Full Terms & Conditions of access and use can be found at
https://www.tandfonline.com/action/journalInformation?journalCode=yhem20
Hematology
ISSN: (Print) 1607-8454 (Online) Journal homepage: https://www.tandfonline.com/loi/yhem20
Recent thymic output function in patients with
hematological malignancy
Yangqiu Li
To cite this article: Yangqiu Li (2005) Recent thymic output function in patients with hematological malignancy, Hematology, 10:4, 297-305, DOI: 10.1080/10245330500093591
To link to this article: https://doi.org/10.1080/10245330500093591
Published online: 04 Sep 2013.
Submit your article to this journal
Article views: 62
View related articles
MALIGNANCY
Recent thymic output function in patients with hematological
malignancy
YANGQIU LI
Institute of Hematology, Medical College, Jinan University, Guangzhou, 510632, China
(Received 10 January 2005; revised 30 January 2005)
Abstract
Thymic function is important for the generation of T-cell diversity in the periphery of both children and adults during both health and disease. Until recently, thymic function could not be monitored, as a consequence of the absence of adequate technology to differentiate recent thymic emigrants (RTEs) from naı¨ve T cells. The generation of TCR diversity occurs in the thymus through recombination of gene segments encoding the variable parts of the TCRa and bchains. During these processes, by-products of the rearrangements are generated in the form of signal joint T-cell receptor excision circles (sjTRECs). As sjTRECs are stable extrachromosomal DNA fragments, they are not replicated during mitosis and thus diluted with each round of cell division, and are therefore most frequent in naı¨ve T cells that have recently left the thymus, their quantification is actually considered as a valuable tool to estimate thymic function. Therefore, quantitative sjTRECs content have been recently used to assess thymic output during both health and disease. In this review, we summarize recent data on the recent thymic output function feature in patients with hematological malignancy and the immune reconstitution after stem cell transplantation and we also characterize factors that may improve the thymic output function.
Keywords: TREC, thymic output function, naı¨ve T cells, leukemia, stem cell transplantation
Introduction
The thymus is considered to atrophy in age with a decline in thymic function. However, this generality is not universally and incontrovertibly true. Many studies performed in animals and humans have proved to the contrary that thymic activity and function appear to be well maintained in the old age and may be indispensable for T-cell reconstitution in different immunological settings. But it has been quite difficult to determine the thymic output function due to lacking of a direct marker for recent thymic emigrants (RTEs), until in 1998, Douek et al. first reported the use of signal joint T-cell receptor excision circles (sjTRECs) to study changes in the frequency of RTEs with age and in the case of human immunodeficiency virus (HIV) 1 infection [1]. This study drew attention to the fact that the level of sjTRECs can be used to
estimate the recent thymic output function. There-fore, sjTRECs was used as a new marker for analysis of thymic output function in different immunodeficiency diseases, cancer patients, patients with leukemia and the immune reconstitution after stem cell transplan-tation (SCT) [1 – 2].
This review summarizes the recent data on the human thymus recent output function and its application in evaluation of thymic function in patients with hematological malignancy, after SCT.
