryos 257
6.16 pome expression in silberblick mutants over expressing shh
in the axial mesendoderm 260
7.2 Acridine orange staining of live embryos 276
7.3 Tubulin and HuC/Elav antibody labeling 278
7.4 Analysis of gene expression in u320'^' embryos 282 7.5 Analysis of the w52(?'^'developing anterior pituitary 286
7.6 Transplantation experiments 290
7.7 A genetic map of zebrafish linkage group 15 293
7.8 Injection of masterblind mRNA into u320'^' 298
LIST OF TABLES
4.1 Comparison of ptx gene expression 131
5.1 Phenotypic classes of mutants selected to maintain from the Tuebingen 2000
Chapter 1. Introduction.
1.1 The study of developmental biology.
Two objectives drive the study of developmental biology. Firstly, a selfish curiosity of our world and ourselves. The complexity o f life all around us and within us requires explanation. The study of development is fundamental to our understanding of life and provides an attempt at an explanation. The ultimate endeavour of developmental biology is to describe how a given cell is able to divide and give rise to a variety o f differentiated cells in a temporally and spatially distinct manor, guided by a single genome. It is this process that allows diversity of cell type within a given organism, leading to complex patterned life forms with multiple organ systems. A complete description eludes us, but developmental biology, combining the traditional observations o f embryology and the ever-expanding repertoire o f molecular genetic techniques, is providing answers at a pleasing pace.
A second objective is the development of medical technology. With knowledge comes an ability to manipulate, prevent and correct. Developmental biology, apart from the obvious contributions it can make to pre-natal care in humans, contributes hugely to the field o f cancer research. Developmental biology is the study of a controlled proliferation, differentiation and apoptosis of cells. Cancer is the result of a break down in the control of these processes. A number o f proteins studied and discussed in this work are not only key regulators o f development but have been identified as molecules playing an active role in tumourgenesis - the Wnt and Sonic Hedgehog proteins being examples (Taipale & Beachy, 2001).
1.2 Model organisms.
in which, under permissive conditions, a single celled organism undergoes an asymmetric cell division, giving rise to two distinct cell types - a mother cell and a pro-spore (Errington, 1992). A complex range of signals (both internal and external to the single cell) are recognised, transduced and integrated via a number of pathways to messenger molecules that then act at various levels within the cell (upon mRNAs, the genomic DNA and proteins) which results in differential gene expression in each o f the two cells. This is one o f the simplest forms o f life, yet provides much information on the molecular mechanisms that control development.
Single celled organisms such as bacteria and yeast are ideal for the study of cell biology, an understanding of the cell cycle and proteiniprotein interactions (Yanagida,
2002). They are irreplaceable in these fields o f research and will continue to contribute, but are limited for the study of inter-cellular communication, patterning and the development of organ systems. More complex organisms are required, such as the Arabidopsis thaliana plant, and animal systems including mouse {Mus musculus), sea urchin (various species including Paracentrotus lividus), chicken {Gallus gallus), frog {Xenopus laevis), zebrafish {Danio rerio), the nematode worm {Caenorhabditis elegans) and the fruit fly {Drosophila melanogaster).
C. elegans is a free-living nematode worm, the adult being 1mm in length and can be either male or hermaphrodite. The hermaphrodites will preferentially mate with males, but in their absence can self fertilize. Their small size and eating habits (consuming bacteria spread onto the agar dishes on which they live) make them ideal organisms for use in the lab, and the self fertilizing gender, combined with simple mechanisms o f DNA uptake makes genetic manipulation relatively simple (Ankeny, 2001). A complete atlas of cell lineage from egg to adult has also been generated (which is highly consistent in this species, Sulston et al., 1983) and the genome has been fully sequenced (The C. elegans sequencing consortium, 1998).
The study of C. elegans itself has a significance impact upon human health with an estimated 1 billion people carrying intestinal parasitic nematode worms (Grencis,
2 0 0 1), and as a research tool, it provides an ideal model for the function of certain
tool to manipulate and study, although the body plan is very simple and thus there are limits to the focus o f research.
Study of the fruit fly Drosophila melanogaster has, more than any other organism, contributed to our understanding o f a fundamental principle of developmental biology: how genes govern the patterning o f the body. The Drosophila adult is a little under 2mm in length, has a generation time of 9 days and as an experimental system, lends itself to some o f the most powerful genetic techniques available (Wolport et al.,
1998). This is partly a reflection o f the long period of time for which it has been the focus o f research, but it now provides a unique system to integrate the study of biochemistry, genetics, molecular and cellular biology - all useful approaches to the study o f development. Gene targeting by homologous recombination (a technique previously exclusive to yeast and mouse) is being developed as an additional tool in Drosophila (Rong & Golic, 2000), and this combined with a completed genome sequence (Adams et a l, 2000) ensures Drosophila an invaluable place in the future of developmental biology research.
Each of the previously mentioned animal models has unique advantages that can be exploited for particular observations or manipulations to study development. The
zebrafish, as one o f the most recent additions to the model systems was perhaps chosen with modern techniques and directions of study in mind. Zebrafish provide a novel (and photogenic) insight into vertebrate development in vivo.
1.3 Zebrafish as a model organism.
The zebrafish Danio rerio (formerly Brachydanio rerio) has a number o f features that makes it an ideal model system for various biological disciplines, and most conspicuously for the study of development (Streisinger et al., 1981; Eisen et al.,
A single female can generate 200-300 eggs per week, and fertilization can be in vivo (natural spawning) or in vitro.
