THE DISTRIBUTION OF CROSSOVERS ALONG UNREPLICATED LAMBDA BACTERIOPHAGE CHROMOSOMES
FRANKLIN W. STAHL, KENNETH D. McMILIN,~ MARY M. STAHL, JEAN M. CRASEMANN AND STEPHEN LAM2
Institute of Molecular Biology, University of Oregon, Eugene, Oregon 97403
Manuscript received December 28, 1973
Present address: Max Plan& Institut fur Molehulare Genetik, Ihnestrasse 63-73, Berlin, West Germany. Present address: Biology Department, University of Utah, Salt Lake City, Utah.
ABSTRACT
The distribution of crossovers along unreplicated chromosomes of bacterio- phage lambda has been examined by determining the density distributions and genotypes of particles in the progenies of crosses of density-labeled by ordinary parents in the presence of genetic blocks to replication. The Red and Rec systems c3,mbined produce crossovers primarily near the ends (especially the right end) of the chromosome. Removal of the generalized lambda recombi- nation functions by red and gam mutations results in loss of these terminal crossovers; coupled with this loss is a disappearance of the differential depen- dence of recombination frequencies in terminal and central intervals on DNA synthesis. Removal of the bacterial system by a r e d mutation results in severe depression of crossing over among unreplicated phage, with the few recombi- nants produced by the lambda system occurring near the right end.
M C M I L I N
andRUSSQ
(1972) showed that crosses could be conducted in phage lambda in the almost complete absence of DNA replication effected by com- bined mutations in the dnaB gene of the bacterial host and either the 0 or P gene of the infecting phages (“double-block” crosses), This technique allowed one to conduct “heavy x light” density-labeled crosses in which the density of each progeny particle is a measure of its material inheritance from each of the two parents(K.
D. MCMILIN, personal communication;WHITE
and Fox 1974; Russo 1973; STAHL et al. 1972a and b).
In STAHL etal.
(197213),
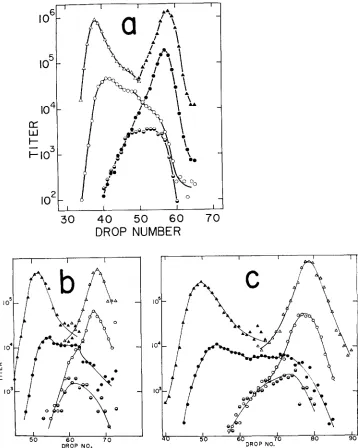
the density distribution of recombinants between near-terminal markers in such a “double-block” cross (a “profile”) was interpreted as a graph of the “cros~over’~ distribution along the unreplicated lambda chromosomes. (See the Introduction in the accompanying paper by MCMILIN et al. (1974) for an explanation of this concept.) Justification for that interpretation lies primarily in the behavior of an unselected plaque- morphology marker in the c l gene (see linkage map in Figure 1 ). In Figure 2a, the distribution of c and C+ markers among recombinant phage of different den-w. al.
A
J
att C IR
I
I
1 I
I I
I I
I
I
FIGURE 1.-The lambda chromosome showing the location of genes and sites of interest to this paper. Locations are taken from the map by SZYBALSKI et al. (1970).
DROP
NUMBER
I I I I I
50 6 0 7 0
CROSSOVER DISTRIBUTION IN LAMBDA 397
ZICHICHI
andEPSTEIN
1962) are found at the density where the c+ phage giveway to the c’s. However, it should be noted that the peak of mottlers is broad, easily reaching the fully light
(L/L)3
position and possibly reaching theH/H
position, too. The shape of the mottler peak implies a large, possibly variable length for overlaps, a condition guaranteed to impose a degree of ambiguity upon the interpretation of our density distributions. The ambiguities will arise pri- marily from the following two sources: (1 ) Heteroduplexes appear to be subject to “mismatch repair”; i.e., they do not invariably segregate both alleles (WHITE and Fox 1974;J.
WILDENBERG and M.MESELSON,
personal communication).(2) Heteroduplexes which are not repaired may have a n artifactual phenotype depending on the markers used and the conditions of assay. Nevertheless, the gross features of our distributions appear interpretable. I n support of this opti- mistic view, we offer a cross like that in Figure 2a but with the coupling of the c marker reversed (Figure 2b). Note that the densities of particles bearing the c ,and c+ markers are reversed, as they should be, and that the heterozygote distri- bution is about the same in the two crosses. A difference in the two sets of dis- tributions can be seen at the point where the c and c+recombinant distributions cross. This may be real; it does occur just where heteroduplexes (and hence “marker effects”) would be most expected to dictate the distributions.
