TH E ROLE OF BONE MORPHOGENETIC PROTEIN-4 IN
THE DEVELOPING CHICK EYE
LI M ING LEONG
THESIS SUBM ITTED TO TH E UNIVERSITY O F LONDON FOR TH E D EG REE OF Ph.D
Department of Biochemistry and M olecular Biology University College London
ProQuest Number: 10044354
All rights reserved
INFORMATION TO ALL USERS
The quality of this reproduction is dependent upon the quality of the copy submitted.
In the unlikely event that the author did not send a complete manuscript
and there are missing pages, th ese will be noted. Also, if material had to be removed, a note will indicate the deletion.
uest.
ProQuest 10044354
Published by ProQuest LLC(2016). Copyright of the Dissertation is held by the Author.
All rights reserved.
This work is protected against unauthorized copying under Title 17, United States Code. Microform Edition © ProQuest LLC.
ProQuest LLC
789 East Eisenhower Parkway P.O. Box 1346
A B S T R A C T
B o n e morphogenetic protein-4 (Bm p-4) is a member of the Transform ing
Growth Factor-6 (TGF-6) superfamily of secreted signalling molecules. In this w ork, I
have investigated its spatiotemporal distribution and possible function in the developing
chick eye. The chick homologue of B m p -4 is expressed throughout the developing
chicken optic vesicle between Ham burger and Hamilton stages 13 and 17. It then
show s a distinctive restriction to the dorsal quadrant o f the eye until stage 30, w hen
expression begins to fade, becom ing undetectable by stage 35. Transcripts o f the
homeobox-containing gene G H 6 show a similar dorsal restriction to B m p -4 , but
between stages 20-35. Overexpression of Bm p-4 in vivo results in no detectable change
in eye m orphology, but enlarges the expression dom ain o f GH6, indicating that the two
genes may form part of a gene cascade involved in patterning the eye. In vitro, BM P-4
appears to stimulate retinal cell proliferation. It has no effect on retinal cell
differentiation in vitro. In the chick, the patterning activities o f sonic hedgehog (shh)
are know n to involve activation of B m p expression in at least two instances; the
developm ent of the limb and midgut. I report here that shh is not expressed in the chick
eye at all during development, but is found in the ventral portion of the diencephalon
(forebrain).
The available data suggests a number o f roles for BMP-4 in chick eye
development. BM P-4 may specify dorso-ventral retinal polarity required fo r proper
retinal axon pathfinding, or establish a dorso-ventral gradient o f cell proliferation in the
developing optic cup. Alternatively, it may act as a hedgehog family member antagonist
TABLE OF CONTENTS
TITLE PAGE
Page number
1
A B S T R A C T
C O N T E N T S
LIST OF ABBR EV IA TIO N S
LIST OF FIGURES
7
11
LIST OF TABLES
A C K N O W L ED G E M EN T S
15
16
CH APTER 1 - INTRO DUCTIO N AND AIM S 18
1.1 The Bone M orphogenetic Proteins 19
1.1.1 H ISTO RY AND RO LE IN BONE FO RM A TIO N 20
1.1.2 FA M ILIES W ITH IN T H E TO F^ SU PERFA M ILY
-A N OVERVIEW 21
1.1.2.1 The T G F-p subfamily 23
1.1.2.2 The A ctivin subfam ily 23
1.1.2.3The D VR group 23
1.1.2.4 O ther m ore divergent genes 26
1.1.3 STR U C TU R E OF TH E BM PS 27
1.1.4 BM PS IN B O N E FO R M A TIO N 31
1.1.5 BM PS AS SIGN A LLIN G M OLECULES IN D EV ELO PM EN T 34
1.1.5.1 BM P-1 34
1.1.5.2 B M P-2 36
1.1.5.3 BM P-3 38
1.1.5.4 B M P-4 38
1.1.5.5 BM P-5 43
1.1.5.6 BM P-6 43
1.1.5.7 BM P-7 44
1.1.5.8 BM P-8, 9 and 10 45
1.1.6 T H E BM P RECEPTO RS 45
1.1.7 D O W N STR EA M CO M PO N ENTS OF BM P SIGN A LLIN G 50
1.2 Eye D evelopm ent 50
1.2.1 EA RLY EV EN TS IN M ORPHOGENESIS O F TH E AVIAN
EYE 51
1.2.2.1 M olecules that affect early phases of eve induction 1.2.2.2 M olecules that affect lens and retina induction 1.2.2.3 Patterning within the developing eve
1.2.2.3.1 The N aso-tem poral axis 1.2.2.3.2 The D orso-ventral axis 1.2.2.3.3 The Lam inar A xis 1.2.2.4 Cell death and patterning 1.2.3 CLUES FRO M D ROSO PHILA
1.2.3.1 hh and shh
1.2.3.2 dpp and B m p -21 B m p -4
1.2.3.3 H om eobox genes
57 62 64 65 70 74 77 77 79 81 82
1.3 A im s 85
C H A P T E R 2 - M A T E R IA L S A N D M E T H O D S
88
2.1 M aterials
2.1.1 EM BRY O S
2.1.2 B A C TER IA L STRAINS 2.1.3 PLA SM ID S
2.1.4 CELL CU LTU RE REA G EN TS 2.1.5 EN ZY M ES
2.1.6 RA DIOCHEM ICALS 2.1.7 SOLUTION S AND M EDIA
2.1.7.1 General M olecular Biologv 2.1.7.2 Plasm id Preparation
2.1.7.3 In-Situ Hybridisation
2.1.7.4 Bacterial Culture Media 2.1.7.5 Cell Culture
2.1.7.6 Im m unofluorescence assay 2.1.7.7 Solutions For Retroviral W ork 2.1.8 M ISC ELLA N EO U S
88
88
88
88
89 89 89 90 90 90 90 91 91 91 92 922 .2 M olecular Biology Techniques 92
2.2.1 RESTRICTIO N EN ZY M E DIGESTION 92
2.2.2 A G A R O SE GEL ELECTROPHORESIS 93
2.2.3 LARGE SCA LE PLAM ID PREPARATION 94
2.2.4 SM ALL SCA LE PLAM ID PREPARATION 96
2.2.5 TRA N SFO RM A TIO N OF E .C O LI W ITH PLASM ID D N A 97
2 .2.6 IN -S IT U HYBRIDISATION TO SECTIONS 98
2.2.6.1 Tissue and section preparation 98
2.2.6.2 Preparation of Slides and Sections 98
2.2.6.3 Probe Preparation 99
2.2.6.4 Pretreatm ent 100
2.2.6.5 H ybridisation 101
2.2.6.6 Post-hybridisation W ashes 102
2.2.6.7 D ipping, Exposure, Developm ent and Staining 102
2.2.7 IN -S IT U HYBRIDISATION TO W HOLE M OUNTS 103
2.2.7.1 Tissue preparation 103
2.2.7.2 Probe preparation 103
2.2.7.3 Pretreatm ent 104
2.2.1 A H ybridisation 104
2.2.7.5 W ashes and antibody reaction 105
2.2.7.6 C olour Reaction 105
2.3 C e ll C u ltu r e T e c h n iq u e s 106
2.3.1 EM B R Y O N IC DISSECTION 107
2.3.2 C U L T U R E OF RETINA L TISSUE 107
2.3.3 RADIO A CTIV E CELL PROLIFERATION A SSA Y 108
2.3.4 A N TIBO D Y DETECTION OF M ATURE NEURONS 108
2.4 R e t r o v i r a l W o r k 109
2.4.1 C U LTU R E OF AVIAN FIBROBLASTS 109
2.4.2 FIB R O B LA ST TRANSFECTION 111
2.4.3 CO N CENTRATION OF VIRAL SU PERN A TEN T 112
2.4.4 GAG STAINING TO ESTIM ATE V IRA L SPREA D 112
2.4.5 T IT E R IN G OF VIRUS 113
2.4.6 IN JE C TIO N OF VIRAL SUPERNATENT 113
2.4.7 G R A FTIN G OF TRANSFECTED FIBRO B LA STS 114
