Structure-Function Relationships of the Viral RNA-dependent
RNA Polymerase
FIDELITY, REPLICATION SPEED, AND INITIATION MECHANISM DETERMINED BY A RESIDUE
IN THE RIBOSE-BINDING POCKET
*Received for publication, October 27, 2006, and in revised form, March 29, 2007 Published, JBC Papers in Press, March 29, 2007, DOI 10.1074/jbc.M610090200
Victoria S. Korneeva and Craig E. Cameron1
From the Department of Biochemistry and Molecular Biology, Pennsylvania State University, University Park, Pennsylvania 16802
Studies of the RNA-dependent RNA polymerase (RdRp) from poliovirus (PV), 3Dpol, have shown that Asn-297 permits this enzyme to distinguish ribose from 2ⴕ-deoxyribose. All animal RNA viruses have Asn at the structurally homologous position of their polymerases, suggesting a conserved function for this residue. However, all prokaryotic RNA viruses have Glu at this position. In the presence of Mg2ⴙ, the apparent affinity of
Glu-297 3Dpol for 2ⴕ-deoxyribonucleotides was decreased by 6-fold relative to wild type without a substantial difference in the fidel-ity of 2ⴕ-dNMP incorporation. The fidelity of ribonucleotide misincorporation for Glu-297 3Dpol was reduced by 14-fold rel-ative to wild type. A 4- to 11-fold reduction in the rate of ribo-nucleotide incorporation was observed. Glu-297 PV was unable to grow in HeLa cells due to a replication defect equivalent to that observed for a mutant PV encoding an inactive polymerase. Evaluation of the protein-(VPg)-primed initiation reaction showed that only half of the Glu-297 3Dpol initiation complexes were capable of producing VPg-pUpU product and that the overall yield of uridylylated VPg products was reduced by 20-fold relative to wild-type enzyme, a circumstance attributa-ble to a reduced affinity for UTP. These studies identify the first RdRp derivative with a mutator phenotype and provide a mech-anistic basis for the elevated mutation frequency of RNA phage relative to animal RNA viruses observed in culture. Although protein-primed initiation and RNA-primed elongation com-plexes employ the same polymerase active site, the functional differences reported here imply significant structural differ-ences between these complexes.
RNA viruses cause a variety of acute and chronic diseases in humans: common cold, summer flu, hepatitis, severe acute res-piratory syndrome, and liver cancer, to name a few (1–5). The genomes of these viruses are transcribed and replicated by a virus-encoded RNA-dependent RNA polymerase (RdRp)2(4, 6,
7). Like other viral polymerases, the RdRp represents an impor-tant target for antiviral drug development (8 –11). The RdRp from poliovirus (PV), 3Dpol, has emerged as an important model for understanding the structure, function, and mecha-nism of this class of nucleic acid polymerases (12–15).
PV 3Dpol will incorporate a ribonucleotide (rNMP) with an incorrect base at a frequency of⬃10⫺7to 10⫺4(16, 17). How-ever, this enzyme is much more tolerant of nucleotides with an incorrect sugar configuration. Both 2⬘- and 3⬘ -deoxynucleoti-des (dNMPs) are incorporated at a frequency of⬃10⫺2(15). It is known that incorporation of more than one incorrect ribo-nucleotide per PV genome decreases the specific infectivity of the RNA (16, 18). Whether or not 2⬘-dNMP incorporation has an effect on viral RNA infectivity is not known.
Several factors likely limit 2⬘-dNMP incorporation into the genomes of RNA viruses of eukaryotes. First, dNTP pools in cells are thought to be low, in the 5–30Mrange (19). Low
dNTP levels are maintained by regulating the activity and local-ization of ribonucleotide reductase (20, 21). In the G1-phase of the cell cycle, very little ribonucleotide reductase holoenzyme exists due to the localization of one subunit in the nucleus and the other in the cytoplasm. During the S-phase of the cell cycle, the ribonucleotide reductase holoenzyme is distributed in both cytoplasm and nucleus; allosteric control mechanisms would likely permit only the nuclear enzyme to function catalytically (20 –22). If nuclear localization of active ribonucleotide reduc-tase prevents eukaryotic RNA virus polymerases from having to select against utilization of 2⬘-dNTPs, then prokaryotic RNA virus polymerases may require a more stringent selection mechanism.
Asn-297 is the primary determinant of ribose specificity in PV 3Dpol (14). This residue is located in the ribose-binding pocket and hydrogen bonds to the ribose 2⬘-OH (Fig. 1A). The capacity for Ala-297 derivative to distinguish between ATP and 2⬘-dATP incorporation is reduced by 10-fold relative to wild-type 3Dpol (14). Asn is present at the structurally homologous position in all plant and animal RNA virus polymerases (Fig. 1B), suggesting that this residue functions in ribose selection for these enzymes as well. Curiously, RNA phage polymerases have a Glu at this position (Fig. 1B), consistent with a difference in ribose/2⬘-deoxyribose selection for these enzymes.
We show that Glu-297 3Dpol does not have a more stringent selection against 2⬘-dNMP incorporation than wild-type (WT) 3Dpol, although the affinity for 2⬘-dNTPs is reduced relative to WT 3Dpol. However, nucleotide incorporation by Glu-297
*This work was supported by NIAID, National Institutes of Health Grant AI053531 (to C. E. C.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1To whom correspondence should be addressed. Tel.: 814-863-8705; Fax:
814-865-7927; E-mail: [email protected].
2The abbreviations used are: RdRp, RNA-dependent RNA polymerase; PV,
3Dpol was slower and less faithful than WT 3Dpol. Glu-297 3Dpol represents the first RdRp derivative with a mutator phe-notype. Glu-297 PV was inviable due to an RNA synthesis defect in cells that was on par with that of a catalytically inactive 3Dpol derivative. This phenotype could be explained by a severe reduction in the yield of product produced by protein-primed initiation of RNA synthesis. We conclude that a requirement for decreased replication speed and/or incorpora-tion fidelity imposes a constraint on the evoluincorpora-tion of the poly-merase active site that restricts the variety of initiation mecha-nisms that can be employed. Thus phage polymerases employ strictly ade novo mechanism for initiation, whereas animal RNA viruses can utilizede novoand protein-primed mecha-nisms for initiation (Fig. 1B). The capacity of a single 3Dpol derivative to exhibit significant differences between RNA- and protein-primed RNA synthesis suggests that the structural organization of the corresponding elongation complexes is dif-ferent. We propose that inhibitors specific for the PV 3Dpol initiation complex may be more efficient than those targeting the elongation complex.
EXPERIMENTAL PROCEDURES
Materials—[␥-32P]ATP (⬎7000 Ci/mmol) was from MP
Bio-medicals (formerly ICN); [␣-32P]UTP (⬎6000 Ci/mmol) was from Amersham Biosciences; DNA oligonucleotides were from Integrated DNA Technologies, Inc.; T4 polynucleotide kinase, Deep Vent DNA polymerase, and restriction enzymes were from New England Biolabs, Inc.; Sephadex G-25 was from Sigma; all nucleotides (ultrapure solutions) were from Amer-sham Biosciences; RNA oligonucleotides were from Dharma-con Research, Inc. (Boulder, CO); synthetic VPg peptide was from Alpha Diagnostic International (San Antonio, TX); all other reagents were of the highest grade available through Sigma, Fisher, or VWR.
Construction, Expression, and Purification of 3Dpol
Derivatives—Mutations were introduced into a modified
3Dpol-coding sequence by using overlap-extension PCR and expressed inEscherichia coliby using a ubiquitin fusion system as described previously (23). In brief, the Glu-297 clone was produced by overlap-extension PCR with oligonucleotides 1 and 7, and 2 and 8 (Table 1), with pET26Ub-3D-BPKN-I92T as template. The final product of second PCR was purified and digested with PstI and NheI and ligated into pET26Ub-3D-BPKN-I92T, which had been digested with the same enzymes. Double and triple mutants (Met-288, 297; Asn-290, Glu-297; and Met-288, Asn-290, Glu-297) were constructed in the same way using oligonucleotide primers 1, 2, and 9 –14 (Table 1), with pET26Ub-3D-BPKNP-I92T plasmid used as template. This plasmid has a silent PmlI site in the 3Dpol coding sequence. Mutations were confirmed by DNA sequencing (Nucleic Acid Facility, Pennsylvania State University). 3Dpol derivatives were expressed and purified as described previously (23). The concentration of the Glu-297 3Dpol was determined by measuring absorbance at 280 nm in 6Mguanidine chloride
solution, using a calculated WT 3Dpol extinction coefficient (71,840M⫺1cm⫺1). For double and triple mutants poly-r(U)
activity assays were performed with partially purified enzymes recovered from ammonium sulfate pellets.