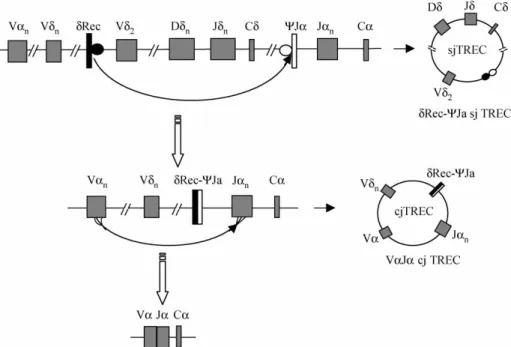
T-cell receptor rearrangements and TCRdgene deletion in thymus
T-cell differentiation in the thymus is characterized by a hierarchical order of rearrangement steps in the T-cell receptor (TCR) genes, resulting in the joining of V, D and J gene segments. During each of
ISSN 1024-5332 print/ISSN 1607-8454 onlineq2005 Taylor & Francis Group Ltd
DOI: 10.1080/10245330500093591
Correspondence: Institute of Hematology, Medical College, Jinan University, Guangzhou 510632, China. Tel: 86 20 85220262. Fax: 86 20 85221343E-mail: [email protected]
the rearrangement steps, DNA fragments between rearranging variable (V), diversity (D, only inbandd), and junctional (J) gene segments are deleted as circular excision products (by products), the so-called sjTRECs. AllV,DandJgene segments are flanked by specific homologous recombination signal sequences (RSS). TCR d gene segments are interspersed with TCRa gene segments along chromosome 14q11 (TCR ad locus), during the process of TCRa
rearrangement, it must delete at first the TCRd
gene, which is an important step in thymic T-cell maturation for TCRablineage. In 1988, DeVillartay et al. identified two so-called TCRddeleting elements,
dRec andcJa, that flank the major part of the TCRd
gene (Figure 1) [1 – 5]. ThedRec andcJa recombina-tion produces a dRec –cJa signal joint TRECs, sjTRECs are assumed to have a high over-time stability, but they can not multiply and consequently, are diluted during T-cell proliferation. A maximum of two sjTRECs can be present within oneab T-cell if the corresponding rearrangement event occurs in both alleles and if the cell does not proliferate following the rearrangement. sjTRECs are exported from thymus to the periphery within RTEs, therefore, the frequency of sjTRECs is considered to be the most accurate marker of T-cell neogenesis, and quantification of dRec –cJa
sjTRECs content in peripheral blood leads to an underestimation of the real frequency of RTEs. It provides an unequivocal way to estimate the RTEs. Although it was reported that the TCRd
locus could be excised through other recombination events that will not generate a sjTREC, at least more
than 70% of the TCRddeletion are thoughdRec –cJa
rearrangement. Since 1998 quantitative detection of sjTRECs has been used for direct measurement of thymic output [1].
This new method was supported by several studies. For example, it was demonstrated that thymectomy during cardiac surgery or for treatment of myasthenia gravis led to a significant decrease in sjTRECs content of purified CD4þand CD8þT cells [1]. Conversely, after successful transplantation of thymic tissue, increasing sjTRECs content with time was found in patients with congenital absence of thymic tissue (complete DiGeorge syndrome) and absence of sjTRECs [6]. Also in pediatric patients with severe combined immunodeficiency who were treated with SCT, sjTRECs were detected as early as one month following transplantation [7].
Quantitative detection of sjTREC in peripheral blood
Several methods have been used to measure thymo-poietic capacity. Thymic size as measured by volumetric computerized tomographic measurement has been correlated with the numbers of CD4 þ-CD45RAþ naı¨ve T cells. The number of phenotypi-cally naı¨ve T cells has been shown to correlate with antigen-specific function. However, there are con-cerns about the limitations of estimating thymic function on the basis of naı¨ve T-cell phenotype alone. T cells expressing a naı¨ve phenotype are not necessarily accurate surrogate markers of thymic Figure 1. Schematic model of recombination processes leading to sjTREC formation. The TCRdlocus is located within the TCRalocus. The major part of TCRdlocus is flanked by two gene segmentsdRec andcJa. During the rearrangement of TCRagene segments, thedRec andcJarearrange to each other and form an extrachromosomal DNA circles, referred to as signal joint T cell receptor excision DNA circles (sjTRECs) (top). The second TRECs, coding joint TRECs forms during the rearrangement of Va– Ja(bottom) [1,2,5].
Y. Li
function. Following thymic emigration, CD45RAþ naı¨ve T cells can have a long quiescent life span, may proliferate in an antigen-independent manner, or may rapidly convert to CD45ROþ memory/effector phenotype T cells. Furthermore, naı¨ve T-cell markers may be acquired by memory T cells. Both naı¨ve and memory cells may have overlapping phenotypes, especially among CD8þT cells [8].
A different molecular biological assay that has been more recently used as a measure of thymopoietic capacity is the frequency of sjTRECs among peripheral blood T cells. Because sjTRECs are circular, stable extrachromosomal DNA fragments, are not replicated during mitosis and are thus diluted with each round of cell division, and are therefore most frequent in naı¨ve T cells that have recently left the thymus, and would not have been affected by peripheral expansion and cellular replication. It has been generally used as an indicator for RTEs in assessing human thymic function [1,2,8].
