Biochemistry
Transduction
of
a
cellular oncogene: The
genesis of Rous
sarcoma
virus
(recombination/splicing/genetic transposition)
RONALD
SWANSTROM*,
RICHARD C. PARKER, HAROLD E. VARMUS, ANDJ. MICHAEL BISHOPtDepartmentofMicrobiology and Immunology,UniversityofCalifornia, School of Medicine,San Francisco,California94143
Contributed byJ. Michael Bishop, January 28, 1983
ABSTRACT Theoncogene ofRous sarcoma virus (v-src) arose by transduction of a cellular gene (c-src). In an effort to explore themechanism of transduction, we have identified the splice ac-ceptor site used in the genesis of mRNA forv-arc,shown that an equivalent site is used in the splicing of mRNA forc-src, and de-termined the nucleotide sequence from the boundaries of ho-mology between v-arc and c-src. Our data indicate that (i) only a portion of c-src isrepresented within v-src, (ii) the leftward re-combination between the genome of thetransducing virus and c-srcoccurred in an intron of the cellular gene, (iii)v-arcisinpart aspliced version of the corresponding portion of c-src, and (iv) nucleotide sequences represented once in the genome of the trans-ducing virus become duplicated to flank v-src. These findings in-dicate thatthe first step in transduction is probablyrecombination between DNA forms of the transducing viral genome and c-src andotherwise support the prevailing model for transduction by retroviruses. The carboxyl termini of the proteins encoded by v-arcandc-src differappreciably. Anunidentified domain of 127 or128nucleotidesis located at differentpositions in the genomes oftwostrains of RSV and givesevidence of being a foreign ele-mentthatentered the viral genomes by genetic transposition in-dependent of thetransduction ofarc.
The oncogenesofretroviruses arose by transduction of cellular genes (1). The mechanism of transduction has yet to be elu-cidated but previous comparisonsof retroviraloncogenes and theircellular progenitorshaveengenderedseveralhypothetical models. Chiefamongtheseis aschemein which transduction ismediatedbytwoseparate steps ofrecombination-the first between the DNAforms ofaretroviralgenomeandacellular gene,the secondbetweenachimericRNAtranscribed fromthe product ofthefirst recombination and theRNA genomeof a retrovirus (seeDiscussion).
The cellularoriginsof oncogenes wereoriginally recognized fromstudies of theoncogene(v-src) ofRous sarcoma virus(RSV) anditscellular progenitor(c-src)(2).In aneffortto testthemodels fortransduction byretroviruses, wehave identifiedthesplice acceptor siteusedinthegenesisofmRNAforv-src, shownthat
anequivalentsite isused inthesplicing ofmRNAforc-src,and determined the nucleotide sequence from the boundaries of
homology
between v-src and c-src. Our resultsprovide
evi-dence that the firststep in transduction isindeed
recombina-tion between viral and cellular DNAs and otherwise support theprevailing model for transduction. In
addition,
wehave found thatproteins encodedbyv-srcandc-srcdifferappreciablyat theircarboxyltermini,andwehave identifiedadomainof127 or128nucleotides (denoted X) that islocated atdifferent po-sitionsinthe genomes of theSchmidt-Ruppin subgroupA (SR-A) andPraguesubgroupC (Pr-C)strains of RSV.MATERIALS ANDMETHODS
Wehave previously described thepreparationof molecularclones ofv-src
(3)
and c-src(4), theidentification ofsplice junctions(5)
by
theuseof nuclease SI (6, 7),theisolation of polyadenyl-ylatedcellularRNA(8), and thedetermination of the nucleo-tidesequence(9) bytheprocedureofMaxamandGilbert(10).
Acorrectedversion(11)ofourprevious sequenceforv-srcand
its environs in SR-A RSV(12) has been used forpresent pur-poses.ThesequenceofthePr-CRSVgenome wasprovidedin
advance of publicationbyD.Schwartz,R. Tizzard,and W. Gil-bert(13).
RESULTS
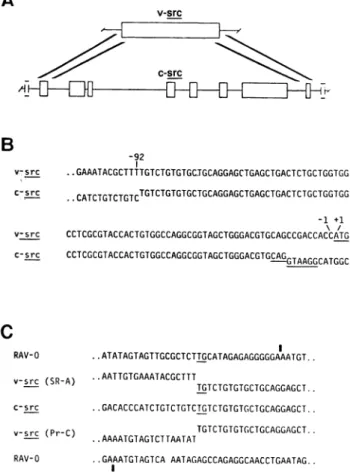
The Boundaries of Homology Betweenv-srcandc-src.
