EXTRACELLULAR MATRIX-DERIVED HYDROGELS FOR DENTAL STEM CELL DELIVERY 1
No benefit of any kind will be received either directly or indirectly by the author(s). 2
Aiswarya Viswanath a, c, 1, Julie Vanacker a, 1, Loïc Germain a, Julian G Leprince a, Anibal. Diogenes b, Kevin 3
Shakesheff c, Lisa J White c, 1, Anne des Rieux a, 1* 4
5
a Louvain Drug Research Institute, Advanced Drug Delivery and Biomaterials Unit, Université Catholique de 6
Louvain, Brussels, Belgium. 7
b Department of Endodontics, University of Texas Health Science Center at San Antonio, San Antonio-TX, 8
USA 9
c School of Pharmacy, University of Nottingham, Nottingham NG7 2RD, UK 10
11
1 These authors contributed equally. 12
* Corresponding author: Anne des Rieux, [email protected] 13
Abstract 16
Decellularised mammalian extracellular matrices (ECM) have been widely accepted as an ideal substrate for 17
repair and remodelling of numerous tissues in clinical and pre-clinical studies. Recent studies have 18
demonstrated the ability of ECM scaffolds derived from site-specific homologous tissues to direct cell 19
differentiation. The present study investigated the suitability of hydrogels derived from different source tissues: 20
bone, spinal cord and dentine, as suitable carriers to deliver human apical papilla derived mesenchymal stem 21
cells (SCAP) for spinal cord regeneration. Bone, spinal cord and dentine ECM hydrogels exhibited distinct 22
structural, mechanical and biological characteristics. All three hydrogels supported SCAP viability and 23
proliferation. However, only spinal cord and bone derived hydrogels promoted the expression of neural lineage 24
markers. The specific environment of ECM scaffolds significantly affected the differentiation of SCAP to a 25
neural lineage, with stronger responses observed with spinal cord ECM hydrogels, suggesting that site-specific 26
tissues are more likely to facilitate optimal stem cell behaviour for constructive spinal cord regeneration. 27
Key words 28
ECM hydrogels, Dental stem cell delivery, Spinal cord, Bone, Dentine 29
1 Introduction 31
Traumatic spinal cord injuries (SCI) cause devastating neurological deficits and disabilities. Initial trauma 32
leads to immediate disruption of neural tissue by axon shearing, blood vessel rupture and cell death. 33
Subsequently a cascade of secondary events occurs composed of ischemic injury, inflammation, cell death, 34
demyelination of axonal tracts and the creation of a glial scar, forming a physical and chemical barrier 1. 35
Treatment options for SCI have been limited in part due to this complex pathophysiology 2. Cell transplantation 36
to replace damaged cells, provide trophic support and promote functional recovery is a favourable strategy for 37
SCI 3. 38
Stem cells derived from dental tissues are an attractive source for cell transplantation in the central nervous 39
system (CNS) due to accessible supply, high proliferation rate and the potential for autologous transplantation 40
4. Human dental stem cells are of neural crest origin, display neural stem cell properties 5, and have recently 41
been shown to induce functional recovery in SCI repair 6. Whilst several different populations of dental stem 42
cells have been identified, stem cells of the apical papilla (SCAP) have been shown to possess greater 43
proliferation potential than dental pulp stem cells 7 and express a variety of neural markers including βIII 44
tubulin, NeuN, nestin, neurofilament M and glial fibrillary acidic protein (GFAP) 5,8. Recently, whole human 45
apical papillae implanted into rat hemisected spinal cords induced a significant improvement of rat motor 46
function 4. However, SCAP cells transplanted in a fibrin hydrogel in the same study did not have the same 47
effect upon motor function 4, highlighting the importance of delivering SCAPs in their original niche or a 48
similarly supportive medium. The goal of the present work was thus to identify a suitable hydrogel to support 49
SCAP delivery for SCI. 50
Biologic scaffolds composed of extracellular matrix (ECM) produced by decellularisation of mammalian 51
tissues largely retain the functional complexity of their tissues of origin 9. Hydrogel forms of ECM do not 52
retain the three dimensional ultrastructure of the native tissue but still maintain in vitro and in vivo biologic 53
activity, including differentiation stimuli for neural stem cells 9,10 and the promotion of an M2 macrophage 54
phenotype 11,12. In addition, ECM hydrogels may supply signalling molecules and growth factors 13,14 required 55
homologous to the repair site, have previously shown the ability to enhance site appropriate cell phenotypes 57
compared to ECM scaffolds derived from non-homologous tissues for a range of complex tissues 16-18. 58
The objectives of the present study were to evaluate the suitability of different ECM derived hydrogels to 59
support SCAPs for delivery into SCI. Previously developed protocols were utilised to produce bone and spinal 60
cord ECM hydrogels (bECMh and scECMh, respectively) and a new method was developed to create a 61
hydrogel form of human dentine ECM (dECMh). The biomolecular composition, mechanical properties, and 62
in vitro cytocompatibility of the hydrogels were evaluated. We hypothesised that the different ECM hydrogels 63
would influence SCAP viability, proliferation and gene expression. 64
2 Material and methods 65
2.1 Chemicals and reagents 66