Fertilization and development of the zebrafish egg is external, and thus the egg is amenable to observation and manipulation from the earliest stages. The embryos are transparent during the early stages o f development, so every step through development can be observed without removing the egg from its normal
microenvironment. This advantage, coupled with tools such as green fluorescent protein (GPP) transgenes and lineage tracers (which can be caged to refine resolution, as used by Rohr & Concha, 2000), allows an in vivo view of the developing embryo at a molecular and cellular level.
Additionally, external development enables transplantation of cells or entire structures (such as the organiser/shield) between embryos, and the microinjection o f functional mRNA, DNA or other biologically active reagents (such as morpholinos). Coupled with caging technology, the temporal and spatial activity of these molecules can be controlled (Ando et al., 2001).
The zebrafish also lend itself to large-scale genetic screening (Mullins et a l, 1994; Haffter et al., 1996 and see chapter 5), a highly productive means o f identifying mutants for the study of development (Nusslein-Volhard & Wieschaus, 1980). As for genetics, the zebrafish field has benefited greatly from the sequencing of thousands of expressed sequence tags (ESTs) and the formulation o f genetic and physical maps
(Geisler et a l, 1999; Kelly et a l, 2000; Hukriede et al., 2001). The zebrafish genome is currently being sequenced in its entirety as a collaboration between The Sanger Institute, Cambridge, UK (www.sanger.ac.uk) and the zebrafish community. This information, applied in combination with emerging technologies such as uncaging and
morpholinos, is likely to greatly accelerate the pace o f research.
The morphological progression o f zebrafish development has been well characterised by Kimmel et al, 1995, (see figure 1.1). The first cell division occurs within 45 minutes of fertilization, and the cleavage period is followed by the blastula period 2V2
Figure 1.1. Stages of zebrafish embryonic development.
AU panels show sketches of embryos.Panel A shows a newly fertilized embryo (O hours post fertilization). The single cell contains mainly yolk, with a cytoplasmic region at the anterior pole.
Panel B is a gastrulating embryo at 50% epiboly in a lateral view (5.5 hours post fertilization).
Panel C is a lateral view of an embryo at tail-bud stage, (10 hours post fertilization). Panel D is a lateral view o f an embryo at the 14-somite stage of development; 16 hours post fertilization, (anterior to the left).
Panel E is a lateral view of an embryo at 25 hours post fertilization (anterior to the top).
Panel F is a lateral view o f an embryo at 35 hours post fertilization (anterior to the top).
Panel G is a lateral view o f an embryo at 42 hours post fertilization (anterior to the top).
Panel H is a lateral view o f an embryo at 60 hours post fertilization (anterior to the left).
Panel I is a lateral view o f an embryo at 72 hours post fertilization (anterior to the left).
Figure 1.1
B
\
T
\
D
a'
G
fertilization (at standard lab conditions, 28.5°C, see section 2.1), and during this period of development (5V4 - 10 hours post fertilization) the axis of the embryo is
established and the neural keel and notochord become apparent. The segmentation period occurs from 1 0 - 2 4 hours, during which time primary organogenesis occurs (including of the pituitary gland and brain). The pharyngula period occurs between 24 - 48 hours o f development, during which pigmentation o f the body and blood circulation commences. This is followed by the hatching period (48 - 72 hours post fertilization), which sees rapid morphogenesis of the primary organ systems. By 3 days post fertilization the swim bladder inflates and the mouth opens. Embryos are now free swimming and feeding (Kimmel et al., 1995).
1.4 Zebrafish forebrain development.
The tissues that compose the forebrain are the eyes, optic stalks, telencephalon and diencephalon (which includes the hypothalamus and posterior lobe o f the pituitary gland). This study is primarily based on the anterior pituitary gland, a non-neural organ. An understanding of forebrain development is however vital for a study of the anterior pituitary. The anterior pituitary is derived from tissue in the immediate proximity of the forebrain. Many of the mechanisms involved in patterning the forebrain similarly influence the developing anterior pituitary (as described in chapter 6). The anterior pituitary, juxtaposed to tissues o f the forebrain throughout its development, also relies on molecular information secreted from tissues within the
forebrain, namely the ventral diencephalon (Gleiberman et al., 1999). These tissues will remain in physical contact into adulthood, thus the anterior pituitary can be seen
as a part of the forebrain itself, albeit derived from non-neural ectoderm.
1.4.1 Specification of neurectoderm.
mesoderm), underlying it. This mesoderm tissue itself is derived from the organiser, or specifically the shield in zebrafish (Shih & Fraser, 1996).
The organiser tissue was first identified in newt, by the experiments o f Spemann & Mangold (Spemann & Mangold, 1924). The organiser tissue (the dorsal blastopore
lip in newt) was able to induce a secondary axis, ultimately resulting in a second, conjoined embryo. The transplanted organiser contributed completely to the mesoderm o f the second embryo, however host cells were also induced by the transplanted tissue and thus also contributed to the second embryo. A second neural plate was induced composed of host cells. This indicates that the transplanted tissue is a source of molecular signals, able to alter the fate of host cells.