Pre-vious studies with heteroduplexes located the approximate
boundaries of homology betweenv-srcandc-src(4,14, 15). We usedmolecular hybridization toidentify and isolate DNA
en-compassingtheseboundaries within molecular clones ofc-src
(Fig. 1A) and then determined the nucleotide sequence of the isolatedDNAs.
Sequencesfromc-srcand thegenomeofSR-A RSVarealigned
acrosstheleftwardboundary of homologyin Fig. 1B. Thetwo sequencesdivergeat apoint92nucleotidesupstreamfromthe
initiationcodonofv-src (11, 12), as reported(13). The diver-gencedefines the point ofrecombination between transducing virusand c-srcand resides in an intron of c-src (seebelow).To
the right of the point of recombination, the nucleotide
se-quencesof c-srcand the genome ofSR-ARSV arehomologous
to apoint 10nucleotidesupstreamfromtheinitiationcodonof v-src, where the sequences again diverge (Fig. 1B). The di-vergence locates the rightward boundary of an exon in c-src (Fig. 1A), manifested bythe presence in c-src of a consensus
sequenceforasplice donorsite(Fig. 1B).
Inasearch forhomologybetween transducingvirusand c-src, we aligned sequences from c-src, SR-A RSV, Pr-C RSV,
andRAV-0-anendogenousretrovirusofchickens that isclosely related toreplicative elementsin RSVbutcarries nooncogene (Fig. 1C). Potential sites ofrecombinationwithc-src were
ap-parent from thealignments. ThesitesforSR-A and Pr-C were atdifferentbutnotwidely separated positionsinthe genome
of RAV-0. Thereis nohomology betweenc-srcand the poten-Abbreviations: RSV, Rous sarcoma virus;SR-A, theSchmidt-Ruppin subgroupA strainofRSV;Pr-C,thePraguesubgroupCstrainofRSV; X, adomain of127or 128nucleotidesfoundatdifferentpositionsinthe genomesofSR-AandPr-CRSV; U3,adomainatthe 3'terminusof the retroviralgenomewhichcomposes partof thelongterminalrepeat in the provirusofretroviruses.
*Presentaddress:DepartmentofBiochemistryandNutritionand Can-cerResearchCenter, Universityof NorthCarolina, Chapel Hill,NC 27514.
tTowhomreprint requestsshould beaddressed.
Thepublicationcostsofthis articleweredefrayedinpartbypagecharge
payment. This article must therefore be hereby marked
A
v-src
< ~~~~~c-src
B
-92
v-s rc . .GAAATACGCTTTTGTCTGTGTGCTGCAGGAGCTGAGCTGACTCTGCTGGTGG c-src CATCTGTCTGTCTGTCTGTGTGCTGCAGGAGCTGAGCTGACTCTGCTGGTGG
-1 +1
v-src CCTCGCGTACCACTGTGGCCAGGCGGTAGCTGGGACGTGCAGCCGACCACCATG..
c-s rc CCTCGCGTACCACTGTGGCCAGGCGGTAGCTGGGACGTGCAGGTAAGGCATGGC..
C
RAV- O
v-src (SR-A) c-src v-src (Pr-C) RAV-0
. ATATAGTAGTTGCGCTCTTGCATAGAGAGGGGGAAATGT..
..AATTGTGAAATACGCTTT
TGTCTGTGTGCTGCAGGAGCT..
.GACACCCATCTGTCTGTCTGTCTGTGTGCTGCAGGAGCT..
TGTCTGTGTGCTGCAGGAGCT..
..AAAATGTAGTCTTAATAT
..GAAATGTAGTCA AATAGAGCCAGAGGCAACCTGAATAG..
FIG. 1. The leftwardboundary ofhomology betweenc-srcandv-src.