The Quant-iT™ Picogreen® assay kit and the Presto Blue kit were from Life Technologies (Carlsbad, US). 67
Bovine serum, L-glutamine, penicillin and streptomycin were from Gemini Bio-Products (Sera Laboratories 68
International, Bolney, UK). All other reagents were purchased from Sigma-Aldrich (Poole, UK). 69
2.2 ECM biologic scaffold production 70
Porcine spinal cord and bovine bone were obtained from market weight animals. Human wisdom teeth were 71
collected from healthy individuals aged between 16 and 18 years. An informed consent was obtained from all 72
donors (Approved by the Commission d’Ethique Biomédicale Hospitalo-Facultaire of UCL, 73
2012/14JUN/283). Female and male donors and animals have been used indiscriminately. Decellularisation of 74
bone (bECM) 19 and spinal cord (scECM) 20 were performed as described previously. Extraction of ECM from 75
human dentine (dECM) was performed using the protocol developed for bECM preparation with modifications. 76
Briefly, teeth were frozen in liquid nitrogen and broken with a Carver press (Carver, #3850, Wabash, US) and 77
the pulp was removed. The enamel was destroyed by sonication in 10% HCl for 20 min 21. Acid neutralisation 78
was performed by immersing the tissue in 2 volumes of water containing penicillin-streptomycin (0.5% w/v) 79
for 1 hr. Demineralisation was performed by incubating tooth fragments in a 0.5M HCl solution under agitation 80
and incubated in a 0.05% trypsin-0.025% EDTA solution for 24 hrs at 37°C at 300rpm for enzymatic 82
decellularisation. Finally, the material was washed with 2 volumes of PBS and freeze dried. 83
2.3 ECM digestion and solubilisation 84
Pepsin digestion and solubilisation was used to obtained pre-gel solutions 22,23. Lyophilised ECM powders 85
were added to 1mg/ml of pepsin in 0.01 N HCl for a final concentration of 10 mg ECM/ml. The suspension 86
was stirred at room temperature for 48 hrs for spinal cord ECM, 72 hrs for dentine ECM and 84 hrs for bone 87
ECM. The resultant pepsin digests were aliquoted and stored at -200C. Gelation of pepsin digests was induced 88
by neutralisation of salt concentration and pH at 4°C followed by warming to 37°C. Briefly, pepsin digests 89
were mixed with one tenth volume of 0.01N NaOH, one ninth volume of 10X PBS and made up to desired 90
volume with 1X PBS. 8 mg/ml of pre-gel solutions were prepared and incubated for 1 hr at 37°C to obtain 91
spinal cord, bone and dentine hydrogels, denoted scECMh, bECMh and dECMh. 92
2.4 DNA, collagen and sGAG quantification 93
Quantitative assessment of DNA in pepsin digests was conducted by an adaptation of previously published 94
method 23. DNA was extracted from pepsin digests by addition of an equal volume of 25:24:1 (v:v:v) 95
phenol/chloroform/isoamyl to 1ml of sample (10 mg/ml). Phenol was removed using an equal volume of 24:1 96
(v:v) chloroform/isoamyl alcohol. DNA was precipitated from the aqueous phase by the addition of 2 volumes 97
of ice-cold ethanol and 0.1 volume of 3M sodium acetate (pH 5.2) and was frozen at -800C for 30 min. The 98
frozen DNA was then centrifuged at 13,000 rpm for 10 min and washed with ethanol, dried at room temperature 99
and resuspended in 0.1 ml of TE buffer. DNA concentration was measured using a Quant-iT™ Picogreen® 100
assay kit following supplier instructions. A standard curve was prepared with known DNA concentrations from 101
0 – 100 ng/ml and samples were read using a Tecan Infinite M200 plate reader (Tecan UK, Reading, UK). 102
Collagen content of pepsin digests was measured by quantifying hydroxyproline 24. Briefly, pepsin digests and 103
a blank pepsin solution were hydrolysed with 1 volume of concentrated HCl (12N) overnight at 120°C. The 104
samples were dried by incubation at 90°C for 2 hrs followed by the addition of 4 volumes of 0.25 M sodium 105
for the assay. The samples were oxidised by incubation with 1 volume of 3-Chloramine T solution (0.07M) at 107
room temperature for 20 min followed by the addition of 4- p-dimethylaminobenzaldehyde (p-DAB) solution 108
[1.16M p-DAB in 60% perchloric acid (6.5ml) and Isopropanol (15ml)] and incubation at 600C for 30 min. A 109
standard curve was prepared using known concentrations of hydroxyproline ranging from 0 – 100 µg/ml and 110
the absorbance was measured at 540nm with a Tecan Infinite M200 plate reader. Collagen content was 111
determined using the relationship that hydroxyproline forms 14.3% of total amount of collagen 24. 112
The sulphated glycosaminoglycan (sGAG) content in pepsin digests was determined using the 1, 9–Dimethyl 113
methylene blue assay (DMMB) 25. One ml of digest was incubated with 0.1mg/ml of Proteinase K (>600U/ml) 114
overnight at 37°C. A solution of blank pepsin was used as negative control as well as diluent for the assay. 50µl 115
of sample was mixed with 200 µl of DMMB solution (0.03 M sodium formate, 0.046M DMMB, ethanol (0.5% 116
v/v) and formic acid (0.2% v/v) in a 96 well plate. A standard curve was constructed using known 117
concentrations of Chondroitin-4-sulphate from 0 – 75 µg/ml. The absorbance was measured immediately at 118
525nm. 119
2.5 ECM-based hydrogel gelation kinetics 120
Turbidimetric gelation kinetics of ECM-based hydrogels was determined as previously described 26. 121
Normalised Absorbance (NA) was calculated as shown in Eq. (1). 122
NA= 𝐴−𝐴0
𝐴𝑚𝑎𝑥−𝐴𝑜 (1)
123
Were A is absorbance at given time, A0 is the initial absorbance and Amax is the maximum absorbance attained 124
