The ribosome is a complex macromolecule responsible for the translation of genetic information into proteins. Experimental data has provided clear evidence that the ribosome undergoes conformational changes during the process of translation. The size of the ribosome coupled with its dynamic nature has made it difficult to determine the detailed structure of the ribosome using traditional structural analysis techniques (e.g. NMR or crystallography). Recently, crystal structures of the ribosome of several organisms have been determined. The crystal structures are consistent with ~90% of the data obtained experimentally. The areas of the crystal structure of the ribosome which are inconsistent with experimental data are concentrated primarily in the areas of the ribosome known to be actively involved in the process of translation. The computational
techniques utilized in this project provide an alternative method for generating structures that are consistent with all the experimental data.
probability base triples in the small subunit of the ribosome. The most likely candidates for base triples in the ribosome were identified as base triples
involving the single stranded nucleotide 121and the 124:237 or the 125:236 base pair. Isomorphic modeling using the algorithm ISOPAIR indicated that a base triple consisting of an interaction between 121 and the 124:237 was the most likely to produce an isomorphic structure for the most frequently occurring
Patricia Babin Fanning
A dissertation submitted to the Graduate Faculty of North Carolina State University
in partial fulfillment of the requirements for the Degree of
Doctor of Philosophy
Biochemistry
Raleigh 2005
APPROVED BY:
________________________________ ________________________________ (Chair of the Advisory Committee)
born and raised in Baton Rouge, Louisiana. After graduating from high school, she attended the University of Dallas and Louisiana State University. She graduated with a Bachelor of Science degree from Louisiana State University in an integrated biology and chemistry program. After working for several years as a lab technician, Patricia completed the equivalent of a Bachelor of Science degree in Computer Science at North Carolina State University. Upon
completion of her training in Computer Science, she began working for IBM. As the 1990's began, it became an evident that cross-functional opportunities in computer science and life sciences would soon be a reality. For Patricia, this integration of disciplines provided an opportunity for her to integrate her two areas of intellectual interest - biochemistry and computer science. Patricia entered the Ph.D. program in Molecular and Structural Biochemistry in 1991 and was admitted to Dr. Paul Wollenzien's lab in 1992 as a graduate student.
I learn the biology as well as the computational techniques. I’ll never forget the day that he gave me a filing box FILLED with papers on the ribosome that he required me to read BEFORE I started the computational work. There were probably 100 papers in that box. I also thank him for giving me time to write the user guide to the first version of MC-SYM. By doing this, he reinforced the importance of contributing to the general good of the scientific community even if you don’t directly profit from it.
In addition, many thanks to Dr. Paul Agris for frequent conversations regarding the structure of the decoding region and to Dr. Stu Maxwell for requiring us to think and not memorize in his Advanced Nucleic Acids course - it was my absolute favorite graduate course. Thanks also to Dr. Jim Brown for his willingness to serve on my committee on such short notice.
PREDICTING THE THREE-DIMENSIONAL STRUCTURE OF 16S
RRNA………...1
LIMITING FACTORS IN PREDICTING THE STRUCTURE... 2
CRITICAL EXPERIMENTS REGARDING 16S RRNA ... 5
ADVANCES IN MODELS... 9
APPENDIX ... 64
REFERENCES... 66
CHAPTER I: STRUCTURAL CHANGES IN BASE-PAIRED REGION 28 IN 16S RRNA CLOSE TO THE DECODING REGION OF THE 30 S RIBOSOMAL SUBUNIT ARE CORRELATED TO CHANGES IN TRNA ABSTRACT ... 80
INTRODUCTION... 82
RESULTS... 85
DISCUSSION... 104
MATERIALS AND METHODS ... 112
ACKNOWLEDGEMENTS ... 119
REFERENCES... 120
CHAPTER II: IDENTITY AND GEOMETRY OF A BASE TRIPLE IN 16S RRNA DETERMINED BY COMPARATIVE SEQUENCE ANALYSIS AND MOLECULAR MODELING ABSTRACT ... 129
INTRODUCTION... 130
RESULTS... 132
DISCUSSION... 155
MATERIALS AND METHODS ... 159
ISOPAIR... 162
ACKNOWLEDGEMENTS ... 164
REFERENCES... 165 CHAPTER III:
CONSTRAINTS, MODELING TECHNIQUES
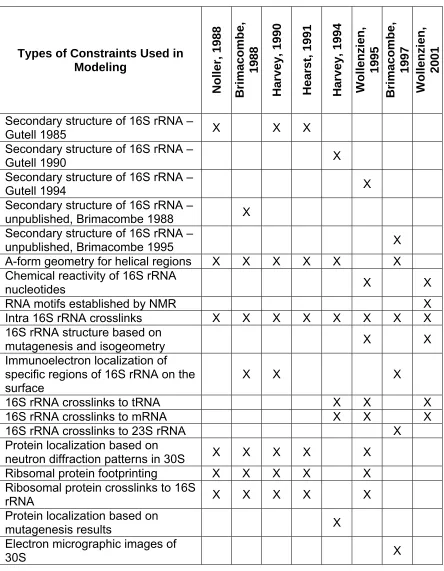
OR MODELS... 7 TABLE I.2 OVERVIEW OF TYPE OF MODELING CONSTRAINTS USED BY MODELLERS ... 8 TABLE I.3 FOOTPRINTING OF 16S RRNA BY RIBOSOMAL PROTEINS. 31 TABLE I.4 16S RIBOSOMAL RNA / RIBOSOMAL PROTEIN
CROSSLINKS…... 45 TABLE I.5 INTRA-RNA CROSSLINKS ... 47 TABLE I.6 HELICAL REGIONS IN 16S RIBOSOMAL RNA IDENTIFIED AS
INSUFFICIENTLY CONSTRAINED FOR UNAMBIGUOUS
PLACEMENT ... 55 TABLE I.7 CROSSLINKS BETWEEN 16S RRNA AND TRNA OR
MRNA……….. ... 57 CHAPTER I
TABLE 1.1 TRNA BINDING BY 30 S SUBUNITS CONTAINING THE 16 S RRNA WILD-TYPE SEQUENCE AND MUTATIONS IN THE
3’-TERMINAL ... 88 TABLE 1.2 POLY(U) BINDING BY RECONSTITUTED 30 S RIBOSOMAL
SUBUNITS CONTAINING MUTATED 16 S RRNA ... 93 CHAPTER II
TABLE 2.1 NUCLEOTIDE FREQUENCIES AT POSITIONS 121(124:237)(A) AND 121(125:236) (B)... 136 TABLE 2.2 DISTRIBUTION (IN PERCENTAGE OF 5,000 PROKARYTIC
SEQUENCES ANALYZED) OF NUCLEOTIDE IDENTITY AT POSITIONS 121(124:237)/(125:236) ... 137 TABLE 2.3 DISTRIBUTION OF SEQUENCE MOTIFS A-D IN BACTERIA AND ARCHEA. ... 140 TABLE 2.4 SUMMARY OF STRUCTURES PREDICTED BY ISOPAIR-MCSYM ANALYSIS. ... 145 TABLE 2.5 RMS DEVIATIONS OF MODELS CONTAINING BASE
FIGURE 1.1 SUCROSE GRADIENT ANALYSIS OF RECONSTITUTED RIBOSOMAL PARTICLES. ... 87 FIGURE 1.2 PATTERNS OF CHEMICAL MODIFICATION IN 16 S RRNA IN 30
S SUBUNITS FOR WILD-TYPE AND MUTANT 16 S RRNA…..96 FIGURE 1.3 SUMMARY OF PATTERNS OF CHEMICAL MODIFICATION IN
THE NATIVE AND RECONSTITUTED 30 S SUBUNITS
CONTAINING THE WILD-TYPE AND MUTATED 16 S RRNA..98 FIGURE 1.4 SECONDARY STRUCTURE IN THE HELIX 28 REGION IN THE
ACTIVE AND INACTIVE CONFORMATIONS AND SUMMARY OF THE SEQUENCE CHANGES THAT RESULT IN REDUCED TRNA BINDING ACTIVITY. ... 102 FIGURE 1.5 REPRESENTATION OF THE THREE-DIMENSIONAL
ARRANGEMENT OF DUPLEX 28 REGION. ... 103 CHAPTER II
FIGURE 2.1 SEQUENCES FOR HELIX 122 REGION USED FOR MODELING. A–D ... 144 FIGURE 2.2 MODELS FOR THE 121(124:237) TRIPLE AND THE
121(125:236) TRIPLE IN MOTIF A. ... 150 FIGURE 2.3 SUPERPOSITION OF THE MODELS FOR THE PROPOSED
HELIX 122 WITH THE BASE TRIPLE 121(125:236) ... 152 FIGURE 2.4 MODELS DEMONSTRATING THE CAUSE OF THE STRONG
NEIGHBOR EFFECT. ... 154 CHAPTER III
FIGURE 3.1 PATTERN OF FE(III)-EDTA CLEAVAGE………178 FIGURE 3.2 DMS AND DEPC REACTIVITY AT N7... 182 FIGURE 3.3 DECODING REGION SECONDARY STRUCTURE AND
CROSSLINK CONSTRAINTS ... 184 FIGURE 3.4 STEREO VIEW OF ALTERNATIVE MODELS FOR HELIX
44A……….189 FIGURE 3.5 STRUCTURAL GEOMETRY OF THE 44A AND 44B REGIONS
FROM THE DELETION OF NUCLEOTIDE 1394 ... 227 FIGURE S.3 MC-SYM MODELS AND THERMUS THERMOPHILUS CRYSTAL
STRUCTURE OF HELIX 28 ... 228 FIGURE S.4 HELIX 44A CHEMICAL REACTIVITY DATA ... 229 FIGURE S.5 MC-SYM MODEL AND THERMUS THERMOPHILUS CRYSTAL STRUCTURE OF HELIX 44A... 231 FIGURE S.6 CHEMICAL REACTIVITY OF THERMUS THERMOPHILUS AND
ESCHERICIA COLI HELIX 44A CRYSTAL STRUCTURE...232 FIGURE S.7 HYDROGEN BONDS BETWEEN NUCLEOTIDE 121 AND BASES
IN HELIX 7A IN THERMUS THERMOPHILUS CRYSTAL
The key relationship between the structure of the ribosome and the function of the ribosome has been so clearly understood from the early days of research into the ribosome that numerous prominent review articles bear both terms –
”structure” and “function” - in their titles (Wittmann 1977; Thompson and Hearst 1983; Brown 1991; Matheson 1992; Bogdanov, Lavrik et al. 1995).