Construction, Expression, and Purification of 3CD—Viral
protein 3CD was expressed and purified as described previously (24).
Transcription of 61-nt 2C-cre— 61-nt 2C-cre RNA was
tran-scribed and gel-purified by denaturing PAGE as detran-scribed pre-viously (24).
Purification, 5⬘-32P Labeling and Annealing of sym/sub—RNA
oligonucleotides were purified, labeled, and annealed as described previously (12).
Poly-r(U) Polymerase Activity Assay—Poly-r(U) activity
assays were performed as described previously with minor modifications (14). Briefly, reactions contained 50 mMHEPES,
pH 7.5, 10 mM2-mercaptoethanol, 5 mMMgCl2, 60MZnCl2,
500MUTP, 0.2Ci/l [␣32-P]UTP, 2MdT15/0.2M
poly-rA467 primer-template complex, and 500 ng of 3Dpol. Reac-tions were carried out in a total volume of 25l at 30 °C for 5 min and then quenched by mixing 5l with an equal volume of 0.1MEDTA. 5l of the quenched reactions was spotted onto
DE81 filter paper discs and dried completely. The discs were washed three times for 10 min in 250 ml of 5% dibasic sodium phosphate and rinsed in absolute ethanol. Bound radioactivity was quantitated by liquid scintillation counting in 5 ml of EcoScint scintillation fluid (National Diagnostics).
3Dpol Kinetic Assays—Reactions contained 50 mMHEPES,
pH 7.5, 10 mM2-mercaptoethanol, 5 mMMgCl2, 60MZnCl2,
500MNTPs (unless specified otherwise), 1Msym/sub (0.5
Mduplex), and 1M3Dpol. Reactions were quenched by
addi-tion of EDTA to a final concentraaddi-tion of 50 mM. 3Dpol was
diluted immediately prior to use in 50 mMHEPES, pH 7.5, 10
mM2-mercaptoethanol, 60MZnCl2, and 20% glycerol. The
volume of enzyme added to any reaction was always less than or equal to one-tenth of the total volume. All reactions were per-formed at 30 °C.
Rapid Chemical-Quench-Flow Experiments—Rapid mixing/
quenching experiments were performed using a Model RQF-3 chemical-quench-flow apparatus (KinTek Corp., Austin, TX). All experiments were performed at 30 °C, with the temperature controlled by a circulating water bath. 3Dpol-sym/sub complex in 50 mMHEPES, pH 7.5, 10 mM2-mercaptoethanol, 60M
ZnCl2, and 5 mMMgCl2was rapidly mixed with the nucleotide
substrate in the same buffer, and the reactions were quenched either by addition of 0.5MEDTA to a final concentration of 0.3 Mor by addition of 2NHCl to a final concentration of 1.2N.
Immediately after addition of HCl, the solution was neutralized by addition of 3MKOH in 1MTris, pH 8.0.
Product Analysis—Products were analyzed by denaturing
PAGE. An equal volume of loading buffer, 5l (90% formam-ide, 0.025% bromphenol blue, and 0.025% xylene cyanol, 1⫻ TBE) (89 mMTris, 89 mMboric acid, 2 mMEDTA) was mixed
with 5l of the quenched reaction mixtures and heated at 65 °C for 2–5 min. prior to loading 5l on a denaturing 23% polyac-rylamide gel containing 1⫻TBE and 7Murea. Electrophoresis
was performed at 90 watts. Gels were visualized by using a PhosphorImager and quantified by using ImageQuaNT soft-ware (Molecular Dynamics).
Data Analysis—Data were fit by non-linear regression using
Time courses at fixed nucleotide concentration were fit to Equation 1,
关Product兴⫽A⫻e⫺kobs⫻t⫹C (Eq. 1)
whereAis the maximal concentration of product formed,kobs is the observed first-order rate constant describing product for-mation,tis the time, andCis a constant. The apparent disso-ciation constant (Kd,app) and maximal rate for nucleotide incor-poration (kpol) were determined using Equation 2.
kobs⫽kpol⫻关NTP兴/共Kd,app⫹关NTP兴兲 (Eq. 2)
Kinetic Simulations—Kinetic simulations were performed by
using KinTekSim version 2.03 (KinTek Corp., Austin TX). Agreement between the experimental data and kinetic simula-tions was determined by visual inspection.
Construction of Mutated Viral cDNA Clones and Mutated
Replicons—To introduce a mutation into the 3Dpol coding
sequence of viral cDNA, overlap extension PCR was performed with oligonucleotides 1 and 3 and the appropriate protein expression plasmid as template. PCR products were purified and digested with AvrII and AflII. The digested PCR product was ligated into pUC18-BglII-EcoRI-3CD vector, a subclone of the viral cDNA (14, 24). From this vector, the fragment between BglII and EcoRI was cloned into the viral cDNA plasmid (pMoRA) (25). To introduce mutations into the subgenomic replicon the fragment between the BglII and AvrII sites from the mutated pUC18-BglII-EcoRI-3CD subclone vector was ligated into pRLucRA (also known as pRLuc31-rib⫹polyAlong (25)). DNA sequencing was used to verify the integrity of all clones.
Infectious Center Assays—RNA transcripts were produced
from the pMoRA-WT and pMoRA-Glu-297 plasmids after lin-earization with EcoRI. Transcription reactions, typically 20l, consisted of 350 mMHEPES, pH 7.5, 32 mMmagnesium acetate,
40 mMdithiothreitol, 2 mMspermidine, 28 mMNTPs, 0.5g of
template, and 0.5g of T7 RNA polymerase. Reactions were incubated at 37 °C for 3 h followed by removal of magnesium pyrophosphate. DNase treatment with RQ1 DNase (Promega,
Madison, WI) was used to remove the template; lithium chlo-ride precipitation was used to remove unincorporated nucleo-tides. RNA concentration was calculated by measuring absorb-ance at 260 nm, assuming anA260of 1 was equivalent to 40 g/ml. HeLa cells were propagated in Dulbecco’s modified Eagle’s medium/F-12 media (Invitrogen) supplemented with 10% (v/v) fetal bovine serum (Invitrogen), and 1% (v/v) penicil-lin and streptomycin (Invitrogen), always keeping the cultures between 20 and 80% confluency. Subconfluent monolayers of HeLa cells were detached from the culture flasks by trypsin treatment, washed with 1⫻phosphate-buffered saline, and cell number was adjusted to 1.2⫻106cells/0.4 ml in phosphate-buffered saline. Cell suspensions (400l) were mixed with 5g of RNA in a microcentrifuge tube, transferred to an electropo-ration cuvette (0.2-cm gap width, Bio-Rad), and subjected to an electric pulse at 500 microfarads and 0.13 V using a GenePulser system (Bio-Rad). 400l of electroporated cells was plated onto appropriate wells of a 6-well plate containing a monolayer of HeLa cells plated with 5⫻105cells the day before. For WT controls, 1/100, 1/1000, and 1/2000 dilutions were plated. Cells were allowed to adsorb to the plate for 1 h at 37 °C, and then the medium/phosphate-buffered saline was aspirated. Cells were then covered with 3 ml of a mixture of 1⫻Dulbecco’s modified Eagle’s medium/F-12 plus 10% fetal bovine serum, 1% penicillin and streptomycin, and 1% low melting point agarose (American Bioanalytical). Plates were then incubated at either 32 °C or 37 °C for 2–3 days. The agarose overlay was removed by using a spatula. Wells were stained with crystal violet, and viral plaques were counted.