For evaluating the sjTRECs level, a real-time quantitative PCR (RQ-PCR) to detect and quantify
dRec –cJa sjTRECs in peripheral blood T cells was established. The RQ-PCR proved to be sensitive with a detection level of approximately one sjTREC copy in 100 ng of DNA [9]. In defining the sjTREC level, one can report the number of sjTRECs per 103– 105cells or per 1mg DNA to compare its values [1].
For RQ-PCR, different techniques may be used, both TaqMan and LightCycler techniques which are based on the hybridization of a dual-fluorescence-labelled probe to the PCR product are employed by most research groups. They are sensitive, rapid and easy to perform. The PCR-ELISA technique that relies on biotin labeling was reported to be useful as a sensitive method for the quantitative detection of sjTRECs, but it is not a true real-time detection method [1,5].
Samples used for sjTRECs detection may be collected from thymic cells, cord blood, peripheral blood mononuclear cells (PBMNCs) and magnetic associated cell sorter (MACS)-sorted cells including CD3þ, CD4þ, CD8þand CD45RAþ cells [10 – 12]. Some results showed that no significant difference was found in individuals when unfractionated PBMCs or MACS-sorted CD45RAþT cells were analyzed for sjTREC counts. However, by analysis of samples from leukemia patients whose peripheral mononuclear cells contain different percentages of leukemic cells and may affect the correct evaluation of sjTRECs levels should be detected the pure T cells (CD3þ, CD4þ, CD8þand others), and it may reflect the true numbers of RTEs in patients with leukemia [10,11].
Recent thymic output function in healthy individuals
The thymus undergoes age-associated involution, with thymic size decreasing from birth at a rate of
approximately 3% per year until middle age, and at a rate of 1% per year thereafter. Age-associated thymic atrophy results in a decline in T lymphocyte output and has been identified as one of the key events that precede inefficient functioning of the immune system in later life [1,2,4]. Soon after it was proposed to use sjTRECs as a measure of thymic output, a series of papers followed, taking sjTRECs analysis as quanti-tative marker for thymus output of RTEs. sjTREC have been detected in individuals over 60 years of age, suggesting continuous thymic output even later in life. Indeed, it was shown that the sjTREC content of PBMNCs, (sorted CD4þor CD8þT cells) is highest in healthy children and declines with age. Some reports showed that the range of number of sjTRECs in healthy individuals was different at varying age ranges, for example, 483^108 sjTRECs copies/105 cells in 1 – 10 year group, 354^77 sjTRECs copies/105 cells in 11 – 40 year group, 65^27 sjTRECs copies/105cells in 41 – 60 year group, 20^ 5 sjTRECs copies/105 cells in 61 – 80þ year group [13]. A number of studies have shown similar results with thymic output function showing an age-dependent decline. During aging, thymic involution primarily induces an age-dependent decline in sjTREC concentrations within both CD4þ and CD8þT-cell populations. Steffens et al. reported the analysis of sjTREC in PBMCs of 148 healthy children and in PBMCs, CD4þ, and CD8þfractions of healthy adults, respectively. It was demonstrated that during the first 5 years of life, thymic output is decreased, but not dramatically. Among adults aged 23 – 58, thymic output was inversely correlated with age. A strong correlation existed between pediatric PBMC sjTRECs and the expression of three naı¨ve phenotypic markers (CD45RAþCD45RO2, CD45RAþCD62Lþ and CD45RO2CD27þCD95low). Adult PBMC sjTRECs correlated only with the expression of CD45RA þ-CD45RO2[14,15].
Using a mathematical model, Ye et al. quantified age-dependent changes both in the number of RTE generated per day and in sjTRECs concentration during an 80-year lifespan. Through their analyses, they demonstrated that RTE and peripheral T-cell division have the same potential to affect sjTRECs concentration at any age in healthy people. T-cell death also influences sjTRECs concentration, but to a lesser extent [15].