(A) Location of theboundary. Thetopographiesofv-srcandc-src are
presentedschematically. The open boxes represent regions ofhomology between v-src andc-src.Thehomology withv-srcin c-src isinterrupted byintronsidentified previously(4,14,15) andperhapsothers. Theheavy diagonallines denote the regions ofc-srcthatcontainthe leftward and rightward boundaries of homology withv-src.Additionalexonsmay be presentinc-srcoutside of theregionsofhomologywithv-src,as de-notedby the incomplete boxes. (B)Identifying the leftwardboundary. The nucleotide sequences of SR-A RSV and c-src have beenalignedto
identify points of divergence (denoted by offsetting of the sequences). Thenumbering schemedesignatesthe first nucleotide5'of the initi-ationcodon forv-src(underlined)as -1.Theunderliningof thec-src
sequencelocatesa consensussequenceforasplicedonorsite(16).(C) Searchingforhomologybetweentransducingvirusandc-src.The
nu-cleotide sequences of the genomes of RAV-0(17), SR-A RSV(11,12), Pr-CRSV (13), and c-src have beenalignedacrossthe leftward bound-aryofhomologybetweenv-srcandc-src.Theboundaryisrepresented by the transition fromhomology with RAV-0tohomology withc-src. A
dinucleotide (T-G) shared by c-src,v-srcinSR-ARSV,andRAV-0atthe putativesiteof recombination forSR-A RSVisunderlined. The vertical barabove the RAV-0 sequence denotes the leftwardboundaryofU3,
adomainatthe3' terminusof the viralRNAgenomethat composes part of thelargeterminalrepeatintheprovirusesof retroviruses(18).
tialrecombinatory siteforPr-C RSVin thegenomeofRAV-0. By contrast, c-srcand RAV-O shareadinucleotide(T-G)atthe
putative siteof recombination forSR-ARSV(Fig. 1C). Whenweexamined therightwardextreme ofhomology
be-tweenv-src andc-src, ambiguities emerged(Fig. 2). The
nu-cleotidesequencesofv-srcandc-sr-cdivergefive residues up-streamfrom the previously definedterminationcodon forv-src
(11, 12). Apotentialopenreading frame ofc-src continuesfor another 12 codons before terminating in TGA. As a conse-quence, aminoacids atthecarboxyl terminus ofv-src are not
foundin c-src and c-src must therefore conclude with amino
acidsnotfoundinv-src.Thesefindingsposeseveralproblems.
First, we cannot accountfor theoriginof thecarboxylterminus
ofv-src ineither SR-AorPr-C RSV.Second,we cannotbe cer-tain thatwehavecorrectly described thecarboxylterminusof c-src. Forexample,ouridentificationof theterminationcodon forc-srcwouldbeincorrectifasplice donorsiteliestothe left of thatcodon, orif the splice acceptorsiteproposed forc-src
inFig. 2 is notused. Third, we cannotasyetidentify the right-ward point of recombination with c-src. The nucleotide se-quenceextending for =95 residues beyond thetermination
co-doninv-srccanbeaccounted for only becombining domains ofhomology with the genomes of RAV-0 and the PRC-II avian
sarcomavirus (unpublished data), andit is thereforeunlikely that anysingleretrovirusfor which the nucleotide sequenceis
currently available could be implicated in the rightward
re-combination with c-src. Ifour interpretation of the available sequencesiscorrect, however, the recombination occurredin
the vicinity of the present termination codon forv-src-very possibly, to its5' side.
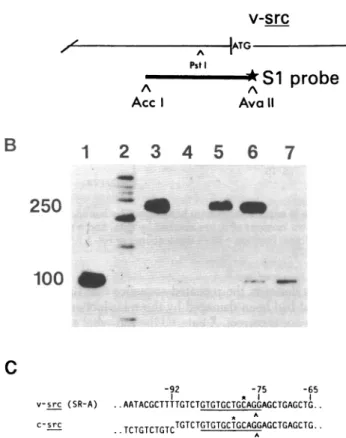
ASplice Acceptor Site Shared Betweenv-srcandc-src.The genesisofmRNAforv-src uses asplice donor sitelocated389 nucleotides from the 5' end ofthe viral genome (5, 9) and a
previously unidentified splice acceptor site upstream of the ini-tiationcodon for v-src(19-21). Wemapped the position of the splice acceptor site by using an end-labeled restriction frag-mentof DNA that spans thesplice acceptorsite in mRNAfor v-src(Fig. 3A). Hybridizationof thefragmenttogenomeRNA of SR-A RSVrendered theDNAresistant tohydrolysis by
nu-clease S1(Fig. 3B, lane 5). By contrast, hybridizationof the re-strictionfragmentto RNAfromcells infected with SR-ARSV produced both DNA that wasfully resistantto nucleaseS1and DNAthat wasreducedto alengthof-100nucleotides by hy-drolysiswith the nuclease(lane 6). WhenRNAfromuninfected
chicken cellswas used in thehybridizations, the smaller (and onlythesmaller) product of hydrolysiswas obtained (lane 7). These results were achieved by usingRNAfromuninfectedcells
inamountsatleast 100 times those used forRNAfrom infected
cells. We therefore attribute the small DNAfragment to hy-bridization with v-src mRNA in thecaseof infectedcells, whereas thefragment of necessity arises from c-srcmRNA inthecase
ofuninfected cells. We conclude that the size of the smaller fragmentdefines the approximate location of thesplice accep-torsiteupstream ofv-srcand thatanequivalentsite isusedin
theproduction ofmRNAfor c-src.
