0095-1137/09/$08.00⫹0 doi:10.1128/JCM.01565-08
Copyright © 2009, American Society for Microbiology. All Rights Reserved.
Utilization of Microsatellite Polymorphism for Differentiating Herpes
Simplex Virus Type 1 Strains
䌤
†
C. Deback,
1,2* D. Boutolleau,
1,2C. Depienne,
3C. E. Luyt,
4P. Bonnafous,
1,2A. Gautheret-Dejean,
1,2,5I. Garrigue,
6and H. Agut
1,2UPMC Universite´ Paris 06, UPRES EA 2387, Paris F-75013, France1; Service de Virologie, Groupe Hospitalier Pitie´-Salpeˆtrie`re, AP-HP,
Paris F-75013, France2; De´partement de Ge´ne´tique et de Cytoge´ne´tique, Centre de Ge´ne´tique Mole´culaire et Chromosomique,
Groupe Hospitalier Pitie´-Salpeˆtrie`re, AP-HP, Paris F-75013, France3; Service de Re´animation Me´dicale, Institut de Cardiologie,
Groupe Hospitalier Pitie´-Salpeˆtrie`re, AP-HP, Paris F-75013, France4; Laboratoire de Microbiologie, UPRES EA 4065,
Universite´ Paris Descartes, Faculte´ des Sciences Pharmaceutiques et Biologiques, Paris F-75006 France5; and
Laboratoire de Virologie EA2968 and IFR66, Universite´ Bordeaux 2, Bordeaux F-33076, France6
Received 12 August 2008/Returned for modification 10 October 2008/Accepted 17 December 2008
The herpes simplex virus type 1 (HSV-1) genome is a linear double-stranded DNA of 152 kpb. It is divided into long and short regions of unique sequences termed ULand US, respectively, and these are flanked by
regions of inverted internal and terminal repeats. Microsatellites are short tandem repeats of 1- to 6-nucleotide motifs; they are often highly variable and polymorphic within the genome, which raises the question of whether they may be used as molecular markers for the precise differentiation of HSV-1 strains. In this study, 79 different microsatellites (mono-, di-, and trinucleotide repeats) in the HSV-1 complete genome were identified by in silico analysis. Among those microsatellites, 45 were found to be distributed in intergenic or noncoding inverted repeat regions, while 34 were in open reading frames. Length polymorphism analysis of the PCR products was used to investigate a set of 12 distinct HSV-1 strains and allowed the identification of 23 polymorphic and 6 monomorphic microsatellites, including two polymorphic trinucleotide repeats (CGT and GGA) within the UL46 and US4 genes, respectively. A multiplex PCR method that amplified 10 polymorphic microsatellites was then developed for the rapid and accurate genetic characterization of HSV-1 strains. Each HSV-1 strain was characterized by its own microsatellite haplotype, which proved to be stable over time in cell culture. This relevant innovative tool was successfully applied both to confirm the close relationship between sequential HSV-1 isolates collected from patients with multiple recurrent infections and to investigate putative nosocomial infections.
Herpes simplex virus type 1 (HSV-1) is a member of the subfamily Alphaherpesvirinae. The seroprevalence of HSV-1 infection increases progressively from childhood and is in-versely proportional to an individual’s socioeconomic back-ground (35). Primary HSV-1 infections in children are typically asymptomatic but can give rise to herpetic gingivostomatitis. After primary infection of the orofacial region, HSV-1 is trans-ported in a retrograde manner to the nuclei of the trigeminal sensory neurons through their axons, which innervate the in-fected area. HSV-1 then establishes a life-long latent infection in the nuclei of sensory neurons, where the genome lies in a nonreplicating chromatin-associated state. Recurrent HSV-1 lesions occur following the reactivation of latent HSV-1, ax-onal transport of the reactivated virus, and HSV-1 replication on the skin and mucous membranes. Recurrent infections typ-ically give rise to herpes labialis or may be responsible for more severe clinical manifestations, including keratitis, meningoen-cephalitis, bronchopneumonitis (22), chronic or disseminated infections in immunosuppressed patients, or eczema
herpeti-cum. Eczema herpeticum, or Kaposi-Juliusberg disease, is an uncommon herpes simplex virus superinfection that occurs in patients with atopic dermatitis. Additionally, HSV-1 accounts for about half of the new cases of genital herpes in developed countries (14).
The HSV-1 genome is a linear double-strand DNA molecule of 152,261 bp (strain 17, GenBank accession number X14112) that encodes at least 77 genes (24, 28). It is composed of two covalently linked regions of unique sequences, termed the unique long (UL) and unique short (US) sequences, which are
bracketed by inverted repeats. Moreover, HSV-1 USand UL
undergo inversions resulting from recombination events that yield four genomic isomers in equimolar accounts (17).
The differentiation of unrelated HSV-1 strains has been used to trace the spread of infection in cases of nosocomial outbreaks (1, 6, 16); pathogenesis investigations (31, 32); and epidemiological studies, which also suggest that HSV-1 strains among the same geographic or ethnic groups are evolutionarily more closely related (5, 37, 39). HSV-1 strains differ slightly in their DNA sequences, which are used as molecular markers of genomic polymorphism. The method generally used to differ-entiate HSV-1 strains is restriction fragment length polymor-phism (RFLP) analysis of total HSV-1 DNA (4, 7, 26, 36) or RFLP-PCR-based analysis (23, 49, 53). Determination of the nucleotide sequences of PCR-amplified DNA fragments en-compassing reiterated regions (33) is another approach cur-rently used to characterize the genomes of HSV-1 strains.
* Corresponding author. Mailing address: Service de Virologie, CERVI, Groupe Hospitalier Pitie´-Salpeˆtrie`re AP-HP, 47-83 Boule-vard de l’Hoˆpital, Paris 75013, France. Phone: 33 1 42 17 74 02. Fax: 33 1 42 17 74 11. E-mail: [email protected].
† Supplemental material for this article may be found at http://jcm .asm.org/.
䌤Published ahead of print on 24 December 2008.
533
on May 16, 2020 by guest
http://jcm.asm.org/
Differences in the sizes of specific fragments among strains are detected in a PCR-based assay and are attributed to a variation in copy numbers or in the nucleotide sequences of these hy-pervariable reiterations, which have been identified within the ICP34.5 gene (8), the “a” sequence (10), the US10 and US11 genes, the introns of the US1 and US12 genes (23, 47, 48), and the glycoprotein G (gG) and gI genes (27).