The decline in the number of RTEs in blood with increasing age is gender-linked. The number of TRECs was higher among healthy girls than among healthy boys [16]. Peripheral blood from females contained significantly higher levels of sjTRECs within CD3þT cells than blood from males, despite there being no significant gender difference in the absolute number of CD3þT cells in the populations analyzed. The findings suggest better thymic function in females compared with males, providing females
with a higher number of RTEs for longer periods of life. Such a finding provides a plausible explanation for the immunological gender differences observed in previous studies and possibly, for the general longer life expectancy in females compared with males [13].
Recent thymic output function in patients with leukemias
It is a common feature that leukemia patients suffer from cellular immunodeficiency which may play a role in initiation of hematological malignancies. Under-standing the mechanisms underlying persistent immunity in leukemia patients might suggest treat-ment strategies to enhance immune competence in all. Research on the TCR Vb repertoire showed that skewed expression of TCR Vb subfamilies is a common feature in patients with leukemia [17]. However, little is known about the state of thymic recent output function in these patients. The first report was described by Petridou et al., who compared the sjTREC values between patients with childhood B-ALL or T-ALL. The analysis was performed by detection of the sjTRECs 207 cases with childhood leukemia and 90 cases with childhood lymphoma. There was a highly significant reduction of sjTREC values in children with T-cell malignancies (median 3100 sjTRECs copies/106PBMCs), whereas children with B-cell origin ALL had slightly but not significantly lower sjTRECs values compared to healthy children (medians 19,300 and 22,500 sjTRECs/106 PBMCs, respectively) [16]. Another description of the sjTRECs level in ALL patients is from Haining et al., who reported that consistent with the reduction in naı¨ve T cells, thymopoiesis (measured by sjTRECs levels) was significantly lower in 73 children with ALL than in normal controls (an age-matched cohort of 805 healthy children) [18].
Our previous research on analysis of sjTRECs-content showed a dramatic reduction of sjTRECs values in patients with AML (including M2, M3, M4, M5 and M6 subtypes). In 4 out of 13 cases, no sjTRECs copies could be detected in 40,000 T cells. The mean value of sjTRECs was 0:814^0:410 copies/1000 T cells in AML patients and 6:837^ 1:571 copies/1000 T cells in normal individuals, respectively [10]. Similar results were found in patients with CML. The mean value of sjTRECs was 0:06^0:16 copies/1000 T cells in CML patients. In 16 out of 20 CML cases, no sjTRECs copies could be detected in peripheral blood, and lower value of sjTRECs could be identified in sorted CD4þor CD8þ cells [11]. These results demonstrate low numbers of T cells with recent recombination of their TCR gene. However, whether this is due to clonal expansion of T cells to antigens, for example, leukemia associated antigens, or reflects impairment of immune function
associated to the malignancy, remains an open question [17].
The number of naı¨ve T lymphocytes was also markedly suppressed in patients with ATL and human T-cell leukemia virus type I (HTLV-I) carriers, in which the immunodeficiency was a common feature, the low number of naı¨ve T lymphocytes was due to suppressed production of T lymphocytes in the thymus, which might account for immunodeficiency observed in HTLV-I-infected individuals [19].
Although the number of sjTRECs was higher among healthy girls than among healthy boys, a similar pattern was evident in T-cell malignancies [13]. The thymic recent output function was decreased in almost all patients with hematological malignancy, but the individual differences in thymic output of naı¨ve T cells were marked. The research from Svaldi et al. showed a high variation in sjTRECs levels was found at diagnosis (median TREC level 136/105 PBMCs; range 1 – 1729) in 25 cases with myeloma [20]. The lower sjTREC level may relate to the poor prognosis, as Petridou et al. showed that during a median follow-up period of about 19 months, only 4 children died. All of them had a T-cell hematopoietic malignancy and relatively low sjTRECs values. And it appears that there is a pattern of concordance of high sjTRECs values with better disease prognosis in hematological childhood malig-nancies. These findings may be used as hypothesis-generating indications that should be confirmed in larger data sets [16].