Similar experiments have been conducted in zebrafish at both a tissue and molecular level. Tissue transplants show that a transplanted shield contributes to the hatching gland, head mesoderm, notochord, somitic mesoderm, endoderm and the ventral neural tissue o f the secondarily induced axis (Shih & Fraser, 1996). These are more tissue types than observed to be derived from the amphibian organiser, however a refinement of this experiment suggests that the donor tissue may be contributing only to the notochord, prechordal plate (head mesendoderm) and floor plate (ventral neural
tissue) (Saude et a l, 2000). Either way, the principle that the transplanted tissue can organise the host cells, which then contribute to the ectopic axis, remains. Neural tissue is induced, composed of host cells.
(Fekany et al., 1999). Loss of functional Dharma results in a loss of various tissues including notochord, the prechordal plate and the forebrain. Functionally, Dharma is believed to interact with the Nodal, Wnt and Bmp pathways (Solnica-Krezel & Driever, 2001).
The dorsal ectoderm will give rise to neural tissue. Bone Morphogenetic Proteins (Bmps) are involved in establishing dorsoventral pattern within the ectoderm.
Zebrafish swirl mutants (lacking bmp2b activity) and snailhouse mutants (lacking bmp7 activity) show a dorsalised phenotype, with increased neurectoderm and a loss of non-neurectoderm (Nguyen et aL, 1998; Schmid et aL, 2000). Over expression of bmp4 causes a ventralisation of the embryo and an expanse of intermediate mesoderm and non-neural ectoderm at the expense o f anterior mesoderm and neural tissue (Neave et aL, 1997). Thus Bmp activity seems to promote a non-neurectoderm fate in the ectodermal tissue.
Bmps are however at least transiently present throughout the ectodermal tissue, with essential activity in patterning the neural tube (Mehler et aL, 1997). For dorsal ectoderm to take on a neural fate. Bmp activity needs to be suppressed during gastrulation. A number of proteins, common across species (Sasai, 2001) have been found to suppress Bmp activity (but not necessarily bmp expression) in the dorsal ectoderm allowing progression to a neural fate, including Chordin, Follistatin and Noggin (Dale & Jones, 1999). Recent conclusions from work on zebrafish bmp, smad (downstream transducers o f Bmp signalling) mutants and noggin mRNA over expression suggests some additional complexities to the model. The decision between a neural and non-neural fate for the ectoderm is not a binary one dependent upon the presence or absence of Bmp, but instead Bmp is active at low concentrations across the neural tissue, influencing fate decisions of neural and non-neural ectoderm cells alike in a concentration dependent manner (Barth et aL, 1999). A low level of Bmp activity may be the requirement for a neural fate, as opposed to an absence.
neural plate, including as mentioned here the induction by the underlying mesoderm and a repression o f non-neural ectoderm factors (Bmps). Both these pathways, though perhaps independent, are reliant upon the organiser. The resulting picture, including the establishment o f the organiser itself involves further signalling pathways, including the Wnt and Nodal pathways. Thus neural induction is a complex process that lies at the ever-retreating boundaries of our understanding.
1.4.2 Patterning the forebrain.
The anterior neural plate relies upon both intrinsic and extrinsic cues (molecular
signals) for correct patterning. External signals include those from the mesendodermal tissues underlying the forebrain, which form the prechordal plate. Row 1 describes a population of cells that lie at the anterior border, within the neural plate, which secrete molecules influencing the identity of cells that will form the most anterior forebrain structures.
Various mutant zebrafish lines show defects in prechordal plate establishment and activity. In most cases, this results in disrupted forebrain patterning, including
cyclopia and a loss of dorsoventral identity (Varga et al 1999; Rohr et al., 2001; Schier & Talbot, 2001). The mutations may lie in the Nodal, Hedgehog or Wnt signalling pathways. Both Nodal and Hedgehog are secreted by the prechordal plate and thus are required for its function. Additionally Nodal and Hedgehog signalling are required for the establishment of the prechordal plate. Nodal may lie upstream of Hedgehog in various aspects of this function (Müller et aL, 2000; Rohr et aL, 2001).
Row 1 plays a role in patterning the anterior most tissues o f the forebrain (Houart et aL, 1998), the eyes and the telencephalon (Varga et aL, 1999). The ablation o f this population of cells causes a loss of telencephalic specific gene expression, (Houart et aL, 1998). This is similar to the phenotype observed in masterblind {mbl) mutants (Heisenberg et aL, 2001). Furthermore, when row 1 cells are transplanted to a more posterior position on the neural plate, they can induce ectopic expression of genes normally expressed in the telencephalon (Houart et aL, 1998).
The m bl locus encodes A xinl, a protein involved in the transduction o f Wnt signalling. It is suggested that mbV^' embryos may have lost the ability to regulate Wnt signalling resulting in the increased expression of Wnt target genes (Heisenberg et aL, 2001). Due to the similarity in phenotype to a row 1 ablation experiment, it can be predicted that row 1 ablation phenotype results from a loss o f Wnt signalling regulation, (or repression). Indeed recent results suggest that row 1 cells express a novel SFRP protein (Wnt inhibitory molecule, Rattner et aL, 1997) termed Tic. Tic activity is able to rescue the row 1 ablation phenotype, and copy the effect of row 1 transplantations (Houart et aL, in press). This data is supported by expression data suggesting SFRPs are involved in telencephalic development o f the mouse (Kim et aL, 2001).