PheGluTyrLeuGlnAlaGlnLeuLeuProAlaCysValLeuGluValAlaGluAm
v-src (SR-A) ..TTCGAGTACCTGCAGGCCCAGCTGCTCCCTGCTTGTGTGTTGGAGGTCGCTGAGTAG.
C-src .GCGAGCCGTTTGCAGGCCCAGCTGCTCCCTGCTTGTGTGTTGGAGGTCGGAGTGATCCATGCACAGCCCGGTGCCCGCAGCAGCTCCCATTGA..
AlaGlnLeuLeuProAlaCysValLeuGluValGlyVa1I1eHisAlaGlnProGlyAlaArgSerSerSerHisOp
FIG. 2. Therightwardboundary ofhomology between v-src and c-src. Nucleotide sequences from the rightward-most region of homology that
wecouldidentify have been aligned. Divergence is indicated by the off-set sequences. Underlining denotes the previously identified termination codon forv-src(TAG),acandidateterminationcodon for c-src(TGA), andaconsensussequencefor a splice acceptor site (16) in c-src. The predicted
A
v-src
ATG
Pst
-L*
S1probe
A A
Acc Ava11
B
1
2
3
4
5
6
7
250
100
c
*
(SR-A) ..AATACGCTTTTGTCTGTGTGCTGCAGGAGCTGAGCTG..
A
c-sc ,TCTGTCTGTCTGTCTGTGTGCTGCAGGAGCTGAGCTG..
FIG. 3. Mappingaspliceacceptorsiteshared byv-srcandc-src.(A)
The strategy. Afragmentof DNAderived from theappropriatedomain
of theRSVgenomeandlabeledon oneendonly(star)wasusedtomap
thespliceacceptorsite.Details ofourprocedures have been described
(5)andfollowtheoriginal design of Weaver and Weissman(7).ATG, Theinitiationcodon forv-src.(B) Analysis of DNAs protected from
hy-drolysis by nuclease S1. Theend-labeledDNA(A)washybridizedwith
viral andcellularRNAs,hydrolyzed with nuclease S1,andanalyzedby
electrophoresis ina4%polyacrylamide/8 Mureagel(10). An
auto-radiograph of the gel after completion of electrophoresisisshown.Lanes: 1,marker DNA, -100nucleotides long, obtained by cleaving the end-labeledDNAfragmentshownin AwithPstI;2,markerobtained by
cleavingtheDNA ofplasmid pBR322with Hin Fand thenby end la-beling;3,DNAhybridized withyeastRNA (25,ag);4,DNAhybridized
with yeast RNAand hydrolyzed with nuclease Si;5,DNAhybridized
with virion RNA of RSV(unmeasured) and hydrolyzedwith nuclease
Si; 6,DNAhybridizedwith 5ggof total RNA from chicken cells in-fected withRSV andhydrolyzedwithnuclease Si;7,DNAhybridized
with 50 ,ugof polyadenylylatedRNAfrom uninfected chicken embryos
andhydrolyzedwith nucleaseS1.(C)Thespliceacceptorsite. The
se-quencesillustratedarelocatedupstreamof theinitiationcodons for v-srcandc-src.Nucleotides arenumberedasinpreviousfigures.The smallestlabeledDNAfragments generatedinthesplice-mapping
pro-cedure terminateatthepositionsmarkedwithasterisks.Arrows in-dicate the locationof thesplicesite,whichisinferred from the
under-linedconsensus sequence(16).