2.5 P h o to m ic r o s c o p y 115
C H A P T E R 3 - B M P -4 AND R E L A T E D S IG N A L L IN G
F A C T O R S IN T H E E Y E 117
3.1 I n tr o d u c t i o n 118
3.2 D is tr ib u tio n O f B m p -4 tr a n s c r ip ts in th e
d e v e lo p in g c h ic k eye 121
3.3 D is tr ib u tio n O f G h6 tr a n s c r ip ts I n th e
d e v e lo p in g c h ic k eye 134
3.4 D is tr ib u tio n o f S h h tr a n s c r ip ts in th e
d e v e lo p in g c h ic k eye 143
3 .5 D is c u s s io n 143
C H A P T E R 4 - E F F E C T O F B M P -4 O N C H IC K
R E T I N A L C E L L S IN C U L T U R E 157
4.1 I n tr o d u c t i o n 157
4.2 R e s u lts 160
4.2.1R E T IN A L CELL CULTURES 160
4.2.2 T H E EFFECTS OF BM P-4 AND BM P-2 ON C H IC K
R ETIN A L CELL CULTURES 161
4.2.2.1 Effects of BM P-4 on cell number 164
4.2.2.2 Cell proliferation experiments 170
4.2.2.3 Effects on retinal cell differentiation 186
4.2.3 CO M PA RISO N OF THE EFFECTS O F V ARIO U S G RO W TH FACTORS WITH THAT OF BM P-4 ON TH E
G R O W TH O F CH ICK RETINAL CELS IN CU LTU RE 197
4.2.3.2 Cell proliferation experiments 220
4.2.3.3 Effects on retinal cell differentiation 220
4 .3 D is c u s s io n 237
4.3.1 G ROW TH OF CH ICK RETINA L CELLS IN CU LTU RE 237
4.3.2 RETINAL CELL REPONSE TO BM P-4 237
4.3.3 DOSE RESPONSE TO BM P-4 SEN SE V IRU S 241
4.3.4 A COM PARISON OF THE EFFECTS OF BM P-2
PROTEIN AND BM P-4 SENSE VIRUS ON R ETIN A L CELLS 243
4.3.5 M ITOGENIC EFFECTS OF BFGF. T G F -b l A N D EO F 244
4.3.6 IN SUM M ARY 248
C H A P T E R 5 - E F F E C T O F B M P -4 O N T H E C H I C K
E Y E I N - V I V O 250
5.1 I n tr o d u c t io n 251
5 .2 R e s u lts 255
5.2.1 USE OF RCASBPfAl VECTORS TO IN FEC T
C H ICK RETINA 255
5.2.1.1 Injections/Grafts into Embryos betw een stages 9-13 258
5.2.1.2 Iniection/Grafts into Embryos between stages 14-20 260
5.2.2 EFFECTS OF IM PLANTING BEADS C O N TA IN IN G
BM P-2 263
5.3 D is c u s s io n 274
5.3.1 RETROVIRAL DATA 274
5.3.2 EFFECTS OF BM P-2 PROTEIN 276
5.3.3 IN SUM M ARY 278
C H A P T E R 6 - D IS C U S S IO N 280
6.1 G e n e r a l D isc u ssio n 281
6.1.1 NON-POSITIO N AL M OLECULES EX PRESSED IN
GRADIENTS ACROSS THE RETINA 282
6.1.2 "SPERRY"-TYPE POSITIONAL M OLECU LES 284
6.1.2.1 “Sperry m olecules” and positional inform ation 284
6.1.2.2 Downstream candidates for BM P-4 positiional
specification 285
6.1.2.2.1The TOP antigens 286
6.1.2.2.2 Eph receptor tyrosine kinases 287
6.1.2.2.3 The 33kD protein 287
6.1.2.2.4 Cell adhesion molecules 288
6.1.3 M OLECULES TH A T INFLUENCE RETIN A L PO LA RITY 291
6.1.4 TH E IN TERACTION OF BMP-4 W ITH S H H A N D G H 6 294
6 .2 C o n c lu s io n s 296
6.3 F u tu r e W o r k 297
7
LIST OF A BBR EV IA TIO N S
ADMP: AER: aFGF: AHD: ALK: ALSV: ALV: AMY: anti-BM P-4/RCAS: A/P: bFG F: BMP: BM P-4/RCAS: Brk: bp: C-terminal: CAM: CEF: CFU: CMS: CRABP: CRBP: DABCO: dhh: dig:
anti-dorsalising-m orphogenetic protein
apical ectodermal ridge
acidic FG F
aldehyde dehydrogenase
activin receptor-like kinase
avian leukosis and sarcoma virus
avian leukosis virus
avian m yeloblastosis virus
RCA SBP(A ) vector containing the m ouse Bm p-4 in the
antisense orientation
anterior-posterior
basic fibroblast growth factor
bone m orphogenetic protein
RCA SBP(A ) vector containing the m ouse Bm p-4 in the
sense orientation
BMP receptor kinase
base pair
carboxy terminal
cell adhesion molecule
chick em bryonic fibroblast
colony-form ing units
central nervous system
cellular retinoic acid binding protein
cellular retinol binding protein
diazobicyclooctane
desert hedgehog
8
Dll: distaless
DMEM: Dulbecco's modified eagle medium
DPP: decapentaplegic
DSL: dorsalin
D/V: dorso-ventral
EGF: epidermal growth factor
Egr: early growth response
ems: empty spiracles
ELAV: embryonic lethal abnormal visual system
ELF: Eph ligand family
en: engrailed
ey\ eyeless
FG F: fibroblast growth factor
GDF: growth and differentiation factor
GDNF: glial-derived-neurotrophic factor
Gly: glycine
H BS: H EPES-buffered saline
H E S S : Hank's balanced saline solution
H H: hedgehog
H N F: hepatic nuclear factor
Hox: homeobox containing gene or protein
IG F: insulin-like growth factor
ihh: Indian hedgehog
IL: interleukin
kb: kilobase
LB: Luria-Bertani
Ld: Limb deformity
LTR: long terminal repeat
MIS: M sx: MAV: N-terminal: NCAM: nrl:
N G F:
OP: otd: Pcix: PBS: PBST: PFA; prd: p.c.: PEG: POU: RA: RAGS: RAR: RCAS: RCASBP: RSV: RTK: RXR: scix: SCW:
Mullerian inhibiting substance
m uscle segm ent hom eobox
myeloblastosis-associated helper virus
amino-terminal
neural cell adhesion molecule
neural retina leucine-zipper
nerve growth factor
osteogenic protein
orthodenticle
paired-box containing
phosphate-buffered saline
phosphate-buffered saline with tween
paraformaldehyde paired box post-coitum polyethylene glycol Pit-Oct-Unc quail retina-specific retinoic acid
repulsive axon guidance signal
retinoic acid receptor
replication competent ALV LTR containing a splice acceptor
replication competent ALV LTR containing a splice acceptor and
p o l gene from Bryan high-titre strain
Rous Sarcom a virus
receptor tyrosine kinase
retinoid X receptor
saxophone
10
se: short ear mutation
Sen serine
Sey: small eye
shn: schmirri
shs: short-sighted
Six: sine ocidis homeobox containing gene
SO Ho: sensory organ hom eobox
SHH: sonic hedgehog
TBE: Tris-Borate-EDTA
TESPA: 3-aminopropyl trietoxysilane
TFR: transferrin receptor
TGF: transforming growth factor
tkv: thick veins
TRAP: temporal retinal axon protein
twhh: tiggy-winkle hedgehog
UV: ultraviolet
VG: vegetal
Xbra: X enopus brachyiiry
Xbar: Xenopiis dualbar
X enopus forkhead
X h o x : X enopus hox
X vent: X enopus ventral
X m o x : X en opus m esoderm derived hox
WD: W D-40 domain of G-protein
LIST OF FIG URES
11
Figure P age number
1.1 Structure o f TGF-P superfamily members
1.2 Type I and type II receptors for TGF-P superfamily members
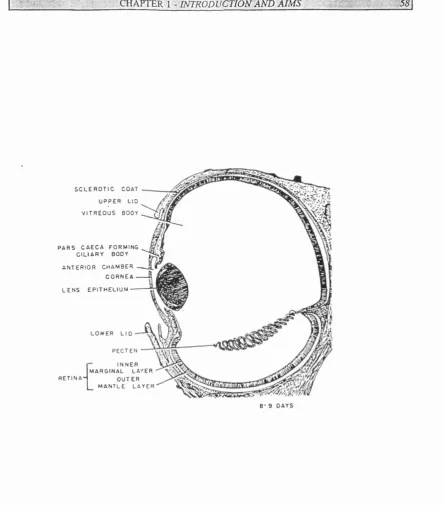
1.3 Schem atic diagrams showing the development o f the chick eye
1.4 Schem atic diagram o f the chick eye at stage 35
1.5 The major axes o f the eye
29