Anterior patterning of the forebrain thus requires inhibition of Wnt ligands, possibly wntSb and w n tl, which are both expressed in the anterior neural plate (Kelly et aL,
1995; Kelly & Moon, 1995). Other genes expressed within the anterior neural plate also influence the identity o f neighbouring cells, including fgfS, which is able to induce the expression of w n tl, engrailed 2 and fg fS itself in the chick midbrain (Crossley et aL, 1996). In zebrafish it was been shown that fgfS also regulates its own expression in the forebrain and is required for the development of specific forebrain midline cell populations (Shanmugalingam et aL, 2000).
morphologically. These territories will contribute to define the identity of neurons, and the migratory pathways of axons.
1.4.3 Neurogenesis and axon guidance.
Segmentation, a common theme in development and within the CNS, is suggested to exist in the process of defining territories within the forebrain. Various models o f segmentation exist, the most recent being the prosomeric model, dividing the forebrain into six prosomeres (Rubenstein et al., 1994). A similar principle of rhombomeric division is an accepted model of hindbrain development (Lumsden &
Keynes, 1989). Neuromeric units are transient developmental structures that relate to gene expression boundaries, as opposed to functional structures of the mature nervous system (Lumsden & Krumlauf, 1996).
The forebrain is suggested to be defined by six neuromeric regions called prosomeres, P1-P3 being diencephalic regions and the more anterior P4-P6 are composed of telencephalic tissue dorsally, and hypothalamic (diencephalic) tissue ventrally (Rubenstein et al., 1994). These boundaries, which can be observed by gene expression domains as a read out, will influence the differentiation of neuronal precursors (Lumsden et al., 1994; Wilson et al., 1997).
Neurogenesis in Drosophila works on the principle o f lateral inhibition. More cells are competent to proceed to a neural fate than are required, and by lateral inhibition a correct number of correctly spaced neurons will differentiate (Artavanis-Tsakonas et al., 1995). The expression of basic helix-loop-helix (bHLH) proneural genes of the achete-scute complex contribute to neuronal differentiation (Gomez-Skarmeta et al, 1995; Skeath & Doe, 1996). A similar process is observed in vertebrate neurogenesis.
activity (as in Drosophila), but also Sonic Hedgehog activity, a signalling molecule expressed in the forebrain (Blader et ah, 1997, and see section 1.5.1). At least three neurogenin genes exist in zebrafish (Wang et al., 2001) and a number of other closely related bHLH factors are also involved in neuronal differentiation, including the NeuroD family of transcription factors (Liao et al., 1999).
Differentiation of early neurons within the zebrafish forebrain occurs in a stereotyped manor, (as described by Wilson et al., 1990), see figure 1.2. The axons projected by these neurons appear to respect the boundaries of gene expression domains within the forebrain, as observed for members o f the eph,forkhead, pax and wnt gene families (Macdonald et al., 1994). Guidance of the axon growth come must be dependent upon factors regulated by these genes.
The axonal outgrowth of the neuron is lead by the growth cone at the tip of the axon. The growth cone moves and senses its environment by the constant extension and retraction of filopodia. There are two main reactions of the growth cone dependent upon molecules sensed in the environment, either attraction or repulsion. This simple mechanism guides the growth cone, and thus the axon to its correct target (Wolport et al, 1998).
At a molecular level, both attractants and repellents can act either in short range or
Figure 1.2. Genetic patterning and axon tract formation.
Panel A shows the positions of early neurons bom within the developing zebrafish brain at around 18 hours post fertilization.
Panels B and C are sketches illustrating the relative expression domains of various genes within the developing neural tissue. Panel B is the anterior neural plate at early neurula stages, around 10 to 12 hours post fertilization. fgfS expression is observed at the anterior border of the neural plate, shh in the midline tissue. These genes in turn influence the expression of other genes such as pax6 (yellow) and pax2 (red).
Panel C shows the expression of fgfS, pax2.1, and shh later in development (24 hours post fertilzation) from a frontal view. This panel also includes a lateral view of the axon scaffold. The gene expression domains represented in panels B and C may influence axon guidance pathways to establish the characteristic axon scaffold.
Panels D, E and F show an embryo at 28 hours post fertilization stained with tubulin antibody. The main components axon scaffold (labelled) are visible in a lateral view (panel D), a dorsal view (panel B) and a rostral view (panel F).
Sketches in panels A, B and C are by Steve Wilson.
Abbreviations:
ap = anterior pituitary d = diencephalon hy = hypothalamus mb = midbrain os = optic stalk
pd = presumptive diencephalon phy = presumptive hypothalamus pos = presumptive optic stalk pr = presumptive retina
pt = presumptive telencephalon r = retina
t = telencephalon
AC = anterior commissure
HCI = hindbrain commissural intemeurons LLP = lateral line projection
OlfB = olfactory bulb ON = optic nerve
PC = posterior commissure POC = postoptic commissure
TAC = tract of the anterior commissure TPC = tract of the posterior commissure
TPOC = tract of the postoptic commissure Trig = trigeminal ganglion
Figure 1.2
l e f X a c c
OS /•
s h h Il
S M \
l)\DT
DVDT
Figure 1.3. Mechanisms of axon guidance.
Panel A illustrates the activity of long-range cues. These are secreted from a source that may be some distance away from the growth cone. The secreted molecules will be detected by the growth cone and will cause it to either be repulsed from the source (e.g. Semaphorins) or attracted to the source (e.g. Netrins).
Panel B illustrates axon guidance by short-range cues. These molecules, such as Ephrin ligands (causing repulsion o f the growth cone) and Cadherins (causing attraction of the growth cone) will be bound to either the outer membranes o f other cells or to the intercellular matrix.