As early as 1968, crystallization of the ribosome was being reported (Barbieri, Laschi et al. 1968; Spencer, Pigram et al. 1969; Thomas and Spencer 1969). However, these crystals were not sufficiently ordered to produce high resolution structures. For example, the first crystallized Thermus thermophilus 70S crystals studied by X-ray crystallography had a diffraction pattern of 20 Å (Trakhanov, Yusupov et al. 1989). Crystals capable of diffracting at low resolutions remained outside the grasp of scientists throughout the 1980’s and 1990’s.
Thus, the notion that the structure of ribosomal RNAs could be successfully predicted was based on previous successful experiences in this area. The widespread availability of computers in the 1990’s gave researchers even more reason to believe that theoretical prediction of the structure of ribosomal RNAs might be possible. However, efforts to predict the three-dimensional structure of ribosomal RNAs were limited by several factors. In the following sections, the factors which limited the ability to predict the structure of the RNA portion of the small subunit of the ribosome (16S rRNA) as well as the history of attempts to generate models of 16S rRNA will be reviewed. The Introduction Section will conclude with a short discussion of MC-SYM, a software package critical to the work discussed in this dissertation.
LIMITING FACTORS IN PREDICTING THE STRUCTURE
The primary factors that limit the ability to successfully predict a three-dimensional structure of 16S rRNA are:
• Insufficient experimental data to constrain theoretical prediction • Insufficient computational power to explore the largely unconstrained
one would need to be able to constrain the locations of a significant percentage of the 1542 nucleotides that comprise E. coli 16S rRNA (E. coli is used as the reference organism.) The most powerful way to constrain the structure of 16S rRNA would be to assume its helical regions were exclusively A-form RNA. However, experimental data that was available in the second half of the twentieth century did not support this assumption. Chemical probing of 16S rRNA and 30S rRNA indicated that only ~20 helical stretches containing ~120 base pairs
showed no chemical reactivity (Moazed, Stern et al. 1986) and did not contain stretches of non-Watson Crick base pairs. Only these regions could be safely assumed to be A-form. Thus, in the absence of being able to assume A-form geometry, a great deal of experimental data was needed to constrain 16S rRNA sufficiently in order to allow its structure to be reliably predicted. In subsequent sections, the experimental data which can be used to constrain 16S rRNA will be discussed; however, the experimental data is insufficient to allow detailed
prediction of the structure of 16S rRNA.
Since insufficient experimental data is available to predict a detailed structure of 16S rRNA, an alternative approach is to identify the entire set of possible
Hadwiger and Fox demonstrated that an entire nucleotide structure can be fixed by specifying the locations of 9 atoms in each nucleotide (Hadwiger and Fox 1991). However, even this simplification is not sufficient to reduce the computational complexity of the problem. Thus, in order to reduce the
computational complexity of predicting the structure of 16S rRNA, most models of 16S rRNA have assumed either that all helical regions of 16S rRNA are A-form RNA or have used fewer atoms to represent the helical regions of 16S rRNA. In subsequent sections, the most significant models and their simplifying
assumptions will be discussed.
choices between modeling 16S rRNA at a non-detailed level that could not truly represent the real structure or generating models using realistic conformations but left a great deal of uncertainty as to which of these realistic conformations was the best choice. No matter what choice was made regarding the level of detail in the modeling, no modeling effort was able to fully explain the dynamics of 16S rRNA. Indeed, even today’s crystal structures of 16S rRNA leave open questions regarding the multiple conformations it assumes.
CRITICAL EXPERIMENTS REGARDING 16S RRNA
Primary
Researcher Year Advancement
Nomura 1968 Reconstitution of functionally active 30S ribosomes Nomura 1970 Assembly mapping of 30S ribosomal proteins Kurland 1970 Identification of binding sites of ribosomal proteins on
16S rRNA
Moore 1972 Determination of quarternary structure by neutron scattering Moore 1975 Neutron scattering study of distribution of protein and
RNA in 30S rRNA Ehresmann,
Zimmerman 1975
Determination of the primary sequence of E. coli 16S rRNA
Noller 1978 Determination of the primary sequence of E. coli 16S rRNA
Spirin 1979 First model of 30S rRNA
Cantor, Hearst 1979 Electron microscopic identification of location of crosslinks within 30S rRNA Noller 1980 First proposed secondary structure model for 16S
rRNA
Crippen 1980 First computer generated model of protein distribution in 30S Cantor 1982 Use of gel electrophoresis to separate crosslinked 16S
rRNA
Cantor 1983 Use of psoralen crosslinking to identify differences between structure of rRNA in 16S and 30S Wollenzien 1985 Incorporation of intramolecular crosslink data into
handbuilt model Noller 1986
Identification of long distance tertiary interactions in 16S rRNA using comparative sequence analysis
(570-866 base pair)
Researcher Year Advancement Wollenzien 1988 Use of reverse transcription to determine the exact position of crosslinks in RNA
Noller 1988 focused on 16S rRNA rather than ribosomal First computer model of small subunit area proteins.
Brimacombe 1989 Crosslinking of mRNA to 16S rRNA
Harvey
(Malhotra, Tan et al.
1990)
First computer model of 16S rRNA that did not require manual intervention. Use of molecular
mechanisms as primary algorithm for RNA folding. Use of “pseudoatoms”.
Lake 1992 Incorporation of additional protein position data into model
Noller
(Noller, Hoffarth et al.
1992)
Widely accepted proof that peptide formation is a result of RNA activity, not ribosomal protein
activity.
Cedergren 1993 Development of MC-SYM
Harvey 1994
Use of surface potential functions to enforce shape of model. Model which included a much
larger set of constraint data than Harvey’s earlier model.