In the present paper we expose the genetic basis for the observed crossover dis- tribution in double-block crosses. Our data are primarily profiles with various components of the recombination systems eliminated by mutation. W e have ex- amined most of the combinations of the following mutations: (1 ) red3: a muta- tion mapping in the lambda redp gene but negative for both beta protein and a Abbreviations used: H/H, L/L, position in an equilibrium density gradient corresponding to the density of lambda particles bearing DN.4 density-labeled on both or neither chain. Also used to refer to the particles themselves; Red, the generalized lambda recombination system; Rec, the coli recombination system which acts on lambda; r e d , one of ihe genetic components of the Rec system; Int, specialized recombination system lambda acting only on a t f ; a f t , locus on the lambda chromosome which is the site of action of Int; Su-, phenotype of a coli strain unable to support the growth of amber ( n m ) mutants of lambda.
FIGURE 2.-The density distributions of phages produced in rec+ red+ gam+ double-block crosses. (a) i n t i cross (From STAHL et al. 1972b, with permission of the National Academy of
Sciences, USA) :
tsA
+
4-
heavy,+ cl tsR light
Key:
A
Total c phage A Total cf phagec tsf recombinants Q c f ts+ recombinants
mottled (c/cf) is+ recombinants
(b) int+ cross:
tsA C l
4-
heavy+
+
tsR lightKey as above.
(c) int- cross. Heavy tsA cl x light tsR. (The number of drops between the H/H and L/L peaks depends on the size of the drops, which vaned from one experiment to another.)
lambda exonuclease, the product of the reda gene
(RADDING
as reported i n SCHUL- MAN et al. 1970); (2) gam210: a suppressible (amber) mutation; (3) int4: a non-conditional mutation which shows none of the interactions withR
mutants characteristic of the int6 mutation which we have used on some previous occa- sions (STAHL and STAHL 1971b) ; (4) the recA56 mutation in our dnaB strainFZ
14, described previously (STAHL et al. 1972b).The data we obtained dictate the manner in which we arrange our results for presentation. The rec+ and rec- results are so different from each other that we separate them into major sections. On the other hand, the int+ and int- results are frequently very similar and in every case show only the differences one would expect from the widely held views of the mechanism of Int action (see GOTTES- MAN and WEISBERG 1971). Therefore, we present the int+ and
inr
crosses side by side where they may be viewed essentially as replicas and used to calibrate reproducibility. Within those two constraints, we present first the crosses with both the red and gam gene products functional, then the red- crosses, then the gam- crosses, and then crosses which are red- gam-. We did perform red- and red- gam- crosses in the rec- host to verify the expectation that no recombillants would be produced in the cl-R interval. The expectations were fulfilled, and the data are not presented. I n the third section, we demonstrate a correlation between the crossover distributions and a differential dependence of crossing over on DNA synthesis in terminal us. central regions of the chromosome.MATERIALS A N D METHODS
Bacterial Sirains: All platings were on strain C600. Host cells were the reef Su- dnaB-
strain FA77 (MCMILIN and Russo 1972) or its recA56 derivative FZ14 ( S T A H L et al., 1972a). Stocks were made, as usual, on K12SH-28.
Phage Strains: The following mutants of phage X were employed:
Mutation Reference
as markers:
tsA14 BROWN and ARBER (1964)
tsR2 BROWN and ARBER (1964)
amJ6 WEIL and SIGNER (1968)
amR5 WEIL and SIGNER (1968)
cl26 MESELSON (1964)
to block replication:
to remove recombination functions:
amp80 C.4MPBELL (1 961)
red3
gam210 ZISSLER et al. (1971)
int4 GINGERY and ECHOLS (1967)
SIGNER and WEIL ( 1968)
Source
W. ARBER W. ARB=
E. SIGNER
E. SIGNER J. WEIGLE
M. MESELSON
E. SIGNER E. SIGNER
E. SIGNER
A map of X showing the location of relevant genes and sites is given in Figure 1.