Microsatellites are short tandem repeats of 1 to 6 bp and are the most mutable DNA sequences found in eukaryote, pro-karyote, and some virus genomes, particularly in intergenic regions and introns (11, 43). While they are hypermutable, microsatellites have been used as molecular markers in numer-ous ecology, evolutionary genetics, and genome mapping stud-ies. Moreover, the instability of these microsatellites, which are mainly homopolymer nucleotide repeats, may improve frame-shift mutations that provide phenotypic changes and an adap-tive advantage in the evolution of some mutated viral strains (12). The identification and genetic mapping of highly poly-morphic microsatellite loci in human cytomegalovirus (HCMV) have been reported previously (9). Therefore, microsatellite polymorphism analysis proved to be a rapid and accurate tool for studying the epidemiology of HCMV infections (29, 51). To our knowledge, this strategy has never been applied to HSV-1 infections.
The aim of our study was to establish the map and charac-terize the short microsatellites within the full-length HSV-1 genome and to confirm the utility of microsatellites as precise molecular markers for the differentiation of HSV-1 strains.
MATERIALS AND METHODS
Cells and virus strains.Vero cells (African green monkey kidney cells) and human embryonic lung fibroblasts (MRC5 cells) were propagated in minimum essential medium (GIBCO, Cergy, France) supplemented with 3% fetal bovine
serum (GIBCO), amikacin (20g/ml; Bristol-Myers Squibb, Rueil-Malmaison,
France), vancomycin (20g/ml; Lilly, Suresnes, France), glutamine (1%;
Euro-bio, Courtaboeuf, France), and a nonessential amino acid solution (1%; Sigma, Saint-Quentin Fallavier, France), referred to hereafter as the culture medium.
A total of 21 isolates and one laboratory strain (strain KOS) were used in this study. The clinical isolates were obtained on Vero cells from oral, bronchoal-veolar lavage, or cutaneous specimens from unrelated patients submitted for the diagnosis of clinical herpes reactivations. Determination of the HSV type of each strain was performed by means of an immunofluorescence assay with specific monoclonal antibodies, as described previously (44).
Microsatellite identification.The MsatFinder program was used to search the HSV-1 whole-genome sequence data (strain 17, GenBank accession number X14112) for microsatellites (45). The output results were filtered to include only short microsatellites with at least four trinucleotide, five dinucleotide, or nine mononucleotide repeats units, as previously described by Davis et al. (9).
Microsatellite characterization by sequencing.DNA was extracted from HSV-1-infected cells exhibiting a full cytopathic effect by using a QIamp DNA blood kit (Qiagen, Courtaboeuf, France), according to the manufacturer’s instructions. We analyzed 29 microsatellites using 25 PCR primer pairs (Table 1). PCRs
containing a reaction mixture with 1⫻buffer (Invitrogen), 2 mM MgCl2, 0.2 mM
each deoxynucleoside triphosphate, 200 nM each forward and reverse primers,
and 0.2 U of proofreadingTaqpolymerase (Platinum, Invitrogen) were
per-formed in a Mastercycler ep thermal cycler (Eppendorf, France). The reaction conditions were 1 cycle of denaturation (2 min at 96°C), 30 cycles (30 s at 96°C, 60 s at 55°C, 60 s at 68°C), and a final elongation step (10 min at 68°C). Negative controls were included in each run, and the specificity of the PCR was evaluated by including in each PCR run DNA extracted from noninfected Vero and MRC5 cells. Sequencing was carried out on a 3100 ABI sequencer, and all reported sequences were determined twice in each direction. Before sequencing of the PCR products, they were purified by using a Microcon kit (Millipore).
Microsatellite characterization by length polymorphism analysis.Each PCR was performed as described above by using a forward primer that was labeled at
the 5⬘end with a fluorescent phosphoramidite dye (6-carboxyfluorescein). One
microliter of each PCR product was diluted with distilled water (1:300) and
prepared by adding 0.5l of an internal size standard (GeneScan-500 ROX or
GeneScan-600 ROX; Applied Biosystems) and highly purified deionized
form-amide (Applied Biosystems) for a final volume of 16l. One previously sized
control sample was amplified and run on each gel to ensure that the amplicons were sized correctly. All products were analyzed on 3730 ABI sequencer (Ap-plied Biosystems), according to the manufacturer’s instructions. Fragment sizing was performed by using GeneMapper software (version 4.0; Applied Biosys-tems).
Multiplex PCR.Ten primer pairs that were able to be multiplexed into the same PCR run and that were able to produce amplicons simultaneously analyz-able within a single gel lane were then selected (Tanalyz-able 1). Multiplex PCRs were performed in a Mastercycler ep thermal cycler (Eppendorf) with a PCR mixture
containing 0.7⫻buffer (Invitrogen); 4 mM MgCl2; 0.2 mM each deoxynucleoside
triphosphate; 240 nM primer M69; 400 nM primers M5 and M22; 500 nM primer
M23; 600 nM primers M30, M10, M41, M26, and M31/32; 1M primer M14; and
0.3 U of proofreadingTaqpolymerase (Platinum; Invitrogen). The reaction
conditions were 1 cycle of denaturation (2 min at 96°C), 30 cycles (30 s at 96°C, 60 s at 65°C, 60 s at 72°C), and a final elongation step (10 min at 72°C). The multiplex PCR products were prepared as described above. The specificity of the multiplex PCR was confirmed by comparing the amplicons obtained by multiplex PCR with those obtained by means of single PCRs whose products were sec-ondarily pooled for capillary electrophoresis. The sizes of the amplicons were determined by using an internal size standard (GeneScan-500 ROX; Applied Biosystems), as described above.