Wollenzien 1995
Full atom model of 16S rRNA, first model of 16S rRNA generated using real nucleotide
conformation sets (MC-SYM) Westhof 1997 Full atom model of core of 16S rRNA Wollenzien 2001 Zero length crosslinks using 4-thiouridine
Westhof 2002
Crystal structure of a complex between aminoglycoside antibiotic and oligo containing A
Modeling
Noller, 1988 Brimacombe,
1988
Harvey, 1990 Hearst, 1991 Harvey, 1994 Wollenzie
1995
Brimacombe,
1997
Wollenzie
2001
Secondary structure of 16S rRNA –
Gutell 1985 X X X
Secondary structure of 16S rRNA –
Gutell 1990 X
Secondary structure of 16S rRNA –
Gutell 1994 X
Secondary structure of 16S rRNA –
unpublished, Brimacombe 1988 X Secondary structure of 16S rRNA –
unpublished, Brimacombe 1995 X
A-form geometry for helical regions X X X X X X Chemical reactivity of 16S rRNA
nucleotides X X
RNA motifs established by NMR X
Intra 16S rRNA crosslinks X X X X X X X X 16S rRNA structure based on
mutagenesis and isogeometry X X Immunoelectron localization of
specific regions of 16S rRNA on the
surface X X X
16S rRNA crosslinks to tRNA X X X 16S rRNA crosslinks to mRNA X X X 16S rRNA crosslinks to 23S rRNA X Protein localization based on
neutron diffraction patterns in 30S X X X X X Ribsomal protein footprinting X X X X X
Ribosomal protein crosslinks to 16S
rRNA X X X X X
Protein localization based on
mutagenesis results X
Electron micrographic images of
earliest models to correctly identify the overall shape of the 16S rRNA in the 30S subunit was published in 1979 (Spirin, Serdyuk et al. 1979) (Figure 1). Several pieces of experimental evidence that had not been available earlier made it possible to generate a more correct model of 16S rRNA:
• A new electron micrograph of 16S rRNA
• New data indicating that many of the ribosomal proteins were compact in structure rather than elongated
• Immunoelectron microscopy data regarding the placement of the ribosomal proteins on the 16S rRNA
Though current models contain considerably more detail, this early handmade model provided the same advantages that computer modeling provides today:
• The model explained a larger set of experimental data than earlier models • It served to generate hypothesis on the structure that could then be tested
comparing the fit to the experimentally measured neutron scattering pattern of 30S subunits tested the validity of the model. Spirin’s team concluded that the model was consistent with the available experimental data.
Noller Model - 1988
In 1988, Harry Noller’s team published the first model of 16S rRNA built with the assistance of computational techniques (Stern, Weiser et al. 1988). (Note: In 1980, Crippen published a computer model of 30S, but the focus of the model was the ribosomal proteins rather than 16S rRNA (Kuntz and Crippen 1980).) This was quite an achievement given that few modeling programs existed and those that did exist required extensive enhancement by Noller’s team in order to accomplish all the tasks needed for the endeavor.
1987)
• The locations of sites in 16 S rRNA that interact with specific ribosomal proteins based on footprinting (Table 4)
• 16S rRNA – protein crosslinks (Table 5) • Intra-RNA crosslinks (Table 6)
Note: See Table for an overview of the constraints used in modeling.
file in FRODO would have resulted in many of the helices being laid over one another in the same general region and requiring the helices to be manually moved in order to scatter them in three dimensional space. In order to overcome this problem, each helical region had a unique value by which its' x and y
coordinates were incremented. The z coordinate for all helical regions was set to 0. As a result, this adjusted .pdb file had the appearance of a secondary
structure diagram when initially displayed in FRODO with the helices all being in the same plane, but with the helices no longer overlapping one another. Once this adjusted .pdb file was created, the file was imported into the graphical display and manipulation software package, FRODO.
Helices were then manually manipulated in three-dimensional space using the software program FBRT to bring them into the best fit for the constraints being applied. Links between helices were broken and then reconnected as necessary in order to facilitate movement of helices. Van der Waals clashes were
prevented using the CONTACTS option in FRODO and distances were
monitored using the DISTANCE option in FRODO. As work progressed, models for two tRNAs were added into FRODO in order to add constraints from
the decision making process as the manual movement of the helices continued in an effort to obtain the best fit for the constraints.
Two methods were used to crosscheck the validity of the resultant model. The first crosscheck method (modeling rule 8) required that the model accommodate homologous segments of smaller and larger RNAs without incurring steric
conflicts. The second crosscheck method was to verify that the overall shape of the resultant molecule resembled the shape of the molecule in electron
micrographs even though the shape from the electron micrographs was not used as a constraint in the modeling.
• Secondary structure of 16S rRNA (Brimacombe’s own secondary structure)
• The three-dimensional co-ordinates for the centers of mass of the 30 S subunit proteins based on neutron diffraction (Capel, Engelman et al. 1987)
• 16S rRNA – protein crosslinks (Table 4) • Intra-RNA crosslinks (Table 5)
• Immunoelectron microscopic localization of specific bases on the surface
of the 30S subunit (Mochalova, Shatsky et al. 1982), (Trempe, Ohgi et al. 1982), (Gornicki, Nurse et al. 1984), (Politz and Glitz 1977), (need
reference for Olson 1979), (Shatsky, Mochalova et al. 1979), (Luhrmann, Stoffler-Meilicke et al. 1981)
Note: See Table 2 for an overview of the constraints used in modeling.
Though there were differences between the Brimacombe secondary structure and the more widely accepted secondary structure of Gutell (Gutell, Weiser et al. 1985), Brimacombe correctly concluded that the differences would be
crosslinks and the neutron scattering data were then used to place the helical regions within the domain in relation to one another. No assumptions regarding interhelical stacking were made in the placement of the helices. Finally, the domains in the wire and tube model were brought together and placed in relation to one another to conform to the immunoelectron microscopic data.
Once the hand built model was completed, the co-ordinates from the mid-points of each helix were measured and used to construct a computer version of the same model. Once the computer version of the model had been constructed, the placement of the helices was refined manually.
The helices that Brimacombe identified as being insufficiently constrained to be accurately placed are shown in Table 6. It is interesting to note that Brimacombe considered the 3’ major domain and the 3’ minor domain to be much more highly constrained than either Noller or Harvey (Table 6).
Brimacombe discussed the consistency of his model with additional data that had not been used to construct the model as a cross validation method. He
dismissed the data as largely being invalid.
Harvey Model - 1990
“Prediction of the three-dimensional structure of Escherichia coli 30S ribosomal subunit: A molecular mechanics approach” (Malhotra, Tan et al. 1990) described the first computer model of 16S rRNA that did not require human manipulation. Instead of manually placing helices in accord with a documented set of modeling rules (Stern, Weiser et al. 1988), Harvey’s group used molecular mechanics to fold a reduced representation of the 16S rRNA chain.
In addition to the difficulties that would be inherent in any large scale modeling activity today, Harvey, like Noller before him, needed to develop molecular modeling tools before even beginning the modeling work itself. To meet the needs of their chosen approach to solving the computational problems, Harvey’s group developed yampp, a molecular mechanics software package.
The experimental constraints used to generate the model included:
• The secondary structure of 16S rRNA as published by Gutell (Gutell, Weiser et al. 1985)
Note: See Table 2 for an overview of the constraints used in modeling.
In Harvey’s approach, 16S rRNA was represented by an increasingly more complex set of pseudoatoms. 16S rRNA was initially represented by 485 pseudoatoms; 1 atom for each helix (“1H”) and 1 atom for each single stranded residue (“1P”) . The size of the pseudoatoms used to represent the helical regions was adjusted based on the varying length of the helical regions in 16S rRNA. The “1H/1P” chain was folded using molecular mechanics to
placed in a model (Table 6). In addition, both groups concluded that
simultaneous satisfaction of all crosslinking and footprinting data could not be accomplished in a single model.
Hearst Model - 1991
In 1991, Hearst produced a computer model which, like Harvey’s model of 16S rRNA, did not require manual intervention (Hubbard and Hearst 1991). Their modeling approach relied primarily on distance geometry. Several commercially available software packages - DSPACE, AMBER and GRAMPS – were utilized in the research. Hearst chose to reduce the computational complexity of the
problem by using pseudohelical structures. In order to ultimately replace the pseudohelices constructs with ideal A-form RNA and to convert the DSPACE files into PDB formats, Hearst’s groups created a new software program, EXPAND.
By 1994, a much larger set of additional constraints were available for modeling 16S rRNA. In addition, Harvey’s group had improved the capabilities of their internally created molecular mechanics program, yammp (Malhotra, Tan et al. 1994). These two factors led Harvey’s group to undertake another 16S rRNA modeling effort (Malhotra and Harvey 1994).
The constraints included in Harvey’s second modeling effort were:
• An updated secondary structure of 16S rRNA (Gutell and Woese 1990) • A subset of the ribosomal protein footprinting data (Stern, Wilson et al.