CROSSOVER DISTRIBUTION I N LAMBDA 3 99
The use of a conditional gam mutation was a valuable aid in the construction of these stocks.
A double mutant red- gam- makes very small plaques under conditions where the mutant pheno- types are expressed. Attempts to make stocks under these conditions frequently fail because of
low titers and a high incidence of particles carrying an extraneous mutation which reverses the small plaque phenotype. This mutation appears to be identical to the mutation described sev- eral years ago by D. HENDERSON (personal communication). HENDERSON'S mutation had the primary phenotype of conferring large plaque size upon Spi- (LINDAHL et al., 1970) deletion mutants, all of which made minute plaques when first isolated. In a companion paper (LAM et al. 1974) we show that the presence of this mutation in red- gam- crosses profoundly alters the distribution of Rec-mediated) recombination along the lambda chromosome. By using the amber gam mutation and the Su+ strain K12SH-28 for making stocks, we obtained lysates which were
OI adequate titer and free of HENDERSON'S mutation.
Labeled stocks: Density-labeled phage stocks were made by techniques described previously (STAHL et al. 197213). Especially difficult stocks were made on plates with IsC-algal hydrolysate (Merck, Sharpe and Dohme) as nutrient source. In most crosses, stocks were not purified before use, but crosses were performed in broth supplemented with ordinary glucose and nucleosides to minimize incorporation of heavy isotopes into progeny particles.
Crosses and Centrifugation techniques have been described previously ( STAHL and STAHL 1971a; MCMILIN and Russo 1972; STAHL et al. 197%).
Plating: The use of BBL-Trypticase plates (STAHL and STAHL 1971a) facilitated scoring of the c marker.
RESULTS
I. recf Profiles
I n all the crosses reported below, one parent is density-labeled and has the genotype tsA cl. The other parent coiitains ordinary isotopes and is genotype tsR. Both parents are blocked for replication by the mutation amP80, and the cross is conducted at restricting temperature (39") in the Su-dnaB- host strain FA77. The lysates from the crosses were centrifuged to equilibrium in a cesium formate gradient, and fractions were collected through a needle hole in the bottom of the tube. Total phage, which were scored with respect to the cl marker when it was easy to do so, were assayed at 30" on the Su+ strain C600. We define as recom- binant those phage which make plaques on C600 at 42". In all crosses, recom- binants were classified by inspection as c, c+ or mottlers (presumably c/c+
heteroduplex particles). Although distinction between c and c+ was rarely in doubt, the scoring of mottlers was occasionally so in some crosses and frequently so in others in which the plaques were small due to the genotypes of the phages (as in the red- crosses described below). In such cases, a given mottler was likely to be scored as c or c+ depending on its predominant component. When inter- preting the data in this and the following section, the reader is asked to recall that phage produced in the absence of replication, whether visibly recombinant o r not, owe their existence as mature infectious particles to the action of the re- combination systems (STAHL et al. 1972a).
400 et
that most of the crossovers resulting in maturable phage occur near the end (s) of the chromosome such that the resulting maturable chromosome has a density only slightly changed from that of one of the parents. (A contrasting result in which recombination is approximately uniformly distributed along the chromo- some can be seen in Figure 3b). I n Figure 2, the LJL peaks are contaminated somewhat with unadsorbed phage particles. I n some of the crosses reported below in which phage yields are small, the extent of contamination is correspondingly greater. (3) The ts+ recombinants are nonuniformly distributed in the gradient indicating a n especially high rate of crossing over just left of R and a somewhat high rate right of A . Thus, some of the exchanges inferred to occur near the chromosome ends do occur between o u r nearly terminal markers i n the
A
andR genes. (4) Among the ts+ recombinants, the c and c+ phage are distributed in the gradient as expected for a break-join mechanism of recombination occurring at a lower rate in the middle of the chromosome than at the ends.
( 5 )
The density distribution of mottled plaques is broad indicating large overlap lengths among the (heteroduplex) recombinants. At n o point on the gradient does the fre- quency of mottlers exceed that of c or of c+ phage. This suggests that one or more of the following situations obtains: (a) mottled plaques are frequently scored mistakenly as c or C+ ; (b) many heteroduplex molecules are “repaired” to homo-duplexes or for other reasons fail to leave two kinds of offspring; (c) in addition to the long overlaps indicated by the breadth of the mottler density distribution, overlaps also occur which, because of short length, are rarely heteroduplex for c/c+. Analogous profiles have been obtained using phage marked with a m muta- tions in the A and R genes. The distributions of crossovers yielding am+ are like those reported here for ts+ recombinants.