Serial Passage and Reverse Transcription-PCR—For serial
passage of Glu-297 PV 1⫻106HeLa cells were electroporated with 5g of RNA (no RNA as negative control), essentially as described above for the infectious center assay. The electropo-rated cells were then plated onto a monolayer of HeLa cells (plated with 5⫻105cells the previous day onto 6-well plates). Plates were incubated at 37 °C. Within 2 days, cells infected with WT RNA started showing signs of cytopathic effect, whereas for Glu-297 infected cells started rounding up only on
TABLE 1
Oligonucleotides used in this study
Oligonucleotide number Oligonucleotide name Sequencea,b
1 3D-PV-AvrII-rev 5⬘-CCT GAG TGT TCC TAG GAT CTT TAG T-3⬘
2 3D-PV-PstI-for 5⬘-GGA GTG ATA ACA GGT TCT GCA GTG GGG TGC GAT-3⬘
3 3D-PV-AflII-for 5⬘-AAC GAT CCC AGG CTT AAG ACA GAT TTT GAG-3⬘
4 3D-BamHI-rev 5⬘-GCG GGA TCC TTA CTA AAA TGAGTC AAG CCA ACG GCG GTA-3⬘
5 3D-SacII-for 5⬘-GCG CCG CGG TGG AGG TGA AAT CCA GTG GAT GAG A-3⬘
6 3Dseq500 5⬘-AGG TTG AGC AGG GGA AA-3⬘
7 3D-PV-N297E-for 5⬘-GGT ACC TCA ATT TTT GAA TCA ATG ATT AAC-3⬘
8 3D-PV-N297E-rev 5⬘-GTT AAT CAT TGA TTC AAA AAT TGA GGT ACC-3⬘
9 3D-PV-S288M-N297E-for 5⬘-AAG GGC GGT ATG CCA ATG GGC TGT TCA GGT ACC TCA ATT TTT GAA TCA ATG ATT AAC AAC-3⬘
10 3D-PV-S288M-N297E-rev 5⬘-GTT GTT AAT CAT TGA TTC AAA AAT TGA GGT ACC TGA ACA GCC CAT TGG CAT ACC GCC CTT-3⬘
11 3D-PV-C290N-N297E-for 5⬘-GGT ATG CCA TCT GGC AAC TCA GGT ACC TCA ATT TTT GAA TCA ATG ATT AAC AAC-3⬘
12 3D-PV-C290N-N297E-rev 5⬘-GTT GTT AAT CAT TGA TTC AAA AAT TGA GGT ACC TGA GTT GCC AGA TGG CAT ACC-3⬘
13 3D-PV-S288M-C290N-N297E-for 5⬘-AAG GGC GGT ATG CCA ATG GGC AAC TCA GGT ACC TCA ATT TTT GAA TCA ATG ATT AAC AAC-3⬘
14 3D-PV-S288M-C290N-N297E-rev 5⬘-GTT GTT AAT CAT TGA TTC AAA AAT TGA GGT ACC TGA GTT GCC CAT TGG CAT ACC GCC CTT-3⬘
a
Codons that introduce amino acid residue change in 3Dpol sequence are in bold.
b
day 4. Cells in mock control looked healthy at that time. For the first passage, 100l of media from the transfected cell were taken on day 2 and transferred onto a fresh monolayer of HeLa cells. Cells infected with the media from WT virus died within 1 day and for Glu-297 within 2 days. Mock control cells did not show any signs of cytopathic effect. 100l of media from day 2 (passage 1) was again transferred onto a fresh monolayer for passage 2. The infected cells from passage 2 showed the same kinetics for cytopathic effect. Passage 3 was done in the same manner as the first two. On the second day, when both WT and Glu-297 PV-infected cells were dead, 1 ml of media was col-lected and viral RNA was extracted with a NucleoSpin nucleic acid purification kit (Macherey-Nagel) according to the manu-facturer’s manual. The 3Dpol cDNA was prepared from puri-fied viral RNA by reverse transcription with Endo-Free RT (Ambion) with oligonucleotide 4 (Table 1). The resulting DNA product was then PCR-amplified using SuperTaq DNA poly-merase (Ambion) and oligonucleotides 4 and 5 as primers. The PCR product for the N297E 3Dpol gene was sequenced with oligonucleotide 6 as primer. Codon GAA that encodes Glu-297 was mutated to GGA, encoding Gly.
Subgenomic Replicon Assays—RNA transcripts were
pro-duced from the pRLucRA plasmids after linearization with ApaI. RNA concentration was calculated as described above. HeLa cells (1.2⫻106) were electroporated with subgenomic replicon (5g) essentially as described above for infectious cen-ter assay. Immediately afcen-ter electroporation 400l of cells was mixed with 6 ml of prewarmed media (Dulbecco’s modified Eagle’s medium/F-12, 10% fetal bovine serum, and 1% penicillin and streptomycin); 0.5 ml of this mixture was aliquoted into a 1.7-ml Eppendorf tube per single time point and transferred into either a 32 °C or 37 °C incubator. At fixed time points, cells were pelleted by centrifugation at 14,000 rpm for 2 min. Cells were lysed in 100l of 1⫻cell culture lysis reagent (Promega). Lysates were left on ice until luciferase and protein concentra-tion assays were performed at which point the lysates were cen-trifuged for 2 min at 14,000 rpm to remove cellular debris and nuclei. Lysates were assayed for luciferase activity by mixing 10 l of lysate supernatant with 10l of luciferase assay substrate (Promega) and quantifying light production for 10 s by using a Junior LB 9509 luminometer (Berthold). Relative light units were adjusted to total protein concentration in the samples. Protein concentration was measured at 595 nm using the Brad-ford reagent (Bio-Rad).
VPg Uridylylation Assay—VPg uridylylation reaction
mix-tures contained 1M3Dpol, 1M61-nt 2C-cre, and 5MVPg
in reaction buffer (50 mMHEPES, pH 7.5, 5 mMmagnesium
acetate, 10MUTP, and 0.04M[␣-32P]UTP (6000 Ci/mmol)).