by the sample after inducing gelation. Normalised gelation was used to calculate the time at which 50% of 125
gelation (t1/2) and 95% of gelation (t 95) occurred. The lag time, tlag, was defined as the intercept of the linear 126
region of the gelation with 0% absorbance 19. Speed of gelation (S) was determined by calculating the slope of 127
the curve (n=8). 128
2.6 Rheological characteristics 129
Instruments, Worcestershire, UK). Cold pre-gel solutions were placed between two pre-cooled (4°C) 25 mm 132
parallel plates separated by a gap of 0.3 mm. For each run, the plates were warmed to reach 37°C (2°C/sec) and 133
the sample was exposed to 1% oscillatory strain with a constant angular frequency of 2 rad/sec. Data were 134
recorded every 30 sec (n=3). 135
2.7 Scanning electron microscopy (SEM) 136
Gel specimens (300 µl per well) were fixed in 5 % glutaraldehyde at room temperature. Samples were 137
dehydrated in a graded series of ethanol (25-100%), dried using a critical point dryer (CPD, Balzers Union, 138
Balzers, FL) and sputter coated with gold using a Cressington 208HR metalizer (ELOISE s.a.r.l., Tremblay, 139
FR). Images were acquired using a JSM-7600F (JEOL, Zaventem, BE) scanning electron microscope at a 140
voltage of 15 kV. 141
2.8 SCAP culture 142
Previously characterised human stem cells of the apical papilla (SCAP) (RP89 cells) 27 were used between 143
passages 6 and 9. SCAP were grown at 37°C in 5% CO2 in Minimum Essential Medium (Sigma-Aldrich) 144
supplemented by 10% bovine serum (Gemini Bio-Products), 1% of L-glutamine (Gemini) and 1% of penicillin 145
and streptomycin (Gemini) until they reached ~80% confluence. SCAP were then incorporated in the pre-gel 146
solutions, placed in 96 well plates and incubated at 37°C for 1 hr. 147
2.9 SCAP viability 148
Pre- gel solution (10µl) containing 500,000 cells/ml were loaded into µ-Slide Angiogenesis ibiTreat 149
microscopy chamber (Ibidi, Proxylab, Beloeil, BE) and were allowed to gel (n=3, N=3). After 2 days, a 150
Live/Dead assay was performed. Pictures (2 fields/sample, Z-stack: 1 picture every 3µm, for a total of 210 151
pictures/stack) were then acquired with a confocal microscope (LSM700, Zeiss, Zaventem, BE). All the 152
pictures were merged to count dead and living cells. Cell viability was expressed as a percentage of the number 153
2.10 SCAP proliferation 155
The effect of ECM origin on SCAP proliferation was analysed when cells were encapsulated in the hydrogels 156
(3D culture) or grown on the surface of hydrogel (2D culture). 157
2.10.1 SCAP proliferation in 3D culture 158
Pre-gel solutions containing 500,000 SCAP/ml were placed in 96 well plates (N=3, n=6) and incubated for 1hr 159
at 37°C to initiate gel formation. Pre-gel solution (100µl) without cells was used as a control. Independent 160
samples were prepared for each time point. After 2, 7 and 14 days, cell proliferation was analysed using a 161
Presto Blue kit. The fluorescence intensities (590 nm) were measured 2 hours after the addition of the Presto 162
Blue reagent. Values obtained for hydrogels without cells were used as background and subtracted from values 163
obtained for hydrogels with cells. A standard curve (from 13,000 to 250,000 cells) was used to correlate the 164
fluorescence intensity with cell number. Cell number fold increase was calculated by dividing the number of 165
cells at day 2, 7 or 14 by the initial number of seeded cells. Samples were collected and frozen at -80°C for 166
further processing for PCR analysis. 167
2.10.2 SCAP proliferation in 2D culture 168
Pre-gel solutions (20µl) were placed in 96 well plates and allowed to gel for 1 hr at 37°C. Two thousands cells 169
were seeded onto the surface of the hydrogels (N=3, n=6). Cell proliferation was evaluated as described for 170
2.10.1. 171
2.11 SCAP gene expression 172
Total mRNA was extracted using the phenol/chloroform extraction method. One microgram of mRNA was 173
reverse transcribed using the Reverse Transcription System kit (Promega, Madison, USA) (n=3). The resulting 174
cDNA was used as template for 30 cycles of semi-quantitative polymerase chain reaction in a T100 TM thermo 175
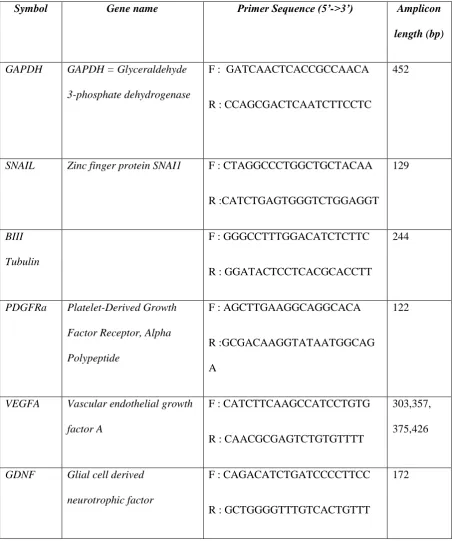
cycler (Bio-Rad, BE). Primer sequences are summarized in Table 1. The PCR products were pooled and 176
subjected to electrophoresis on Syber™Safe (Invitrogen, BE) stained 2% agarose gel. mRNA relative 177
2.12 Statistical analysis 179
Statistical analyses were performed using PRISM (GraphPad software, CA, USA) and JMP10 (SAS, NC, 180
USA). Two-way ANOVA with post hoc Bonferroni’s multiple comparison tests as well as Tukey’s HSD were 181
performed with a P-value of 0.05. Errors bars represent the standard deviation (SD) in all figures. 182
3 Results 183
3.1 Influence of ECM origin on cellular, collagen and sGAG content 184
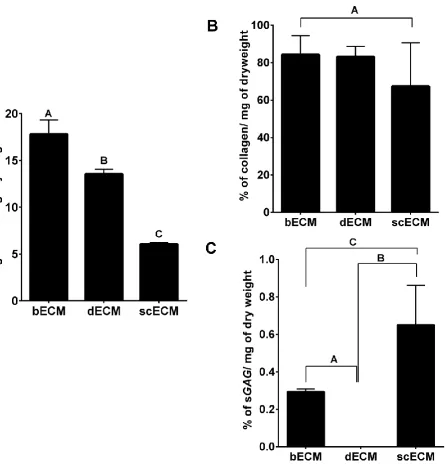
Determination of decellularisation efficiency was based upon quantification of dsDNA. Regardless of origin, 185
all three ECMs contained less than 50 ng/mg of dsDNA (Figure 1A). Concentrations of nucleic acid of bECM, 186
dECM and scECM were significantly different with the lowest quantity detected in scECM; bECM, dECM 187
and scECM contained 17.8±1.5 ng of DNA/mg, 13.56±0.5 ng of DNA/mg and 6.8±0.17 ng of DNA/mg, 188
respectively. No significant difference was observed between the collagen content of the three ECMs, although 189