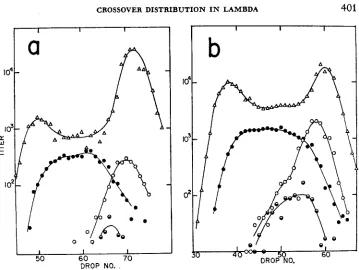
2. rec+red-gam+. In these crosses (Figure 3a and 3b) both parents were red-; markers and conditions of the cross are otherwise the same as in the crosses de- scribed above. The following features of the distributions should be noted: (I)
A
large fraction of the total phage are distributed in regions of the gradient between the two extreme positions. (In Figure 3a, the height of the
L/L
peak is probably due in part to unadsorbed parental phage in that peak.) (2) I n agreement with the total phage distribution, the ts+ recombinants are approximately uniformly distributed suggesting an approximately constant rate of recombination along the chromosome. (3) Int has a detectable, but slight, influence on the recombinant distribution, apparently contributing ts+ recombinants slightly lighter than half heavy as expected from the location of its site of action ( a t t ) slightly to the right of middle of the lambda chromosome (see Figure 1). (4) c/cf heterozygotes are present and rather broadly distributed as in the cross above.W e may conclude that removal of red function reduces the high rates of cross- ing over a t the chromosome ends (relative to the middle) which characterize the rec + red + gam + crosses.
CROSSOVER DISTRIBUTION IN LAMBDA 401
IO
IC
E
W
I-
k
IC
,
, 1 I I 1
50 60 70
DROP NO..
b
A
o f
.
0FIGURE 3.-The density distributions of phages produced in ret+ red- gam+ double-block crosses-heavy tsA c l crossed with light tsR. (a) i n t f , (b) int-. (From STAHL et al. 1973, with permission of Academic Press.)
A Total phage c tsf recombinants @ c+ ts+ recombinants
mottled (c/c+) ts+ recombinants
binant phage reflect a relative reduction in the rate of crossing over near the ends of the chromosome. The reduction at the right end is less severe than that ob- served in the red- crosses above; the reduction at the left end appcars to be as great as that in the red- crosses.
F. w. STAHL et al.
I
50 60 DROP NO.
,
-
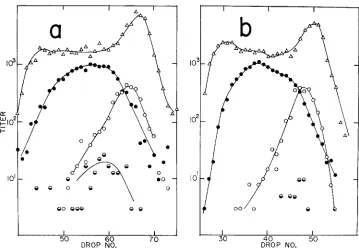
40FIGURE 4.-The density distributions of phages produced i n r e d red+ gam- double-block crosses-heavy tsA cl X light tsR. (a) int+, (b) inr. Key as in Figure 3.
,
50 60 7 0DROP NO.
10:
I 0'
I 10
I I I
I I I
30 40 50
DROP NO.
CROSSOVER D I S T R I B U T I O N IN L A M B D A 403
11. rec- Profiles
The following crosses were conducted in FZ14, a recA mutant derivative of the dnaB mutant strain used in the crosses above.