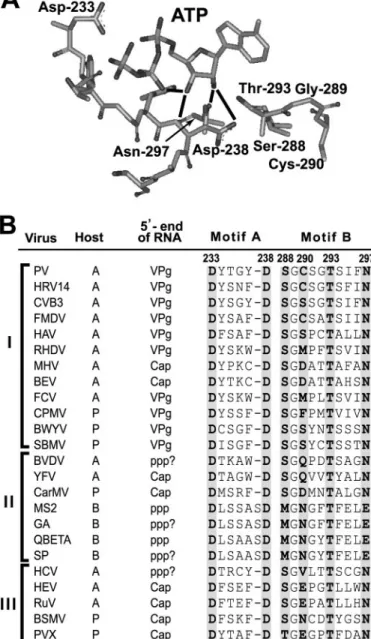
FIGURE 1.Differences in ribose/2ⴕ-deoxyribose specificity for RNA phage polymerases relative to animal and plant RNA virus polymerases predicted from structure/sequence comparison.A, structural model for the ribose-binding pocket of the RdRp from PV, 3Dpol (14). The 3Dpol ribo-nucleotide-binding pocket is shown with bound ATP. This structural model is based on homology modeling, because a crystal structure for this complex is still not available for any RdRp (14). This pocket is composed of residues from conserved structural motifs A (Asp-238) and B (Ser-288 to Asn-297). The extensive hydrogen bonding network shown links the position of the triphos-phate (catalytic efficiency) to the nature of the bound nucleotide, that is, whether or not a correct base and sugar configuration are present. Asn-297 permits the enzyme to distinguish between ribose and 2⬘-deoxyribose by hydrogen bonding to the 2⬘-OH group of ribose. A 3Dpol derivative contain-ing Ala at this position instead of Asn fails to distcontain-inguish rNTPs from 2⬘-dNTP as well as wild-type 3Dpol (14). The image was created by using the program WebLab Viewer (Molecular Simulations Ins., San Diego, CA).B, conserved Asn in animal and plant RNA virus polymerases is a Glu in RNA phage polymerases. RNA virus polymerases can be organized into three supergroups: I, II, and III. Representative viruses, their host (A, animal;P, plant; andB, bacteria) and mechanism of initiation of RNA synthesis based on the structure of the 5⬘-end of the genome (VPg, protein primed;Capandppp,de novo) are indicated. In those instances in which the 5⬘-end has not been analyzed directly, the struc-ture of the 5⬘-end appears with aquestion markand represents the most logical structure based on available biochemical data or relationship to other viruses. Residues 233 and 238 (3Dpol numbering) of conserved structural motif A are conserved in all animal and plant RNA virus polymerases. Residues 288, 293, and 297 (3Dpol numbering) of conserved structural motif B are
All reactions were adjusted to a final NaCl concentration of 20 mM. 3Dpol and 3CD were diluted immediately prior to use in
buffer containing 50 mMHEPES, 10 mM2-mercaptoethanol, and 20% glycerol. First, VPg was mixed with 2C-cre RNA in 10 l of the reaction buffer described above. This mixture was incubated at 30 °C for 5 min prior to addition of 3CD to a final concentration of 1M. After addition of 3CD the reaction
mix-ture was incubated at 30 °C for 5 min prior to adding 3Dpol to a final concentration of 1 M. The uridylylation reaction was
allowed to proceed for 30 min, after which 5l of reaction were quenched by addition of an equal volume of 100 mMEDTA in a loading dye (90% formamide, 1⫻ TBE, 0.025% bromphenol blue). Products were separated by Tris-Tricine SDS-PAGE. Gels contained 15% acrylamide and 0.4% bisacrylamide. The cathode buffer (upper chamber) contained 0.1MTris, 0.1 M
Tricine, and 0.1% (w/v) SDS; the anode buffer contained 0.2M
Tris, pH 8.9. Gels were run at 80 watts. Products were visualized by using a PhosphorImager and quantified by using Image-Quant software.
RESULTS
Rationale—Previous studies identified Asn-297 as a key
player in ribonucleotide selection by the RdRp from PV, 3Dpol. Asn-297 hydrogen bonds to the 2⬘-hydroxyl of the incoming ribonucleotide (rNTP) (Fig. 1A). Changing Asn-297 to Ala pre-vented 3Dpol from distinguishing an rNTP with correct base pairing capacity from a 2⬘-dNTP with correct base pairing capacity, a lethal change for PV (Table 2) (14). Asn-297 is con-served in all plant and animal RNA virus polymerases; however, this residue is a Glu in RNA phage polymerases (Fig. 1B). This study was initiated to determine the consequence(s) of this dif-ference in the ribose binding pocket of eukaryotic and prokary-otic RNA viruses.
Glu-297 3Dpol Incorporates 2⬘-Deoxyribonucleotides as
Effi-ciently as WT 3Dpol—3Dpol-coding sequence was changed to
encode a glutamic acid at position 297 (Glu-297 3Dpol). This derivative was purified to⬎90% purity exactly as described for WT 3Dpol, suggesting no significant change in the structure of
the enzyme (23). In addition, this derivative retained at least half of the poly(rU) polymerase activity of WT 3Dpol (Table 3). To evaluate the kinetics and mechanism of 3Dpol, we use a 10-nt symmetrical RNA substrate referred to as sym/sub (S/S). S/S-U, for example, has uracil as the first templating base (Fig. 2A) (12). 3Dpol binds tightly to the duplex portion of this sub-strate and can utilize either 3⬘-end as a primer for nucleotide addition. The rate constant for dissociation of Glu-297 3Dpol was 0.0001 s⫺1, corresponding to a half-life for the complex of ⬃2 h as observed for WT 3Dpol (12).
We evaluated the dependence of the rate constant for AMP incorporation on ATP concentration. For this experiment, 2 M3Dpol was first incubated with 2M5⬘-32P-labeled S/S-U (1
M duplex) in reaction buffer to establish the enzyme-RNA
complex. The assembled complex was then rapidly mixed with ATP of the appropriate concentration in reaction buffer, as described under “Experimental Procedures.” Mixing resulted in a 2-fold dilution. The reaction was stopped at various times by addition of EDTA to 0.3Mfinal concentration. The product,
11-mer RNA, was separated from the substrate, 10-mer RNA, by electrophoresis through a denaturing, 23% polyacrylamide gel. Substrate and product were visualized by using a Phosphor-Imager and quantitated by using the ImageQuant software (Molecular Dynamics) (Fig. 2B).
TABLE 2
Glu-297 3Dpol incorporates 2ⴕ-deoxyribonucleotides as efficiently as WT 3Dpol
Enzyme Substrates Kinetic parameters
Nucleic acid Nucleotide Kd,app kpol kpol/Kd,app
M s⫺1 s⫺1/
M
sym/sub-U
GCAUGGGCCC
WTa CCCGGGUACG ATP 130⫾20 90⫾10 0.7
Glu-297 530⫾120 8⫾1 0.02
Ala-297b 180⫾30 5⫾1 0.03
WTa dATP 280⫾60 0.8⫾0.1 0.003
Glu-297 1700⫾400 0.4⫾0.02 0.0002
Ala-297b 260⫾30 0.2⫾0.05 0.0008
WTa GTP 210⫾70 0.01⫾0.001 0.00005
Glu-297 290⫾90 0.005⫾0.001 0.00002
sym/sub-A
GCUAGGGCCC
WTa CCCGGGAUCG UTP 100⫾10 270⫾20 2.7
Glu-297 380⫾80 70⫾6 0.2
WT dUTP 190⫾40 6⫾1 0.03
Glu-297 1100⫾200 2⫾0.2 0.002
aValues are taken from Ref. 26. bValues are taken from Ref. 14.
TABLE 3
Activity of Glu-297 3Dpol is not enhanced by introduction of residues conserved in phage RdRps
Poly-r(U) polymerase activity was measured in purified or partially purified prepa-rations of the indicated 3Dpol derivatives as described under “Experimental Proce-dures.” The specific activity is reported to one significant figure; the standard error of the measurement was⬍20%.
Enzyme Specific activity
pmol/min/g
WT 800
WTa
400
Glu-297 300
Met-288 and Glu-297a ⬍5
Asn-290 and Glu-297a
10 Met-288, Asn-290, and Glu-297a
5
The concentration of 11-nt product formed was plotted as a function of time and fit to a single exponential equation (Equa-tion 1), yielding the observed rate constant for AMP
incorpo-ration templated by U (Fig. 2C). The observed rate constants were then plotted as a function of ATP concentration, and the data were fit to a hyperbola (Equation 2), yielding an apparent dissociation constant for ATP,Kd,app, of 530⫾120M, and a maximal rate constant for AMP incorporation,kpol, of 8⫾1 s⫺1 (Fig. 2Dand Table 2). The ability to fit the data to a hyperbola is in good agreement with the minimal kinetic mechanism pro-posed for WT 3Dpol (Scheme 1) (26). TheKd,appvalue for ATP was increased by 4-fold, and the maximal rate constant for AMP incorporation by Glu-297 3Dpol was reduced by 11-fold relative to WT 3Dpol (Table 2).
Next, we evaluated the kinetics of dAMP incorporation for Glu-297 3Dpol. TheKd,appandkpolvalues for dAMP incorpo-ration into S/S-U were 1700⫾ 400Mand 0.4 ⫾ 0.02 s⫺1,
respectively (Table 2). Relative to WT 3Dpol, these values reflect a 6-fold increase in theKd,appvalue for dATP and a 2-fold decrease in thekpolvalue for dAMP incorporation. These data are consistent with Glu-297 3Dpol selecting against dAMP incorporation better than WT 3Dpol (Table 2). To show that this observation was not unique to dAMP incorporation, we evaluated dUMP incorporation into S/S-A. For Glu-297 3Dpol, theKd,appvalue for dUTP increased by 6-fold, and thekpolvalue decreased by 3-fold relative to WT 3Dpol (Table 2), consistent again with this derivative selecting against dNMP incorpora-tion better than WT 3Dpol. If we define the overall efficiency of nucleotide incorporation askpol/Kd,app, then the efficiency of dNMP incorporation relative to the efficiency of AMP incorpo-ration for Glu-297 3Dpol was essentially the same as WT 3Dpol (Table 2).