Welocated theacceptorsiteinv-srcandc-srcmRNAswith
greaterprecisionby fractionating the products of hydrolysis with nuclease S1 inagel designed for nucleotidesequence analysis
(datanotshown).Asmarkers, weusedfragments ofDNA
gen-erated by the procedure ofMaxam and Gilbert(10) from the
sameend-labeledDNAaswasusedfor thesplicemapping.The
resultsindicated that the splice acceptorsite islocated 75
nu-cleotidesupstreamof theinitiationcodon forv-src,inthe midst
ofasuitableconsensussequencesharedbyv-srcandc-src(Fig.
ANucleotide Sequence with Variable Locations inthe
Ge-nomes of RSV.Comparison of the genomes ofSR-Aand Pr-C RSVupstream ofthe transduced domain of c-src revealedan
absenceofhomology (Fig. IC). Further inspectionof
nucleo-tidesequences uncovered a sequence of127 or128nucleotides that is located to the left of v-srcin SR-A RSVbutto theright of v-src in Pr-C RSV. The enigmatic natureof this sequence prompted us to give it the arbitrary designation of X. Other
workershave also notedthe presence of X in SR-A and Pr-C
RSV, but their definition ofthe boundaries ofX is different from ours(22).
TheversionsofXand theirimmediate environs asfoundin
thegenomes of SR-A andPr-C RSV arealignedinFig. 4.
Sev-eral notable featuresareapparent.(i)Thetwo versionsof X
dis-playmore than 85%homology. (ii)Itappearsthat arepetition of12nucleotides may haveat one time flanked X both in SR-AandPr-CRSV.Thecompletesequence (C-G-C-T-C-T-T-G-C-A-T-A)isfoundattheleftwardboundaryofX in SR-A RSV
andattherightward boundaryofX inPr-C RSV. In SR-ARSV, therightwardcopyof the repeatlackssixnucleotides whereX
abuts the transducedportionofc-src. By contrast,the leftward copyofthe repeat inPr-C RSVlacks three nucleotides and is
interrupted byfivenucleotidesnotfoundintheotherversions
ofthe sequence. Asinglecopyof thesamesequence of12
nu-cleotides occurs in the genome of RAV-O, just upstream from the U3domain (Fig. 4). Since the genome of RAV-O doesnot contain X, we presume that the 12-nucleotide sequence may representthe siteatwhichXwasinserted
(see Discussion).
Wehave usedamolecularsubelonecontainingaportion of
X tosearchfor the sequenceinthe genomes of otheravian ret-roviruses. Bymolecularhybridizationwith cloned viralDNAs
immobilizedonnitrocellulose,wefound evidence forX inthe genomesofavianerythroblastosisvirusand myelocytomatosis virusbut not in the genomesof RAV-0, RAV-1, RAV-2, and PRC-II avian sarcoma virus (datanotshown).
The Viral Boundaries ofv-src. Havingrecognizedand de-finedX,wecouldnowdescribe the
topography
surrounding
v-src in somedetail. Itappearslikely that thesameportionof c-srchas been transducedin bothSR-AandPr-C RSV(Fig.
5).Inbothstrainsofvirus,the transducedunit isflankedbyalarge direct repeat
recognized previously
(12, 13, 23); thesamese-quenceispresent as asinglecopyinthe genome ofRAV-0. In
Pr-C,therepetitionsituatedtothe left ofv-srcincludesasmall portionoftheU3domain(AU3) andthe
rightward
versionof therepeat isinterrupted by
X. InSR-A,Xintervenesbetween theleftwardversionof therepeatandv-src,whereas the right-ward copyofthe repeat is undisturbed.DISCUSSION
TheTopographies ofv-srcandc-src. Wehave
compared
v-srcandc-src toexplore themechanismbywhich retrovirusestransduce cellular genes. Our
analysis
produced
several find-ingsthat bearontherelationship betweenthetwogenes. First, less than theentire c-srcgenehas beentransducedintoRSV.The domain between 10 and 75nucleotides upstream ofthe
initiationcodonin v-srcrepresents anuntranslatedexon in c-src. Tothe left ofthisexon,v-src contains aportionofanintron
fromc-srcthatisjoinedtothe
presumed
siteofrecombination with atransducing
virus. At leastone additional untranslatedexon mustlie upstream ofthis intron in c-srcbut itdoesnot
appearinv-src.
Second,
theaminoacidsequencesatthecar-boxyl
termini ofv-srcand c-srcmustdiffer fromeach other.- amoso
I
SR-A ..AGGATATAGTAGACACGCTTTTGC ATATGATACATAACTTCCCTGTTTTGCCCTTAGACTATTCAAGTTGCCTCTGTGGATTAGGGCTGGA Pr-C ..AGGACA A TTCTGCTTGGAATATGATGGCGT CTTCCCTGTTTTGCCCTTAGACTATTCGAGTTGCCTCTGTGGATTAGGGCTGGA
RAV-O ..AGGATATAGTAGTTGCGCTCTTGC ATA
GAGA..