47
52
58
59
3.1 Expression o f B m p-4 and GH6 in frontal head sections as detected by
Francis-West et a/(1994) and Stadler and Solursh (1994)
3.2 Diagram illustrating plane o f sectioning
3.3 B m p-4 expression in stage 13 eyes
3 .4 B m p -4 expression in a stage 15 eye
3.5 B m p -4 expression in a stage 17 eye
3.6 B m p-4 expression in a stage 19-20 eye
3.7 Diagrams illustrating B m p-4 expression in stage 20 and 24 eyes
3.8 B m p -4 expression in a stage 24 eye
3 .9 B m p-4 expression in a stage 24 eye (closeup)
2.1Q B m p -4 expression in a stage 2 8 eye
3.1 1 B m p -4 expression in a stage 3 0 eye
3.12 B m p -4 expression in a stage 35 eye
3.1 3 G H 6 expression in a stage 17 eye
3 .1 4 G H 6 expression in a stage 20 eye
3.15 G H 6 and B m p-4 expression in a stage 24 eye
3 .16 G H 6 expression in a stage 24 eye (closeup)
3 .17 G H 6 and Brnp-4 expression in a stage 28 eye
3 .18 G H 6 and Brnp-4 expression in a stage 30 eye
12
3 .1 9 G H 6 and B m p-4 expression in a stage 35 eye 146
3 .2 0 shh expression in stage 24 forebrain 147
3.21 shh expression in stage 30 forebrain 148
3 .22 shh expression in stage 35 forebrain 149
3.2 3 shh expression in stage 17 forebrain 150
4.1 Neurofilam ent-labelled chick retinal cell culture 162
4.2 Graph sh ow ing changes in cell number over 8 days in culture 163
4.3 Graphs com paring cell number in stage 26 cultures treated with BM P-4/RC A S and controls 165
4.4 Bar charts sh ow ing changes in cell number in stage 26
BM P-4/RC AS treated cultures and controls 167
4.5 Graphs com paring cell number in stage 35 cultures treated with BM P-4/RC A S and controls 171
4.6 Bar charts show ing changes in cell number in stage 35
BM P-4/R C A S treated cultures and controls 173
4.7 Graphs com paring cell proliferation in stage 26 cultures treated with
BM P-2 protein, BM P-4/R C A S and controls 176
4.8 Bar charts show ing changes in cell proliferation in stage 26 cultures treated
with B M P -2 protein, BM P-4/RCA S and controls 178
4.9 Graphs com paring cell proliferation in stage 35 cultures treated with
BM P-2 protein, B M P-4/R C A S and controls 181
4.10 Bar charts show ing changes in cell proliferation in stage 35 cultures treated
with B M P -2 protein, BM P-4/RC A S and controls 183
4.11 Graphs comparing differentiation in stage 26 cultures treated with
BM P-2 protein, BM P-4/R C A S and controls 187
4.12 Bar chart and photographs showing differentiation in stage 26 cultures treated with
BM P-2 protein, BM P-4/R C A S and controls 1 day after treatment 189
4.13 Bar chart show ing differentiation in stage 26 cultures treated with
BM P-2 protein, B M P-4/RC A S and controls 2 days after treatment 192
13
BM P-2 protein, BM P-4/RCA S and controls 3 days after treatment 193
4.15 Bar chart show ing differentiation in stage 26 cultures treated with
BM P-2 protein, BM P-4/RCA S and controls 4 days after treatment 196
4.16 Bar chart and photographs show ing differentiation in stage 26 cultures treated with
BM P-2 protein, BM P-4/RCA S and controls 5 days after treatment 198
4.17 Graphs comparing differentiation in stage 35 cultures treated with
BM P-2 protein, BM P-4/RC AS and controls 201
4.18 Bar chart show ing differentiation in stage 35 cultures treated with
BM P-2 protein, BM P-4/RC A S and controls immediately after treatment 203
4.19 Bar chart and photographs showing differentiation in stage 35 cultures treated with
BM P-2 protein, BM P-4/RC A S and controls 1 day after treatment 204
4.20 Bar chart show ing differentiation in stage 35 cultures treated with
BM P-2 protein, BM P-4/RC A S and controls 2 days after treatment 207
4.21 Bar chart and photographs showing differentiation in stage 35 cultures treated with
BM P-2 protein, BM P-4/RC AS and controls 3 days after treatment 208
4.22 Bar chart show ing differentiation in stage 35 cultures treated with
BM P-2 protein, BM P-4/RC AS and controls 4 days after treatment 211
4.23 Bar chart and photographs showing differentiation in stage 35 cultures treated with
BM P-2 protein, BM P-4/RC AS and controls 5 days after treatment 212
4.24 Graphs comparing changes in cell number in cultures treated
with various growth factors 216
4.25 Graphs comparing changes in cell proliferation in cultures treated
with various growth factors 221
4.26 Graphs comparing changes in differentiation in cultures treated
with various growth factors 225
4.27 Phase contrast and neurofilament-labelled photographs o f cultures
treated with various growth factors 1 day after treatment 229
4.28 Phase contrast and neurofilament-labelled photographs o f cultures
14
5.1 Schem atic diagram o f RCASBP proviral genom e 253
5.2 Bar chart depicting percentage mortality o f embryos 3 days after injections/grafts 25 7
5.3 Stage 12 embryos microinjected with BM P-4/RCAS viral
supernatant and stained for virus-positive cells 259
5.4 Stage 24 embryo grafted with BM P-4/RC AS virus-producing
cells and probed for virus-positive cells 261
5.5 Stage 21 embryo grafted with control virus-producing cells and
probed for virus-positive cells 2 6 2
5.6 Stage 25 embryo grafted with BM P-4/RCAS virus-producing
cells and probed for virus-positive cells 2 6 4
5.7 Stage 28 embryo microinjected with BM P-4/RCAS viral
supernatant and stained for virus-positive cells 2 6 5
5.8 Stage 30 embryo microinjected with BM P-4/RCAS viral
supernatant and stained for virus-positive cells 2 6 6
5.9 Stage 35 embryos microinjected with BM P-4/RCAS viral
supernatant and stained for virus-positive cells 2 6 7
5.10 Stage 28 eye with BM P-2 soaked bead inserted and probed for G H 6 2 7 0
15
LIST OF TABLES
Table Page number
1.1 Organisation o f the TGF-P superfamily 22
1.2 R ole o f BM Ps in vertebrate bone formation 33
1.3 Multiple roles o f BM Ps in patterning the developing embryo 35
1.4 Known positional m olecules in the developing retina 66
16
A C K N O W L E D G E M E N T S
Firstly, m any thanks to my supervisor, Prof. Paul Brickell, for all his support,
guidance and advice during these three years.