scECM tended to contain less collagen (Figure 1B); bECM, dECM and scECM contained 84.28±10.20 %, 190
83.16±5.55 % and 67.41±23.16 % of collagen, respectively. Significantly more sGAG was detected in scECM 191
(0.65 % of dry weight) compared to bECM and dECM (0.3 % and 0%, respectively) (Figure 1C). 192
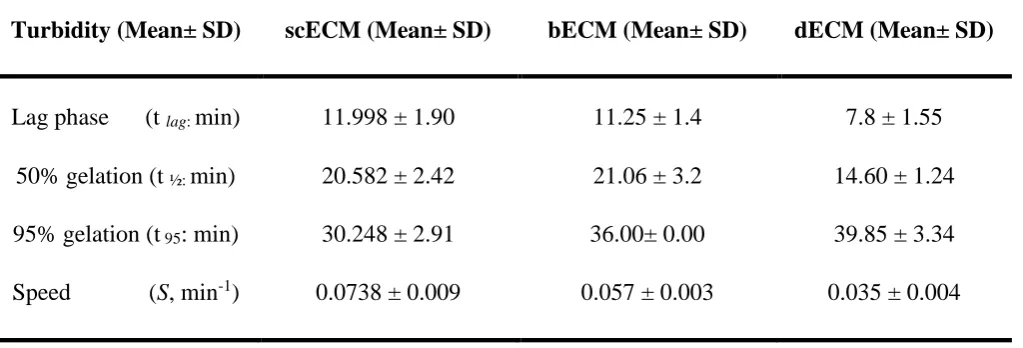
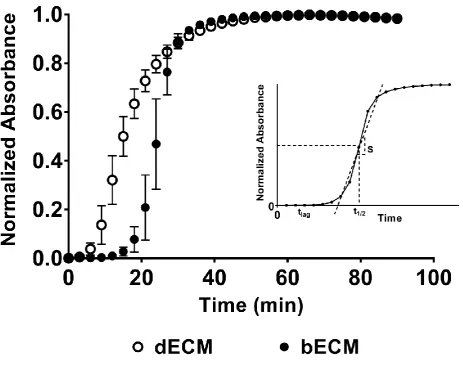
3.2 Influence of ECM origin on hydrogel gelation kinetics 193
The gelation kinetics of scECMh, bECMh and dECMh were sigmoidal, and gelation occurred within 40 min 194
(Figure 2). The tlag and t1/2 was 1.40-1.53 fold lower for dECMh compared to sc ECMh and bECMh. However, 195
the time required to reach 95% of the final turbidity is 0.75-0.84 fold lower for scECMh with respect to bECMh 196
and dECMh. The gelation speed was 1.3 and 2.1 fold faster for scECMh compared to bECMh, dECMh 197
respectively (Table 2). 198
3.3 Influence of ECM origin on hydrogel mechanical properties 199
Storage modulus (Gʹ) and loss modulus (Ǵ́́́́́́́́ʹ) of all the pre-gel solutions increased 5 min after pH neutralisation 200
and temperature elevation and reached a maximum after 1 hr (Figure 3). G’ was consistently higher than G”, 201
reflecting the formation of a solid hydrogel. Regardless of origin, hydrogels showed similar gelation profiles, 202
highest storage modulus (287.17 +/- 8.43 Pa); while scECMh and dECMh were significantly lower (19.35 +/- 204
1.16 and 4.79 +/- 0.33 Pa, respectively). 205
3.4 Influence of ECM origin on hydrogel microstructure 206
The origin of hydrogel markedly influenced the hydrogel morphology. While bECMh and dECMh exhibited 207
a fibrillary structure with disorganised thin fibres (Figure 4A and B); scECMh showed a honeycomb structure 208
with thick walls and large pores (Figure 4C). dECMh appeared to be composed of thicker fibres and smaller 209
pores in comparison to bECMh. 210
3.5 Influence of ECM origin on dental stem cell proliferation 211
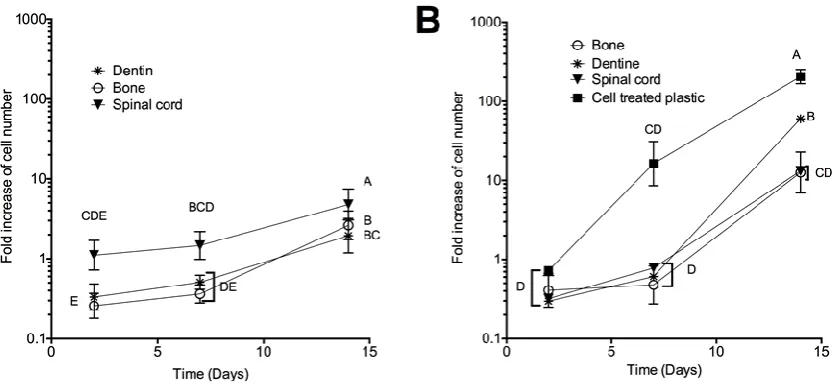
Regardless of the ECM origin, SCAP proliferated when incorporated in the ECM hydrogels. When cultured in 212
3D, the number of SCAP increased between 0.36 and 1.47-fold 7 days post seeding and 2 to 4.78-fold 14 days 213
post seeding compared to the initial seeding density (Figure 5A). No significant difference was observed 214
between day 2 and 7 for any of the hydrogels, whilst SCAP proliferation significantly increased between day 215
7 and day 14 for all of the hydrogels. The SCAP proliferation was 1.8 to 2.5-fold higher in scECMh compared 216
to the dECMh and bECMh on day 14, respectively. 217
When grown on the hydrogels (2D culture), the SCAP proliferation on various ECM hydrogels increased 218
between 0.48 to 1-fold 7 days of post seeding and between 12.6 to 60-folds 14 days post seeding (Figure 5B). 219
SCAP proliferation on tissue culture treated plastic (TCP) was much higher than either on 2D or in 3D 220
hydrogels with a fold increases of 16.28 and 205.7 at day 7 and 14, respectively. No significant difference was 221
observed between Day 2 and 7. 222
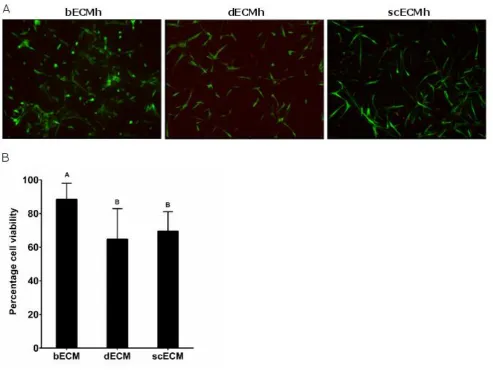
3.6 Influence of SCAP incorporation into different ECM on cell viability 223
SCAP viability 2 days after incorporation in all ECM hydrogels was above 65 % (Figure 6B). Viability was 224
significantly higher in bECMh than in dECMh and scECMh (85 %, 65 % and 68 %, respectively). In general, 225
SCAP morphology was elongated in all three ECM hydrogels, however some rounded cells were also observed 226
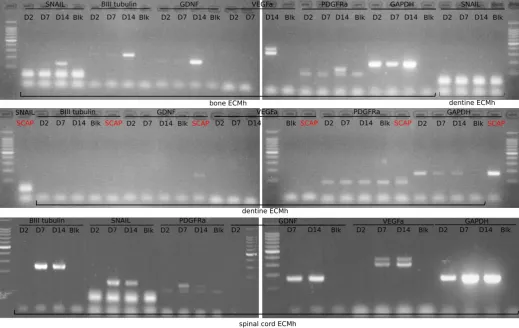
3.7 Influence of ECM origin on SCAP gene expression 228
The expression of genes related to the neural lineage (SNAIL, βIIITubulin, GDNF, VEGFA, PDGFRa, and 229
GAPDH as a housekeeping gene) were analysed to determine whether ECM origin would elicit or support 230
SCAP orientation. When SCAP were cultured routinely (Figure 7), GDNF and PDGFRa were slightly 231
expressed but no expression of SNAIL, βIIITubulin or VEGFA was detected. When SCAP were incorporated 232
in bECM, no expression of the gene of interest was detected at Day 2 and Day 7. SNAIL, βIIITubulin, GDNF, 233
VEGFA and PDGFRa were expressed after 14 days of incubation in the hydrogel. SCAP incorporated in dECM 234
showed no expression of the genes of interest. Only when SCAP were grown in scECM a marked expression 235
of all the tested genes was detected after 7 days. 236
4 Discussion 237
The present study illustrates the influence of biological scaffolds derived from homologous and non-238
homologous tissue on the gene expression profile of dental stem cells from apical papilla (SCAP). Previously 239
published methods were employed to prepare ECM scaffolds derived from spinal cord (scECM) 20 and bone 240
(bECM) 19. A new decellularisation method was developed for the preparation of dentine ECM scaffolds 241
(dECM) based on the material characteristics and its similarity towards the bone tissue. 242
The acellular nature of bECM, dECM, and scECM was determined by quantifying residual dsDNA content. 243
The amount of dsDNA per mg of dry ECM present in bECM, dECM and scECM was considerably lower than 244
the <50 ng threshold previously determined as an effective criteria for decellularisation 28. The presence of 245
DNA and other cellular material, either xenogeneic or allogeneic in nature, within in the ECM scaffold due to 246
inefficient decellularisation could potentially elicit host immunological response. However, it is unlikely that 247