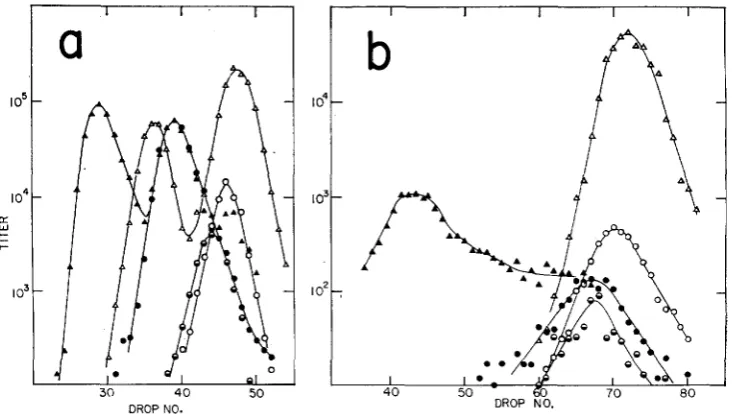
1. rec-red+gam+. Figures 6a and b reveal that removal of the recA function severely depresses generalized recombination over the entire length of the bac- terial chromosome. This conclusion is indicated (weakly) by the low yields of phage obtained in recA- crosses under conditions of blocked
DNA
duplication and(strongly) by the relatively large ikaction of the yield which results from recom- bination by the Int system. (The low yields are not directly reflected in the titers shown in the figures. For data relating yields OI unduplicated phage to the pres- ence of the different recombination systems, see STAHL et al. 1972a.) The follow- ing features of the density distributions should be noted: (1) The int+ and int- distributions are profoundly different indicating that a large fraction of the phage produced results from action of the Int system. (2) I n the i n t cross it can be seen that recombinants are produced only at the right end of the chromosome. ( 3 )
Heterozygotes in the int- cross are a large fraction of all particles bearing the c marker. The constancy of this fraction along the gradient suggests that all of the particles bearing the c marker may be overlapped at the c l locus.
(4)
The heights of the light peaks indicate heavy contamination with unadsorbed phage.2. rec-red+gam-. Removal o f the gam function (Figures 7a and b) reduces the amount of generalized recombination; the phage yield is so low as a result that little can be said except that the few recombinants found arise by crossing over near the right end of the chromosome.
/a'
I I I IDROP NO.
w.
b
A
,
,
“no
80 90
DROP NO. 70
FIGURE 7.-The density distributions af phages produced in rec- red+ g a n - double-block crosses-heavy tsA c l x light tsR. (a) int+, (b) i n t . Key as in Figure 3.
111. recf crosses with some DNA synthesis allowed
STAHL and STAHL (1971a and b) demonstrated that DNA synthesis is associ- ated to a variable degree
with
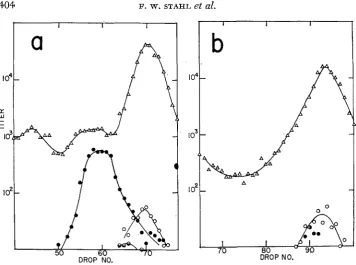
recombination in different regions of the lambda chromosome. The differential association of synthesis with different regions was shown ( 1971 b) to characterize recombination when all recombination functions were present, to characterize Red recombination in r e d - cells, and, to a lesser extent. to be true of Rec recombination in re& infections. This section extends those observatiom and reveals an involvement of the gam gene product. The ef- fects observed here correlate with those of the preceding recf section. Combina- tions of recombination mutations which repress the frequency of crossing over near termini in DNA-minus crosses reduce the differential association of DNA synthesis with different regions of the chromosome.The crosses in this section differ in three respects from those described in sec- tion 1. (1 ) Infecting phages carry no genetic block for DNA synthesis, (2) the marked intervals are J-cZ and cl-R, and (3) both parents are heavy. The conse- quence of the first difference is that the amount of DNA synthesis manifest by the progenies of the crosses can be adjusted by controlling the temperature at which the infection is made in the temperature sensitive dnaB- strain FA77. The second and third changes permit us to compare the amounts of DNA synthesis associated with recombination in a pair of intervals whose behavior in this regard is sharply contrasting.
CROSSOVER DISTRIBUTION IN LAMBDA 405
and the host was rec+. In the first cross, the lambda generalized recombination functions were intact; in the second, the infecting phage was red-; in the third the phage was both red- and gam-. This progression resulted in a progressive loss of the difference in amount of
DNA
synthesis associated with recombination i n the two marked intervals, J-cZ andCZ-R
(Figure 8). This loss of difference is revealed as a convergence of the mean densities of recombinants in the two intervals.DISCUSSION
In previous reports we have shown that in phage lambda, recombination, repli- cation, and maturation of chromosomes are interdependent processes. TO be encapsidated, a chromosome must either replicate more than once, or recombine
(STAHL
et al. 1972a). Any of the three recombination systems, Red, Rec, or Int, can promote maturation. The production of most Red-mediated recombinant phage is associated with extensive DNA synthesis(STAHL
and STAHL 1971b; STAHL et al. 1972a and b).