1986), (Stern, Powers et al. 1988), (Stern, Changchien et al. 1988) (Svensson, Changchien et al. 1988) (Powers, Changchien et al. 1988) (Powers, Stern et al. 1988) (Table 3)
• 16S rRNA – ribosomal protein crosslinks (Brimacombe, Atmadja et al. 1988) (Table 4)
• Intra-16S rRNA crosslinks (Brimacombe, Atmadja et al. 1988) (Table 5) • Neutron scattering data for 30S ribosomal proteins (Capel and
Ramakrishnan 1988)
Mitchell et al. 1993) (Table 7)
Clearly, this is a much larger constraint set than was used in the earlier model. In addition, yampp had been modified to allow for modeling within the envelope of the shape of 30S provided by electron microscopic data.
The researchers were able to assess the extent to which their models met the constraints by measuring the potential energy ranges of the structures. A potential of zero would indicate that the structure had met all of the constraints completely. However, small deviations in the ideal A-RNA geometry of the chains themselves resulted in an average potential energy of 100 to 200 kcal per mole even when the constraints were not applied to the model. Models that were subjected to all the constraints had an average potential energy of 280 to 380 kcal per mole, indicating that attempts to meet the constraint added on
~additional 180 kcal per mole of potential. The single greatest source of
additional potential energy from the constraints derived from the imposition of the surface shape constraints. These constraints added an average of 104.1 kcal per mole to the potential energy.
18, 21, 35, 36, 37, 39, 40 and 44. For example, in Brimacombe’s model, helix 18 was pointed toward the solvent interface; in Harvey’s model, helix 18 was pointed toward the 50S subunit interface. Harvey’s 1994 model also differed from
Noller’s 1988 model in the placement of helices 18, 21, 24, 34, 35, 36, 37, 40 and 44. It is interesting to note that most of the helices represented differently in Brimacombe and Noller’s model are not helices that were considered ambiguous in placement.
Wollenzien Model – 1995
The 1995 Wollenzien model was the first model of 16S rRNA that used “real” nucleotide geometries. As such, it was the first model to address the structure of helical regions within 16S rRNA that could not reasonably be considered to be A-form RNA based on the available experimental data. Examples of regions that provide likely candidates for non A-form geometry include:
• Helices containing Watson-Crick base pairs with reactivity at Watson-Crick hydrogen bonding positions (Moazed, Stern et al. 1986)
• Helices containing non Watson-Crick base pairs • Helices containing consecutive wobble base pairs
• Helices containing single stranded nucleotides which lack a single
chemical probes for hydrogen bonding positions (e.g. nucleotide 926 in helix 18).
In addition to the use of real nucleotide geometries, the model was the first to explicitly model the single stranded regions of 16S rRNA.
The model originated as a wire model that was handbuilt to satisfy the following constraint set:
• Secondary structure of 16S rRNA (Gutell, Larsen et al. 1994) • Footprints of ribosomal proteins on 16S rRNA (Table 3) • Ribosomal RNA – 16S rRNA crosslinks (Table 4)
• Intra-16S rRNA crosslinks (Stiege, Atmadja et al. 1986; Wilms, Noah et al. 1997). (Note: the Wilms crosslinks were unpublished at the time the model was built, but were published in 1997.) (Table 5)
• mRNA – 16S rRNA crosslinks (Rinke-Appel, Junke et al. 1993; Juzumiene, Shapkina et al. 1995) Table 7
• Ribosomal protein - ribosomal protein distances (Capel, Kjeldgaard et al. 1988)
model, the use of real nucleotide geometries allowed chemical reactivity data (Moazed, Stern et al. 1986) and the results of site directed mutagenesis results (Ericson, Minchew et al. 1995) to be as constraints in the construction of the model.
The model was constructed by using MC-SYM to create models for each helix in 16S rRNA based on their secondary structure and reactivity of individual
nucleotides to chemical probes at the Watson Crick hydrogen bonding positions (Moazed and Noller 1986). The goal of the modeling was to create models that were as close to A-form RNA as possible (using the assumption that A-form RNA is the lowest energy conformation and therefore, the most likely form). In
minimized.
P sites and 70S in the post translational state containing tRNA in the P and E sites (Stark, Orlova et al. 1997) as well as improved image processing
techniques(Serysheva, Orlova et al. 1995). Brimacombe indicated that the micrographs contained sufficient density in the areas corresponding to the
expected locations of the tRNA to allow for more precise placement of the helices surrounding the decoding region of 16S rRNA. In addition, new constraints for the placement of the helices within the decoding region were available from new site-directed crosslinking experiments between 16S rRNA and mRNA or between 16S rRNA and tRNA. Finally, Brimacombe’s team developed a new software program, ERNA-3D, which allowed them to explicitly model the single stranded regions of 16S rRNA (Mueller, Doring et al. 1995).
The constraint set used in Brimacombe’s original modeling effort was enhanced by the inclusion of the following constraint sets in the second modeling effort:
• An updated secondary structure of 16S rRNA (Brimacombe 1995) • 16S rRNA crosslinks to 23S rRNA (Mitchell, Osswald et al. 1992) • Fitting of the 16S model to cryo-electron micrographs of 70S containing
structure for the RNA model being built; the output of ERNA-3D was a full atom model of the RNA being built. ERNA-3D supported interactive manipulation of the helical and single stranded regions of the RNA being modeled. As a helical region was moved, its’ accompanying single stranded regions were moved accordingly. Brimacombe’s group began the modeling with the decoding region and utilized the concept of “building up” around the site of docking of the mRNA and tRNA on 16S rRNA. Brimacombe considered the final model to be too far removed from atomic resolution to justify minimization of the model.
In Brimacombe’s 1988 model, he indicated that a significant number of helices in the 5’ domain of 16S rRNA lacked sufficient constraints to precisely define their location (Table 6). In the 1997 model, the only helices that Brimacombe
identified as being placed ambiguously due to lack of constraints were helices 17, 33 and 45 (Table 6).
Wollenzien Model – 2001
1997) Note: See Table 5 for an explicit identification of the crosslink constraints used in the 2001 model, but not the 1995 model.
• NMR structure for GNRA hairpin loops (Heus and Pardi 1991) • NMR structure for CUUG hairpin loops (Jucker and Pardi 1995)
• NMR structure for methylated tetraloops in 16S rRNA (Rife and Moore 1998)
• NMR structure for S8 binding site on 16S rRNA (Kalurachchi, Uma et al. 1997)
• Loop E RNA motif identified in 5S rRNA by crystallography (Dallas and Moore 1997)
• Structure for helix 28 based on mutagenesis data (Ericson, Minchew et al. 1995)
Note: See Table 2 for an overview of the constraints used in modeling.
underlying MC-SYM.
In the mid 1980’s, Robert Cedergren had the foresight to realize that ever-increasing number of biologically relevant small RNAs coupled with the inability of crystal and NMR technology to keep up with the need to determine the
structure of these RNAs created the need for computational tools with the ability to predict the structure of RNA (Gautheret, Major et al. 1993). Dr. Cedergren formed a ground-breaking collaboration between his department and the
Computer Science department at the University of Montreal to create the needed tools. The result was MC-SYM, the first computer algorithm capable of dealing with non-A-form geometries in RNA (Major, Turcotte et al. 1991).
was sufficient to generate models consistent with the experimental data
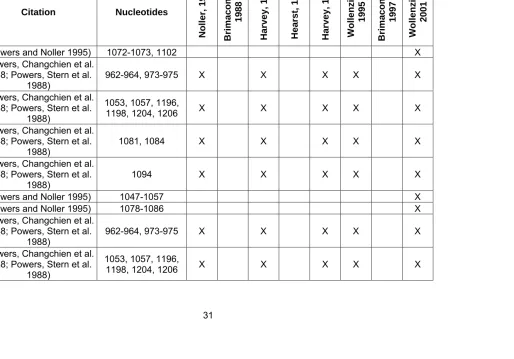
S2 (Powers and Noller 1995) 1072-1073, 1102 X
S2 (Powers, Changchien et al. 1988; Powers, Stern et al. 1988)
962-964, 973-975 X X X X X
S2
(Powers, Changchien et al. 1988; Powers, Stern et al.
1988)
1053, 1057, 1196,
1198, 1204, 1206 X X X X X
S2
(Powers, Changchien et al. 1988; Powers, Stern et al.
1988)
1081, 1084 X X X X X
S2 (Powers, Changchien et al. 1988; Powers, Stern et al. 1988)
1094 X X X X X
S3 (Powers and Noller 1995) 1047-1057 X
S3 (Powers and Noller 1995) 1078-1086 X
S3
(Powers, Changchien et al. 1988; Powers, Stern et al.