RESULTS
Short microsatellite distribution in the complete sequence of HSV-1 genome.The in silico analysis by use of the Msat-Finder algorithm revealed that the HSV-1 genome (strain 17, GenBank accession number X14112) contained at least 79 targeted short repeat sites (arbitrarily numbered HSV1-M1 to HSV1-M79), including 35 mononucleotide repeat units, 16 dinucleotide repeats, and 28 trinucleotide repeats (Fig. 1). As expected, they were mainly localized in noncoding sequences, including terminal repeat regions (TRL, TRS) and inverted
internal repeat regions (IRL, IRS) as well as intergenic regions,
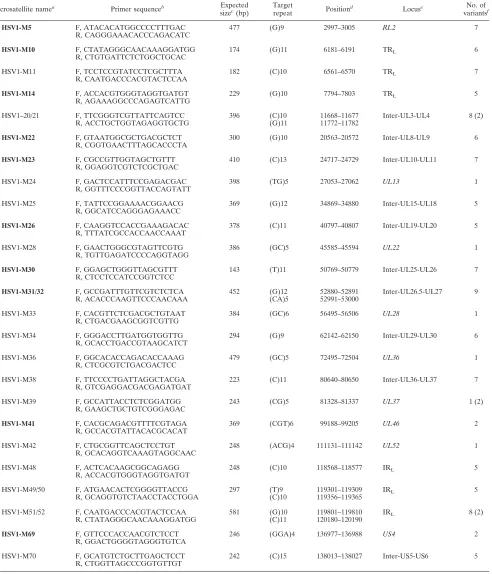
while 34 were in open reading frames (Fig. 1). The maximum number of repeat units of dinucleotide or trinucleotide motif was 6 [M33-(GC)6 and M41-(CGT)6], while the longest mono-nucleotide tract was 15 bases [M70-(C)15] (Table 2). Among these microsatellites, 29 were selected and 25 primer pairs were designed in order to synthesize amplicons containing these target repeats (Table 1).
Characterization of HSV-1 microsatellites. We first sur-veyed the 29 selected microsatellites in strain KOS and 11 HSV-1 clinical isolates (isolates A to K) using the 25 different PCR primer pairs separately (Table 1). The study of the cor-responding amplified fragments by means of capillary electro-phoresis showed that 19 of these loci were polymorphic, with microsatellites M24, M28, M33, M36, M39, and M42 being monomorphic among the panel of HSV-1 isolates tested. The polymorphic alleles are presented in Table S1 in the supple-mental material. At the same time, the variability of each microsatellite was confirmed by sequencing methods. As ex-pected, mononucleotide repeats were more variable, with 5 to 7 variants detected among the 12 strains tested. M21, M39, and M51 were considered impure microsatellites since they pre-sented polymorphisms in both their lengths and sequences. Polymorphic trinucleotide microsatellites were found within two protein-coding genes: M41(CGT)6, which encodes a stretch of aspartic acid residues within the UL46 gene, which encodes the tegument protein, and M69(GGA)4, which codes
on May 16, 2020 by guest
http://jcm.asm.org/
TABLE 1. Characteristics of amplicons containing HSV-1 microsatellite sequences
Microsatellite namea Primer sequenceb Expected
sizec(bp)
Target
repeat Position
d Locuse No. of
variantsf
HSV1-M5 F, ATACACATGGCCCCTTTGAC 477 (G)9 2997–3005 RL2 7
R, CAGGGAAACACCCAGACATC
HSV1-M10 F, CTATAGGGCAACAAAGGATGG 174 (G)11 6181–6191 TRL 6
R, CTGTGATTCTCTGGCTGCAC
HSV1-M11 F, TCCTCCGTATCCTCGCTTTA 182 (C)10 6561–6570 TRL 7
R, CAATGACCCACGTACTCCAA
HSV1-M14 F, ACCACGTGGGTAGGTGATGT 229 (G)10 7794–7803 TRL 5
R, AGAAAGGCCCAGAGTCATTG
HSV1–20/21 F, TTCGGGTCGTTATTCAGTCC 396 (C)10 11668–11677 Inter-UL3-UL4 8 (2)
R, ACCTGCTGGTAGAGGTGCTG (G)11 11772–11782
HSV1-M22 F, GTAATGGCGCTGACGCTCT 300 (G)10 20563–20572 Inter-UL8-UL9 6
R, CGGTGAACTTTAGCACCCTA
HSV1-M23 F, CGCCGTTGGTAGCTGTTT 410 (C)13 24717–24729 Inter-UL10-UL11 7
R, GGAGGTCGTCTCGCTGAC
HSV1-M24 F, GACTCCATTTCCGAGACGAC 398 (TG)5 27053–27062 UL13 1
R, GGTTTCCCGGTTACCAGTATT
HSV1-M25 F, TATTCCGGAAAACGGAACG 369 (G)12 34869–34880 Inter-UL15-UL18 5
R, GGCATCCAGGGAGAAACC
HSV1-M26 F, CAAGGTCCACCGAAAGACAC 378 (C)11 40797–40807 Inter-UL19-UL20 5 R, TTTATCGCCACCAACCAAAT
HSV1-M28 F, GAACTGGGCGTAGTTCGTG 386 (GC)5 45585–45594 UL22 1
R, TGTTGAGATCCCCAGGTAGG
HSV1-M30 F, GGAGCTGGGTTAGCGTTT 143 (T)11 50769–50779 Inter-UL25-UL26 7
R, CTCCTCCATCCGGTCTCC
HSV1-M31/32 F, GCCGATTTGTTCGTCTCTCA 452 (G)12 52880–52891 Inter-UL26.5-UL27 9
R, ACACCCAAGTTCCCAACAAA (CA)5 52991–53000
HSV1-M33 F, CACGTTCTCGACGCTGTAAT 384 (GC)6 56495–56506 UL28 1
R, CTGACGAAGCGGTCGTTG
HSV1-M34 F, GGGACCTTGATGGTGGTTG 294 (G)9 62142–62150 Inter-UL29-UL30 6
R, GCACCTGACCGTAAGCATCT
HSV1-M36 F, GGCACACCAGACACCAAAG 479 (GC)5 72495–72504 UL36 1
R, CTCGCGTCTGACGACTCC
HSV1-M38 F, TTCCCCTGATTAGGCTACGA 223 (C)11 80640–80650 Inter-UL36-UL37 7
R, GTCGAGGACGACGAGATGAT
HSV1-M39 F, GCCATTACCTCTCGGATGG 243 (CG)5 81328–81337 UL37 1 (2)
R, GAAGCTGCTGTCGGGAGAC
HSV1-M41 F, CACGCAGACGTTTTCGTAGA 369 (CGT)6 99188–99205 UL46 2
R, GCCACGTATTACACGCACAT
HSV1-M42 F, CTGCGGTTCAGCTCCTGT 248 (ACG)4 111131–111142 UL52 1
R, GCACAGGTCAAAGTAGGCAAC
HSV1-M48 F, ACTCACAAGCGGCAGAGG 248 (C)10 118568–118577 IRL 5
R, ACCACGTGGGTAGGTGATGT
HSV1-M49/50 F, ATGAACACTCGGGGTTACCG 297 (T)9 119301–119309 IRL 5
R, GCAGGTGTCTAACCTACCTGGA (C)10 119356–119365
HSV1-M51/52 F, CAATGACCCACGTACTCCAA 581 (G)10 119801–119810 IRL 8 (2)
R, CTATAGGGCAACAAAGGATGG (C)11 120180–120190
HSV1-M69 F, GTTCCCACCAACGTCTCCT 246 (GGA)4 136977–136988 US4 2
R, GGACTGGGGTAGGGTGTCA
HSV1-M70 F, GCATGTCTGCTTGAGCTCCT 242 (C)15 138013–138027 Inter-US5-US6 5
R, CTGGTTAGCCCGGTGTTGT
aThe 10 microsatellites in boldface are those that were selected for testing by multiplex PCR.