I am grateful also to Prof. Cheryll Tickle of A natom y who not only allowed me
to w ork in her department but also provided wise counsel. Prof. David Latchmann
(M olecular Pathology) and Dr. Benny Chain (Imm unology) are also offered thanks for
kindly allow ing m e to work in their departments during the course of my PhD.
Thanks go out to all the researchers w ho have so kindly contributed reagents,
techniques and technical advice to this thesis. In particular, I would like to thank Dr.
Philippa Francis-W est for all her help with m any diverse aspects of my project and for
her vision and skill. I would also like to acknowledge Dr. Esther Bell, M iss Anne
Cizmeci (photographic techniques). Dr. Jim Cohen (immunocytochemistry), Dr. David
Darling, Dr. Peter M obbs (retinal cell culture). Dr. H erm ann Rohrer, Dr. Eduardo
Seleiro and Dr. Scott Stadler (GH6 DNA).
A s alw ays, thanks to all my friends in various departments of U C L for good
advice and camaraderie; Esther T., Amanda, Anne, Shanie and Nat plus others too
num erous to m ention.
M ost o f all, my deepest gratitude to friends not directly in the field w ho have
had to put up with me throughout these three years. My special thanks to my dearest
friend. M iss M argaret Marchetti, w hose loving support extended to listening to my
ranting diatribes on obscure embryological problem s in num erous London cafes. In a
sim ilar vein, I am grateful to Miss Sophia Prevezanou, M iss Neo Soek Y ing, Dr. H o
Kwok Ki and M iss Lim Meei Shan; thanks all for your patience and understanding - it
was m uch appreciated.
Last and best o f all, I would like to thank my parents for their love, patience and
w isdom. Y ou w ere always with me in spirit if not physically and I couldn't have done
17
For my parents in loving dedication
"All xue can do in life is try,
CHAPTER 1
I N T R O D U C T I O N
- ■ C ^ k ¥ m ^ \ ‘ W m O D Ü G TlO N AND AIMS . ' 19
Over the past few years, great strides have been made in identifying the many
signalling factors that regulate cell-cell interactions in embryogenesis. Among the many
large families o f grow th factor molecules that have been extensively researched is the
Transform ing G row th Factor-13 (TG F-6) superfamily which includes the Bone
M orphogenetic Proteins (BMPs).
These secreted factors have been implicated as regulators of m orphogenesis in
diverse em bryonic systems ranging from mouse skeletal element formation (Kingsley et
al, 1992; K ingsley, 1994a) to epiderm is formation in X enopus gastrula (W ilson and
H em m ati-Brivanlou, 1995) to chick limb patterning (Francis et al, 1994).
This thesis examines the role of one member of the BMP subgroup, B M P-4, in
the development o f the chick eye. I sought to do this by firstly investigating the
distribution of B m p-4 RNA in the developing eye and then looking at the effects of
BM P-4 on retinal cells in vitro and in vivo.
1.1 T he Bone M orphogenetic Proteins
The Bone M orphogenetic Proteins are a group o f at least ten signalling proteins
some o f w hich w ere originally identified by their ability to induce ectopic bone
form ation (Urist, 1965). Because of this functional approach to their identification, one
m em ber of this group is structurally unrelated to the others; BMP-1 has been identified
as Procollagen C-Proteinase (K essler et al, 1996; Reddi, 1996) whereas the other
BM Ps are members of the TG F-6 superfamily and share a closely-related carboxy-
m k Y m K l -i n t r o d u c t i o n AND AIMS X 20
1.1.1 HISTORY AND ROLE IN BONE FORMATION
The majority of embryonic bone development takes place by the process of
endochondral bone formation which involves the laying dow n of a cartilage matrix
w hich then calcifies. This process can be mimicked by the implantation of
demineralised bovine bone extract in sites that would not normally give rise to bone,
such as rabbit intram uscular pouches (Urist, 1965).
Urist (1965) believed that a proteinaceous agent in the extract was responsible
for its bone-form ing activity and named this agent bone morphogenetic protein (B M P ).
At this time, it w as still unclear as to w hether BMP ability w as due to one or several
interacting com ponents of the extract (W ozney et al, 1988).
The process o f bone formation as mediated by BM Ps takes place as follows. By
day 3 after implantation of the extract, invasion by surrounding mesenchymal cells
takes place. The mesenchymal cells differentiate into chondroblasts and chondrocytes
producing a m ass o f cartilage (a condensation) w hich undergoes proliferation,
hypertrophy and finally calcification (Carrington and Reddi, 1991). At day 10, the
cartilage matrix begins to break down due to the activity of osteoblasts, derived from
mesenchymal cells, and osteoclasts, derived from bone m arrow haematopoietic stem
cells, w hich go on to form bone on the remnants of the cartilage matrix. At the same
time, the m arrow o f the developing bone becomes filled with blood vessels and
haematopoietic cells. The entire process o f ossicle formation is complete by day
21 (Zheng et al, 1994).
Largely because this bioassay of ossification remained the defining criteria for
BM P activity, separation of this activity from the rest of the extract remained
problem atic W ang and coworkers succeeded in extracting BM Ps in guanidine (W ang et
al, 1990). The initial cloning of the first three members of the BMP subgroup, BM Ps -
2, -3 and -4, (W ozney et al, 1988) led shortly to the cloning of another three B M Ps,
BM P-5, -6 and -7 (Celeste et a l , 1990). BM P-8 or Osteogenic Protein-2 w as later
. r - C H A P T E R 1 - INTRODirCTlOMAm AIMS'': ; ; , / - 2 /
discovered, BM P-9 and BM P-10, has not yet been published, but patents have been
applied for in 1993 and 1994 respectively.
The BMP activities of BM P-1, -2 and -3 were demonstrated by expressing the
proteins in COS cells, showing that implanted cells resulted in cartilage formation
(Luyten et al, 1989). Similarly, Wang et at (1990) show ed that recombinant BM P-2
expressed in Chinese hamster ovary cells was capable of inducing cartilage and bone
form ation. Secreted BMP-4 has also been shown to have osteogenic activity
(H am m onds et al, 1991) as has BMP-5 in cell lines(Baylink et al, 1996).
Another BMP that has been shown to have bone-inducing potential is BM P-7
w hich has been shown to induce differentiation of chondroblasts and osteoblasts in
m urine clonal cell lines (Asahina et al, 1996). The other BM Ps have not been show n to
induce ectopic bone or cartilage, but several are thought to have various functions as
signalling molecules in morphogenesis (see later).
1.1.2 FAM ILIES W ITH IN THE TG F-p SUPERFAM ILY-AN OVERVIEW
All the BM Ps, except BM P-1, are members o f the Transform ing G row th
Factor-p superfamily. This is a large group of molecules (at least 36 members at tim e of
w riting), which are peptide growth factors capable o f regulating m orphogenesis at
m any stages o f embryonic development. Members o f the TG F-P superfamily have
been divided into distinct subgroups according to the sequence homology o f their
mature C-terminal domains (see Table 1.1). The major categories consist of : 1) The
TG F-p subfam ily 2) The Activin subfamily 3) The large dpp-Vg-related (DVR) group
Members of the TGF-jg superfamily
GROUP
FAMILY
SUBFAMILY
MEMBERS
TGF TGF 1.2,3,4 & 5
Activin Inhibin /3A, B
DVR
<
BMP/OP
<
dpp
60A
Radar
BMP-2, BMP-4 & dpp
BMP 5-8. 60A, UNIVIN. OP-3
Radar, GDF 5-7
V g l, dsl. BMP-9
BMP-3, GDF1, GDF3, nodal, screw, ADMP, BMP-10, GDF-10
Inhibin a, MIS, GDNF
T a b le 1 .1 The organisation of the TG F-6 superfamily into families, subfam ilies and
groups according to sequence identity. M ost o f the groupings are taken from K ingsley
(1994), while the organization o f the B M P /O P family is from Griffith (1996), Radar
1.1.2.1 The T G F-P subfamily
The first discovered m em ber o f this family, T G F -p i, w as isolated from human
platelets and placenta, and bovine kidney as a 25kD homodimer with a unique N-
terminal sequence. Other closely related isoforms have been found in vertebrates,
namely T G F -(32, p3, p4 and p 5 (reviewed in Sporn and Roberts, 1992; Mosesgf al,
1990). TG F-p4 is thought to be the chick homologue o f m am m alian T G F -pi (Burt and
Paton, 1992).