complete removal of cells will be obtained even with harsh decellularisation procedures 29. 248
The percentage of collagen and glycosaminoglycan in the ECM scaffolds derived from bone, spinal cord and 249
dentine varied depending upon the function and nature of tissue source. In addition to the intrinsic ECM 250
composition influencing sGAG content, the higher sGAG content in scECMh could be due to the 251
sterilised with peracetic acid induced a greater loss of cellular components such as lipids, nucleic acids, and 253
other undesirable elements ensuing more sGAG and FGF-2 compared to ECM non-treated with peracetic acid. 254
We postulate that the higher sGAG content in scECM with respect to bECM and dECM might be due to the 255
peracetic acid treatment during decellularization30. Also, we hypothesize that the differences observed between 256
ECM of different origins might be due to i) either different amounts of sGAG in the tissue before treatment or 257
ii) to the different treatments used to obtain each ECM30,31. 258
The gelation kinetics based on turbidimetry demonstrated that scECM, bECM and dECM hydrogels (Figure 259
2) exhibited sigmoidal gelation kinetics. Although the speed to complete gelation as well as t95 were greater 260
for scECMh and bECMh, the lag time as well as t1/2 of dECMh was considerably lower with respect to bECMh; 261
this is likely to be the effect of glycosaminoglycan presence during the in vitro self-assembly of collagen 262
fibres32. 263
It is noteworthy that the individual ECMs demonstrate distinct morphologies when analysed via scanning 264
electron microscopy. The basal lamina of the spinal cord is comprised predominantly of collagen IV though 265
types I and III are also present 33 whereas bone and dentine are composed primarily of collagen type I, with 266
traces of type III, V, IX, XII, XIV, XIX, and XXI 34. We hypothesise that scECMh contains Type IV collagen 267
that tends to form a flexible triple helix that self-assembles to incorporate structural glycoproteins 35, resulting 268
in the sheet-like structure observed in Figure 4C. However in bECMh and dECMh collagen XII, XIV, XIX, 269
and XXI serve as molecular bridges between collagen I monomers to produce fibrils. 270
Rheology was used to study the viscoelastic properties of ECM hydrogels. It is interesting to note that 271
regardless of the tissue origin, hydrogels exhibited similar sigmoidal shaped gelation profiles. However, the 272
storage modulus for dECMh is considerably lower than scECMh and bECMh. We postulate that this difference 273
is due to the disparity of source of tissue as well as the decellularisation processes. The potential impact of 274
hydrogel solid content has been evaluated but no difference was visible between hydrogels of different origins 275
of moduli, especially when considering the different morphologies observed with SEM36,37. Although the final 277
moduli of dECM and scECM are lower than 0.1kPa, the final modulus of bECMh was in the range of 0.1-278
1.0kPa. Hydrogels with elastic moduli within this range have been reported to have a positive influence on the 279
probability of neuronal marker expression and neuronal differentiation38. 280
Several authors have emphasised the intrinsic ability of ECM derived scaffolds to aid viability, migration and 281
proliferation of numerous cell types 9,39. Nevertheless, the present study is the first time ECM scaffolds from 282
decellularised bone, dentine and spinal cord have been used as a 3D culture system for apical papilla stem cells. 283
High cell viability (Figure 6) was observed in bECMh, scECMh and dECMh, confirming the suitability of 284
these scaffolds as delivery vehicles for SCAP. Greater cell viability was observed in bECMh compared to 285
dECMh and scECMh. The choice of detergents used for the decellularisation procedure could have a marked 286
deleterious effect up on the ECM 40. We hypothesise that the lower cell viability observed in scECMh may be 287
due to traces of residual detergent present after decellularisation. This is the first time that dentine has been 288
decellularised and further studies may be needed to optimise the duration as well as the concentration of HCl 289
solution used in the enamel removal step. We postulate that too harsh treatment applied during 290
decellularisation procedure may have led to the lower cell viability observed in dECMh 291
In both 2D and 3D culture environment, SCAP proliferated till day 7 irrespective of the culture conditions 292
(Figure 5). However, in the 2D environment, there was a remarkable increase in the proliferation between day 293
7 and day 14 for SCAP grown on TCP and dECMh. We postulate that after the initial phase of cell and ECM 294
hydrogel interaction, the surface with higher elasticity (dECMh) facilitated quicker cell migration and 295
proliferation compared to the more rigid surfaces of bECMh and scECMh. SCAP grown in 2D culture 296
proliferated faster than SCAP grown in 3D. Cell movement on 2D environment is relatively unrestricted; 297
whereas in a 3D environment, cells need to degrade their adjacent material enzymatically in order to migrate 298
41. Furthermore, in 2D culture, the cells are exposed to a homogenous concentration of nutrients which 299
environment 42. We also postulate that the porous structure of scECMh (Figure 4C) facilitated greater 301