bF, forward; R, reverse.
cExpected length of the PCR fragment according to the annotation of the reference HSV-1 complete sequence (strain 17, GenBank accession number X14112).
dAccording the annotation of the same reference sequence.
eA gene name in italics indicates that the target repeat was located within an open reading frame, according to the annotation of the same reference sequence.
fNumber of variant alleles among the panel of the first 12 HSV-1 strains studied, as determined by both length analysis and nucleotide sequencing. M21, M39, and
M51 were considered impure microsatellites, since some strains presented both length and sequence polymorphisms. The numbers in parentheses indicate the number of sequence variants.
on May 16, 2020 by guest
http://jcm.asm.org/
for a polyglutamyl stretch within US4, which encodes virion gG (Table 1). At these two sites, two variant sequences were found among all strains tested in this study (Fig. 2). From this pre-liminary analysis, it was concluded that short microsatellites corresponding to mono-, di-, and trinucleotide repeats were highly polymorphic in the HSV-1 genome and can be
consid-ered potential molecular markers for the precise differentia-tion of HSV-1 strains.
[image:4.585.135.453.67.284.2]Microsatellite haplotypes obtained by multiplex assay for different HSV-1 strains.Ten primer pairs amplifying polymor-phic sites that could be multiplexed in the same PCR run were chosen in order to facilitate the rapid differentiation of HSV-1
FIG. 1. Linear map of genomic distribution of HSV-1 microsatellites. The linear double-stranded DNA is represented with the nucleotide scale at the bottom according to the sequence of strain 17 (GenBank accession number X14112). The unique ULand USsequences are shown as heavy solid lines, and the terminal and internal inverted repeats (TRL, IRL, IRS, and TRS) are shown as hatched boxes. The open reading frames on both the forward and the reverse strands are represented by open arrows. Each vertical line represents one microsatellite locus; the dotted lines and the dashed lines represent monomorphic and polymorphic microsatellites in the samples tested, respectively. This map is restricted to sites with at least four trinucleotide, five dinucleotide, or nine mononucleotide repeats units.
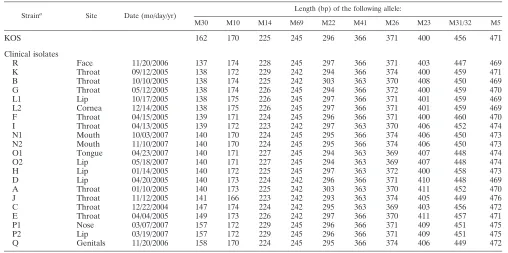
TABLE 2. Determination of microsatellite haplotypes for strain KOS and the 21 clinical isolates studied
Straina Site Date (mo/day/yr) Length (bp) of the following allele:
M30 M10 M14 M69 M22 M41 M26 M23 M31/32 M5
KOS 162 170 225 245 296 366 371 400 456 471
Clinical isolates
R Face 11/20/2006 137 174 228 245 297 366 371 403 447 469
K Throat 09/12/2005 138 172 229 242 294 366 374 400 459 471
B Throat 10/10/2005 138 174 225 242 303 363 370 408 450 469
G Throat 05/12/2005 138 174 226 245 294 366 372 400 459 470
L1 Lip 10/17/2005 138 175 226 245 297 366 371 401 459 469
L2 Cornea 12/14/2005 138 175 226 245 297 366 371 401 459 469
F Throat 04/15/2005 139 171 224 245 296 366 371 400 460 470
I Throat 04/13/2005 139 172 223 242 297 363 370 406 452 474
N1 Mouth 10/03/2007 140 170 224 245 295 366 374 406 450 473
N2 Mouth 11/10/2007 140 170 224 245 295 366 374 406 450 473
O1 Tongue 04/23/2007 140 171 227 245 294 363 369 407 448 474
O2 Lip 05/18/2007 140 171 227 245 294 363 369 407 448 474
H Lip 01/14/2005 140 172 225 245 297 363 372 400 458 473
D Lip 04/20/2005 140 173 224 242 296 366 371 410 448 469
A Throat 01/10/2005 140 173 225 242 303 363 370 411 452 470
J Throat 11/12/2005 141 166 223 242 293 363 374 405 449 476
C Throat 12/22/2004 147 174 224 242 295 363 369 403 456 472
E Throat 04/04/2005 149 173 226 242 297 366 370 411 457 471
P1 Nose 03/07/2007 157 172 229 245 296 366 371 409 451 475
P2 Lip 03/19/2007 157 172 229 245 296 366 371 409 451 475
Q Genitals 11/20/2006 158 170 224 245 295 366 374 406 449 472
aStrain pairs L1-L2, N1-N2, O1-O2, and P1-P2 were obtained from the same patient at two different times.