1988) 962-964, 973-975 X X X X X
S3
(Powers, Changchien et al. 1988; Powers, Stern et al.
1988)
1053, 1057, 1196,
S3 (Powers, Changchien et al. 1988; Powers, Stern et al. 1988)
1094 X X X X X
S4 (Powers and Noller 1995) 406-411, 427 X
S4 (Powers and Noller 1995) 437-440 X
S4 (Powers and Noller 1995) 500-504, 541-543 X
S4
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
31, 38 X X X X X
S4
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
497-498 X X X X X
S4
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
S4
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
724-725, 727 X X X X X
S4
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
893 X X X X X
S5
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
26 X X X X X
S6 see S18 X
S7 (Powers and Noller 1995) 938-939 X
S7 (Powers and Noller 1995) 946, 1235 X
S7 (Powers and Noller 1995) 1236-1241, 1289-1290 X
S7 (Powers, Changchien et al. 1988; Powers, Stern et al. 1988)
1382 X X X X X
S7
(Powers, Changchien et al. 1988; Powers, Stern et al.
1988) 935-940, 1346 X X X X X
S7
(Powers, Changchien et al. 1988; Powers, Stern et al.
1988)
944-945, 949-951 X X X X X
S7 (Powers, Changchien et al. 1988; Powers, Stern et al. 1988)
1238, 1248-1251, 1256, 1286-1288,
1299-1300
X X X X X
S7
(Powers, Changchien et al. 1988; Powers, Stern et al.
1988)
1302, 1316,
1331-1334 X X X X X
S7
(Powers, Changchien et al. 1988; Powers, Stern et al.
1988)
1349, 1360, 1362,
1365, 1374, 1377 X X X X X
S8 (Powers and Noller 1995) 598-599, 642-643 X
S8 (Svensson, Changchien et al. 1988) 573, 575, 583 X X X X X
S8 (Svensson, Changchien et al. 1988) 811 X X X X X
S8 (Svensson, Changchien et
al. 1988) 858-859 X X X X X
S9 (Powers and Noller 1995) 1114-1118 X
S9 (Powers and Noller 1995) 1126-1130, 1148-1150 X
S9 (Powers and Noller 1995) 1248-1252, 1279-1280 X
S9
(Powers, Changchien et al. 1988; Powers, Stern et al.
1988)
1126-1127, 1130,
1145-1146 X X X X X
S9
(Powers, Changchien et al. 1988; Powers, Stern et al.
1988)
1279-1281 X X X X X
S10 (Powers, Changchien et al. 1988; Powers, Stern et al. 1988)
1150 X X X X X
S11 (Powers and Noller 1995) 706-708 X
S11
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
687, 694-695, 698,
703-705 X X X X X
S11
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
776, 778 X X X X X
S11
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
819 X X X X X
S11
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
890 X X X X X
S12 (Powers and Noller 1995) 517, 522-524, 536 X
S12
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
21 X X X X X
S12
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
558 X X X X X
S12
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
521 X X X X X
S12
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
563, 881, 886-887 X X X X X
S12
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
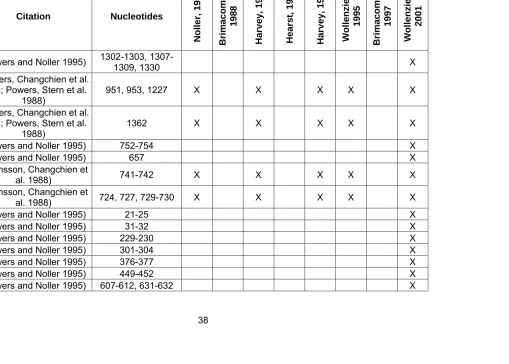
S13 (Powers and Noller 1995) 1302-1303, 1307-1309, 1330 X
S13
(Powers, Changchien et al. 1988; Powers, Stern et al.
1988)
951, 953, 1227 X X X X X
S14 (Powers, Changchien et al. 1988; Powers, Stern et al. 1988)
1362 X X X X X
S15 (Powers and Noller 1995) 752-754 X
S15 (Powers and Noller 1995) 657 X
S15 (Svensson, Changchien et
al. 1988) 741-742 X X X X X
S15 (Svensson, Changchien et al. 1988) 724, 727, 729-730 X X X X X
S16 (Powers and Noller 1995) 21-25 X
S16 (Powers and Noller 1995) 31-32 X
S16 (Powers and Noller 1995) 229-230 X
S16 (Powers and Noller 1995) 301-304 X
S16 (Powers and Noller 1995) 376-377 X
S16 (Powers and Noller 1995) 449-452 X
S16
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
31 X X X X X
S16
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
51, 362-364 X X X X X
S16
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
119, 315 X X X X X
S16
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
301-302 X X X X X
S16
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
S16
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
607-609, 619-622 X X X X X
S16
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
718 X X X X X
S17 (Powers and Noller 1995) 234-235 X
S17 (Powers and Noller 1995) 254-255, 264-265, 279 X
S17
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
246, 251, 262-266,
274, 279-281 X X X X X
S18
+S6 (Powers and Noller 1995) 736, 743 X
S18
+S6 (Powers and Noller 1995) 673-675 X
S18
S18
+S6 (Svensson, Changchien et al. 1988) 664 X X X X X
S18 +S6
(Svensson, Changchien et al. 1988)
674-676, 687,
694-695, 698, 703-705 X X X X X
S18 +S6
(Svensson, Changchien et
al. 1988) 715-718 X X X X X
S18
+S6 (Svensson, Changchien et al. 1988) 728 X X X X X
S18 +S6
(Svensson, Changchien et
al. 1988) 778, 803, 812 X X X X X
S19 (Powers and Noller 1995) 955 X
S19 (Powers and Noller 1995) 958-959, 976-978 X
S19 (Powers and Noller 1995) 1013 X
S19 (Powers and Noller 1995) 1046-1047 X
S19 (Powers and Noller 1995) 1312-1314 X
S19 (Powers, Changchien et al. 1988; Powers, Stern et al. 1988)
1395-1398 X X X X X
S19
(Powers, Changchien et al. 1988; Powers, Stern et al.
S19 (Powers, Changchien et al. 1988; Powers, Stern et al. 1988)
958-959, 976-980 X X X X X
S19
(Powers, Changchien et al. 1988; Powers, Stern et al.
1988) 983 X X X X X
S19
(Powers, Changchien et al. 1988; Powers, Stern et al.
1988)
993-994, 996, 1014,
1016, 1036 X X X X X
S19 (Powers, Changchien et al. 1988; Powers, Stern et al. 1988)
1046, 1213 X X X X X
S19
(Powers, Changchien et al. 1988; Powers, Stern et al.
1988) 1316-1319 X X X X X
S19
(Powers, Changchien et al. 1988; Powers, Stern et al.
1988)
1361 X X X X X
S20 (Powers and Noller 1995) 172-174 X
S20 (Powers and Noller 1995) 184-187 X
S20
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
108 X X X X X
S20
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
194 X X X X X
S20
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
246, 251, 274 X X X X X
S20
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
329 X X X X X
S20
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
1433-1434,
S21
(Stern, Wilson et al. 1986; Stern, Changchien et al. 1988; Stern, Powers et al.