proliferation rate in 3D environment compared to bECMh and dECMh. 302
A slight expression of some neural lineage markers by non-differentiated SCAP, attributed to their neural crest 303
origin, was reported previously 43. When SCAP were embedded into scECMh, there was a remarkable 304
expression of key neural lineage markers on day 7 and day 14 (Figure 7). We hypothesise that the potential 305
neurotrophic factors as well as proteoglycans embedded within scECMh have induced a neural-like phenotype. 306
ECM scaffolds derived from CNS tissues have already been reported to induce neuronal cell differentiation of 307
neuronal progenitor cells 9,20,23. Furthermore, decellularisation using peracetic acid might have resulted in a 308
higher concentration of FGF-2 as well as sGAG 44, which in turn enhanced the gene expression of neuronal 309
lineage markers in scECMh. On day 14, SCAP embedded in bECMh also showed an expression of neural 310
lineage genes. We hypothesise that the by-products of ECM degradation including soluble biomolecules in 311
conjunction with scaffold elasticity may influence SCAP differentiation towards neural phenotype. 312
5 Conclusion 313
Solubilised forms of decellularised ECM from spinal cord, dentine and bone tissues were prepared by a 314
combination of enzymatic and chemical processes and induced to form hydrogels. These acellular ECM 315
hydrogels had discrete structural, mechanical and biological characteristics. Human stem cells from apical 316
papilla exhibited a strong positive response to scECMh by proliferating within and on the hydrogel and by 317
expressing neural genes; substantiating the influence of neurotrophic factors present in decellularized spinal 318
cord. SCAP exhibited a greater proliferation rate in 2D in dECMh, compared to bECMh and scECMh, however 319
no neuronal differentiation occurred. Neuronal lineage markers were expressed by SCAP when cultured with 320
bECMh and it is hypothesized that the hydrogel mechanical properties combined with active biomolecules 321
provided the stimulus for this behaviour. The results of the present study show that decellularised ECM 322
hydrogels facilitate SCAP viability and proliferation. However, ECM hydrogels from site-specific tissues 323
325
Acknowledgements 326
The authors would like to thank Dr Christopher Medberry and Professor Stephen Badylak for assistance with 327
preparation of the spinal cord ECM. The authors also would like to thank MICA - Université Catholique de 328
Louvain platform for Scanning Electron Microscope facility. Aiswarya Viswanath is supported by NanoFar 329
"European Doctorate in Nanomedicine" EMJD programme funded by EACEA. This work is supported by 330
grants from the Université Catholique de Louvain (F.S.R), Fonds National de la Recherché Scientifique 331
(F.R.S.-FNRS). A. des Rieux is a F.R.S.-FNRS Research Associate and J. Vanacker is a F.R.S.-FNRS 332
Postdoctoral Researcher. J. Leprince is supported by la Fondation Saint Luc. 333
References 334
1. McCreedy DA, Sakiyama-Elbert SE. Combination therapies in the CNS: engineering the environment. Neurosci 335
Lett 2012;519(2):115-21. 336
2. Tator CH. Review of treatment trials in human spinal cord injury: issues, difficulties, and recommendations. 337
Neurosurgery 2006;59(5):957-82; discussion 982-7. 338
3. Mothe AJ, Tator CH. Advances in stem cell therapy for spinal cord injury. J Clin Invest 2012;122(11):3824-34. 339
4. De Berdt P, Vanacker J, Ucakar B, Elens L, Diogenes A, Leprince JG, Deumens R, des Rieux A. Dental Apical Papilla 340
as Therapy for Spinal Cord Injury. J Dent Res 2015;94(11):1575-81. 341
5. Huang GT-J, Gronthos S, Shi S. Mesenchymal Stem Cells Derived from Dental Tissues vs. Those from Other 342
Sources: Their Biology and Role in Regenerative Medicine. Journal of Dental Research 2009;88(9):792-806. 343
6. Sakai K, Yamamoto A, Matsubara K, Nakamura S, Naruse M, Yamagata M, Sakamoto K, Tauchi R, Wakao N, 344
Imagama S and others. Human dental pulp-derived stem cells promote locomotor recovery after complete 345
transection of the rat spinal cord by multiple neuro-regenerative mechanisms. J Clin Invest 2012;122(1):80-90. 346
7. Martens W, Bronckaers A, Politis C, Jacobs R, Lambrichts I. Dental stem cells and their promising role in neural 347
8. Sonoyama W, Liu Y, Yamaza T, Tuan RS, Wang S, Shi S, Huang GT. Characterization of the apical papilla and its 349
residing stem cells from human immature permanent teeth: a pilot study. J Endod 2008;34(2):166-71. 350
9. Crapo PM, Tottey S, Slivka PF, Badylak SF. Effects of biologic scaffolds on human stem cells and implications for 351
CNS tissue engineering. Tissue Eng Part A 2014;20(1-2):313-23. 352
10. Bible E, Dell'Acqua F, Solanky B, Balducci A, Crapo PM, Badylak SF, Ahrens ET, Modo M. Non-invasive imaging 353
of transplanted human neural stem cells and ECM scaffold remodeling in the stroke-damaged rat brain by (19)F- 354
and diffusion-MRI. Biomaterials 2012;33(10):2858-71. 355
11. Sicari BM, Dziki JL, Siu BF, Medberry CJ, Dearth CL, Badylak SF. The promotion of a constructive macrophage 356
phenotype by solubilized extracellular matrix. Biomaterials 2014;35(30):8605-8612. 357
12. Wolf MT, Carruthers CA, Dearth CL, Crapo PM, Huber A, Burnsed OA, Londono R, Johnson SA, Daly KA, Stahl EC 358
and others. Polypropylene surgical mesh coated with extracellular matrix mitigates the host foreign body 359
response. J Biomed Mater Res A 2014;102(1):234-46. 360