Glu-297 3Dpol exhibited a 14- to 35-fold reduction in the efficiency of rNMP incorporation relative to WT 3Dpol (Table 2). Unexpectedly, the fidelity of base selection (GMP incorpo-ration into S/S-U) by Glu-297 3Dpol was decreased by 10-fold relative to WT 3Dpol. This is the first RdRp derivative with a mutator phenotype.
Glu-297 PV Is Quasi-infectious—The rate constant for rNMP
incorporation by Glu-297 3Dpol was decreased by 4- to 11-fold relative to WT 3Dpol. Therefore, this mutant virus should reveal the impact, if any, of replication speed and fidelity on virus multiplication and fitness. Glu-297-encoding changes were engineered into the infectious cDNA for PV. RNA was produced byin vitrotranscription, and the infectivity of the RNA was evaluated by infectious center assay at both 37 °C and 32 °C (14). Glu-297 PV RNA did not produce virus at either temperature (Fig. 3A). Under the same conditions, WT PV RNA was able to establish a productive infection (Fig. 3A).
If the change of Asn-297 to Glu is indeed lethal, then viable virus will never be isolated. However, if this change permits some low level of replication (quasi-infectious), then viable virus may be recovered after serial passage. In this experiment, FIGURE 2.Kinetic analysis of Glu-297 3Dpol.A, RNA primer-template substrate
employed. A 10-nt self-complementary RNA referred to as sym/sub (S/S). Shown is S/S-U. The “U” indicates that uracil is the templating base.B, product analysis. In this experiment, 2MGlu-297 3Dpol was incubated with 2Msym/sub (1M duplex) and rapidly mixed with 200MATP. Reactions were quenched at vari-ous times up to 1 s by addition of EDTA. Reaction products were resolved by electrophoresis on a denaturing 23% polyacrylamide gel and visualized by using a PhosphorImager (Amersham Biosciences).C, concentration depend-ence of the kinetics of nucleotide incorporation. Experiments were per-formed as described above in the presence of: 50 (F), 100 (E), 250 (f), 500 (䡺), 1000 (Œ), or 2000M(‚) ATP. Thesolid linerepresents the fit of the data to a single exponential (Equation 1), yieldingkobsvalues of 0.6⫾0.03, 1.3⫾
0.1, 2.3⫾0.15, 3.53⫾0.3, 5.6⫾0.5, and 5.7⫾0.4 s⫺1, for 50 –2000
MATP, respectively.D, kinetic constants for AMP incorporation by Glu-297 3Dpol. Values forkobswere plotted as a function of ATP concentration. Thesolid line
represents the fit of data to a hyperbola (Equation 2), yielding values forKd,app
for ATP of 530⫾120Mandkpolof 8⫾1 s⫺ 1.
HeLa cells were electroporated with Glu-297 PV RNA and plated onto a fresh monolayer of HeLa cells. Plates were incu-bated at 37 °C. Every 2 days, a fraction of the media was used to infect a fresh monolayer. This process was repeated reitera-tively until the monolayer was killed in 2 days. This took three serial passages. The 3Dpol gene from this virus was amplified by using reverse transcription-PCR and sequenced. A single nucleotide substitution was observed that converted the GAA codon encoding Glu-297 to a GGA codon encoding Gly, a virus isolated by us previously (Fig. 3B) (27). Together, these data permit us to conclude that Glu-297 PV is quasi-infectious.
Glu-297 PV Is Severely Impaired for RNA Synthesis in Cells—
The polymerase domain (3D) has functions in the context of precursors that are unrelated to RNA elongation. For example, the 3CD precursor is a protease that processes the polyprotein into “mature” forms required for virus multiplication (7, 24, 28, 29). 3CD is an RNA-binding protein that binds to the cis-acting element required for initiation of genome replication, possibly recruiting 3Dpol to this location (30 –32). Finally, 3CD appears to function after RNA synthesis as well in a step related to virion maturation that does not require 3CD protease activity (33). To determine to what extent RNA synthesis was attenuated for Glu-297 PV, mutations encoding this change were engineered into the PV subgenomic replicon. This replicon consists of a PV
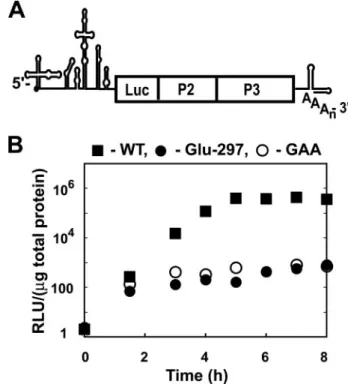
genome in which capsid-coding sequence has been replaced by a luciferase reporter gene (Fig. 4A), permitting RNA synthesis to be monitored indirectly by measuring luciferase activity (14). WT and Glu-297 PV replicons were evaluated at 37 and 32 °C (Fig. 4Band data not shown). Exponential replication of the WT PV replicon was observed for 5 h after transfection, leading to a⬎105-fold increase in luciferase activity. In contrast, only a 2-log increase in luciferase activity was observed for the Glu-297 PV replicon. This level of luciferase activity was also observed for the GAA PV replicon that encodes a polymerase completely lacking polymerase activity due to a change in the active site GDD motif to GAA (34, 35). Luciferase activity observed for the GAA PV replicon derives from translation of the transfected RNA. Such a substantial replication defect was not expected for Glu-297 PV given the significant polymerase activity associated with this Glu-297 3Dpol (Tables 2 and 3).
The Product Yield and Processivity of VPg-primed RNA
Syn-thesis by Glu-297 3Dpol Is Severely Impaired—Another aspect
of PV genome replication in which 3Dpol plays a crucial role is VPg uridylylation. VPg is a 22-amino acid-long peptide that is found covalently attached to the 5⬘-end of PV RNA. VPg or some precursor thereof is used to prime RNA synthesis (35– 38). Uridylylation of VPg leads to production of VPg-pUpU, FIGURE 3.Glu-297 PV is quasi-infectious.A, PV genomic RNA encoding
Glu-297 3Dpol is not infectious. An infectious center assay (see “Experimental Procedures”) was performed to evaluate the infectivity of WT or Glu-297 PV RNA after electroporation into HeLa cells. Virus was not recovered from cells containing Glu-297 PV at 37 °C or 32 °C.B, isolation of Gly-297 PV as a pseudo-revertant of Glu-297. After transfection of Glu-297 PV RNA into HeLa cells, a fraction of the media was transferred to a fresh monolayer every 2 days for 6 days at which point cytopathic effect was observed. Therefore, Glu-297 PV is quasi-infectious. Shown is the electropherogram and sequence for cDNA pre-pared from viral RNA by using reverse transcription-PCR. Theunderlined codon is for position 297 and encodes a Gly. The original sequence was GAA.
which serves as a primer for viral RNA synthesis (31, 36, 39). This reaction is catalyzed by 3Dpol, templated by an adenylate residue in the 2C-cre (cis-replicating element) by using a slide-back mechanism, and is stimulated by protein 3CD (31, 40, 41). This reaction can be mimickedin vitroas shown in Scheme 2.