The significance of measurement of thymic recent output function in patients with stem cell transplantation
Successful allogeneic hematopoietic stem cell trans-plantation (allo-HSCT) requires reconstitution of normal T-cell immunity. Recipient thymic activity, biologic features of the allograft, and preparative regimens all contribute to immune reconstitution. Measurement of sjTRECs in peripheral blood is a means of quantifying recent thymic T-cell production and has been used after transplantation in many studies to estimate thymus-dependent T-cell recon-stitution. Following HSCT, there is a prolonged period of profound posttransplantation immune deficiency. Recovery of the immune function after SCT depends on 2 mechanisms, peripheral expansion of T cells and naı¨ve T cells production by thymus. The peripheral expansion of T cells may relate to graft-vs-host disease (GVHD), and the thymic-dependent pathways of immune reconstitution may indicate the true recovery of T cells neogenesis and may be used as a marker for predictor of prognosis.
Although phenotypically mature T cells had recovered by 1 – 2 months after bone marrow transplantation (BMT), sjTRECs levels remained
Y. Li
low for 3 months after BMT. The duration of immunodeficiency following marrow transplantation is not yet clear. T-cell neogenesis became evident by 6 months, and normal levels of adult thymic function were restored at 6 – 12 months after BMT [21]. The immunity of patients surviving 20 – 30 years after transplantation is normal or near normal [22]. Low sjTRECs values are significantly associated with morbidity and mortality after transplantation [23]. The extent and rapidity with which T cells are regenerated from graft-derived precursor cells directly influences the incidence of infection and the T-cell-based graft-versus-tumor effect. GVHD and age were the most important predictors of low sjTRECs levels after HSCT, but the administration of standard immunosuppressive medications did not affect sjTRECs levels [8]. Some factors affected thymic function after HSCT were identified. Increasing patient age, chronic GVHD and T-cell depletion predicted low sjTRECs levels and slow naı¨ve T-cell recovery [24].
The quality of thymic function before transplantation
The quality of thymic function before transplantation affects thymus-dependent T-cell reconstitution after transplantation. Healthy donors were found to contain 1200 – 155,000 sjTRECs copies/ml blood, Chen et al. found patients who had more than 1200 copies of sjTRECs per ml of blood before allo-HSCT showed early recovery of sjTREC numbers and TCRb
repertoire diversity. In contrast, patients who had fewer than 1200 sjTRECs copies/ml blood before transplantation demonstrated significantly slower restoration of thymus-dependent T cells. Similar results were found in myeloma patients after autologous peripheral blood stem cell transplantation (auto-PBSCT). Patients with more than 136 sjTRECs/105 PBMCs at diagnosis had a statistically significant better overall survival and event-free survival, whereas low TREC levels correlated with a higher incidence of infectious complications after auto-PBSCT. Indicating that the rate of reconstitution of thymus-dependent T cells is dependent on the competence of thymic function in the recipients before transplantation [20,25]. These data support the concept that pre-transplant measurement of sjTREC may provide important information for predicting thymus-dependent T-cell reconstitution after trans-plantation, both in allo-HSCT and auto-PBSCT.
sjTRECs levels in patients with GVHD: Thymopoiesis is inhibited by GVHD
Allo-HSCT leads to a prolonged state of immuno-deficiency characterized by low peripheral naı¨ve T-cell counts. Many research groups have measured quantitative and qualitative thymic function, TCR
spectratyping and the numbers of phenotypically naı¨ve T cells or sjTRECs level and concluded that active chronic GVHD is the important factor in thymopoiesis.
Efficient reconstitution of the peripheral T-cell pool after HSCT is dependent on normal thymic function. However, the development of GVHD in the context of allo-HSCT is associated with injurious effects on thymocyte development. Patients with a history of resolved GVHD had decreased numbers of sjTRECs compared with those with no GVHD [8]. Active chronic GVHD was the most important factor that predicted low sjTRECs levels even years after HSCT. Age did not affect sjTRECs levels in patients with active chronic GVDH all ages [8]. sjTREC levels were significantly lower among both the CD4þand CD8þ T cells in the active chronic GVHD groups. Weinberg’s group showed that sjTREC levels were typically lower than 2000 sjTRECs per 105 T cells [8,23]. Poulin et al. analyzed quantitative and qualitative thymic function through quantification of sjTRECs frequencies (both the sjTRECs and six different Db– JbTRECs, respectively) in a cohort of patients sampled from 1 to 10 years following allo-HSCT and showed that reduced thymic function was associated only with ongoing clinical chronic GVHD in this cohort. Nonetheless, the diversity of thymic production remained unchanged irrespective of the patient’s chronic GVHD status. The data indicate that the inability to reconstitute the naı¨ve T-cell compart-ment for several years after allo-HSCT, in the absence of chronic GVHD, is a consequence of impaired naı¨ve T-cell survival rather than thymic dysfunction [26].