Figure adapted from Wolport et a l, 1998.
Figure 1.3
e.g. Semaphorins e.g. Netrins
B
e.g. Uph ligands e.g. Cadherinsr"
+The transparent composition of the developing zebrafish embryo combined with the availability of many mutants makes for an attractive model on which to study axon guidance (Karlstrom et al., 1997). The guidance o f the retinal axons (which form the optic nerve) from the retina to the tectum (retino-tectal pathfinding) has been a major focus. Various classes of mutants exist (reviewed by Hutson et ah, 2002). In some mutants, {belladonna, chameleon, space cadet and sonic you) the axons fail to leave the retina. Another common phenotype is a failure of the axons to cross the midline,
(observed in mutants including blow out, detour, iguana and smooth-muscle-omitted). In all six classes of phenotype are described. A huge number of genes involved in axon guidance have been identified in zebrafish, both encoding guidance molecules (such as roundabout, Lee et ah, 2001, and various Eph ligands, Brennan et ah, 1997) and those encoding proteins upstream o f the guidance molecules (such as the fgfS gene, Shanmugalingam et ah, 2000, and the gU2 gene, Karlstrom et ah, 1999)
Due to the stereotyped nature of early neurogenesis and axonal guidance in the zebrafish brain, this system is ideal as a readout o f not only the process involved directly in axon formation but also those involved in the general patterning and integrity of the forebrain. Thus the axon scaffold is an ideal marker for large-scale mutation screening. The early axon scaffold in relation to genetic screening is discussed in greater depth in chapter 5.
1.5 Molecular pathways involved in forebrain development.
Signalling mechanisms between cells specify regional identities within the developing brain (reviewed in Wilson et ah, 2002). Anterior-posterior (AP), dorso-ventral (DV) and left-right (LR) patterning events within the early neural tissue provide
1.5.1 The Hedgehog pathway.
The Hedgehog (HH) pathway is typical o f molecular pathways identified in developmental biology in that it functions in a variety o f tissues at different times throughout development. It is able to direct cells to a variety o f states at different points in development (both temporal and spatial) primarily by the differential response in recipient cells, dependent upon their own status. Following is an introduction to the hedgehog genes of zebrafish and current thoughts on the molecular pathway in which they function.
1.5.1.1 Zebrafish hedgehog genes.
The first hedgehog gene identified was the D rosophila segment polarity gene hedgehog (hh) (Nusslein-Volhard & Wieschaus, 1980; Lee et al, 1992). All hh genes encode two peptides, an N-terminal product which initiates Hedgehog signalling (and is referred to as the HH protein herein), and a C-terminal product that has protease activity (Bumcrot et aL, 1995). The C-terminal product functions to modify the secreted HH protein with the addition o f cholesterol to the N-terminal. This is suggested to be critical for the long-range signalling activity of HH, facilitating its regulated secretion (Lewis et aL, 2001; Zeng et aL, 2001), although the exact mechanism is unclear (Ingham, 2001). The activity o f the secreted HH protein is further enhanced by the addition o f palmitoyl to the C-terminal, by an as yet unidentified enzyme (Kohtz et aL, 2001).
Three homologues of the Drosophila hedgehog gene have been identified in both zebrafish and mammals. The mammalian hedgehog genes, (identified in mouse) are called sonic, Indian and desert hedgehog (Echelard et a l, 1993). In zebrafish, the three hedgehog family genes are called sonic, echidna and tiggywinkle hedgehog (Krauss et al, 1993).
(reviewed by Marti & Bovolenta, 2002). The most detailed analysis o f vertebrate HH function is focused on its role in specification of ventral structures in the neural tube.
Underlying the ventral midline of the neural plate is the notochord, a rod o f axial mesoderm cells providing structural support to the early, boneless embryo. The notochord tissue o f chick was shown, by grafting experiments, to influence the identity o f cells within the neural tube. Notochord tissue is responsible for establishing a population o f cells in the ventral midline o f the neural tube, called floor plate cells. Floor plate cells in turn have a role in patterning the dorsoventral axis of the neural tube (Placzek et aL, 1990). The Sonic Hedgehog protein was found to be the responsible agent, (Roelink et aL, 1994), secreted from the notochord with the ability to induce both floor plate at the ventral extreme, and motor neurons more dorsally. Subsequently, Shh expression is induced in the floor plate itself. Many cell types (both neuronal and non-neuronal) are found to differentiate in a HH concentration dependent manner along the dorsoventral axis of the neural tube (Patten & Placzek, 2000).
The induction of the zebrafish floor plate however may be regulated by a different mechanism. Fish that lack both Sonic and Tiggy-winkle Hedgehog activity still form a subset of medial floor plate cells in the ventral neural tube (Etheridge et aL, 2001). Instead, Nodal signalling may be the key molecule in zebrafish, as various mutants in this pathway lack all floor plate (Gritsman et aL, 1999). The Nodal and Hedgehog pathways may also be interacting in the zebrafish axial mesoderm (Varga et aL, 1999; Rohr et aL, 2001).