-91
| TGTCTGTGTGCTGCAGGAGCTGAGCT..
SR-A GGCAGCTCGGATGGTCTGACGGCCAGATAAGGCAGGAAAGACAGCTATTGGTAATTGTGAAATACGCTTT
Pr-C GGCAGCACGGATAGrCTGATGGCCAAATAAGGCAGGCAAGACAGCTATTTGTAACTGCGAAATACGCTTTTGCATAGGGAGGGGGAAATGTAGTCT..
RAV-O CGCTCTTGCATAGAGAGGGGGAAATGTAGTCA..
..GTTG |
|(U3/LTR)
FIG. 4. X unitsofSR-A and Pr-C RSV. The sequences ofXhave been alignedsoas toachieve maximumhomology. The boundariesassigned
toXaredenotedbyboldverticalbars,positionsoccupiedbydifferent nucleotidesintwo versionsofXaremarked by dots, andasequencethat bracketsXtoformdirect repeats, andoccursbutonceinthe genomeof RAV-0,isunderlined. Position-91 inthe sequence of SR-ARSV represents thebeginningofv-src.Theleftwardboundaryof U3 is marked for RAV-1 and Pr-C RSV.
Third, the single splicea for v-src was derived: Moreover, a precise d4 mRNAforv-srchasshc of the initiation codonf in the same readingfra allfour of the upstrear nationcodons locatedsi ingsjoinagrowing list fromeukaryoticmRNA the 5' end of theRNA ANucleotide Sequer ofRSV. Wehave iden tides (designated X) th genomes of SR-A and quencesarepresentin roviruses. Theoriginsc
retroviral genomesare is the now
imperfect
r elementin atleast twoarosefrom a
single
cop
retroviral genomes dev
during
theinsertion of When didX enter t] catedatdifferentposit junction betweenXanuA
B
U5 gag pol
U5 gag pol
c
FIG. 5. Topographyof C.(C)RAV-0.Symbols:h tides thatisrepeatedto bri
0;shadedbox,theunident
coding regionofc-srctra
coding regionofunknown
theRSVgenome.
cceptor siteusedinthegenesisofmRNA
from a functional splice site in c-src. escription of the spliced leader in the )wnthatfourAUGcodonslie upstream Forv-src(5, 9). Twoofthesecodonsare
ime asv-src. Translation of v-src from
n AUG codons is precluded by
termi-hortdistances downstream. These find-)f exceptions tothe rule that translation
displaysadefect in therepeated sequence thatflanksX, asif
the repeathad been damaged bythe transduction ofc-src. If
this inferenceiscorrect,Xentered the retroviralgenome prior to src.The different locations ofX inSR-A and Pr-C RSVcould then beexplained in at least twoways:either one copyofthe
sequence has been deleted from each strainofvirus orthe
se-quencewaspresentinonlyoneof thetwo retroviruses partic-ipatingin theseparate stepsof transduction(seebelow andFig. 6).
sgenerally begins at the AUG closest to Transduction of CellularGenesbyRetroviruses. The
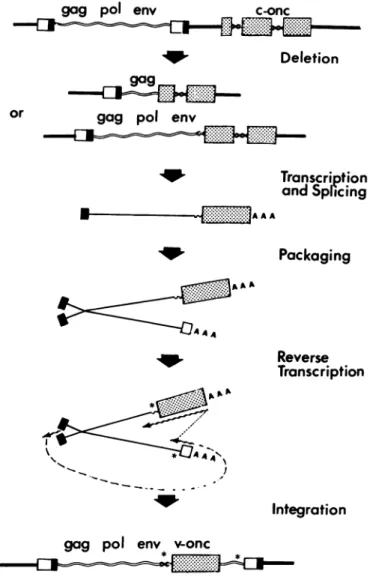
pre-(25). vailing model fortransductionof cellular genesbyretroviruses
icewithVariableLocationsin Genomes (18, 27-29) is outlined in Fig. 6. Fortuitous insertion of a retro-tified a sequence of 127 or 128 nucleo- virus provirus upstream from a cellular oncogene isfollowedby sat is found at different positions in the rearrangement of DNA that joins a leftward domain of the pro-Pr-C RSV. The same or related se- virus and a portion of the cellular oncogene into a hybrid tran-atleast twoother (butnotall)avian ret- scriptional unit. Transcriptionof the unit andsplicingproduce Af Xandthe meansbywhichXreached chimeric RNA that issuitable forpackagingintoheterozygous for themomentobscure. Ouronly clue retrovirus virions. Recombination duringasubsequent infec-'epeat of 12 nucleotides that flanks the tion generates a retrovirus genomethatnowincludes the
trans-strains ofRSV. Therepeats apparently duced portion of thecellular oncogene.