Comparison of the ability of immune reconstitution after transplantation from different source stem cells
The quantitative analysis of sjTRECs may be used as an important factor to compare the ability of immune reconstitution after different sources of SCT. For example, Talvensaari et al. compared the ability of immune reconstitution after SCT with cord blood (CB) stem cells and bone marrow stem cells. CB is used increasingly as a source of hematopoietic stem cells because of a lower risk of acute and chronic GVHD. However, there is some concern regarding the ability to adequately reconstitute host immune response due to the immaturity and naivety of CB T cells. During the first year after CB transplantation for high-risk hematological disorders, TCR reper-toires were highly abnormal and sjTRECs values low in both groups. Notably, from 2 years after transplantation onward, sjTRECs values as well as TCR diversity were higher in CB recipients than in recipients of bone marrow transplants. These data indicate an efficient thymic regeneration pathway from CB lymphoid progenitors despite the low number of cells infused compared to bone marrow,
arguing for a complete clinical immune recovery after CB transplantation [27].
Comparison of the ability of immune reconstitution in different types of SCT
The quantitative analysis of sjTRECs contents is also used as a common mark for evaluation of immune reconstitution after different type of SCT. Adult patients receiving auto-PBSCT following myelo-ablative chemotherapy could rapidly reconstitute normal sjTRECs content [28]. The production of substantial numbers of new naı¨ve T cells by the thymus could be detected by 100 days post-transplant. In comparison to the CD34-selected group, patients who received unselected PBSCs showed a delay T-cell reconstitution, in the CD34-unselected group, numbers of sjTRECs returned to baseline after 2 years, whereas in the CD34-selected group, numbers at 2 years were significantly higher than both baseline numbers, and 2-year numbers in the unselected group [28,29].
In comparison to myeloablative regimens, patients who underwent non-myeloablative transplantation showed increased recovery of thymic output function. But the T cells recovery after non-myeloablative allo-PBSCT (NST) was shown at different time point from different research groups. Some groups reported that sjTRECs could be detected as early as at day 45 after NST [30], whereas another groups showed that sjTRECs were detected at 12 months after NST [31]. This difference may relate to the source of naı¨ve T cells, and whether they are donor or recipient origin. Generally, thymic-dependent pathways were activated between 6 and 12 months in the majority of patients after allo-HSCT with naı¨ve T-cell numbers increasing in parallel with sjTRECs levels [24]. Chao et al. measured sjTREC levels in adult recipients of umbilical cord blood transplants following a novel non-myeloablative regimen. CD3þ T cells reached normal levels in adults 6 to 12 months following non-myeloablative transplantation compared with 24 months in adults receiving a myeloablative regimen. sjTRECs were detected 12 months after transplan-tation in the non-myeloablative recipients, whereas sjTRECs were not detected in adults until 18 – 24 months in those receiving myeloablative regimens. Indicating that thymic recovery is likewise accelerated in non-myeloablative compared to myeloablative regimens [31]. Lewin et al. detected sjTRECs levels in 158 patients who had undergone allo-HSCT. All patients had received myeloablative conditioning regimens and were full donor chimeras in remission. Younger patients exhibited more rapid recovery and higher sjTRECs. Recipients of T-cell-depleted allo-grafts initially had lower sjTRECs than unmodified allograft recipients, but the difference abated beyond 9 months. sjTRECs level disparities did not achieve
significance among adults with respect to type of allograft. Recipients of all ages of either unmodified or T-cell-depleted allografts therefore actively generate new T cells [23]. In conclusion, sjTRECs quantitative detection is a sensitive method for evaluation the immune recovery after different methods of SCT, but it may analyze with the other influencing factors, as well as age and the source of stem cells and so on.