1.5.1.2 Hedgehog signal transduction.
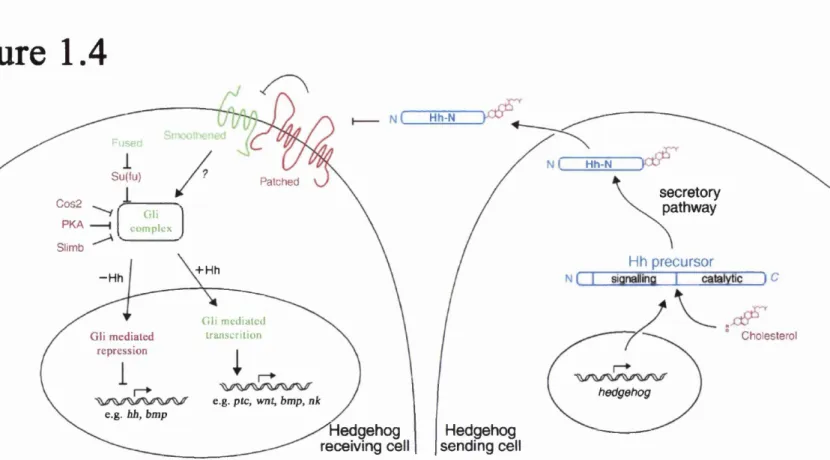
A summary of the interactions between some of the key proteins in the HH pathway is illustrated in figure 1.4. Hedgehog proteins across species signal by binding to membrane bound Patched (Ptc) proteins (Ingham et aL, 1991; Stone et aL, 1996; Goodrich et aL, 1996). Two Ptc proteins have been identified in both mouse and zebrafish (Lewis et aL, 1999). This Ptc-HH complex is then internalised by the cell, (Strutt et aL, 2001), where it alters the status of the Smoothened (Smu) protein.
In the resting state o f the pathway, without HH signalling, Ptc proteins repress the activity o f the Smu protein, a function described as tumour suppressor activity (Wicking & McGlinn, 2001). Activation of the pathway causes the Ptc protein to lift this repression. Although Ptc is in a complex with HH, there is no interaction between HH and Smu (Ingham & McMahon, 2001). Smoothened is a G-protein coupled receptor, related to the Frizzled family o f Wnt receptors (Dann et aL, 2001) that is essential for classical HH activity (Chen et aL, 2001).
Evidence is also emerging o f a Ptc-independent mechanism o f Hedgehog signal
transduction (Ramirez-Weber et aL, 2000; Testaz et aL, 2001), but the molecules responsible for intracellular transduction are as yet elusive. Potential candidates exist such as Hedgehog Interacting Protein (HIP), which binds all vertebrate HH proteins, but it is likely to be involved in regulation of ligand secretion as opposed to activating a reception response (Chuang & McMahon, 1999; Ingham & McMahon, 2001).
The biochemical workings of Smu are unclear, but the transcriptional target of Smu in Drosophila is the cubitus interruptus {ci) protein (Forbes et aL, 1993). Ci is a transcription factor that mediates expression of HH target genes, although mutant
Figure 1.4. Major components of the Hedgehog signalling pathway.
This figure illustrates the main components of the Hedgehog signalling pathway. The cell on the right is expressing hedgehog. The resulting Hedgehog peptide is then cleaved. The N-terminal part is the active signalling molecule. The C-terminal part shows enzymatic activity, attaching cholesterol to the N-terminal part. This modified protein is then secreted by the cell.The cell on the left is the recipient cell. Hedgehog binds to the receptor molecule Patched on the cell surface. This binding event lifts the repression of Smoothened by Patched. Smoothened acts upon Gli proteins, which then enter the nucleus to direct expression of target genes. The target mentioned genes are a few examples o f many.
When Hedgehog signalling is not occurring. Patched represses Smoothened, which ultimately leads to both a lack of Gli-mediated activation and Gli mediated repression of Hedgehog target genes.
Other factors mediate the pathway by interacting with Gli proteins. PKA, Su(Fu) are examples o f proteins that repress Gli activation. Other factors such as Fused then regulate these proteins. This results in a pathway which can receive multiple inputs and thus can direct a wide array of responses.
Figure 1.4
H h -N
Fused
H h - N S u ( f u )
Patched
secretory pathway
C o s 2
PKA
Gli
c o m p l e x
S lim b
Hh precursor
+ H h - H h
C h o l e s t e r o l
G li m ediated transcrition G li m ediated
rep ression
hedgehog
e .g . p tc, wnf, b m p , nk
e .g . hh, b m p
^Hedgehog receiving cell
Gli-2 appears to have both activation and repression activity over the expression of HH target genes (Altaba, 1998; Sasaki et aL, 1999). A zebrafish line, you-too, carries a mutation in the gli-2 gene, which interestingly lacks lateral floor plate (Karlstrom et aL, 1999). As discussed in the previous section, HH signalling plays an important role in floor plate specification (though not definitive in zebrafish). A suggested target gene of HH signalling in the floor plate is the wnt4b gene, which is not expressed m. you-too mutants (Liu et aL, 2000). Furthermore, the developing anterior pituitary in you-too mutants is trans-fated and expresses proteins characteristic of lens tissue (Kondoh et aL, 2000).
The nature of the mutation in you-too may result in a protein exhibiting dominant- negative function (Karlstrom et al., 1999). This indicates that target genes o f gli-2 are continually activated, even if HH signalling no longer prevails. It is speculated that anterior pituitary development requires a cessation of HH activity, as opposed to lens tissue, which requires continuous activation of the pathway (Kondoh et a l, 2000), thus leading to the observed fate transition. The implication on the floor plate phenotype and loss of wnt4b expression in you-too may be due to a repressive function of Gli-2, blocking transcription activation by other Gli-proteins.