13. Singelyn JM, Sundaramurthy P, Johnson TD, Schup-Magoffin PJ, Hu DP, Faulk DM, Wang J, Mayle KM, Bartels 361
K, Salvatore M and others. Catheter-deliverable hydrogel derived from decellularized ventricular extracellular 362
matrix increases endogenous cardiomyocytes and preserves cardiac function post-myocardial infarction. J Am 363
Coll Cardiol 2012;59(8):751-63. 364
14. Wang RM, Christman KL. Decellularized myocardial matrix hydrogels: In basic research and preclinical studies. 365
Adv Drug Deliv Rev 2015. 366
15. Ballios BG, Cooke MJ, Donaldson L, Coles BL, Morshead CM, van der Kooy D, Shoichet MS. A Hyaluronan-Based 367
Injectable Hydrogel Improves the Survival and Integration of Stem Cell Progeny following Transplantation. Stem 368
Cell Reports 2015;4(6):1031-45. 369
16. DeQuach JA, Yuan SH, Goldstein LS, Christman KL. Decellularized porcine brain matrix for cell culture and tissue 370
engineering scaffolds. Tissue Eng Part A 2011;17(21-22):2583-92. 371
17. Sellaro TL, Ranade A, Faulk DM, McCabe GP, Dorko K, Badylak SF, Strom SC. Maintenance of Human Hepatocyte 372
18. Sellaro TL, Ravindra AK, Stolz DB, Badylak SF. Maintenance of hepatic sinusoidal endothelial cell phenotype in 374
vitro using organ-specific extracellular matrix scaffolds. Tissue Eng 2007;13(9):2301-10. 375
19. Sawkins MJ, Bowen W, Dhadda P, Markides H, Sidney LE, Taylor AJ, Rose FR, Badylak SF, Shakesheff KM, White 376
LJ. Hydrogels derived from demineralized and decellularized bone extracellular matrix. Acta Biomater 377
2013;9(8):7865-73. 378
20. Medberry CJ, Crapo PM, Siu BF, Carruthers CA, Wolf MT, Nagarkar SP, Agrawal V, Jones KE, Kelly J, Johnson SA 379
and others. Hydrogels derived from central nervous system extracellular matrix. Biomaterials 2013;34(4):1033-380
40. 381
21. Bazos P, Magne P. Bio-emulation: biomimetically emulating nature utilizing a histo-anatomic approach; 382
structural analysis. Eur J Esthet Dent 2011;6(1):8-19. 383
22. Hong Y, Huber A, Takanari K, Amoroso NJ, Hashizume R, Badylak SF, Wagner WR. Mechanical properties and in 384
vivo behavior of a biodegradable synthetic polymer microfiber-extracellular matrix hydrogel biohybrid scaffold. 385
Biomaterials 2011;32(13):3387-94. 386
23. Crapo PM, Medberry CJ, Reing JE, Tottey S, van der Merwe Y, Jones KE, Badylak SF. Biologic scaffolds composed 387
of central nervous system extracellular matrix. Biomaterials 2012;33(13):3539-3547. 388
24. Woessner JF, Jr. The determination of hydroxyproline in tissue and protein samples containing small 389
proportions of this imino acid. Arch Biochem Biophys 1961;93:440-7. 390
25. Farndale RW, Sayers CA, Barrett AJ. A Direct Spectrophotometric Micro-Assay for Sulfated Glycosaminoglycans 391
in Cartilage Cultures. Connective Tissue Research 1982;9(4):247-248. 392
26. Wolf MT, Daly KA, Brennan-Pierce EP, Johnson SA, Carruthers CA, D'Amore A, Nagarkar SP, Velankar SS, Badylak 393
SF. A hydrogel derived from decellularized dermal extracellular matrix. Biomaterials 2012;33(29):7028-38. 394
27. Ruparel NB, de Almeida JF, Henry MA, Diogenes A. Characterization of a stem cell of apical papilla cell line: 395
effect of passage on cellular phenotype. J Endod 2013;39(3):357-63. 396
28. Crapo PM, Gilbert TW, Badylak SF. An overview of tissue and whole organ decellularization processes. 397
29. Badylak SF, Gilbert TW. Immune response to biologic scaffold materials. Seminars in Immunology 399
2008;20(2):109-116. 400
30. Engfeldt B, Hjerpe A. Glycosaminoglycans of dentine and predentine. Calcif Tissue Res 1972;10(2):152-9. 401
31. Liu Z, Masuko S, Solakyildirim K, Pu D, Linhardt RJ, Zhang F. Glycosaminoglycans of the porcine central nervous 402
system. Biochemistry 2010;49(45):9839-47. 403
32. Stuart K, Panitch A. Influence of chondroitin sulfate on collagen gel structure and mechanical properties at 404
physiologically relevant levels. Biopolymers 2008;89(10):841-51. 405
33. Weidner N, Grill RJ, Tuszynski MH. Elimination of basal lamina and the collagen "scar" after spinal cord injury 406
fails to augment corticospinal tract regeneration. Exp Neurol 1999;160(1):40-50. 407
34. Clarke B. Normal bone anatomy and physiology. Clin J Am Soc Nephrol 2008;3 Suppl 3:S131-9. 408
35. Burnside ER, Bradbury EJ. Manipulating the extracellular matrix and its role in brain and spinal cord plasticity 409
and repair. Neuropathol Appl Neurobiol 2014;40(1):26-59. 410
36. Black LD, Allen PG, Morris SM, Stone PJ, Suki B. Mechanical and failure properties of extracellular matrix sheets 411
as a function of structural protein composition. Biophys J 2008;94(5):1916-29. 412
37. Marinkovic M, Block TJ, Rakian R, Li Q, Wang E, Reilly MA, Dean DD, Chen XD. One size does not fit all: 413
developing a cell-specific niche for in vitro study of cell behavior. Matrix Biol 2016;52-54:426-41. 414
38. Engler AJ, Sen S, Sweeney HL, Discher DE. Matrix elasticity directs stem cell lineage specification. Cell 415
2006;126(4):677-89. 416
39. Tukmachev D, Forostyak S, Koci Z, Zaviskova K, Vackova I, Vyborny K, Sandvig I, Sandvig A, Medberry CJ, Badylak 417
SF and others. Injectable Extracellular Matrix Hydrogels as Scaffolds for Spinal Cord Injury Repair. Tissue Eng 418
Part A 2016;22(3-4):306-17. 419
40. Faulk DM, Carruthers CA, Warner HJ, Kramer CR, Reing JE, Zhang L, D'Amore A, Badylak SF. The effect of 420
detergents on the basement membrane complex of a biologic scaffold material. Acta Biomater 2014;10(1):183-421