on May 16, 2020 by guest
http://jcm.asm.org/
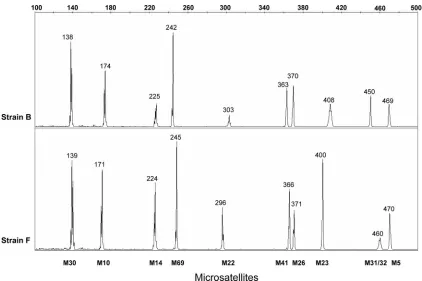
[image:4.585.39.547.464.717.2]strains by their microsatellite haplotypes (Table 1). In the present study, the haplotype was defined as the combination of the lengths of the 10 PCR products obtained from the multi-plex PCR assay and analyzed by polyacrylamide capillary elec-trophoresis (Fig. 3). The optimization of the multiplex reaction conditions led to the final definition of the technique described in Materials and Methods. The interassay reproducibility of
the multiplex PCR was assessed by amplifying sample A in 50 independent reactions, and no variation in the haplotype was observed. The microsatellite haplotypes of the 21 clinical iso-lates and strain KOS are presented in Table 2. As expected, the haplotypes corresponding to distinct HSV-1 isolates obtained from different individuals were all different. Considering the number of variants described for each of these 10
microsatel-FIG. 2. Sequences of two polymorphic trinucleotide microsatellites present in HSV-1 genes. (A) Microsatellite analysis of M41(CGT)6, which codes for a stretch of aspartic acid residues (residues 582 to 587) within the UL46 gene, showed two variants, M41(CGT)6 and M41(CGT)5, that led to alleles 366 and 363, respectively. The two variants represented 70% and 30% of the HSV-1 strains tested in this study, respectively. (B) Microsatellite analysis of M69(GGA)4, which codes for a polyglutamyl stretch (residues 79 to 82) within the US4 gene showed two variants, M69(GGA)4 and M69(GGA)5, that led to alleles 242 and 245, respectively. The two variants represented 42% and 58% of the HSV-1 strains tested in this study, respectively.
FIG. 3. Electropherograms of multiplex PCR products illustrating the haplotype characteristics used to differentiate the HSV-1 strains. The haplotype was defined as the combination of the lengths of 10 amplicons labeled at the 5⬘end with 6-carboxyfluorescein and containing HSV-1 microsatellite sequences obtained from the multiplex assay and analyzed by polyacrylamide capillary electrophoresis. The numbers 100 to 500 indicate the sizes (in base pairs) of the PCR amplicons. The haplotypes of strain B (top panel) and strain F (bottom panel) represented here display the length polymorphisms for the 10 loci.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:5.585.80.502.396.677.2]lites, it was computed that this strategy would be able, in theory, to identify more than 107different haplotypes.
There-fore, the multiplex assay was found to be simple, reproducible, and highly discriminative.
The stability of the microsatellites was investigated by prolonged serial propagation of HSV-1 in cell cultures. Strain KOS and four clinical isolates (isolates A, B, C, and F) were plaque purified three times and were then serially propagated for 15 passages on Vero cells. In order to take the impact of the cellular environment into account, we also used different cellular systems. For that purpose, two strains (strains KOS and A) were propagated in separate trials: on Vero cells alone, on human MRC5 fibroblasts alone, and under a third condition that used the repetitive rotation of Vero and MRC5 cells. At each of the 15 passages and in the contexts of the different host cells, the microsatellite haplo-types of the strains were analyzed. No variation was ob-served, with each of the five strains keeping its own haplo-type, depicted in Table 2, at each passage (data not shown). These data suggest that the HSV-1 microsatellites remained stable during viral replication. This led us to conclude that the multiplex PCR and length polymorphism analysis could be used as a rapid and precise tool for the identification of HSV-1 strains.
Differentiation of clinical HSV-1 isolates.The microsatellite haplotypes of two serial isolates from the same individual were compared (Table 2). Isolate L1 was isolated from a patient’s lip vesicles; isolate L2 was isolated 59 days later from the same patient’s herpetic keratitis lesions. Isolates N1 and N2 were isolated 39 days apart from a patient with a stem cell allograft who presented with chronic HSV-1 infection of the oral mu-cosa. Isolates O1 and O2 were isolated from the same individ-ual from two different herpetic recurrence lesions on the tongue and on the inferior lip, respectively. Isolates P1 and P2 came from a patient who developed herpetic vesicles 2 weeks apart, first on the skin around the nose and then on the lip. The microsatellite haplotype was identical in each case, even though the locations of the herpes lesions slightly differed over time. These results indicate that in these cases the strain hap-lotype remained stable in vivo through distinct or chronic her-petic episodes in the same individual.
Finally, strains Q and R were from two different patients attending the same care unit. The first patient, an immunosup-pressed 80-year-old woman, was hospitalized with a case of severe genital herpes due to HSV-1 that required intravenous acyclovir treatment. The second patient was hospitalized for acute eczematous lesions of facial atopic dermatitis and who developed a HSV-1 superinfection at exactly the same time. While this might have suggested the transmission of HSV-1 by hand carriage from the first patient to the second, the haplo-types of the two isolates were very different, thus indicating, on the contrary, that the infections originated from different viral strains (Table 2).
DISCUSSION
This study reports for the first time the presence of numer-ous microsatellite loci across the HSV-1 genome, found mostly, as expected, in noncoding regions and characterized by a high level of length polymorphism. Short microsatellite
poly-morphisms have already been used for the precise character-ization of HCMV strains (9, 29, 51), and a similar proposal has been made for human herpesvirus 6 strains regarding the spe-cific copy number of telomeric repeat sequences located at the ends of the genome (25). In comparison with reiterated regions in the US component still currently used for HSV-1 strain
differentiation (33), short microsatellites have the advantage of being distributed across all parts of the HSV-1 genome. The HSV-1 microsatellites remain stable for any given strain after serial propagation in cell culture in vitro and in vivo. The results of this preliminary longitudinal study indicate that se-quential HSV-1 isolates obtained from one individual present the same microsatellite profiles. Moreover, the multiplex PCR-based analysis of microsatellites presented here is a simple, specific, precise, and reproducible method that rapidly pro-vides results that can be used to compare HSV-1 strains. Al-together, these results establish the potential use of microsat-ellite polymorphism analysis in epidemiological studies of HSV-1 infections.