The HH pathway in relation to anterior pituitary development is discussed further in chapter 3, and manipulated experimentally in chapter 6.
1.5.2 The Nodal pathway.
1.5.2.1 Zebrafish nodal genes.
nodal genes are involved in a variety o f pattering events during development (Whitman, 2001). Mouse nodal is expressed in the epiblast prior to gastrulation, signalling to both the extra-embryonic ectoderm and visceral endoderm, tissues that
subsequently function to establish the AP axis o f the embryo (Lu et aL, 2001; Beddington & Robertson, 1999). During gastrulation, nodal is expressed in the presumptive mesendoderm (primitive streak) and following gastrulation, it is
expressed asymmetrically, exclusively in the left side of the lateral plate mesoderm, where it regulates the expression of genes implicated in LR patterning (Concha et aL, 2000; Wright, 2001).
Expression of zebrafish nodal genes {eye and sqt, previously nodal related! (ndr2) and nodal relatedl {ndrl) respectively) is similar to mouse Nodal expression, sqt is maternally expressed, and both eye and sqt are expressed prior to gastrulation in the marginal cells, then in the germ ring and later subdivide the organiser tissue into two zones, sqt is expressed in a small population of cells called the forerunner cells which will not involute, while eye is expressed in the hypoblast layer of the shield and is later also expressed later in the prechordal plate (Rebagliati et aL, 1998; Feldman et aL, 1998; Sampath et aL, 1998). Loss of eye and sqt expression (double mutants) causes a change o f fate specification for dorsal marginal cells, from endoderm and
axial mesoderm to a neurectoderm fate. The involution o f these cells during gastrulation does not occur in mutants (Feldman et aL, 2000) and embryos show a lack o f all dorsal mesoderm and endoderm derived structures (reviewed in Schier & Talbot, 2001).
1.5.2.2 The Nodal signalling pathway
progression of the nodal signalling pathway in both mouse and zebrafish is shown in figure 1,5. There are two types of receptors: type I receptors, which activate intracellular proteins for signal transduction, and type II receptors, which activate type I receptors to transduce the signal (Massagué, 1998). Type I receptors include the structurally similar Alk4 and TARAM-A proteins (Reissmann et aL, 2001); type II receptors include ActRIIA and ActRIIB (Song et aL, 1999).
Additionally, co-receptors are involved in reception of the intercellular Nodal signal, which are members of the EGF-CFC family of proteins. One-eyed pinhead (Oep), a zebrafish co-receptor of Cyc and Sqt proteins, is required for effective Nodal signalling and its loss results in defective mesoderm and endoderm formation,
disrupted organiser development and mis-positioning of the AP axis (Gritsman et aL, 1999). Unsurprisingly, this phenotype is identical to that observed in cyc!sqt double mutants, which lack functional nodal ligands (Feldman et aL, 1998), and oep'^' mutants are unresponsive to the over expression o f nodal genes (Gritsman et aL,
1999).
The Nodal signal is transduced from the membrane bound receptors to the nucleus by Smad proteins (Heldin et aL, 1997). In response to the activation of type I receptors by TGF13 signalling, Smad proteins are phosphorylated. Smad2 and SmadS are specific to TGFB (except BMP) and activin response, and Smadl, Smad5 and Smad8
are associated with BMP signalling. Smad4 appears to be an essential component of
the transduction o f all TGFBs, including BMP, by forming heterodimers with other Smads. Smad6, Smad? and Smad9 have inhibitory functions (Baker & Harland,
1997; Kretzschmar & Massagué, 1998).
Figure 1.5. The progression of the Nodal pathway.
This figure shows the sequential molecular progression between the major components of the Nodal signalling pathway from ligand through to target genes. Mouse components are on the left, zebrafish on the right. Where homologous genes between species have different names, these are indicated in a similar colour.
Zebrafish have two Nodal ligands (Cyclops and Squint) compared to one in mouse. The receptors and co-receptors are similar in both species. Smad proteins are involved in transduction and also transcription o f target genes, on co-operation with transcription co-factors.
Target genes o f the Nodal pathway include those encoding the Nodal ligands themselves (thus the pathway up-regulates itself), goosecoid and the asymmetrically expressed activin {Lefty2 in mouse). The target genes listed are an example and many other genes are also implicated to be directly regulated by the Nodal pathway.
Figure 1.5
MOUSE
Medal Lefty2
I
AciRllA Alk4 C n ete AclKfIB AIK/ 0 « p
I
Sm ad2 Sm ad3 Sm ad4
;
I A S I? (F iix H l)
I
Nodal Lefty2G ee LIGAND(S) i RECEPTORS/ CO-RECEPTORS
I
INTRACELLULAR TRANSDUCTION TRANSCRIPTION CO-FACTORSi
TARGET GENES
ZEBRAFISH
Antivin---S^uiet
I
ActRIIA Alk4 Oep AcrRIIB TARAM A
I
S m ad ? Sm ad3 Sm*d4
Schm »tx|tur (F oxH I)
I
mesoderm is absent, (although other mesodermal tissue is present (Waldrip et aL, 1998). A double heterozygote knockout mouse lacking partial Nodal and Smad2 activity shows a phenotype similar to zebrafish oep mutants, including asymmetry defects and cyclopia (Nomura & Li, 1998). A mouse knockout of Smad4, the ‘global’ Smad protein causes a very dramatic phenotype that may result from consequential disruption of multiple signalling pathways (Sirard et aL, 1988).
