Development and Application of a Novel Peptide Nucleic Acid Probe for the Specific Detection of Helicobacter pylori in Gastric Biopsy Specimens
Full text
Figure
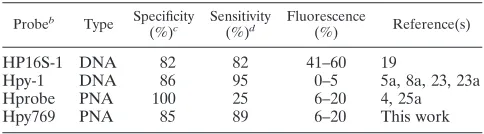
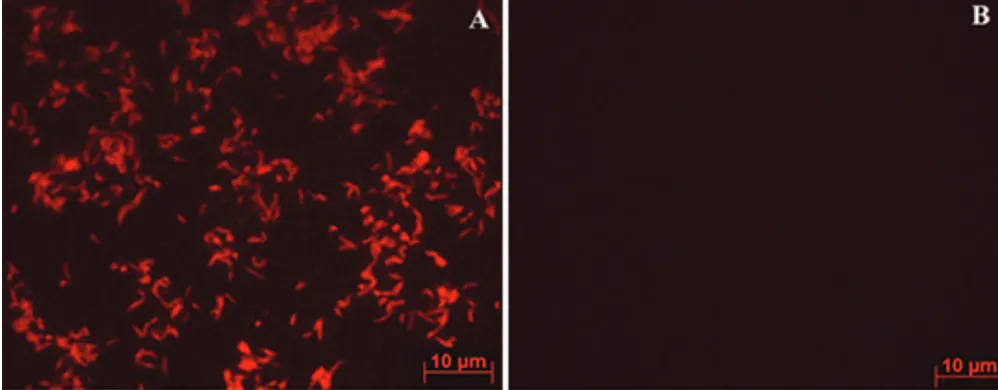


Related documents
On the other hand, there are many parameters which we have to decide.Those are the coefficients of the spline functions and the total number of knots, and positions of the
Bayesian Cross-Sectional Analysis of the Conditional Distribution of Earnings of Men in the United States, 1967-1996: Appendices.. Geweke, John and
The addition of continuous pulse oximetry surveillance to intermittent or continuous respiratory rate monitoring can be very helpful when detecting Type II RECC events unless
Conclusions: A realistic high protein weight-reducing diet was associated with greater fat loss and lower blood pressure when compared with a high carbohydrate, high fiber diet in
Medicinal plants extract of Cocculus pendulus , Malva neglecta , Rhazya stricta , Jaubertia aucheri, Corchorus depressus , Salvia bucharica, Microcephala
In this study, based on the Rep protein sequence, some gemycircularviruses from giant panda samples showed high sequence identity to the viruses detected in insects or feces from
diagnostic testing, vector-borne and zoonotic infectious agents were frequently identi fi ed, representing 17% (23 out of 133) of identi fi ed causes of encephalitis in our
or more abnormal diagnostic test results, including autonomic function testing, sensory nerve biopsy, or IENFD in skin biopsy, were required to establish the