93. 422
41. Walters NJ, Gentleman E. Evolving insights in cell-matrix interactions: elucidating how non-soluble properties 423
42. Tibbitt MW, Anseth KS. Hydrogels as extracellular matrix mimics for 3D cell culture. Biotechnol Bioeng 425
2009;103(4):655-63. 426
43. Germain L, De Berdt P, Vanacker J, Leprince J, Diogenes A, Jacobs D, Vandermeulen G, Bouzin C, Préat V, 427
Dupont-Gillain C and others. Fibrin hydrogels to deliver dental stem cells of the apical papilla for regenerative 428
medicine. Regenerative Medicine 2015;10(2):153-167. 429
44. Hodde J, Janis A, Ernst D, Zopf D, Sherman D, Johnson C. Effects of sterilization on an extracellular matrix 430
scaffold: part I. Composition and matrix architecture. J Mater Sci Mater Med 2007;18(4):537-43. 431
Tables and Figures 434
[image:20.612.43.498.135.677.2]435
Table 1: Human genes and primers used for RT-PCR analysis 436
437
Symbol Gene name Primer Sequence (5’->3’) Amplicon
length (bp)
GAPDH GAPDH = Glyceraldehyde
3-phosphate dehydrogenase
F : GATCAACTCACCGCCAACA
R : CCAGCGACTCAATCTTCCTC
452
SNAIL Zinc finger protein SNAI1 F : CTAGGCCCTGGCTGCTACAA
R :CATCTGAGTGGGTCTGGAGGT 129
BIII
Tubulin
F : GGGCCTTTGGACATCTCTTC
R : GGATACTCCTCACGCACCTT
244
PDGFRa Platelet-Derived Growth
Factor Receptor, Alpha
Polypeptide
F : AGCTTGAAGGCAGGCACA
R :GCGACAAGGTATAATGGCAG
A
122
VEGFA Vascular endothelial growth
factor A
F : CATCTTCAAGCCATCCTGTG
R : CAACGCGAGTCTGTGTTTT
303,357,
375,426
GDNF Glial cell derived
neurotrophic factor
F : CAGACATCTGATCCCCTTCC
R : GCTGGGGTTTGTCACTGTTT
172
441
Table 2: Gelation kinetics for scECM, bECM and dECM hydrogels 442
443
Turbidity (Mean± SD) scECM (Mean± SD) bECM (Mean± SD) dECM (Mean± SD)
Lag phase (t lag: min) 11.998 ± 1.90 11.25 ± 1.4 7.8 ± 1.55
50% gelation (t ½: min) 20.582 ± 2.42 21.06 ± 3.2 14.60 ± 1.24
95% gelation (t 95: min) 30.248 ± 2.91 36.00± 0.00 39.85 ± 3.34
Speed (S, min-1) 0.0738 ± 0.009 0.057 ± 0.003 0.035 ± 0.004
n=8, p<0.05 444
Table 3: Influence of ECM origin on hydrogel final moduli 446
Material Storage modulus (G’, Pa) Loss modulus (G’’, Pa)
447
bECM (8mg/ml) 287.17 ± 8.43 A 42.61 ± 3.72 A 448
dECM (8mg/ml) 4.79 ± 0.33 B 0.56 ± 0.128 B 449
scECM (8mg/ml) 19.35 ± 1.16 C 2.40 ± 0.385 C 450
ECM not related by the same letter are significantly different (G’ and G’’ separately) (p<0.05; n=3). 451
453
[image:23.612.67.513.70.543.2]454
Figure 1: Influence of ECM origin on cellular, collagen and sGAG content. 455
(A) Efficiency of decellularisation was assessed by quantifying the amount of dsDNA/mg dry weight. B) 456
Collagen content and (C) sGAG content of ECM scaffolds were evaluated by quantifying hydroxyproline and 457
using the 1, 9 –Dimethyl methylene blue assay, respectively. Data is presented as Mean ± standard 458
deviation (n=3). Conditions not linked by the same letter are significantly different (p<0.05). 459
461
Figure 2: Turbidimetric gelation kinetics of scECM, bECM, and dECM scaffolds were compared at 8mg/ml 462
concentrations. The neutralised pre-gel solution was added to a 96 well plate and gelation was induced at 463
370C. The absorbance was measured at 405nm and recorded at 3min intervals. The data was normalised 464
between 0 (initial absorbance) and 1 (maximum absorbance). Data represents mean ± SD (n=8). Inset: 465
Figure 3: Influence of ECM origin on hydrogel mechanical properties. 468
Rheological properties of hydrogels were investigated by small amplitude oscillatory shear rheology. 469
The storage (G′) and loss moduli (G′′) were measured using a Kinexus Pro rotational rheometer and 470
rSpace software (n=3). 471
473
[image:27.612.54.529.61.492.2]474
Figure 4: Influence of ECM origin on the hydrogel microstructure. 475
ECM hydrogels were formed and processed for SEM analysis, scale bar = 1 μm with (A) bECM, (B) dECM 476
478
Figure 5: Influence of ECM origin on SCAP proliferation. 479
A- 500,000 SCAP/ml were incorporated in different ECM hydrogels and incubated at 37°C. Presto blue 480
test was performed at different time points (2, 7 and 14 days). Fluorescence intensities were measured 481
at 590 nm (n=6, N=3). B- Pre-gel solutions were allowed to gel at 37°C. Then 2000 SCAP were seeded 482
on the ECM. After 2, 7 and 14 days, cell proliferation was analysed using a Presto Blue kit (n=6, N=3). 483
As a positive control, the same amount of cells was seeded on tissue culture plastic. Conditions not 484
related by the same letter are significantly different (p < 0.05). 485
487
Figure 6: Influence of ECM origin on SCAP viability. 488
500,000 SCAP/ml were incorporated in different ECM hydrogels and incubated for 2 days. A 489
Live/Dead assay was performed. Living and dead cells were observed by confocal microscopy (488 490
and 568 nm, respectively) (n=3, N=3). (A) Live cells were stained with Calcein (green) and dead cells 491
were stained with ethidium homodimer-I (red). (B) The images were processed to quantify the number 492
of live cells and dead cells using ImageJ (n = 3–5 acquisitions/sample). Conditions not related by the 493
same letter are significantly different (p< 0.05). 494
496
Figure 7: Influence of ECM origin on SCAP gene expression. 497
500,000 SCAP/ml were incorporated in different ECM hydrogels and incubated for 2, 7 and 14 days 498
(D2, D7 and D14). mRNA were extracted and gene expression was analysed by RT-PCR (n=3, N=3). 499
Blanks (Blk) were obtained by replacing mRNA by RNAse free water. Gene expression of SCAP 500