TG F-Ps have been show n to regulate multiple functions in animal cells,
including cell cycle progression, differentiation, adhesion, migration and extracellular
matrix production (reviewed in Sporn and Roberts, 1992). T G F -p i null mutant mice
suffer defects in yolk sac haematopoiesis and endothelial differentiation w ith those
surviving past birth dying a few weeks later from an inflammatory syndrome (Dickson
et al, 1995; Kulkarni et al, 1993).
1.1.2.2 The Activin subfamilv
A ctivin protein is fromed either by disulphide-linked homodimers of Inhibin pA
or pB subunits or by a heterodim er composed of pA and p B subunit proteins (Mitrani
et al, 1990). The pA and pB subunits can also form dimers with a more distantly
related subunit called Inhibin a (Kingsley, 1994).
The activins are thought to regulate the formation of axial structures in the chick
(M itrani et al, 1990) and mesoderm induction and neurulation in X en opus (N ishim atsu
et al, 1992a) but not in mice (M atzuk et al, 1995).
1.1.2.3 The DVR group
The DVR group is the largest subdivision o f the TG F-P superfamily and
P-3, G D F-1, GDF-P-3, etc (see Table 1.1). Kingsley (1994) has pointed out that the
grouping is supported by percentage similarity scores but that the borders of the group
are som ew hat blurry.
The BMP family itself can be further broken dow n into three subfamilies, the
dpp fam ily, the 60A family and the newly discovered radar family. Each of these will
be discussed in turn. Members within each subfamily share 12-92% identity, while
m em bers of different subfamilies share only 40-60% identity (Storm et al, 1994).
The dpp subfamily is named after the Drosophila decapentaplegic (dpp ) gene
w hich w as originally identified as a series o f allelic mutations affecting imaginai disc
developm ent (Spencer et al, 1982). This gene regulates several aspects o f Drosophila
developm ent and is discussed in greater detail in section 1.2.3.2.
BM P-2 and BMP-4 are vertebrate homologues o f dpp and are 75% identical to
the D rosophila gene. The fly and vertebrate proteins are able to substitute for each other
on a functional level. A chimeric dpp-Bmp-4 transgene w as able to rescue dorso-ventral
patterning of null dpp mutants flies (Padgett et al, 1993), while dpp and the related
protein 60A (see later) can induce bone formation in rats (Sam path et al, 1993).
Like the dpp subfam ily, the 60A subfamily consists o f one Drosophila gene
and a num ber of vertebrate BM Ps. The original member, 60A, w as isolated as part o f a
search for TG F-p family m em bers in the fruit fly genome (D octor et al, 1992; W harton
et al, 1991). 60A protein is cleaved and then secreted as an N-terminal domain and C-
term inal homodimers and may play a role in mesoderm form ation and gut development
(Doctor et al, 1992). Bm p-5, -6, -7 and -8 are members o f this subfamily and show
approximately 75% homology between themselves and approximately 60% hom ology
to 60A. The recently discovered m olecules Univin (in sea urchins; Stenzel, 1994) and
OP-3 (Griffith et al, 1996) also belong to this subfamily.
It has been suggested that members of the 60A subfam ily may exert their effects
' ; ' :/;v. > .ICHAPTER 1 y im m m C T tO N AN D AIM S _ x'. ' " /: n
heterodimers with potent bone-inducing capacity (Aono et al, 1995) and B m p -2 and
B m p -7 h?Lve overlapping expression domains in vivo (Francis-W est et al, 1995; Lyons
et al, 1995). Furtherm ore, BM P-2, -4 and -7 appear to bind to common cell surface
receptors (see section 1.1.5).
The third family, radar, was named after the recently discovered zebrafish gene
product of the same name and contains Radar and G rowth and dijferenatiation factors
5-7 (G d f 5-7). Radar is most closely related to G df-6 and is expressed in the dorsal
region of the neural plate and retina. It is also found in neural crest and ventral posterior
m esenchym e (Rissi et al, 1995). Mutations in the G df-5 gene cause a condition called
brachypodism in mice and in hum ans, characterised mainly by shortening o f limb
skeletal elements due to faulty chondrogenesis (Storm et al, 1994; Thom as et al, 1996).
Apart from these three subfamilies, the BMP/OP family also contains the V g-1,
dorsalin (dsl) and Bmp-9 genes. The Vg-1 gene w as originally isolated due to its
asym m etric distribution in oocytes (W eeks and M elton, 1987) and is thought
to be involved in mesoderm and axis formation in the oocytes (Thom sen and M elton,
1993)D s l has been implicated in the regulation of cell differentiation in the neural tube
(Basler et al, 1993) while B m p-9 is thought to be involved in liver m orphogenesis
(Song et al, 1995).
A number of diverse molecules lie within the DVR grouping but outside the
BM P fam ily. This group is more heterogenous than the others discussed so far. B m p
-3, for exam ple, bears only 40-48% homology to the other BM Ps. Gdf-1 w as isolated
during a screen for novel (mammalian) members of the T G F-P subfam ily. The gene
has been show n to comprise two non-overlapping open reading frames encoding two
different proteins. One of these was detected exclusively in brain, spinal cord and
peripheral neurons in mouse embryos. The other is highly conserved between mice and
Another GDF, GDF-3, lacks a cysteine residue important for the process of
dimer formation in other members o f the T G F -p superfam ily. It is expressed in
developing skeletal tissue (Jones et al, 1992b) and in adult thymus, spleen, bone
m arrow and adipose tissue.
N od al has been localised to the node at the anterior portion o f the primitive
streak in mouse embryos and is thought to encode a signalling molecule essential for
m esoderm formation and organisation of axial structure in early mouse development
(Zhou et al, 1993).
Unlike the other genes in this group, screw (sew) is a D rosophila gene thought
to interact with dpp in specifying dorsal cell fates in the developing em bryo. Like sew ,
Anti-Dorsalising-M orphogenetic-Protein (ADMP) is thought to be involved in setting
up em bryonic polarity, but as its name implies, overexpression of admp m R N A in
X enopu s suppresses dorsal and anterior structures. Dorsalising factors like activin and
LiCl induce A d m p gene expression suggesting that the gene may act to restrict the
dorsalising activity of activin(M oos et al, 1995).
G df-10 is expressed in developing murine uterus, adipose tissue, brain, liver
and spleen, suggesting that it has multiple roles in em bryogenesis (Cunningham et al,
1995).