By
of thesame sequencethat resides in Several features of this model are evident from ourcom-7oid of X and may therefore have arisen parisonof v-src and c-src. (i) Theleftward recombination with atransposablegeneticelement (26). c-srcapparently occurred within anintron, an eventthatcan
he genome(s)ofRSV, andwhyis itlo- be easily explained only ifthe initialstep in transduction is a
ions inthe SR-AandPr-C strains?The rearrangementofDNAsuchas thedeletion illustratedinFig. itheleftboundary ofv-src in SR-A RSV 6. (ii) v-src represents in part asplicedversion of the homol-ogous regions of c-src. (iii)The second recombination is pos-tulatedto occur between the spliced chimeric RNAand viral
env U3 RNA during the course of reverse transcription (Fig. 6). The
src recombination should thereforeoccurwithinan exonofc-src,
andhomology betweenc-srcand the transducingviruswould facilitate the crossing-over. We were not able to locate the AU3 rightward point of recombination with c-src with any
assur-env
M-
U3 ance, but it appears to have taken place within an exon of c--<j src jtI.fr
src-either a coding domain or an untranslated region of the gene. Previous evidence indicates that recombination of the sortconjectured can occur duringthe course of retroviralin-fection (28). (iv) Thelargedirect repeat that brackets v-src pre-U5 gag pol env U3 sumably mirrors the occurrence of two separate recombinatory e events, thefirst downstream of the progenitor for therepeated sequenceintransducingvirus, thesecond upstreamof the
se-avianretrovirus genomes. (A) SR-A. (B) Pr- quence (see asterisks in Fig. 6).
Leavyarrows, the sequence of 120 nucleo- Ourpresentconclusions are inaccord with recent
findings
acketsrc inRSV butoccursonly onceinRAV- forOurbpre
conlusions ar
in acorwith
recent
fn gified sequence X; sinusoidal lines, the 5' non- oc-myb- thecellulargene that gaverise tothe oncogene (v-Lnsduced into RSV; brokenline, the 3' non- myb) of avianmyeloblastosis virus (30). (i) The leftward recoi-origin;AU3,aportion of theU3 domain in bination with c-myb occurred within an intron (ref. 30;
por-gag
c-onc
Deletion
gagpol
envTranscription and Splicing
A AA
Packaging
Reverse
Transcription
Integration
gag
pol
env voncFIG. 6. Model for the transduction of cellularoncogenesby
retro-viruses.
tionofc-mybareprecisely splicedoutofv-myb (30). (iii)The rightward recombinationwith c-myb occurredwithin anexon
ofc-myband without benefit ofextensivehomology between c-mybandthegenomeof thetransducing virus.
WeretheSRand Prstrains ofRSVformedbyindependent transductions ofc-src,or wereboth derived fromasingle trans-duction?Although thetwo strains ofviruswere isolated
sep-arately(31), itis possiblethattheyare merely divergent
rep-resentationsof thesametransduction. The datapresented above
providecircumstantialevidence both for and againstthis
pos-sibility. We see no way to reachadecisive conclusion unless
newandmoretellinginformation canbe obtained.
We thank J. Marinos for assistance and C. C. Huang and K.-H. Klempnauer for unpublisheddata. This workwassupportedbygrants
from the National Institutesof Health and the American Cancer So-ciety. R.S.wassupportedbyGrant 1T32 CA 09043 from theNational
Institutes of Health. R.C.P. was supportedbythe American Cancer
Society (postdoctoralfellow) and theAmerican CancerSociety, Cali-forniaDivision(senior postdoctoralfellow).
1. Bishop, J. M. &Varmus, H. (1982)inMolecularBiology of
Tu-morViruses:RNA TumorViruses,eds. Weiss, R., Teich, N., Var-mus, H. E. &Coffin, J. (Cold SpringHarborLaboratory, Cold
Spring Harbor, NY),2ndEd.,pp. 999-1108.