As far as the mechanism of microsatellite variability is con-cerned, the changes in repeat numbers at microsatellite loci are generally caused by slip-strand mispairing errors during DNA replication (42). It has been recognized that in prokaryotic cells, as well as in eukaryotic cells, the integrity of the genome is under the control of complex DNA repair systems, including the mismatch repair system. Their functions are used to rec-ognize and repair erroneous insertions, deletions, and the mis-incorporation of bases that can arise during DNA replication and recombination, as well as to repair some forms of DNA damage (2, 15, 50). It is known that some mutational events are normally proofread during replication by the HSV-1 DNA polymerase, which possesses an intrinsic 3⬘ to 5⬘exonuclease activity that attends to the removal of incorrectly incorporated deoxyribonucleoside triphosphates (41). Furthermore, during HSV-1 infection, some host cell DNA repair proteins are cruited to nuclear replication compartments (20, 52). The re-cruitment of host DNA repair proteins seems to benefit viral replication, but their actual roles need to be confirmed. The influence of the recruitment of the mismatch repair system by HSV-1 on the stability or instability of its microsatellite se-quences has not yet been studied. Additionally, some authors have demonstrated that the reduced DNA binding of the UL42 processivity subunit of the HSV-1 DNA polymerase is associ-ated with a significant decrease in replication fidelity (18). All these elements could underlie the diversity of HSV-1 micro-satellites observed in our study.
Regarding their impact on the physiology of HSV-1, micro-satellites might exert subtle influences on the regulation of DNA metabolic processes and viral gene expression as well as chromatin organization, as shown in eukaryote and prokaryote genomes. There is abundant evidence indicating that changes in gene sequence length could inactivate, affect, or change protein function, as shown in human cancer and genetic dis-orders and by the modification of pathogenesis in bacteria (19). Microsatellites may provide a molecular basis for fast adaptation to environmental changes or selective pressure. For instance, many of the mutations associated with the acyclovir-resistant HSV-1 isolates contain insertions or deletions in short homopolymeric G or C sequences in the thymidine kinase (TK) gene (40). Some of these regions are remarkably instable
on May 16, 2020 by guest
http://jcm.asm.org/
and likewise give to the TK-negative mutants the possibility to revert to a TK-positive phenotype, while the phenotype gives the virus the advantage of being able to reactivate and grow in ganglia (13). These variations in the length of the sequence of the TK gene of HSV-1 illustrate the adaptive mechanisms that permit the virus to evade acyclovir therapy yet retain its patho-genicity. In our study, the screen that identified 79 microsat-ellites does not yet constitute an exhaustive survey of the HSV-1 genome; we looked only for total repeats longer than the chosen cutoff (microsatellites of four trinucleotides, five dinucleotides, or nine mononucleotides). This cutoff explains why homopolymeric G or C sequences in the TK gene were not included. In our study, two polymorphic trinucleotide micro-satellites were found within two protein sequences. This may result in changes in the UL46 tegument protein, which mod-ulates the transcriptional induction of alpha genes (3, 54), and US4, which encodes gG, which is a viral envelope glycoprotein that is assumed to contribute to viral entry through the apical surfaces of polarized cells (46). In the second case, our results confirmed the sequence variations of the GAG tandem repeat flanking GAA in gG that were already reported by others (27, 30). To date, these amino acid sequence modifications and their biological significance have not yet been reported.
Otherwise, although the effects of polymorphisms in micro-satellites on the physiology of HSV-1 are largely unknown, the fact that microsatellites were more often found in noncoding regions than in open reading frames suggests that they have no influence on either the virulence of the strain or the evolution of the virus. Since these variations must likely occur at random without offering any replicative or adaptive advantage, these microsatellites may thereby be considered neutral molecular markers. In every instance, they might be used in studies in which HSV-1 strain tracing is necessary: in physiological and epidemiological studies (34) or when nosocomial HSV-1 infec-tions (21, 38) or reinfection with a novel strain (31) are sus-pected. Our study has indentified 23 polymorphic microsatel-lite loci that may be used as molecular markers for epidemiological analysis of HSV-1 strains and HSV-1 strain differentiation. Furthermore, microsatellite-based phylogeny and principal-component analyses, which are the methods usu-ally described in the literature, might be applied to show the clustering of HSV-1 strains. Microsatellites accurately charac-terized the HSV-1 strains from patients evaluated in the present study and were used to ascertain the identity between strains isolated sequentially from the same patient, as well as to differentiate strains in the case of the hypothetical nosocomial transmission of HSV-1 strains. Our preliminary results are promising and should have implications for further epidemio-logical studies of the transmission and the evolution of HSV-1 strains.
ACKNOWLEDGMENTS
This work was supported in part by a grant from IFR113 Immunite´, Cancer et Infection Groupe Hospitalier Pitie´-Salpeˆtrie`re.
We thank Flore Rozenberg for providing strain KOS and Ariel Bardi for proofreading the manuscript.
The authors have no conflicting financial interests.
REFERENCES
1.Adams, G., B. H. Stover, R. A. Keenlyside, T. M. Hooton, T. G. Buchman, B. Roizman, and J. A. Stewart.1981. Nosocomial herpetic infections in a
pe-diatric intensive care unit. Am. J. Epidemiol.113:126–132.
2.Baranovskaya, S., J. L. Soto, M. Perucho, and S. R. Malkhosyan.2001. Functional significance of concomitant inactivation of hMLH1 and hMSH6 in tumor cells of the microsatellite mutator phenotype. Proc. Natl. Acad. Sci.
USA98:15107–15112.
3.Barker, D. E., and B. Roizman.1990. Identification of three genes nones-sential for growth in cell culture near the right terminus of the unique
sequences of long component of herpes simplex virus 1. Virology177:684–
691.
4.Bowden, R., H. Sakaoka, P. Donnelly, and R. Ward.2004. High recombina-tion rate in herpes simplex virus type 1 natural popularecombina-tions suggests
signif-icant co-infection. Infect. Genet. Evol.4:115–123.
5.Bowden, R., H. Sakaoka, R. Ward, and P. Donnelly. 2006. Patterns of Eurasian HSV-1 molecular diversity and inferences of human migrations.
Infect. Genet. Evol.6:63–74.
6.Buchman, T. G., B. Roizman, G. Adams, and B. H. Stover.1978. Restriction endonuclease fingerprinting of herpes simplex virus DNA: a novel
epidemi-ological tool applied to a nosocomial outbreak. J. Infect. Dis.138:488–498.
7.Chaney, S. M., K. G. Warren, and J. H. Subak-Sharpe.1983. Variable restriction endonuclease sites of herpes simplex virus type 1 isolates from
encephalitic, facial and genital lesions and ganglia. J. Gen. Virol. 64(Pt
12):2717–2733.