1.1.2.4 Other more divergent genes
The last group of molecules I will discuss are divergent molecules that do not
fall into any particular group within the TG F-p s u p e r f a m i l y . / « / i fa has already been
mentioned. Mullerian inhibiting substance (M IS) is a gonadal hormone and M IS
deficient male mice develop as pseudohermaphrodites w ith oviducts and uteri. Four
receptors for MIS have been cloned (He et al, 1993). A nother molecule, glial-derived-
neurotrophic factor (GDNF) was isolated due to its ability to promote the survival and
X^fAPTBR 1 > um O D Ü C T lO N AND AIMS f : : : :/ 27
1.1.3 STR U C T U R E OF THE BM PS
A s already mentioned, BM Ps are members of the TGF-B superfamily. With the
exception of GDF-3 which has six cysteine residues, the T G F-p C-terminal domain has
seven conserved cysteines. The members of this family are initially synthesised as
large precursor proteins containing an amino terminal signal sequence and a pro
dom ain. This pre-pro precursor is cleaved at a dibasic or RX X R site to release a mature
C -term inal peptide of 110-140 am ino acids (Fig. 1.1 A). Proteolytic cleavage of the pro
dom ain and dimer formation are required for activity (Dale et al, 1993; Thom sen and
M elton, 1994) and may be an important method o f regulation o f this class of
m olecules. The BM Ps are capable of both homodimer and heterodimer formation
(A ono et al, 1995). The hom odim eric form o f X enopus B M P-2 has been purified from
X e n o p u s em bryos and shown to induce alkaline phosphatase expression in osteoblast
cells (Shoda et al, 1994), while homodimers of X en o p u s BM P-4 and BM P-7, as well
as heterodim ers o f BM P-4/-7 have osteoinductive capabilities in rats, with the
heterodim er having the strongest activity (Aono et al, 1995). Aono et al (1995) also
suggested that native bone-inducing activity in bony matrix w as due to heterodimeric
B M P.
The N-terminal region (leader and pro-domain) o f TGF-6 family members
exhibits a great am ount of variability (Kingsley, 1994) and can be disrupted in at least
one family m em ber, D SL, w ithout affecting biological activity (Basler et al, 1993).
Even w ithin the B M Ps, it is the C-terminal amino acid region that is most highly
conserved (Feng et al, 1994; Rosen and Thies, 1992). H ow ever, the pro-domain is
thought to be necessary for normal synthesis and secretion (Gray and M ason, 1990;
H am m onds et al, 1991; Thom sen and Melton, 1993). The im portance of the pro-region
is illustrated by the fact that creation o f a fusion protein containing the pro-region of
BM P-2 fused to the mature region of BMP-4 dramatically improved secretion of BMP-
ICHAFTERI '"'V'r2<5
The conserved cysteines in the mature protein play an important role in its
tertiary structure - the creation of the cystine or TGF-13 knot (Daopin et al, 1992;
Schlunegger and Grutter, 1992). This is an eight-member ring at the core of the
molecule formed by two disulphide bridges connecting four cysteine residues, w ith a
third disulphide bridge pointing directly through the ring (see Fig. I .I B ) . Finally, the
last cysteine residue in each m onom er subunit is necessary for the formation of
disulphide bonds between biologically active homodimers or heterodimers (M assague,
1990). Tw o TG F-6 family m em bers, GDF-3 and G D F-9 (Jones et al, 1992b;
M cPherron and Lee, 1993), lack this last cysteine but may be able to dimerise by virtue
of their hydrophobic contacts.
The three-dimensional structure of BM P-7 has been reported (Fig. I .I B ;
G riffith et al, 1996) and, like TG F-6, possesses the cystine motif and the beta-folds,
suggesting a common mechanism for receptor interaction.
It has been suggested that the protease activity of BMP-1 may have a role in the
activation o f the other BMPs, since its sequence is closely related to that of Drosophila
tolloid w hich has been implicated in the processing pathway of DPP, the Drosophila
ortholog o f BM P-2 and -4 (Childs and O 'C onnor, 1994; Finelli et al, 1994; Shimell et
al, 1991). This prospect seems less likely now that BM P-1 has been identified as
procollagen C-proteinase and is thought to play a more direct role in ossification
(K essler et al, 1996; Reddi, 1996). However, the BMP-1 protease may have m ore than
one function. Also, the Bm p-1 gene encodes two alternatively spliced transcripts, not
only for the BMP-1 pro-collagen protease, but also for mammalian Tolloid (which
encodes a putative astacin metalloendopeptidase domain). It has been suggested that
BM P-1 belongs to a separate class of structurally-related proteins containing CUB
: / -:v;=.<CHAPXER' 1 - m R O D U C T lO N A m AIMS = %.y:,.
Fig. 1.1 Structure o f TGF-B superfam ily members. (A) D iagram m atic view of TGF-B
related protein (from Kingsley, 1994). All members of the family are synthesised as
larger precursor molecules. The leader sequence targets the precursor to the secretory
pathway; while the variable pro-domain may assist in folding, synthesis or secretion.
The signalling molecule consists of homo- or hetero-dimers o f a mature w ell-conserved
C-terminal domain which is cleaved from the rest of the molecule at a RX XR site. The
mature fragment contains seven invariant cysteines. (B) Schematic draw ing o f the 3-D
structure of BM P-7 (from Griffith et al, 1996). Six cysteines form three disulphide
bonds characteristic of the cystine knot motif. Two at C ys-67-Cys-136 and C ys-71-
C ys-138, form a ring through which the third, Cys-38-Cys-104, passes. The four
strands of the antiparallel B-sheet, which emanate from the knot, form two finger-like
projections. The heel of the hand is formed by an a-h elix at the opposite end o f the
knot. The tube marked a l is the a helix while the solid thin lines indicate the
intrasubunit disulphide bonds. The N -term inus lacks the disulphide bond characteristic
CHAPTER \^lN T m D U C T J O N AND% ÎM S
m
Leader
(15-25 @a)
Pro-do main
(50-75 aa)
c c c c c c c
Mature domain
(110-140 aa)
H eel
Finger
1.1.4 BM PS IN BONE FORMATION
The initial discovery of the BM Ps w as due to their ability to induce the
form ation of bone when implanted in ectopic sites in the rat (see section 1.1.1) and their
effects on bone morphogenesis continue to be the subject of m uch research.
BMP-1 has recently been shown to be identical to Procollagen C-proteinase and
cleaves the C-propeptides of procollagen I, II and III to produce the collagen fibres of
the extracellular matrix (Kessler et al, 1996).
B m p-2 expression is localised to the precondensing mesenchyme o f the
developing mouse limb (Lyons et al, 1990) and both B m p -2 and -4 are found in the
developing cartilage elements of the chick limb (Francis et al, 1994), suggesting a role
for these BM Ps in embryonic skeletogenesis. B m p -2 and-4 have also been implicated
in orofacial primordia development (Bennett e ta l, 1995; Francis-W est et al, 1994).
B m p-3 is expressed in developing human perichondrium , periosteum and
osteoblasts and has been show n to have cartilage-inducing activity when expressed in
COS cells (Luyten et al, 1989; Vukicevic et al, 1994).
Mice carrying the short-ear {se) mutation have deletions in the B m p -5 gene
(Kingsley et al, 1992; Kingsley, 1994a). The m utant m ice suffer from a num ber o f soft
tissue alterations and skeletal defects. Kingsley (1994a) traced these defects to the early
stages of cartilage formation based on the alteration in the size and shape o f cartilage
condensations in the mutant mice. H ow ever, B m p -5 is also expressed later in
osteogenesis in the osteogenic stem cell layer surrounding the cartilage matrix
(K ingsley, 1994a) and Baylink et al (1996) have proposed that the defects in se mice
are due to impaired osteoblast differentiation.
The B m p -6 gene is expressed in hypertrophic cartilage in developing m ouse
em bryos(Lyons et al, 1989) and together with Bm p-1, 2 and 4 is expressed by cultures
o f foetal rat calvarial osteoblasts as they form mineralised bone nodules (H arris et al,
Q ^ \ Y I E K l B rm O D V C T IO N AN D A IM S
-B m p -7 m R N A is expressed at multiple sites in the m ouse embryo and is often
colocalised w ith Bm p-2, suggesting that the two BM Ps function co-operatively (Lyons
et al, 1995), perhaps in heterodimers. Mice deficient in B m p -7 have skeletal defects in
the skull, rib cage and hindlimbs (Luo et al, 1995; Dudley et al, 1996), show ing that
B m p -7 is required for proper skeletal developm ent. In addition, BM P-7 induces
chondroblastic and osteoblastic differentiation in murine clonal cell lines (Asahina et al,
1996).