1988)
Nucleotides Noller, 1988
Brimacombe,1988
Harvey, 1990 Hearst, 1991 Harvey, 1994
Wollenzien, 1995 Brimacombe, 1997 Wollenzien, 2001
S3 (Osswald, Greuer et al. 1987) 5’ X X X
S3 (Greuer, Osswald et al. 1987) 1155-1158 X X X X X X X
S4 (Osswald, Greuer et al. 1987) 5’ end X X X
S4 (Greuer, Osswald et al. 1987) 413 X X X X X X X
S5 (Osswald, Greuer et al. 1987) 5’ end X X X X X X X
S5 (Osswald, Greuer et al. 1987) 559-561 X X X X X X X
S7 (Osswald, Greuer et al. 1987) 1238-1240 X X X X X X X
S7 (Wower and Brimacombe 1983) 1377-1378 X X X X X X X
S7 (Greuer, Osswald et al. 1987) 3’ end X X X
S8 (Osswald, Greuer et al. 1987) 5’ end X X X
S8 (Wower and Brimacombe 1983) 593-597 X X X X X X X
S8 (Wower and Brimacombe 1983) 629-633 X X X X X X X
S8 (Wower and Brimacombe 1983) 651-654 X X X X X X X
S9 (Greuer, Osswald et al. 1987) 954 X X X X X X X
Nucleotides Noller, 1988
Brimacombe,1988
Harvey, 1990 Hearst, 1991 Harvey, 1994
Wollenzien, 1995 Brimacombe, 1997 Wollenzien, 2001
S11 (Osswald, Greuer et al. 1987) 702-705 X X X X X X X
S13 (Osswald, Greuer et al. 1987) 189-191 X X X X
S13 (Osswald, Greuer et al. 1987) 1337-1338 X X X X X X X S17 (Greuer, Osswald et al. 1987) 278-280 X X X X X X X S17 (Kyriatsoulis, Maly et al. 1986) 629-633 X X X X X X X S18 (Greuer, Osswald et al. 1987) 845-851 X X X X X X X
S18 (Kyriatsoulis, Maly et al. 1986) 3’ end X X X X
S19 (Wower and Brimacombe 1983) 1223-1231 X X X X X X X
S21 (Greuer, Osswald et al. 1987) 693-697 X X X X
S21 (Wower and Brimacombe 1983) 723-724 X X X X
Crosslink Number
Noller, 1988
Brimacombe,1988
Harvey, 1990 Hearst, 1991 Harvey, 1994
Wollenzien, 1995 Brimacombe, 1997 Wollenzien, 2001
1 Wollenzien, Murphy et al. 1985) (Wollenzien and Cantor 1982; 5':3' X
2 (Wollenzien, Murphy et al. 1985) 5':240 X
3 (Wollenzien, Murphy et al. 1985) 5':340 X
4 (Wollenzien, Murphy et al. 1985) 5':440 X
5 (Wollenzien, Murphy et al. 1985) 5':690 X
6 (Wollenzien, Murphy et al. 1985) 5':800 X
7 (Juzumiene and Wollenzien 2000) 9:298-300 X
8 (Juzumiene and Wollenzien 2000) 9:313-314 X
9 (Juzumiene and Wollenzien 2000) 9:334-338 X
10 (Juzumiene and Wollenzien 2000) 9:343-347 X
11 (Juzumiene and Wollenzien 2000) 9:556-563 X
12 (Juzumiene and Wollenzien 2000) 9:570-577 X
13 (Juzumiene and Wollenzien 2000) 9:892 X
14 (Juzumiene and Wollenzien 2000) 9:901 X
15 (Juzumiene and Wollenzien 2000) 9:906 X
Crosslink Number
Noller, 1988
Brimacombe,1988
Harvey, 1990 Hearst, 1991 Harvey, 1994
Wollenzien, 1995 Brimacombe, 1997 Wollenzien, 2001
17 (Juzumiene and Wollenzien 2000) 9:921-926 X
18 (Juzumiene and Wollenzien 2000) 9:1071-1072 X
19 (Juzumiene and Wollenzien 2000) 9:1080-1083 X
20 (Juzumiene and Wollenzien 2000) 9:1092 X
21 (Juzumiene and Wollenzien 2000) 9:1149-1151 X
22 (Juzumiene and Wollenzien 2000) 9:1330 X
23 (Juzumiene and Wollenzien 2000) 9:1392-1398 X
24 (Juzumiene and Wollenzien 2000) 9:1400-1401 X
25 (Juzumiene and Wollenzien 2000) 9:1413 X
26 (Juzumiene and Wollenzien 2000) 9:1492-1493 X
27 (Juzumiene and Wollenzien 2001) 14:560-562 X
28 (Juzumiene and Wollenzien 2001) 14:570-571 X
29 (Juzumiene and Wollenzien 2001) 14:903-907 X
30 (Juzumiene and Wollenzien 2001) 14:917-920 X
31 (Juzumiene and Wollenzien 2001) 14:1081-1082 X
32 (Juzumiene and Wollenzien 2001) 14:1164-1168 X
Crosslink Number
Noller, 1988
Brimacombe,1988
Harvey, 1990 Hearst, 1991 Harvey, 1994
Wollenzien, 1995 Brimacombe, 1997 Wollenzien, 2001
34 (Juzumiene and Wollenzien 2001) 14:1492 X
35 (Juzumiene and Wollenzien 2001) 17:560-562 X
36 (Juzumiene and Wollenzien 2001) 17:570-571 X
37 (Juzumiene and Wollenzien 2001) 17:903-904 X
38 (Juzumiene and Wollenzien 2001) 17:1082-1084 X
39 (Juzumiene and Wollenzien 2001) 17:1121-1125 X
40 (Juzumiene and Wollenzien 2001) 17:1130-1131 X
41 (Atmadja, Brimacombe et al. 1985) 31:48 X X X X X X X X
42 (Atmadja, Stiege et al. 1986) 31:306 X X X X X X X X
43 (Wilms, Noah et al. 1997) 54:353 X X
44 (Wollenzien, Murphy et al. 1985) 110:310 X
45 (Stiege, Atmadja et al. 1986) 118:288-289 X X X X X X X X
46 (Wollenzien, Murphy et al. 1985) 180:540 X
47 (Wollenzien, Murphy et al. 1985) 240:3' X
48 (Wilms, Noah et al. 1997) 244:894 X X
Crosslink Number
Noller, 1988
Brimacombe,1988
Harvey, 1990 Hearst, 1991 Harvey, 1994
Wollenzien, 1995 Brimacombe, 1997 Wollenzien, 2001
50 (Wollenzien, Murphy et al. 1985) 270:700 X
51 (Wollenzien, Murphy et al. 1985) 300:800 X
52 (Wollenzien, Murphy et al. 1985) 330:3' X
53 (Stiege, Atmadja et al. 1986) 366-369:397-399 X
54 (Wollenzien, Murphy et al. 1985) 420:3' X
55 (Wilms, Noah et al. 1997) 441:494 X X
56 (Wollenzien, Murphy et al. 1985) 450:3' X
57 (Wollenzien, Murphy et al. 1985) 470:3' X
58 (Wollenzien and Cantor 1982) 510:3' X
59 (Mundus & Wollenzien, unpubl.) 533:1359 X
60 (Mundus & Wollenzien, unpubl.) 533:1384 X
61 (Mundus & Wollenzien, unpubl.) 533:1397 X
62 (Mundus & Wollenzien, unpubl.) 533:1401 X
63 (Mundus & Wollenzien, unpubl.) 533:1406 X
64 (Mundus & Wollenzien, unpubl.) 533:1409 X
65 (Mundus & Wollenzien, unpubl.) 561:884 X
Crosslink Number
Noller, 1988
Brimacombe,1988
Harvey, 1990 Hearst, 1991 Harvey, 1994
Wollenzien, 1995 Brimacombe, 1997 Wollenzien, 2001
67 (Mundus & Wollenzien, unpubl.) 561:935 X
68 (Mundus & Wollenzien, unpubl.) 561:1380 X
69 (Wilms, Noah et al. 1997) 562:884 X X
70 (Wollenzien, Murphy et al. 1985) 570:780 X
71 (Wilms, Noah et al. 1997) 582:760 X X
72 (Wollenzien, Murphy et al. 1985) 620:1240 X
73 (Thompson and Hearst 1983) 625:1420 X X
74 (Wollenzien, Murphy et al. 1985) 660:3' X
75 (Atmadja, Stiege et al. 1986) 690 to 790 loop X X X X X X X X 76 (Atmadja, Stiege et al. 1986) 693-696:794&799 X X X X X X X X
77 (Wollenzien, Murphy et al. 1985) 710:920 X
78 (Wollenzien, Murphy et al. 1985) 720:1360 X
79 (Wollenzien, Murphy et al. 1985) 760:1120 X
80 (Mundus and Wollenzien 1998) 788:561 X
81 (Mundus and Wollenzien 1998) 788:693 X
82 (Mundus and Wollenzien 1998) 788:723 X
Crosslink Number
Noller, 1988
Brimacombe,1988
Harvey, 1990 Hearst, 1991 Harvey, 1994
Wollenzien, 1995 Brimacombe, 1997 Wollenzien, 2001
84 (Mundus and Wollenzien 1998) 788:920 X
85 (Atmadja, Stiege et al. 1986) 793:1517 X X X X X X X X
86 (Wollenzien, Murphy et al. 1985) 850:1240 X
87 (Wollenzien, Murphy et al. 1985) 880:1310 X
88 (Wollenzien, Murphy et al. 1985) 894:1468 X
89 (Wollenzien, Murphy et al. 1985) 920:3' X
90 (Mundus & Wollenzien, unpubl.) 920:555 X
91 (Mundus & Wollenzien, unpubl.) 920:1396 X
92 (Mundus & Wollenzien, unpubl.) 920:1409 X