8.Chou, J., and B. Roizman. 1990. The herpes simplex virus 1 gene for ICP34.5, which maps in inverted repeats, is conserved in several
limited-passage isolates but not in strain 17syn⫹. J. Virol.64:1014–1020.
9.Davis, C. L., D. Field, D. Metzgar, R. Saiz, P. A. Morin, I. L. Smith, S. A. Spector, and C. Wills.1999. Numerous length polymorphisms at short
tan-dem repeats in human cytomegalovirus. J. Virol.73:6265–6270.
10.Davison, A. J., and N. M. Wilkie.1981. Nucleotide sequences of the joint between the L and S segments of herpes simplex virus types 1 and 2. J. Gen.
Virol.55:315–331.
11.Field, D., and C. Wills.1996. Long, polymorphic microsatellites in simple
organisms. Proc. Biol. Sci.263:209–215.
12.Garcia-Barreno, B., T. Delgado, and J. A. Melero.1994. Oligo(A) sequences of human respiratory syncytial virus G protein gene: assessment of their
genetic stability in frameshift mutants. J. Virol.68:5460–5468.
13.Griffiths, A., M. A. Link, C. L. Furness, and D. M. Coen.2006. Low-level expression and reversion both contribute to reactivation of herpes simplex virus drug-resistant mutants with mutations on homopolymeric sequences in
thymidine kinase. J. Virol.80:6568–6574.
14.Gupta, R., T. Warren, and A. Wald.2007. Genital herpes. Lancet370:2127– 2137.
15.Ham, M. F., T. Takakuwa, W. J. Luo, A. Liu, A. Horii, and K. Aozasa.2006. Impairment of double-strand breaks repair and aberrant splicing of ATM and MRE11 in leukemia-lymphoma cell lines with microsatellite instability.
Cancer Sci.97:226–234.
16.Hammer, S. M., T. G. Buchman, L. J. D’Angelo, A. W. Karchmer, B. Roiz-man, and M. S. Hirsch.1980. Temporal cluster of herpes simplex enceph-alitis: investigation by restriction endonuclease cleavage of viral DNA. J.
In-fect. Dis.141:436–440.
17.Hayward, G. S., R. J. Jacob, S. C. Wadsworth, and B. Roizman.1975. Anatomy of herpes simplex virus DNA: evidence for four populations of molecules that differ in the relative orientations of their long and short
components. Proc. Natl. Acad. Sci. USA72:4243–4247.
18.Jiang, C., Y. T. Hwang, J. C. Randell, D. M. Coen, and C. B. Hwang.2007. Mutations that decrease DNA binding of the processivity factor of the herpes simplex virus DNA polymerase reduce viral yield, alter the kinetics of viral DNA replication, and decrease the fidelity of DNA replication. J. Virol.
81:3495–3502.
19.Li, Y. C., A. B. Korol, T. Fahima, and E. Nevo.2004. Microsatellites within
genes: structure, function, and evolution. Mol. Biol. Evol.21:991–1007.
20.Lilley, C. E., C. T. Carson, A. R. Muotri, F. H. Gage, and M. D. Weitzman.
2005. DNA repair proteins affect the lifecycle of herpes simplex virus 1. Proc.
Natl. Acad. Sci. USA102:5844–5849.
21.Linnemann, C. C., Jr., T. G. Buchman, I. J. Light, and J. L. Ballard.1978. Transmission of herpes-simplex virus type 1 in a nursery for the newborn.
Identification of viral isolates by D.N.A. “fingerprinting.” Lanceti:964–966.
22.Luyt, C. E., A. Combes, C. Deback, M. H. Aubriot-Lorton, A. Nieszkowska, J. L. Trouillet, F. Capron, H. Agut, C. Gibert, and J. Chastre.2007. Herpes simplex virus lung infection in patients undergoing prolonged mechanical
ventilation. Am. J. Respir. Crit. Care Med.175:935–942.
23.Maertzdorf, J., L. Remeijer, A. Van Der Lelij, J. Buitenwerf, H. G. Niesters, A. D. Osterhaus, and G. M. Verjans. 1999. Amplification of reiterated sequences of herpes simplex virus type 1 (HSV-1) genome to discriminate
between clinical HSV-1 isolates. J. Clin. Microbiol.37:3518–3523.
24.McGeoch, D. J., M. A. Dalrymple, A. J. Davison, A. Dolan, M. C. Frame, D. McNab, L. J. Perry, J. E. Scott, and P. Taylor.1988. The complete DNA sequence of the long unique region in the genome of herpes simplex virus
type 1. J. Gen. Virol.69(Pt 7):1531–1574.
25.Mirandola, P., T. Ravaioli, and E. Cassai.1997. PCR analysis of human
telomeric repeats present on HHV-6A viral strains. Virus Genes15:29–32.
26.Norberg, P., T. Bergstrom, and J. A. Liljeqvist.2006. Genotyping of clinical
on May 16, 2020 by guest
http://jcm.asm.org/
herpes simplex virus type 1 isolates by use of restriction enzymes. J. Clin.
Microbiol.44:4511–4514.
27.Norberg, P., T. Bergstrom, E. Rekabdar, M. Lindh, and J. A. Liljeqvist.2004. Phylogenetic analysis of clinical herpes simplex virus type 1 isolates identified
three genetic groups and recombinant viruses. J. Virol.78:10755–10764.
28.Perry, L. J., and D. J. McGeoch.1988. The DNA sequences of the long repeat region and adjoining parts of the long unique region in the genome of
herpes simplex virus type 1. J. Gen. Virol.69(Pt 11):2831–2846.
29.Picone, O., Y. Ville, J. M. Costa, C. Rouzioux, and M. Leruez-Ville.2005. Human cytomegalovirus (HCMV) short tandem repeats analysis in
congen-ital infection. J. Clin. Virol.32:254–256.
30.Rekabdar, E., P. Tunback, J. A. Liljeqvist, and T. Bergstrom.1999. Vari-ability of the glycoprotein G gene in clinical isolates of herpes simplex virus
type 1. Clin. Diagn. Lab. Immunol.6:826–831.
31.Remeijer, L., J. Maertzdorf, J. Buitenwerf, A. D. Osterhaus, and G. M. Verjans.2002. Corneal herpes simplex virus type 1 superinfection in patients
with recrudescent herpetic keratitis. Investig. Ophthalmol. Vis. Sci.43:358–
363.