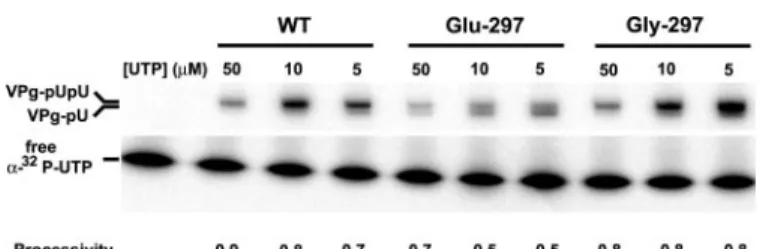
For WT 3Dpol, both VPg-pU and VPg-pUpU products are observed at UTP concentrations⬍5 M. Above 5 MUTP,
however, VPg-pUpU is the primary product (Fig. 5A). For Glu-297 3Dpol, VPg-pU is the primary product at low UTP concen-trations, and both VPg-pU and VPg-pUpU are still equimolar at concentrations of UTP as high as 300M(Fig. 5B). Quantitative
evaluation of the UTP concentration dependence of UMP incorporation into VPg showed that the maximal value of UMP incorporated by Glu-297 3Dpol was reduced by approximately
half relative to WT 3Dpol (Fig. 5,C and D). Moreover, the concentra-tion of UTP required for half-maxi-mal UMP incorporation (K0.5) by Glu-297 3Dpol was increased by 14-fold relative to WT 3Dpol (Fig. 5, CandD). These data suggest that the capacity for the Glu-297 3Dpol uridylylation complex to bind UTP is compromised significantly relative to WT 3Dpol, whereas the incorporation rate of Glu-297 3Dpol uridylylation complex was much less affected. These observa-tions are different from those made for the Glu-297 3Dpol RNA elongation complex where the incorporation rate constant (kpol) was affected more than UTP bind-ing (Kd,app) (Table 2).
The 5⬘-ends of PV plus and minus strands contain VPg-pUpU, likely the result VPg-pUpU produced at the 2C-cre being used as the primer for extension from the 3⬘-end of PV plus and minus strand RNAs (36, 37, 39, 42). Although WT 3Dpol pro-duced as much VPg-pUpU product at saturating concentrations of UTP (Fig. 5E), Glu-297 3Dpol produced half as much VPg-pUpU product, even at apparently saturating con-centrations of UTP (Fig. 5F). More-over, theK0.5value for UTP for pro-duction of VPg-pUpU by Glu-297 3Dpol was still 7-fold higher than that observed for WT 3Dpol. We conclude that both the overall prod-uct yield and the processivity of the Glu-297 3Dpol uridylylation com-plex is reduced relative to WT 3Dpol and that, when coupled with the reduced elongation rate of Glu-297 3Dpol, provides a biochemical explanation for the severe RNA synthesis defect observed biologically.
The Processivity of RNA-primed RNA Synthesis by Glu-297
3Dpol Appears Normal—The finding that processive
VPg-primed RNA synthesis was reduced for Glu-297 3Dpol sug-gested the possibility that processive RNA-primed RNA syn-thesis might also be reduced, contributing to the lack of RNA synthesis observed in cells. Processive RNA synthesis was eval-uated by using S/S-U, the same RNA substrate utilized for sin-gle-nucleotide-addition experiments, in the presence of all four ribonucleotides (125Meach). Upon rapid mixing of
Glu-297 3Dpol-S/S-U complex with all four nucleotides, for-mation of 11-, 12-, 13-, and 14-mer RNA products were observed (Fig. 6). The net rate constants for sequential incor-FIGURE 5.VPg-primed RNA synthesis by Glu-297 3Dpol is impaired.AandB, VPg uridylylation assay. The
reaction described in Scheme 2 was performed as described under “Experimental Procedures.” Tyr-3 of the VPg peptide is used as the nucleophile for nucleotidyl transfer of UMP templated by the 2C-cre to yield VPg-pU, which undergoes another round of nucleotidyl transfer to yield VPg-pUpU. Uridylylated products are sepa-rated from labeled UTP by electrophoresis through a 15% Tris-Tricine gel and visualized by phosphorimaging. WT (A) and Glu-297 (B) 3Dpol uridylylation activity was monitored as a function of UTP concentration. Both VPg-pU and VPg-pUpU were produced by both enzymes.CandD, decreased affinity of Glu-297 3Dpol for UTP relative to WT 3Dpol. Total product formed was plotted as a function of UTP concentration and fit to a hyper-bola (Equation 2,solid line) for WT (C) and Glu-297 (D) 3Dpol. TheK0.5values for UTP were 3.9⫾1.2Mfor WT
3Dpol and 56⫾11Mfor Glu-297 3Dpol. The maximum values for UMP incorporated were 5.0⫾0.5MUMP incorporated for WT 3Dpol and 3⫾0.2MUMP incorporated for Glu-297 3Dpol.EandF, decreased processivity for Glu-297 3Dpol. Processivity is defined as the fraction of product that appears as VPg-pUpU relative to all products. Processivity is plotted as a function of UTP concentration and fit to a hyperbola (Equation 2,solid line) for WT (E) and Glu-297 (F) 3Dpol. TheK0.5values for UTP were 0.8⫾0.2Mfor WT 3Dpol and 5.7⫾1.6Mfor
poration of all four nucleotides were comparable, and no strong stops (Fig. 6A) or long lags (inset to Fig. 6B) were observed. We conclude that any change in processivity of RNA-primed RNA synthesis by Glu-297 3Dpol is below the limit of detection of this assay. Because the rate constant for dissociation of 3Dpol from S/S-U is ⬍0.0001 s⫺1 (12), an incorporation rate constant in the 0.0001– 0.001 s⫺1regime would be required to observe a difference in processivity by using this assay.
Efficient and Processive Uridylylation of VPg by Gly-297
3Dpol—Serial passage of Glu-297 PV uncovered a
pseudo-re-vertant virus, Gly-297 PV, that we isolated previously (27). This virus exhibits delayed growth kinetics but reaches titers within one log of wild type (27). Interestingly, growth of this virus could be stimulated at least 5-fold by growth in the presence of Mn2⫹(27). RNA-primed RNA synthesis by Gly-297 3Dpol was similar to WT 3Dpol; VPg-primed RNA synthesis was not eval-uated (27). If the reduced yield of uridylylated VPg products and/or processivity of VPg uridylylation by Glu-297 3Dpol was indeed the reason that Glu-297 PV did not replicate, then Gly-297 3Dpol should exhibit enhanced production of uridylylated VPg products, processivity, or both. Indeed, VPg uridylylation by Gly-297 3Dpol was essentially indistinguishable from WT 3Dpol (Fig. 7).
Attempts to Rescue Glu-297 3Dpol RNA-primed Elongation
Rate by Rational Design—The initial observation that the rate
constant for AMP incorporation into S/S-U by Glu-297 3Dpol
was 10-fold slower than WT 3Dpol was first interpreted to mean that the context into which Glu-297 was placed was sub-optimal. Structure/sequence comparisons identified two other changes in the ribose-binding pocket between RdRps from RNA viruses of eukaryotes and prokaryotes. These changes were at positions 288 and 290 using the PV 3Dpol numbering (Fig. 1). We changed Ser-288 to methionine and Cys-290 to asparagine individually and in combination in the context of Glu-297 3Dpol. None of these derivatives rescued the activity of Glu-297 3Dpol, because all changes yielded derivatives that retained⬍3% of the poly(rU) polymerase activity observed for WT 3Dpol (Table 3).
DISCUSSION
Previous studies of the RdRp from PV, 3Dpol, established a role for Asn-297 in ribonucleotide selection (14). By changing Asn-297 to Ala, a 3Dpol derivative was created that was inca-pable of distinguishing an rNTP with correct base pairing capacity from a 2⬘-dNTP with the same capacity (14). A struc-tural model for the 3Dpol elongation complex has been devel-oped based on homology modeling, because a crystal structure for this complex is still not available (14). Many of the features of this model are consistent with biochemical data, in particular a hydrogen bonding interaction between Asn-297 and the 2⬘-hydroxyl of the rNTP (Fig. 1A) (14). Interestingly, PV con-taining substitutions at position 297 of 3Dpol, including Ala, Gln, and Asp, were all inviable, despite the fact that all of these 3Dpol derivatives exhibited demonstrable levels of RNA-primed elongation activity (14). Consistent with the essential nature of this residue was the finding that all animal and plant RNA virus polymerases have an Asn at this position (Fig. 1B). Therefore, it was quite surprising to find that RNA phage poly-merases have a Glu at this position (Fig. 1B).