Zebrafish smad2 and smad4 are ubiquitously expressed in early cleavage and early blastula stages (maternal expression) and expression of both is maintained through gastrulation. Zebrafish smad3 is also maternally expressed, and zygotic expression does not commence until tailbud stage (Dick et aL, 2000).
1.5.2.3 Nodal activity and forebrain development.
Following organiser development, mesendoderm formation and establishment o f the
AP axis, nodal genes are expressed in the prechordal plate where Nodal proteins have a function in DV specification o f cells during forebrain development. Homeobox
transcription factors Nk2.1a and Nk2.1b are homologues o f mouse Nkx2.1/TTF1. These genes act as good markers o f DV identity within the zebrafish forebrain; nk2.1a is expressed in the ventral diencephalon/hypothalamic tissue, and nk2.1b is expressed primarily in the ventral telencephalon. Expression of both these genes is lost in oep'^' mutant embryos (Rohr et a l, 2001).
It is unlikely Nodal activity in the prechordal plate induces nk2.1 gene expression directly, but the nk2.1 gene in mouse has been shown to be inducible by Shh (Gaiano et aL, 1999 and section 6,2.5 in this thesis), and another nk gene, nk2.2, can be induced ectopically by shh over-expression in zebrafish (Barth & Wilson, 1995). Experimental evidence suggests that nodal activity (specifically Cyc) can induce shh expression, at least in the ventral neural tube (Müller et al., 2000), while Cyclops and Hh activity are seen to be interacting synergistically in the specification of primary motor neurons (Odenthal et aL, 2000).
Ectopic expression of shh is able to restore lost nk2. lb expression in the telencephalon o f oep'^' mutants, but not the diencephalic expression o f nk2.Ia (Rohr et aL, 2001). This indicates that nodal signalling may be acting upstream of hedgehog signalling for
some of its activity, but independently of hedgehog for other functions.
from the prechordal plate at the anterior extreme of the embryo that leads to the fused- eye phenotype (Varga et al., 1999), analogous to the hum an condition holoprosencephaly, which is also associated with defects in NODAL signalling (Gripp et aL, 2000). The failure of correct anterior cell migration is discussed in more detail in section 6.2.2, in relation to the silberblick mutation.
1.6 The endocrine system.
The endocrine system is a communication network relaying information between cells and organ systems across the body. The endocrine system works hand-in-hand with the nervous system to maintain homeostasis (an internal steady state o f the body maintained by regulation of blood pressure, nutrition, metabolism, excretion, water retention and salt balance), to provide an appropriate physiological response to external stimuli and to regulate growth, development and reproduction.
Activity of the endocrine system can be reduced to three functional units: glands, hormones and target cells. Glands release hormones into the blood in response to a number of mechanisms including direct stimulation from nerve cells, as a response to
hormones detected in the blood or by direct monitoring of the physiological internal environment, e.g. concentration of ions in the plasma.
Hormones, the messenger molecules of the endocrine system, can be divided into two groups: peptide hormones, composed o f amino acids, and steroid hormones, synthesised from lipids. The two groups differ not only in composition but also by mode of action.
Steroid hormones are able to pass through the cell membrane o f the target cell and enter the cytoplasm. The hormone will bind then bind a receptor complex that enters the nucleus to activate transcription. Generally steroid hormones generate a response more slowly than peptide hormones, as they require the transcription and translation of genes to generate the desired physiological response.
The third functional unit of the endocrine system is a target cell. These are cells that carry receptors to the hormone signal, be it on the cell surface (for peptide hormones) or intracellular receptors (for steroid hormones). Target cells may lie in other endocrine organs, allowing communication within the endocrine system, or in specialised cells within organs such as the kidney, controlling water retention, or in vast numbers of cells throughout the body that respond to the secretion o f growth hormone.
Function of the endocrine system relies heavily on the principle of negative feedback. Many hormones, including all hypothalamic hormones, are secreted when levels in the blood are detected to be below a threshold level (specific to each hormone), and secretion is inhibited when levels of circulating hormone are high. The hormone being detected in the blood may not be the same hormone that is being controlled by the negative feedback loop, as will become apparent when the hormones secreted by the hypothalamus are discussed in more detail in section 1.7.1.
1.7 Organs of the endocrine system.
The teleost endocrine system is very similar to the human system (illustrated in figure
1.6). A description of some of the major glands within the endocrine system follows. The endocrine system acts downstream o f the nervous system, altering the physiological state of the body to cope with environmental demands as observed by sensory organs and perceived by the brain. (The endocrine system can also feed back to the brain altering perception and behaviour, discussed later). Because o f the functional relationship between the central nervous system and the endocrine system, a communication link between then is of great importance. This is largely achieved by the hypothalamus, a neural structure lying ventrally in the zebrafish forebrain. This brain tissue also fimctions within the endocrine system.
The pituitary gland is widely referred to as the ‘master’ organ o f the endocrine system, as many o f its hormone secretions directly effect the secretions o f other glands. This point is entirely valid, however the hypothalamus directly controls secretions from the pituitary gland and so directs the activity of the master gland. The relationship between these two organs is referred to as the hypothalamic-pituitary axis.
1.7.1 The hypothalamus.
Figure 1.6. The endocrine system of Human and Fish.
Panel A shows the major organs of the endocrine system in human males and females. Panel B shows the major tissues of the teleost endocrine system.
Panel A is adapted from Purves et aL, 2001.