The production of recombinants by the Rec system is associated with less DNA synthesis (STAHL et al. 1972a and 1973). I n the present paper we demonstrate that in the absence of replication, the Rec system (in red-crosses) produces recombinants at a higher rate than does the Red system (in recA- crosses). This result is harmonious with the conclusion (STAHL et al, 197213) that (Red) recombination-associated synthesis is essential for recom- binant production.
Under conditions of blocked replication we have determined the density dis- tributions of wild-type recombinants arising in Heavy
x
Light density-labeled crosses between phages bearing terminal t s markers. The behavior of a non- selected plaque morphology marker (c) justified the view that the density dis- tributions observed are appropriate indicators of the distribution of crossovers along the length of the lambda chromosome, with a n appreciable ambiguity rricing from long heterozygous overlaps,W e reached the following conclusions with respect to the distribution of cross- overs along unduplicated chromosomes: (1) I n recA- cells, the Red system pro- duces few recombinants, and the few produced are due to crossovers near the right end. Elimination of gamma function further reduces the number of crossovers. (2) In rec+ cells, the Ked system produces many recombinants, and most of those are produced by crossovers near the right end, while some are due to crossovers near the left end. The number of Red-dependent crossovers in a recf cell is far greater than in a recA- cell suggesting a n interaction between the two generalized recombination systems. (3) I n recf cells, elimination of either gam function or red function reduces the rate of crossing over near the ends. Removal of both the g a m and red functions has cumulative consequences.
406
w.
et al. 0 r- n a 0 D 0 U 3 0 d4 ? %
2 . e c a . a 0
!7 '3
s
g . 2.e Z i E 4 4 4
b o * &
m Z &
.F
k
3
i
2
s
43"m
L32
.-
$ + Z
,.
::
2 . 1 4 E
2
g z : %
B.B
2
2 %
!3
z
7: 5
52
E L P & c
3
.E: (I) 20 a, E i 3
k 2 2 E
.3g:
a h
p
; % U ,
,^s 4-
04
Q
-
5
.,o
6
22
.r2 2 2 1
-a
..
S E ,3 8 8
e
p &
8
E = &5 z m . 2
:z
01"
'x""&J$i:
# + k % m ,o m s
$ E < h % : & -
a, Q h G u t . + %
- 2 +
E $
& x . 5 5
-
a y d E u . zE+ % m U 0 0 u +
.-
J B e C h F u u
8
2
& r p d 00;h . & Z r ? g m
E
v) c
m
c
v)
>-a',p
% x. + ..
g e - 3 -
E:
L P
h f , U a,.- 3
9 . 3 h w a,
2
p,-g
4
bt
CROSSOVER DISTRIBUTION I N LAMBDA
407
had enjoyed extensive DNA synthesis. A conclusion of this paper is that the Rec system in the absence o€ the lambda functions red and g a m displays none of the regional differences in recombination-associated DNA synthesis charactcristic of infections by red+gam+ phage. However, the Rec system in the presence of the g a m function does show regional differences, albeit slight, in the amount of syn- thesis associated with recombination. This behavior correlates with the distribu- tion of crossovers along unreplicated chromosomes. The loss of the terminal con- centration of crossovers is accompanied by loss of regional differences in amounts of DNA synthesis associated with recombination. The set of facts described here- in begs a model. In a subsequent communication we will propose a modification of one previously put forth ( STAHL et al. 1973).
DAVID HENDERSON’S generous communication of unpublished observations saved us from disaster. JEFF HAEMER provided valuable criticism of the manuscript.
This work was supported by Research grant number GB 8109 from the National Science Foundation and by Program Project grant number GM15423 (AARON NOVICK, Project Director) from the National Institutes of Health. K.D.M. is a Predoctoral Trainee on Training grant number GMOQ715 from the National Institutes of Health.
LITERATURE CITED
BROWN, A. and W. ARBER, 1964
CAMPBELL, A., 1961
GINGERY, R. and H. ECHOLS, 1967 Mutants of bacteriophage lambda unable to integrate into the host chromosome. Proc. Natl. Acad. Sci. U.S. 58: 1507-1514.
GOTTESMAN, M. E. and R. A. WEISBERG, 1971 Prophage insertion and excision. pp. 113-138. In: The Bacteriophage Lambda. Edited by A. D. HERSHEY. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
KELLENBERGER, G., M. L. ZICHICHI and H. T. EPSTEIN, 1962 Heterozygosis and recombination of bacteriophage. Virology 17: 44-45.