We hypothesized that the cytosolic availability of 2⬘-dNTPs in prokaryotes may account for this difference. Perhaps a Glu at this position would confer upon the polymerase the capacity to be more stringent in its selection against 2⬘-dNTP utilization and perhaps even permit the virus to grow. A Glu-297 3Dpol was indeed active, and a decrease in 2⬘-dNTP affinity was noted. However, a decrease in efficiency of 2⬘-dNMP incorporation relative to WT 3Dpol was not observed (Table 2). Glu-297 3Dpol exhibited two differences relative to WT 3Dpol in the RNA-primed synthesis of RNA. The observed rate constant for correct rNMP incorporation was decreased by 4- to 11-fold to the 10 –100 s⫺1regime (Table 2). The efficiency of GMP mis-FIGURE 6.Processive RNA-primed RNA synthesis by Glu-297 3Dpol is
nor-mal.Nucleotide-addition experiments were performed as described in the legend to Fig. 2 in the presence of all four nucleotides. The phosphorimaging of the gel (A) and quantification thereof (B) are shown. The kinetics of forma-tion and disappearance of 11-mer (E), 12-mer (䡺), 13-mer (F), and 14-mer (f) were simulated (solid lines) to net rate constants of 1 s⫺1, 4 s⫺1, 7 s⫺1, and 2 s⫺1
for formation of each of the individual products. Theinsetis an enlargement of the data between 0 and 2.5 s.
incorporation was increased by 14-fold, corresponding to a mutation frequency of 1⫻10⫺3.
Recent studies of the kinetics of nucleotide incorporation by the phage QRdRp have shown that the average rate constant for nucleotide incorporation by this enzyme is⬃10 s⫺1(43), with projected maximal rates being on the order of 30 s⫺1.3In addition, Drake (44) has shown that the mutation rate of Qin cells is 1.5⫻10⫺3, 10- to 100-fold higher than measured for PV in the same study. Our data would suggest that the presence of Glu instead of Asn in the ribose-binding pocket contributes to both the reduction in replication speed and fidelity.
Do RNA bacteriophage require a slower replication rate, greater population diversity, both, or neither? Given the sub-stantially faster doubling time of prokaryotic cells relative to mammalian cells, it appears unlikely that a slower replication rate for RNA bacteriophage would increase the yield of progeny virus per cell at a given time. Unlike mammalian host cells, prokaryotic host cells exhibit substantial phenotypic diversity, for example the presence of phage-resistant cells (45). It is pos-sible that the increased mutation frequency of the RNA phage permits phage evolution to keep pace with host cell evolution, thus precluding virus extinction caused by the selection of phage-resistant cells. Because viral population dynamics theory predicts a log-linear relationship between virus titer and muta-tion frequency, RNA phage will be able to absorb the increased mutational load and avoid extinction catastrophe by growing to high titer (46).
To our knowledge, Glu-297 3Dpol represents the first RdRp derivative with a mutator phenotype. As discussed below, studying the biological consequences of decreased fidelity is not possible in the PV system. However, construction of this deriv-ative in the context of other animal RNA viruses may be possi-ble. Similarly, construction of a Qmutant encoding an RdRp with an Asn in the ribose-binding pocket instead of Glu should alter phage evolutionary capacity relative to the host, providing important insight into the role of parasite/pathogen-evolving ability in host-parasite co-evolution.
Our previous biochemical and biological studies of position 297 of PV 3Dpol led us to conclude that subtle (2.5- to 5-fold) decreases in the elongation rate measuredin vitrocause sub-stantial RNA replication defectsin vivodue to tight coupling between translation and replication (14). Since these initial studies were published, there have been two advances in the field. First, anin vitroreaction that mimics VPg-primed initia-tion of PV genome replicainitia-tion has been developed (Scheme 2)
(31, 47). Second, it has become clear that not all 3Dpol alleles that confer a reduced elongation ratein vitroconfer a substan-tial defect to RNA replicationin vivo(48).
Evaluation of Glu-297 3Dpol in the VPg uridylylation reac-tion revealed that this derivative had a substantially reduced affinity for UTP (14-fold) relative to WT 3Dpol (Fig. 5, compare CtoD), without a significant difference in the observed rate of uridylylation, perhaps a reflection of the fact that nucleotidyl transfer is not rate-limiting in this reaction. In addition, there was a decrease in uridylylation processivity in that only approx-imately half of the Glu-297 3Dpol complexes yielded VPg-pUpU even under apparently saturating concentrations of UTP (Fig. 5F). In the Glu-297 3Dpol elongation complex, affinity for UTP was only affected minimally (Table 2), elongation rate was decreased substantially (10-fold) (Table 2), and processivity was not affected at all (Fig. 6).
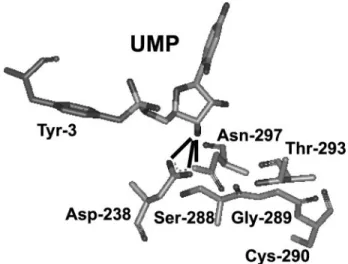
The opposite effects of the Glu-297 substitution on the two different reactions, VPg uridylylation and RNA elongation, sug-gest differences in the structural role and biochemical function of Asn-297 in these reactions. The structure of the foot-and-mouth disease virus 3Dpol-VPg-pU complex has been solved (49). Based upon this structure, Asn-297 of PV 3Dpol would interact with the 3⬘-OH in the uridylylation complex (Fig. 8) but with the 2⬘-OH in the elongation complex (Fig. 1A). The find-ing, that relatively modest reductions in yield of uridylylated VPg (10- to 15-fold) and processivity (2-fold) have such dra-matic effects on genome replication and virus production, sug-gests that drugs targeting initiation may be more efficacious than those targeting RNA elongation.
The inability of Glu-297 3Dpol to support both protein-primed RNA synthesis and RNA-protein-primed RNA synthesis may suggest that the pressure to be more selective against dNTP utilization also limits the nature of the initiation mecha-nisms employed by RNA phages. For example, both plant and animal RNA viruses employ both protein-primed (VPg
3K. Hosada, personal communication.
SCHEME 2.VPg uridylylation reaction.Incorporation of two UMP residues into VPg by 3Dpol is templated by a single AMP residue in the 2C-cre RNA using a slide-back mechanism (40). The reaction is capable of processive syn-thesis of VPg-pUpU but under sub-optimal conditions VPg-pU also accumu-lates (31, 41). The reaction is stimulated by 3CD, presumably as a recruitment/ retention factor for 3Dpol (31, 35, 41).
in Fig. 1B) andde novo(Capandpppin Fig. 1B) mechanisms of initiation; however, RNA phages employ only ade novo mechanism (Fig. 1B).
Glu-297 PV was quasi-infectious as a pseudo-revertant PV (Gly-297 PV) was isolated (Fig. 3B). This virus was also isolated in the past and shown to be an Mn2⫹-stimulated PV mutant (27). Gly-297 3Dpol restored the defects to uridylylation observed for Glu-297 3Dpol, suggesting that the uridylylation defect is the cause of the severe RNA synthesis defect in cells observed here and with other 297 alleles (14). Why do Gly-297 3Dpol and PV function (27) and Ala-297 3Dpol and PV not function (14)? We propose that a water molecule can be accom-modated in the pocket formed by Gly-297 but not in the pocket formed by Ala-297, thus permitting hydrogen bonding as observed for Asn-297. If our interpretation is correct, then the temperature sensitivity of RNA synthesis observed for Asp-297 PV (14) and the Mn2⫹dependence of RNA synthesis observed for Gly-239 and Ala-297 PV (27) are a result of an initiation (2C-cre-dependent VPg uridylylation) defect. To our knowl-edge, these PV mutants represent the first polymerase mutants with a specific defect to VPg uridylylation for any picornavirus or any RNA virus that employs a protein-primed mechanism of initiation. These mutants will be very useful for studying initi-ation and clarifying some of the controversies surrounding the role of the 2C-cre-templated uridylylation in picornavirus minus-strand and plus-strand syntheses (37, 50).
Acknowledgment—We thank Dr. Harsh B. Pathak for purification of the 3CD and 2C-cre RNA.