2. Stehelin, D., Varmus,H.E., Bishop, J. M. &Vogt,P. K.(1976)
Nature(London)268,170-173.
3. DeLorbe,W.J., Luciw,P.A.,Goodman,H.M., Varmus,H. E.
&Bishop, J.M. (1980)1.Virol.36,50-61.
4. Parker, R.C., Varmus, H.E. & Bishop,J. M. (1981) Proc. Natl. Acad. Sci. USA78,5842-5846.
5. Hackett, P. B., Swanstrom,R., Varmus, H. E. &Bishop, J. M. (1982)J.Virol.41,527-534.
6. Berk,A. J. &Sharp, P. A.(1978)Proc. Natl. Acad. Sci. USA78, 1274-1278.
7. Weaver,R. F. &Weissmann,C. (1979) Nucleic Acids Res. 7, 1175-1193.
8. Gonda, T.J., Sheiness,D. K. & Bishop, J. M. (1982) Mol. Cell. Biol.2,617-624.
9. Swanstrom, R., Varmus, H. E. &Bishop, J. M. (1982)J. Virol.
41,535-541.
10. Maxam,A. M. &Gilbert,W. (1980) MethodsEnzymol. 65, 499-560.
11. Czernilofsky, A. P., Levinson,A. D., Varmus, H.E., Bishop,J. M., Tischer, E. &Goodman,H. M. (1983)Nature(London)301, 736-738.
12. Czernilofsky,A. P., Levinson,A. D.,Varmus, H. E.,Bishop,J.
M., Tischer,E. &Goodman, H.M.(1980) Nature (London)287, 198-203.
13. Schwartz, D., Tizzard, R. &Gilbert, W. (1982) in Molecular
Bi-ology of Tumor Viruses: RNA TumorViruses, eds. Weiss, R., Teich, N., Varmus,H. E. &Coffin,J. (ColdSpringHarbor Lab-oratory, ColdSpring Harbor, NY), 2ndEd.,pp. 1338-1348. 14. Shalloway, D.,Zelenetz, A. D. &Cooper, G. M. (1981) Cell24,
531-541.
15. Takeya,T., Hanafusa, H.,Junghans, R. P., Ju, G. &Skalka,A. M.(1981)Mol. Cell. Biol. 1, 1024-1037.
16. Mount, S. M.(1982)NucleicAcidsRes.10, 459-472. 17. Hughes, S. H.(1982)J.Virol.43, 191-200.
18. Varmus, H. E.(1982)Science 216,97-108.
19. Weiss,S.R., Varmus,H. E. &Bishop, J. M. (1977) Cell12, 983-992.
20. Mellon,P. &Duesberg, P. H.(1977)Nature(London) 270, 631-634.
21. Krzyzek, R.A., Collett, M.S., Lau,A. F.,Perdue,M.L.,Leis,
J. P. &Faras,A. J. (1978)Proc.Natl. Acad. Sci. USA75, 1284-1288.
22. Tsichlis, P. N., Donehower, L., Hayer, G., Zeller, N.,
Mala-vanca, R., Astrin, S. &Skalka,A.(1982)Mol. Cell. Biol. 2, 1331-1338.
23. Yamamoto,T.,Tyagi, J. S., Fagan,J.,Jay, G., deCrombrugghe,
B. &Pastan, I. (1980)J.Virol.35, 436-443.
24. Smart, J. E.,Oppermann, H., Czernilofsky, A. P., Purchio, A.
F.,Erickson, R. L. &Bishop, J. M. (1981)Proc.Natl. Acad.Sci. USA 78,6013-6017.
25. Kozak, M.(1978)Cell15, 1109-1123. 26. Calos, M. &Miller,J. (1980)Cell 20,579-595.
27. Goff, S. P.,Gilboa, E.,Witte,0. N. &Baltimore,D.(1980)Cell
22,777-785.
28. Goldfarb,M. P. &Weinberg,R. A.(1981)J. Virol.38,136-150. 29. Bishop, J. M.(1982) Adv. CancerRes.37, 1-32.
30. Klempnauer, K.-H.,Gonda,T.J. &Bishop,J.M.(1982)Cell31,
453-463.
31. Morgan,H. R. &Traub, W.(1964) Natl.Cancer Inst.Monogr.17,
392-393.