32.Remeijer, L., J. Maertzdorf, P. Doornenbal, G. M. Verjans, and A. D. Osterhaus.2001. Herpes simplex virus 1 transmission through corneal
trans-plantation. Lancet357:442.
33.Rixon, F. J., M. E. Campbell, and J. B. Clements.1984. A tandemly reiter-ated DNA sequence in the long repeat region of herpes simplex virus type 1
found in close proximity to immediate-early mRNA 1. J. Virol.52:715–718.
34.Roest, R. W., W. F. Carman, J. Maertzdorf, A. Scoular, J. Harvey, M. Kant, W. I. Van Der Meijden, G. M. Verjans, and A. D. Osterhaus.2004. Genotypic analysis of sequential genital herpes simplex virus type 1 (HSV-1) isolates of patients with recurrent HSV-1 associated genital herpes. J. Med. Virol.
73:601–604.
35.Roizman, B., D. M. Knipe, and R. Whitley.2007. Herpes simplex viruses, p.
2501–2601.InD. M. Knipe, P. M. Howley, et al. (ed.), Fields virology, 5th ed.
Lippincott Williams & Wilkins, Philadelphia, PA.
36.Roizman, B., and M. Tognon.1983. Restriction endonuclease patterns of herpes simplex virus DNA: application to diagnosis and molecular
epidemi-ology. Curr. Top. Microbiol. Immunol.104:273–286.
37.Sakaoka, H., K. Kurita, Y. Iida, S. Takada, K. Umene, Y. T. Kim, C. S. Ren, and A. J. Nahmias.1994. Quantitative analysis of genomic polymorphism of herpes simplex virus type 1 strains from six countries: studies of molecular
evolution and molecular epidemiology of the virus. J. Gen. Virol. 75(Pt
3):513–527.
38.Sakaoka, H., Y. Saheki, K. Uzuki, T. Nakakita, H. Saito, K. Sekine, and K. Fujinaga.1986. Two outbreaks of herpes simplex virus type 1 nosocomial
infection among newborns. J. Clin. Microbiol.24:36–40.
39.Sakaoka, H., H. Saito, K. Sekine, T. Aomori, L. Grillner, G. Wadell, and K. Fujinaga.1987. Genomic comparison of herpes simplex virus type 1 isolates
from Japan, Sweden and Kenya. J. Gen. Virol.68(Pt 3):749–764.
40.Sasadeusz, J. J., F. Tufaro, S. Safrin, K. Schubert, M. M. Hubinette, P. K.
Cheung, and S. L. Sacks.1997. Homopolymer mutational hot spots mediate
herpes simplex virus resistance to acyclovir. J. Virol.71:3872–3878.
41.Song, L., M. Chaudhuri, C. W. Knopf, and D. S. Parris.2004. Contribution
of the 3⬘- to 5⬘-exonuclease activity of herpes simplex virus type 1 DNA
polymerase to the fidelity of DNA synthesis. J. Biol. Chem.279:18535–18543.
42.Tachida, H., and M. Iizuka.1992. Persistence of repeated sequences that
evolve by replication slippage. Genetics131:471–478.
43.Tautz, D., and M. Renz.1984. Simple sequences are ubiquitous repetitive
components of eukaryotic genomes. Nucleic Acids Res.12:4127–4138.
44.Thi, T. N., C. Deback, I. Malet, P. Bonnafous, Z. Ait-Arkoub, and H. Agut.
2006. Rapid determination of antiviral drug susceptibility of herpes simplex
virus types 1 and 2 by real-time PCR. Antivir. Res.69:152–157.
45.Thurston, M. I., and D. Field.2005. MsatFinder: detection and character-ization of microsatellites. CEH Oxford, Oxford, United Kingdom. http//www .genomics.ceh.ac.uk/msatfinder/.
46.Tran, L. C., J. M. Kissner, L. E. Westerman, and A. E. Sears.2000. A herpes simplex virus 1 recombinant lacking the glycoprotein G coding sequences is defective in entry through apical surfaces of polarized epithelial cells in
culture and in vivo. Proc. Natl. Acad. Sci. USA97:1818–1822.
47.Umene, K., and T. Kawana.2003. Divergence of reiterated sequences in a series of genital isolates of herpes simplex virus type 1 from individual
patients. J. Gen. Virol.84:917–923.
48.Umene, K., and M. Yoshida.1989. Reiterated sequences of herpes simplex virus type 1 (HSV-1) genome can serve as physical markers for the
differ-entiation of HSV-1 strains. Arch. Virol.106:281–299.
49.Vogel, J. U., B. Weber, and H. W. Doerr.1994. Typing and strain differen-tiation of clinical herpes simplex virus type 1 and 2 isolates by polymerase chain reaction and subsequent restriction fragment length polymorphism analysis. Zentralbl. Bakteriol. Parasitenkd. Infektkrankh. Hyg. Abt. 1 Orig.
281:502–512.
50.Wagner, R., Jr., and M. Meselson.1976. Repair tracts in mismatched DNA
heteroduplexes. Proc. Natl. Acad. Sci. USA73:4135–4139.
51.Walker, A., S. J. Petheram, L. Ballard, J. R. Murph, G. J. Demmler, and J. F. Bale, Jr.2001. Characterization of human cytomegalovirus strains by analysis
of short tandem repeat polymorphisms. J. Clin. Microbiol.39:2219–2226.
52.Wilkinson, D. E., and S. K. Weller.2004. Recruitment of cellular recombi-nation and repair proteins to sites of herpes simplex virus type 1 DNA replication is dependent on the composition of viral proteins within prerep-licative sites and correlates with the induction of the DNA damage response.
J. Virol.78:4783–4796.
53.Yoshida, M., and K. Umene.2003. Close association of predominant geno-type of herpes simplex virus geno-type 1 with eczema herpeticum analyzed using restriction fragment length polymorphism of polymerase chain reaction.
J. Virol. Methods109:11–16.
54.Zhang, Y., and J. L. McKnight.1993. Herpes simplex virus type 1 UL46 and UL47 deletion mutants lack VP11 and VP12 or VP13 and VP14,
respec-tively, and exhibit altered viral thymidine kinase expression. J. Virol.67:
1482–1492.