REFERENCES
1. Hayden, F. G. (2006)Curr. Opin. Infect Dis.19,169 –178
2. Weiss, S. R., and Navas-Martin, S. (2005)Microbiol. Mol. Biol. Rev.69,
635– 664
3. Kim, W. R. (2002)Hepatology36,Suppl. 1, S30 –S34
4. Behrens, S. E., Tomei, L., and De Francesco, R. (1996)EMBO J.15,12–22 5. Hayden, F. G. (1999)Adv. Exp. Med. Biol.458,55– 67
6. Kitamura, N., Semler, B. L., Rothberg, P. G., Larsen, G. R., Adler, C. J., Dorner, A. J., Emini, E. A., Hanecak, R., Lee, J. J., van der Werf, S.,
Ander-son, C. W., and Wimmer, E. (1981)Nature291,547–553
7. Semler, B. L., and Wimmer, E. (eds) (2002)Molecular Biology of Picorna-viruses, ASM Press, Washington, DC
8. Vignuzzi, M., Stone, J. K., and Andino, R. (2005)Virus Res.107,173–181 9. Harki, D. A., Graci, J. D., Korneeva, V. S., Ghosh, S. K., Hong, Z., Cameron,
C. E., and Peterson, B. R. (2002)Biochemistry41,9026 –9033
10. Harki, D. A., Graci, J. D., Galarraga, J. E., Chain, W. J., Cameron, C. E., and Peterson, B. R. (2006)J. Med. Chem.49,6166 – 6169
11. Jochmans, D., Deval, J., Kesteleyn, B., Van Marck, H., Bettens, E., De Baere, I., Dehertogh, P., Ivens, T., Van Ginderen, M., Van Schoubroeck, B., Ehteshami, M., Wigerinck, P., Gotte, M., and Hertogs, K. (2006)J. Virol.
80,12283–12292
12. Arnold, J. J., and Cameron, C. E. (2000)J. Biol. Chem.275,5329 –5336
13. Thompson, A. A., and Peersen, O. B. (2004)EMBO J.23,3462–3471
14. Gohara, D. W., Crotty, S., Arnold, J. J., Yoder, J. D., Andino, R., and Cam-eron, C. E. (2000)J. Biol. Chem.275,25523–25532
15. Gohara, D. W., Arnold, J. J., and Cameron, C. E. (2004)Biochemistry43,
5149 –5158
16. Crotty, S., Cameron, C. E., and Andino, R. (2001)Proc. Natl. Acad. Sci. U. S. A.98,6895– 6900
17. Castro, C., Arnold, J. J., and Cameron, C. E. (2005)Virus Res.107,141–149 18. Crotty, S., Maag, D., Arnold, J. J., Zhong, W., Lau, J. Y., Hong, Z., Andino,
R., and Cameron, C. E. (2000)Nat. Med.6,1375–1379 19. Traut, T. W. (1994)Mol. Cell Biochem.140,1–22 20. Lee, Y. D., and Elledge, S. J. (2006)Genes Dev.20,334 –344
21. Yao, R., Zhang, Z., An, X., Bucci, B., Perlstein, D. L., Stubbe, J., and Huang, M. (2003)Proc. Natl. Acad. Sci. U. S. A.100,6628 – 6633
22. An, X., Zhang, Z., Yang, K., and Huang, M. (2006)Genetics173,63–73 23. Gohara, D. W., Ha, C. S., Kumar, S., Ghosh, B., Arnold, J. J., Wisniewski,
T. J., and Cameron, C. E. (1999)Protein Expr. Purif17,128 –138 24. Pathak, H. B., Ghosh, S. K., Roberts, A. W., Sharma, S. D., Yoder, J. D.,
Arnold, J. J., Gohara, D. W., Barton, D. J., Paul, A. V., and Cameron, C. E. (2002)J. Biol. Chem.277,31551–31562
25. Herold, J., and Andino, R. (2000)J. Virol.74,6394 – 6400
26. Arnold, J. J., and Cameron, C. E. (2004)Biochemistry43,5126 –5137 27. Crotty, S., Gohara, D., Gilligan, D. K., Karelsky, S., Cameron, C. E., and
Andino, R. (2003)J. Virol.77,5378 –5388
28. Parsley, T. B., Cornell, C. T., and Semler, B. L. (1999)J. Biol. Chem.274,
12867–12876
29. Cornell, C. T., and Semler, B. L. (2002)Virology298,200 –213
30. Xiang, W., Harris, K. S., Alexander, L., and Wimmer, E. (1995)J. Virol.69,
3658 –3667
31. Paul, A. V., Rieder, E., Kim, D. W., van Boom, J. H., and Wimmer, E. (2000)
J. Virol.74,10359 –10370
32. Yang, Y., Rijnbrand, R., Watowich, S., and Lemon, S. M. (2004)J. Biol. Chem.279,12659 –12667
33. Franco, D., Pathak, H. B., Cameron, C. E., Rombaut, B., Wimmer, E., and Paul, A. V. (2005)Virol. J.2,86
34. Lyle, J. M., Clewell, A., Richmond, K., Richards, O. C., Hope, D. A., Schultz, S. C., and Kirkegaard, K. (2002)J. Biol. Chem.277,16324 –16331 35. Richards, O. C., Spagnolo, J. F., Lyle, J. M., Vleck, S. E., Kuchta, R. D., and
Kirkegaard, K. (2006)J. Virol.80,7405–7415
36. Ambros, V., and Baltimore, D. (1978)J. Biol. Chem.253,5263–5266 37. Murray, K. E., and Barton, D. J. (2003)J. Virol.77,4739 – 4750
38. Tellez, A. B., Crowder, S., Spagnolo, J. F., Thompson, A. A., Peersen, O. B., Brutlag, D. L., and Kirkegaard, K. (2006)J. Mol. Biol.357,665– 675 39. Crawford, N. M., and Baltimore, D. (1983)Proc. Natl. Acad. Sci. U. S. A.
80,7452–7455
40. Paul, A. V., Yin, J., Mugavero, J., Rieder, E., Liu, Y., and Wimmer, E. (2003)
J. Biol. Chem.278,43951– 43960
41. Rieder, E., Paul, A. V., Kim, D. W., van Boom, J. H., and Wimmer, E. (2000)
J. Virol.74,10371–10380
42. Lyons, T., Murray, K. E., Roberts, A. W., and Barton, D. J. (2001)J. Virol.
75,10696 –10708
43. Nakaishi, T., Iio, K., Yamamoto, K., Urabe, I., and Yomo, T. (2002)J. Biosci. Bioeng.93,322–327
44. Drake, J. W. (1993)Proc. Natl. Acad. Sci. U. S. A.90,4171– 4175 45. Jain, R., Knorr, A. L., Bernacki, J., and Srivastava, R. (2006)Biotechnol. Prog.
22,1650 –1658
46. Bull, J. J., Sanjuan, R., and Wilke, C. O. (2007)J. Virol.81,2930 –2939 47. Paul, A. V., van Boom, J. H., Filippov, D., and Wimmer, E. (1998)Nature
393,280 –284
48. Arnold, J. J., Vignuzzi, M., Stone, J. K., Andino, R., and Cameron, C. E. (2005)J. Biol. Chem.280,36417–36428
49. Ferrer-Orta, C., Arias, A., Agudo, R., Perez-Luque, R., Escarmis, C., Do-mingo, E., and Verdaguer, N. (2006)EMBO J.25,880 – 888
50. Sharma, N., O’Donnell, B. J., and Flanegan, J. B. (2005)J. Virol. 79,
3565–3577
Victoria S. Korneeva and Craig E. Cameron
DETERMINED BY A RESIDUE IN THE RIBOSE-BINDING POCKET
doi: 10.1074/jbc.M610090200 originally published online March 29, 2007 2007, 282:16135-16145.
J. Biol. Chem.
10.1074/jbc.M610090200
Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted
•
When this article is cited
•
to choose from all of JBC's e-mail alerts
Click here
http://www.jbc.org/content/282/22/16135.full.html#ref-list-1