93 (Wollenzien and Cantor 1982) 930:3' X
94 (Wollenzien, Murphy et al. 1985) 930:1420 X
95 (Wilms, Noah et al. 1997) 934:1345 X X
96 (Thompson and Hearst 1983) 956:1506 X
97 (Mundus & Wollenzien, unpubl.) 965:552 X
98 (Mundus & Wollenzien, unpubl.) 965:562 X
99 (Mundus & Wollenzien, unpubl.) 965:1049 X
Crosslink Number
Noller, 1988
Brimacombe,1988
Harvey, 1990 Hearst, 1991 Harvey, 1994
Wollenzien, 1995 Brimacombe, 1997 Wollenzien, 2001
101 (Mundus & Wollenzien, unpubl.) 965:1183 X
102 (Mundus & Wollenzien, unpubl.) 965:1364 X
103 (Mundus & Wollenzien, unpubl.) 965:1381 X
104 (Mundus & Wollenzien, unpubl.) 965:1400 X
105 (Mundus & Wollenzien, unpubl.) 965:1409 X
106 (Mundus & Wollenzien, unpubl.) 965:1420 X
107 (Wilms, Noah et al. 1997) 967:1400 X X
108 (Shapkina & Wollenzien, unpubl.) 976:1362 X
109 (Wollenzien, Murphy et al. 1985) 980:3' X
110 (Wollenzien, Murphy et al. 1985) 980:1300 X
111 (Wilms, Noah et al. 1997) 991:1212 X X
112 (Wollenzien, Murphy et al. 1985) 1030:1360 X
113 (Wollenzien, Murphy et al. 1985) 1030:1410 X
114 (Wilms, Noah et al. 1997) 1052:1200 X X
115 (Shapkina & Wollenzien, unpubl.) 1064:1190 X
116 (Wollenzien, Murphy et al. 1985) 1080:1280 X
Crosslink Number
Noller, 1988
Brimacombe,1988
Harvey, 1990 Hearst, 1991 Harvey, 1994
Wollenzien, 1995 Brimacombe, 1997 Wollenzien, 2001
118 (Wilms, Noah et al. 1997) 1093:1182 X X
119 (Wollenzien, Murphy et al. 1985) 1120:3' X
120 (Wilms, Noah et al. 1997) 1126:1281 X X
121 (Wollenzien, Murphy et al. 1985) 1140:1370 X
122 (Wollenzien, Murphy et al. 1985) 1240:3' X
123 (Wollenzien, Murphy et al. 1985) 1300:3' X
124 (Expert-Bezancon and Wollenzien 1985) 1316:1361 X
125 (Wollenzien, Murphy et al. 1985) 1350:3' X
126 (Wilms, Noah et al. 1997) 1402:1501 X X
127 (Mundus & Wollenzien, unpubl.) 1406:792 X
5’ 7a X X X
5’ 7b X X X
5’ 8a X X X X
5’ 8b X X X X
5’ 9 X X X
5’ 10 X X X
5’ 13 X X
5’ 14 X X
5’ 15 X
5’ 16 X X
5’ 17a X X X
5’ 17b X X X X
Central 20 X
Central 26 X
3’ major 31 X
3’ major 33a X X X X
3’ major 33b X X X X
3’ major 33c X X X
3’ major 41 X
3’ minor 44a X X X 3’ minor
domain 44b X X X
3’ minor
Interaction Distance
Interact Uncertainty Noller, 1988
Brimacombe,1988
Harvey, 1990 Hearst, 1991 Harvey, 1994
Wollenzien, 1995 Brimacombe, 1997 Wollenzien, 2001
(Brimacombe, Mitchell et al. 1993); Richard Brimacombe (personal
communication)
966-1388 10 4
Cross-link between P-site tRNA nucleotide 32 and 16S nucleotides 966, 1338 and 1378. The cross-link
to 1378 may be from the A-site
X X X
(Brimacombe, Mitchell et al. 1993); Richard Brimacombe (personal
communication)
966-1378 10 4
Cross-link between P-site tRNA nucleotide 32 and 16S nucleotides 966, 1338 and 1378. The cross-link
to 1378 may be from the A-site
X X X
(Brimacombe, Mitchell et al. 1993); Richard Brimacombe (personal
communication)
1338-1378 10 4
Cross-link between P-site tRNA nucleotide 32 and 16S nucleotides 966, 1338 and 1378. The cross-link
to 1378 may be from the A-site
X X X
(Brimacombe, Mitchell et al. 1993); Richard Brimacombe (personal communication) mRNA/tRNA sphere-966, 1338, 1378 5 4
Cross-link between P-site tRNA nucleotide 32 and 16S nucleotides 966, 1338 and 1378. The cross-link
to 1378 may be from the A-site
Interaction Distance
Interact Uncertainty Noller, 1988
Brimacombe,1988
Harvey, 1990 Hearst, 1991 Harvey, 1994
Wollenzien, 1995 Brimacombe, 1997 Wollenzien, 2001
(Prince, Taylor et al. 1982) 1400-966 20 4
Cross-link between P-site tRNA nucleotide 34 and nucleotide 1400 on the 16 S RNA, and the previous entry
in this table
X X X
(Prince, Taylor et al. 1982) 1400-1338 20 4
Cross-link between P-site tRNA nucleotide 34 and nucleotide 1400 on the 16 S RNA, and the previous entry
in this table
X X X
(Prince, Taylor et al. 1982) 1400-1378 20 4
Cross-link between P-site tRNA nucleotide 34 and nucleotide 1400 on the 16 S RNA, and the previous entry
in this table
X X X
(Prince, Taylor et al. 1982) mRNA/tRNA sphere-1400 5 4
Cross-link between P-site tRNA nucleotide 34 and nucleotide 1400 on the 16 S RNA, and the previous entry
in this table
Interaction Distance
Interact Uncertainty Noller, 1988
Brimacombe,1988
Harvey, 1990 Hearst, 1991 Harvey, 1994
Wollenzien, 1995 Brimacombe, 1997 Wollenzien, 2001
(Tate, Greuer et al. 1990) 1408-966 30 4 mRNA and nucleotide 1408 on the 16 Cross-link between +4 nucleotide on S RNA, and the first entry in this table
X X X
(Tate, Greuer et al. 1990) 1408-1338 30 4
Cross-link between +4 nucleotide on mRNA and nucleotide 1408 on the 16
S RNA, and the first entry in this table X X X
(Tate, Greuer et al. 1990) 1408-1378 30 4
Cross-link between +4 nucleotide on mRNA and nucleotide 1408 on the 16 S RNA, and the first entry in this table
X X X
(Tate, Greuer et al. 1990; Dontsova, Kopylov et al. 1991; Rinke-Appel, Junke
et al. 1991; Dontsova, Dokudovskaya et al. 1992)
1052-1408 10 4
Cross-links between mRNA and 16 S RNA (+11 to 532, +7 to 1395, +6 to
1052, +6 to 1408)
Interaction Distance
Interact Uncertainty Noller, 1988
Brimacombe,1988
Harvey, 1990 Hearst, 1991 Harvey, 1994
Wollenzien, 1995 Brimacombe, 1997 Wollenzien, 2001
(Tate, Greuer et al. 1990; Dontsova, Kopylov et al. 1991; Rinke-Appel, Junke
et al. 1991; Dontsova, Dokudovskaya et al. 1992)
1052-1395 10 4
Cross-links between mRNA and 16 S RNA (+11 to 532, +7 to 1395, +6 to
1052, +6 to 1408)
X X X
(Tate, Greuer et al. 1990; Dontsova, Kopylov et al. 1991; Rinke-Appel, Junke
et al. 1991; Dontsova, Dokudovskaya et al. 1992)
1052-532 25 4 Cross-links between mRNA and 16 S RNA (+11 to 532, +7 to 1395, +6 to 1052, +6 to 1408)
X X X
(Tate, Greuer et al. 1990; Dontsova, Kopylov et al. 1991; Rinke-Appel, Junke
et al. 1991; Dontsova, Dokudovskaya et al. 1992)
1395-532 20 4
Cross-links between mRNA and 16 S RNA (+11 to 532, +7 to 1395, +6 to
Interaction Distance
Interact Uncertainty Noller, 1988
Brimacombe,1988
Harvey, 1990 Hearst, 1991 Harvey, 1994
Wollenzien, 1995 Brimacombe, 1997 Wollenzien, 2001
(Tate, Greuer et al. 1990; Dontsova, Kopylov et al. 1991; Rinke-Appel, Junke
et al. 1991; Dontsova, Dokudovskaya et al. 1992)
1395-1408 15 4
Cross-links between mRNA and 16 S RNA (+11 to 532, +7 to 1395, +6 to
1052, +6 to 1408)
X X X
(Tate, Greuer et al. 1990; Dontsova, Kopylov et al. 1991; Rinke-Appel, Junke
et al. 1991; Dontsova, Dokudovskaya et al. 1992)
mRNA/tRNA sphere-532, 1052, 1395,
1408
5 4 Cross-links between mRNA and 16 S RNA (+11 to 532, +7 to 1395, +6 to 1052, +6 to 1408)
X X X