LINDAHL, G., G. SIRONI, H. BIALY and R. CALENDAR, 1970 Bacteriophage lambda; abortive in- fection of bacteria lysogenic for phage P2. Proc. Natl. Acad. Sci. U.S. 66: 587-594. MCMILIN, K. D. and V. E. A. Russo, 1972 Maturation and recombination of bacteriophage
lambda DNA molecules in the absence of DNA duplication. J. Mol. Biol. 68: 59-65. MCMILAN, K. D., M. M. STAHL and F. W. STAHL, 1974 Rec-mediated recombinational hot spot
activity in bacteriophage lambda. I. Hot spot activity associated with spi- deletions and bio substitutions. Genetics 77: 409-423.
On the mechanism of genetic recombination between DNA molecules. J. Mol. Biol. 9: 734-745. ~ , 1967 TLe molecular basis of genetic recombination.
pp. 61-104. In: Heritage from Mendel, Edited by R. A. BRINK. The Umversity of Wisconsin Press, Madism, Wisconsin.
On the physical structure of lambda recombinant DNA. Molec. Gen. Genet. 122 : 353-366.
Properties of recombination- Temperature-sensitive mutants of coliphage lambda. Virology
24: 237-239.
Sensitive mutants of bacteriophage lambda. Virology 14: 22-32.
MESELSON, M., 1964
Russo, V. E. A., 1973
SHULMAN, M., L. M. HALLICK, H. ECHOLS and E. R. SIGNER, 1970
SIGNER, E. and J. WEIL, 1968
deficient mutants of bacteriophage lambda. J. Mol. Biol. 52: 501-520.
general recombination. J. Mol. Biol. 34: 261-271.
408
STAHL, M. M. and F. W. STAHL, 1971a
w. et al.
DNA synthesis associated with recombination. I. Re- combination in a DNA-negative host. pp. 431-442. In: The Bacteriophage Lambda. Edited by A. D. HERSHEY. Cold Spring Harbor, New York.
STAHL, F. W. and M. M. STAHL, 1971b DNA synthesis associated with recombination. 11. Re- combination between repressed chromosomes. pp. 443-453. In: The Bacteriophage Lambda. Edited by A. D. HERSHEY. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York. STAHL, F. W., K. D. MCMILIN, M. M. STAHL, R. E. MALONE, Y. Nozu and V. E. A. Russo, 1972a A role for recombination in the production of “free-loader” lambda bacteriophage particles. J. Mol. Biol. 68: 57-67.
An enhancing role for DNA synthesis in formation of bacteriophage lambda recombinants. Proc. Natl. Acad. Sci. US.
69: 3598-3601.
STAHL, F. W., S. CHUNG, J. CRASEMANN, D. FAULDS, J. &EMER, S. LAM, R. E. MALONE, K. D.
MCMILIN, Y. Nozu, J. SIEGEL, J. STRATHERN and M. STAHL, 1973 Recombination, replica- tion, and maturation in phage lambda. pp. 487-503. In: Virus Research. Edited by C. FRED
Fox and W. S. ROBINSON. Academic Press, New York.
SZYBALSKI, W., K. BE”, M. FIANDT, S. HAYES, Z. HRADECNA, S. KIMAR, H. A. LOZERON, H. J. J. NIJKAMP and W. STEVENS, 1970 Transcriptional units and their controls in ESCheriChiQ coli phage lambda: operons and scriptons. Cold Spring Harbor Symp. Quant. Biol. 35: 341- 353.
Recombination in bacteriophage 1. 11. Site-specific recombina- tion promoted by the integration system. J. Mol. Biol. 34: 273-279.
On the molecular basis of high negative interference. Proc. Natl. Acad. Sci. US. (In press.)
The role of recombination in growth of bacterio- phage lambda. I. The gamma gene. pp. 455-4433. In: The Bacteriophage Lambda. Edited by A. D. HERSHEY. Cold Spring-Harbor Laboratory.
Corresponding editor: D. KAISER STAHL, F. W., K. D. MCMILIN, M. M. STAHL and Y. Nozu, 1972b
WEIL, J. and E. R. SIGNER, 1968
WHITE, R. L. and M. S. Fox, 1974