BM P-8, 9 and 10 have not yet been show n to induce bone or cartilage
formation in vivo and their localisation in developing skeletal structures has not been
reported.
W hat is currently known about the osteoinductive properties of the BM Ps is
sum m arised in Table 1.1.
The exact mechanism of endochondral ossification induced by the B M Ps is yet
unknow n, but a number of clues to the pathway exist (for review, see R osen and
Thies, 1992).
The BM Ps are secreted signalling molecules and are thought to act in an
autocrine or paracrine manner to regulate skeletal development in the embryo (Chen et
al, 1995). Recombinant human BM P-2 has been demonstrated to enhance cell
proliferation (Yamaguchi et al, 1991) and stimulate gene expression of interleukin-6
(IL-6) and TG F-B l in a human osteoblast-like cell line (Zheng et al, 19 9 4 ). TG F-61
production by monocytes in response to recombinant human BM P-4 is thought to be
instrumental in the differentiation of mesenchymal cells into chondrocytes
(Cunningham et al, 1992); while IL-6 stimulates osteoclast maturation (K urihara, 1990)
and bone resorption (Lowick et al, 1989).
Recently, osteoblast treatment with BM P-2 has been shown to shift TGF-13
action away from DNA synthesis and towards stimulation o f collagen synthesis by
altering TGF-B binding to its receptors (Centrella et al, 19 9 5 ). TG F-Bl and BM P-2
have also been shown to stimulate chondrogenesis in a chondroblast cell line by altering
I ' ' / / - --y ; i Q n h Y ï m X - m R O D Ü C T l O H Â m AIM S
BM P g » n a I n d u e * * b o n e / c a r ti la g a '’ E x p r e s s io n in d e v e lo p in g s k e le ta l s tr u c t u r e s E ffects of k n o c k o u t o n b o n e fo rm a tio n BM P-1 C a r t ila g e
B M P -2 B o th
B M P-3
B M P - 4
B M P-5
BMP-6
B o th B o th
B o th
N at rep o rted
BMP-8 Not re p o rte d BMP-9 Not rep o rted BMP-10 Not rep o rte d
C o n d e n s in g m e s e n c h y m e C o n d e n s in g p r e c a r til a g in o u s m e s e n c h y m e , lim b a p ic a l e c to d e r m a l rid g e . In te rd ig its, o r o f a c ia l p r im o r d ia a n d to o t h b u d s . L a ter in p e r ic h o n d r iu m a n d p e r i o s t e u m
P e r ic h o n d r iu m , p e r i o s t e u m , o s t e o b l a s t s A p ica l e c to d e r m a l rid g e a n d
m e s e n c h y m e of early lim b b u d s . A lso In o ro fa c ia l p r im o r d ia
C o e x te n s iv e w ith m a n y r e g i o n s of c o n d e n s in g m e s e n c h y m e . L a ter fo u n d in p e r ic h o n d r iu m a n d p e r io s te u m o f d iffe re n tia tin g sk e le ta l e le m e n ts
H y p e r t r o p h i c c a r t i l a g e a n d
o s t a o b l a s t s
C o n d e n s i n g p r e c a r t i l a g i n o u s
m e s e n c h y m e , i n t e r d i g i t s . L a t e r
in p e r i c h o n d r i u m .
Not rep o rted Not rep o rted Not rep o rted
N ot re p o rte d N ot re p o rte d
N ot r e p o r te d No sk ele tal- sp e c ific d e f e c ts
Sm all ex tern al ea r. s c o llo p in g of th y ro id c a rtila g e , th in n in g of tr a c h e a rings. S h o rt w id e sk u ll, v e r te b r a l p r o c e s s e s r e d u c e d o r a b s e n t, lo n g b o n e s h a v e a lte r e d c u r v e s but s m a lle r w id th s
Not reported
Hind limb polydactyly. failure of seventh pair of ribs to fuse to sternum , cranial m e m b ra n o u s b o n e s not fully developed
Not reported Not reported
Not reported
T a b le 1 .2 Table summarising the role o f the Bone Morphogenetic Proteins in
vertebrate bone formation. Their ability to induce ectopic formation of bone and/or
cartilage, the pattern of expression of their transcripts in developing skeletal tissue and
I--- ' 1 - INTRODVCTION A m ÂÎMS
bone formation BM P-2 has also been show n to stimulate the in vitro migration of
osteoblasts and osteosarcoma cells and might therefore play a role in chemotactic
recruitm ent of osteoblasts during bone formation (Lind et al, 1996).
In addition, the Hox, P ax and Gli genes have been implicated in skeletal
developm ent and BM Ps have been show n to induce sequential H ox and M sx gene
expression in the process of ectopic bone formation (limura et al, 1994). If a parallel
may be drawn to related members of the TG F-6 family, TG F-6 itself has been show n
to induce the expression of a novel zinc-finger protein which is restricted to osteoblasts
and skeletal muscle cells (Subramaniam et al, 1995) and may mediate downstream
signalling in bone formation.
The BM Ps are of great interest because of their potential clinical applications in
stimulating bone healing (Rosen and Thies, 1992), how ever, much remains to be
discovered about the complex molecular cascades that accompanies endochondral
ossification.
1.1.5 BM PS AS SIG N A LLIN G M OLECULES IN DEVELOPMENT
A part from their function in the formation o f the skeleton, the BM Ps have been
im plicated in numerous other developmental processes.
Table 1.3 summarises the known m R N A expression patterns, effects o f m ouse
knockout/dom inant negative experim ents, naturally-occurring mutants and postulated
roles o f B M Ps in em bryogenesis outside of endochondral ossification.
1.1.5.1 BMP-1
BM P-1, the only non-TG F-6 related member of the family, may play a role in
axis formation in the em bryo. Transcripts o f the murine gene are distributed at low
BMP g e n * E x p re s s io n p a tte rn in so ft ti s s u e Effects of m o u s e k n o ck o u ts P r o p o s e d r o le s B M P. I
B M P-2
B M P .3
8 M P -4
B M P-5
B M P-6
B M P-7
B M P-8
B M P-9
B M P-10
M e s o d e r m , d e v e lo p in g s p in a l c o r d a n d m id /tiin d b ra in
M y o g en ic la y er of h e a rt c u s h io n s , to o th b u d s , hair & w h is k e r follicles, s e n s o ry p la c o d e s , d e v e lo p in g ovary a n d te stis .
In lu n g ep ith eliu m , kidney tu b u le s a n d in testin a l m u c o s a
M e s o d e rm , e p id e rm a l e c to d e rm , d o r s a l n e u ra l tu b e , lateral s o m ite s . L a ter in h e a rt, w h isk er follicles, to o th b u d s , b ra n c h ia l a rc h e s , d ie n c e p h a lo n , h in d b ra in , ey e.
D ev e lo p in g lu n g m e se n c h y m e , u re te r, b la d d e r , in testin e, a n d te le n c e p h a lo n . A dult lu n g e n d liver.
D e v e lo p in g o o c y te s , m e n in g e s , h e a rt, p la c e n ta , n eu ral tu b e a n d sk in & g u t ep ith elia. A dult a n d fetal b rain .
D e v e lo p in g n o to c h o rd , s u r f a c e e c to d e r m , h e a rt, g u t, kidney, e y e a n d b ra n c h ia l a rc h e s.