(Tate, Greuer et al. 1990; Dontsova, Kopylov et al. 1991; Rinke-Appel, Junke
et al. 1991; Dontsova, Dokudovskaya et al. 1992)
1400-1408 15 4
mRNA cross-links to the 16 S RNA, and cross-link between P-site tRNA nucleotide 34 and nucleotide 1400 on
the 16 S RNA
Interaction Distance
Interact Uncertainty Noller, 1988
Brimacombe,1988
Harvey, 1990 Hearst, 1991 Harvey, 1994
Wollenzien, 1995 Brimacombe, 1997 Wollenzien, 2001
(Tate, Greuer et al. 1990; Dontsova, Kopylov et al. 1991; Rinke-Appel, Junke
et al. 1991; Dontsova, Dokudovskaya et al. 1992)
1400-1052 20 4
mRNA cross-links to the 16 S RNA, and cross-link between P-site tRNA nucleotide 34 and nucleotide 1400 on
the 16 S RNA
X X X
(Tate, Greuer et al. 1990; Dontsova, Kopylov et al. 1991; Rinke-Appel, Junke
et al. 1991; Dontsova, Dokudovskaya et al. 1992)
1400-1395 25 4
mRNA cross-links to the 16 S RNA, and cross-link between P-site tRNA nucleotide 34 and nucleotide 1400 on
the 16 S RNA
X X X
(Podkowinski and Gornicki
1991) S7-1400 20 4
Cross-link between S7 and P-site
Interaction Distance
Interact Uncertainty Noller, 1988
Brimacombe,1988
Harvey, 1990 Hearst, 1991 Harvey, 1994
Wollenzien, 1995 Brimacombe, 1997 Wollenzien, 2001
(Moazed and Noller 1986; Moazed and Noller 1990)
mRNA/tRNA sphere-529, 530, 531, 892, 1405, 1408, 1492, 1493, 1494
5 4 Protection of the 16 S RNA
nucleotides by tRNA at the A-site X X X
(Moazed and Noller 1986; Moazed and Noller 1990)
mRNA/tRNA sphere-532, 693, 794, 795, 926, 966, 1338, 1339, 1381, 1399, 1400, 1401
5 4 nucleotides by tRNA at the P-site Protection of the 16 S RNA X X X
(Weitzmann, Tumminia et al. 1991)
mRNA/tRNA sphere-S7,
S19
• Residues 1258-1259/1276-1277 are paired
• Residues 554-556/884-886 are unpaired
• Residues 888-891/909-912 are unpaired
Rule 2: Base-paired elements are modeled as A-form RNA
Rule 3: 30S ribosomal proteins are modeled as spheres whose volume is determined by their molecular weight using a partial specific volume of 0.74 cm3/g. The co-ordinates for the center of mass of the spherical proteins were those published by Capel et al. (Capel, Engelman et al. 1987)
Rule 4: Strong protection of a nucleotide by a specific protein creates a
constraint that the nucleotide must be placed within the sphere of the protein in the model.
Rule 5: Protein-RNA and RNA-RNA crosslinking data are used, if and only if: • The crosslinking data was obtained in intact 30S or 70S ribosomal
particles known to retain their functional properties • The RNA crosslinks were verified by direct sequencing.
Rule 6: In general, single stranded regions of RNA utilize P-P backbone
mitochondrial) should be superimposable on the 16S model without needing to exceed P-P backbone lengths of 7Å in the smaller 16S-like RNA. Homologous segments of larger 16S-like RNAs (e.g. mammalian cytoplasmic 18S rRNA) should be superimposable on the model without steric clashes occurring within the larger 16S-like RNA.
Rule 9: When adjacent helices are by 0 or 1 residue, they are coaxially stacked unless there is evidence to indicate that they should not be. This rule is based upon coaxial stacking patterns observed in tRNA.
Rule 10: Helices for which there is not sufficient constraint data are not included in the model, but can be accommodated in the model. As a result, the following regions are not included in the model:
• 59-112 (helix 6 and associated single stranded regions)
• 116-239 (helices 7, 8 and 9 and associated single stranded regions) • 404-499 (helices 16 and 17 and associated single stranded regions) • 991-1045 (helix 33 and associated single stranded regions)
• 1297-1339 (helix 42 and associated single stranded regions) • 1346-1383 (helix 43 and associated single stranded regions)
Atmadja, J., R. Brimacombe, H. Blocker and R. Frank (1985). "Investigation of the tertiary folding of Escherichia coli 16S RNA by in situ intra-RNA cross-linking within 30S ribosomal subunits." Nucleic Acids Res 13(19): 6919-36. Atmadja, J., W. Stiege, M. Zobawa, B. Greuer, M. Osswald and R. Brimacombe
(1986). "The tertiary folding of Escherichia coli 16S RNA, as studied by in situ intra-RNA cross-linking of 30S ribosomal subunits with
bis-(2-chloroethyl)-methylamine." Nucleic Acids Res 14(2): 659-73.
Barbieri, M., R. Laschi and C. Rizzoli (1968). "[Electron microscopic studies of crystallized ribosomes]." Boll Soc Ital Biol Sper 44(8): 778-9.
Bogdanov, A. A., I. N. Lavrik, S. S. Dokudovskaia and O. A. Dontsova (1995). "[Structure and function of 5S rRNA in ribosomes]." Mol Biol (Mosk) 29(6): 1218-27.
Brimacombe, R. (1995). "The structure of ribosomal RNA: a three-dimensional jigsaw puzzle." Eur J Biochem 230(2): 365-83.
Capel, M. S., D. M. Engelman, B. R. Freeborn, M. Kjeldgaard, J. A. Langer, V. Ramakrishnan, D. G. Schindler, D. K. Schneider, B. P. Schoenborn, I. Y. Sillers and et al. (1987). "A complete mapping of the proteins in the small ribosomal subunit of Escherichia coli." Science 238(4832): 1403-6.
Capel, M. S. and V. Ramakrishnan (1988). "Neutron-scattering topography of proteins of the small ribosomal subunit." Methods Enzymol 164: 117-31. Dallas, A. and P. B. Moore (1997). "The loop E-loop D region of Escherichia coli
5S rRNA: the solution structure reveals an unusual loop that may be important for binding ribosomal proteins." Structure 5(12): 1639-53. Dontsova, O., S. Dokudovskaya, A. Kopylov, A. Bogdanov, J. Rinke-Appel, N.
Junke and R. Brimacombe (1992). "Three widely separated positions in the 16S RNA lie in or close to the ribosomal decoding region; a site-directed cross-linking study with mRNA analogues." Embo J 11(8): 3105-16.
Expert-Bezancon, A. and P. L. Wollenzien (1985). "Three-dimensional arrangement of the Escherichia coli 16 S ribosomal RNA." J Mol Biol 184(1): 53-66.
Gautheret, D., F. Major and R. Cedergren (1993). "Modeling the
three-dimensional structure of RNA using discrete nucleotide conformational sets." J Mol Biol 229(4): 1049-64.
Gornicki, P., K. Nurse, W. Hellmann, M. Boublik and J. Ofengand (1984). "High resolution localization of the tRNA anticodon interaction site on the Escherichia coli 30 S ribosomal subunit." J Biol Chem 259(16): 10493-8. Greuer, B., M. Osswald, R. Brimacombe and G. Stoffler (1987). "RNA-protein
cross-linking in Escherichia coli 30S ribosomal subunits; determination of sites on 16S RNA that are cross-linked to proteins S3, S4, S7, S9, S10, S11, S17, S18 and S21 by treatment with
bis-(2-chloroethyl)-methylamine." Nucleic Acids Res 15(8): 3241-55.