P a tte r n n o t re p o rte d
D e v e lo p in g h e p a to c y te s
N o t r e p o r te d
N ot re p o rte d
N ot re p o rte d
N ot re p o rte d
A b n o rm al m e s o d e r m a n d yolk s a c b lo o d v e s s e l form ation. D is o rg a n isa tio n of s tr u c tu r e s p o s te rio r to th e heart.
I n c r e a s e d I n c id e n c e of lu n g c y s ts , liver g ra n u lo m a s , h y d ro tic k id n e y s & u reter, d is p la c e d o v a r ie s & ir re g u la r in te s tin a l folding
N ot re p o rte d
F ailu re of n e p h r o g e n e s is in th e d e v e lo p in g k id n e y , m icro p h th a lm ia or a n o lp h th a lm ia
N ot re p o rte d
N ot re p o rte d
N ot r e p o rte d
E m b ry o n ic ax is form ation
M e s o d e rm in d u c tio n , n e u ro n a l sp e c ific a tio n , e p ith e lia l- m e s e n c h y m a l in te ra c tio n s , cell fate d e te rm in a tio n
S ig n allin g m o le c u le s in lu n g , k id n e y a n d in testin a l m o r p h o g e n e s is
Call fate sp e c ific a tio n , e p ith e lia l- m e s e n c h y m a l in te ra c tio n s , a p o p to s is , s ig n a llin g m o le c u le s in axis fo rm a tio n , n e u ro tro p h ic factor, m e s o d e r m a n d a n d e p id e r m a l in d u c e r
M e d ia te s
e p ith e lia l- m e s e n c h y m a l in te ra c tio n s
N e rv o u s ti s s u e m o r p h o g e n e s is
S ig n a llin g m o le c u le in k id n e y a n d e y e d e v e lo p m e n t
U nknow n
R e g u la to ry ro le in h e p a tic g ro w th a n d fu n ctio n
U nknow n
Table 1.3 Table summarising the multiple roles of Bone M orphogenetic Proteins in
I
\
6%
4"/Cr(lIAPTEKa
vUMS;
_____
2ZÊÉ
in the floor plate o f the spinal cord and mid/hindbrain from about 9.5 days p.c. The
latter pattern o f expression is regulated by the transcription factor H N F-33 (Sasaki and
Hogan, 1994). H ow ever, the recent discovery o f th e identity o f B M P -1 as Procollagen
C-proteinase (K essler et al, 1996) has raised the possibility that BMP-1 has no role in
patterning at all.
1.1.5.2 B M P-2
B M P-2 expression has been detected in a myriad o f non-skeletal tissues
including the limb, heart, to o th buds, hair and w hisker follicles (Lyons et al, 1990),
sensory placodes (A nthony Graham, unpublished com munication) and ovary and testis
(Nishimatsu É?/a/, 1992).
In the developing limb-bud, there are three m ajor sources o f signalling
molecules responsible for dictating outgrow th and axis specification. The apical
ectoderm al ridge (A ER) is thought to sustain outgrow th o f the limb by maintaining a
zone o f undifferentiated, proliferating mesenchymal cells at the tip o f the bud; the
progress zone (reviewed in Tickle, 1995). FGF-4 has been implicated as a ridge signal
candidate m olecule that can maintain outgrow th following removal o f the A E R (C ohn
et al, 1995; N isw ander and M artin, 1993; V ogel and Tickle, 1993).However, the
expression of F g f-4 is detected only after considerable outgrow th has occured and
another FGF, F gf-8, has been show n to be expressed prior to limb outgrow th in the
m ouse and chick and can also substitute for the A ER in stimulating limb outgrow th
(M ahm ood et al, 1995). Interestingly, B M P-2 counteracts the effect o f FG Fs in limb
buds devoid o f ectoderm (N isw ander and M artin, 1992).
A group o f cells in the posterior mesenchyme, the polarising region, are
thought to pattern the anterior-posterior axis. Bm p-2 transcripts are found in posterior
limb mesenchyme and can be activated in anterior mesenchyme by the application o f
retinoic acid o r polarising region grafts which also induce mirror-image duplication
X -ÎN rR O D U C n O N AND AIMS:::
---recom binant human BM P-2 using a retroviral vector is able to induce ectopic F G F -4 in
the anterior ridge, followed by ectopic expression o f the H ox-D genes in the anterior
mesenchym e (Duprez et al, 1996). Partial digit duplications were also observed. This
may imply that BM P-2 has a role in maintenance o f the ridge, and possibly in
patterning the anterior-posterior axis.
B M P-2 transcripts in the heart are localised to the myocardium at the time of
formation o f the endocardial cushions (Lyons et al, 1990). The formation o f the
cushions is thought to be initiated by a signal from the myocardium (Potts and R unyon,
1989) and Lyons et al (1990) have postulated that this signal is BM P-2. Interestingly
enough, the closely related gene dpp has been show n to determine the com petency of
mesoderm al cells to develop into cardiac tissue (Frasch, 1995) implying that its
vertebrate counterparts may well be important morphogens in heart development.
BM P-2 is detected fairly late in tooth development and together w ith B M P-4, its
expression pattern is suggestive of a role in epithelial-mesenchymal interactions (Vainio
et al, 1993). It has been shown to induce odontoblast differentiation w hen applied
together w ith dentin and postulated to induce ameloblast differentiation as well (Vainio
et al, 1993).
In the process of hair follicle formation, BM P-2 is first detected in the epiderm al
placode and becom es restricted to non-dividing cells around the base of the hair shaft in
the mature follicle (Lyons et al, 1990). W hen the closely-related BM P-4 protein is
ectopically expressed in the outer root sheath, animals display progressive balding after
the first grow th cycle suggesting that the role of BM P-2 in normal follicle developm ent
is to term inate cell proliferation and turn on trichocytic markers (Blessing et al, 1993).
B M P-2 is able to induce the expression o f the neural cell adhesion molecules
N CA M and L I in neuroblastoma cells (Perides, 1994) and can induce a distinct set o f
neuropeptide m arkers in cultured sympathetic neurons (Fann and Patterson, 1994). The
neuropeptide genes induced change with membrane depolarisation possibly allowing
I : c h X p x e r i m rR O D U cnoH a n d a i m s
-BM P-2 has also been put forward as a mesoderm -inducing factor synthesised
by the extraembryonic endoderm (Rogers et al, 1992). F9 cells resemble cells o f the
blastocyst inner cell mass when undifferentiated and extraembryonic endoderm w hen
differentiated. When differentiation is induced by retinoic acid (RA), B m p-2 m RN A
levels increase and BMP-2 treatment o f such cells can alter growth and m orphology.
BM P-2 also represses its own expression in a negative feedback loop (Rogers et al,
1992). The action of RA on BM P may be mediated by the RAR-alpha and the RAR-
gam m a receptors (Rogers, 1996).
X enopus Bm p-2 transcripts are detected in the oocyte and blastula, but not after
gastrulation (Nishimatsu et al, 1992) and the gene, like BM P-4, may play a role in
m esoderm form ation in the X enopus embryo.
1.1.5.3 BMP-3
BM P-3 is expressed in a hum an lung carcinoma cell line (Wozney et al, 1988),
and has been detected in human foetal lung tissue . It has also been found in developing
human intestinal mucosa and kidney mesonephric tubules (Vukicevic et al, 1994). This
suggests that it may have a regulatory role in the m orphogenesis of hum an lung, kidney
and intestine.
1.1.5.4 BM P-4
B m p-4 transcripts are found in many of the same organs that contain B m p -2
transcripts, but usually the spatiotemporal localisation within the organ is ju st slightly
different. Outside o f the developing skeleton, B m p -4 has been found in the heart, the
w hisker follicles (Lyons et al, 1991), the teeth (Vainio et al, 1993), the branchial arches
(Francis-W est et al, 1994; Wall and Hogan, 1995), the diencephalon and hindbrain
(Lyons et al, 1991; Graham et al, 1994) and the eye (this thesis; also Francis-W est et