Detection of sjTRECs is also used to evaluate the ability of immune recovery after donor lymphocyte infusion (DLI), which has been used to enhance graft-versus-leukemia activity after SCT, based on the delayed reconstitution of T-cell immunity after SCT. Bellucci et al. showed that CD4þ DLI was also associated with increased numbers of sjTRECs in CD3þT cells [32].
Recent thymic output function after chemotherapy
The thymus appears to play a role in immune recovery in adults after receiving chemotherapy. Previous studies demonstrate that naı¨ve and memory T cells show differential sensitivity to chemotherapy. Aquino et al. showed that 13 patients received a mean of 3:8^ 0:4 courses of fludarabine and 10 of them had detectable sjTREC levels in sorted CD4þand CD8þ T cells, in which the samples were obtained at a average of 18.8 (range 3.6 – 40.3) months after completing chemotherapy [33]. Another prospective study from Haining et al. showed that 73 children with ALL receiving a protracted, 2-year chemotherapy regimen and compared to an age-matched cohort of 805 healthy children sjTRECs levels was significantly lower in ALL patients than in normal controls, consistent with the reduction in naı¨ve T cells [18]. There was a positive correlation between the degree of sjTRECs expression and the percentage of naı¨ve T cells, naı¨ve CD4þ and naı¨ve CD8þ cells indicating that increased thymic output as demonstrated by an increase in sjTRECs levels correlated with, and was predictive of, increased numbers of naı¨ve T-cell following fludarabine administration [33].
Increase in thymic output function
Age-associated thymic atrophy is a key event preceding the inefficient functioning of the immune system, resulting in a diminished capacity to generate new T cells. Moreover, the number of naı¨ve T cells was markedly suppressed in patients after chemotherapy or SCT. Despite recent progress in our understanding of the biology of T-cell homeo-stasis, clinically available therapies to substantially improve immune reconstitution in patients sustaining T-cell depletion are lacking. Studies have been conducted to improve thymic function. It is proposed that cytokines may play a key role in the early stages
Y. Li
of thymocyte development. Some cytokines (IL-2 and IL-7) and other factors have been shown to increase the thymic output function during immune recovery [34 – 37]. More recently, Haining et al. showed that specifically protect thymopoiesis and/or naı¨ve T cells such as IL-7, KGF or androgen inhibitors may lessen the impact of chemotherapy on the peripheral T-cell pool and improve functional immunity in cancer patients [18].
IL-2
Interleukin-2 (IL-2) was used as immunotherapy with highly active antiretroviral therapy (HAART) to HIV-positive patients and could increase sjTRECs levels more significantly in CD4þcells [34], indicating that IL-2 immunotherapy could be considered for use in improvement of thymic output function in leukemia patients. It may further explain the utility of immunotherapy with IL-2 in improvement in the cellular immune function in the treatment of cancer patients.
IL-7
Thymic atrophy is thought to result from a failure of the thymic microenvironment to support thymo-poiesis in old age and recent evidence suggests that a decline in interleukin-7 (IL-7) expression may limit thymocyte development by restricting combinations of survival, proliferation and rearrangement of the TCRb chain. Although the ability of IL-7 to restore the immune system by enhancing thymic output remains controversial [38], in a thymic organ culture system, exogenous IL-7 has been shown to increase the sjTRECs frequency in fetal as well as infant thymus. The in vivo experiments using NOD/LtSz-scid mice given transplants of human fetal thymus and liver suggested that IL-7 can also directly enhance sjTRECs generation. The results provide compelling evidence that IL-7 has a direct effect on increasing TCRa and TCRb rearrangement and indicate the potential use of IL-7 for enhancing new naı¨ve T-cell generation in immunocompromised patients [35]. It is remarkable that IL-7 is a key cytokine in the early stages of thymocyte develop-ment and expression levels are greatly reduced with age [39]. It is a critical factor limiting the survival for naı¨ve T cells, and elevated levels of IL-7 serve to increase expansion of naı¨ve T cells [40]. Further-more, IL-7 can improve T-cell recovery after experimental T-cell-depleted bone marrow trans-plantation (TCD-BMT) in T-cell-deficient mice by strong expansion of RTEs. Quantification of sjTRECs in mice receiving a TCD-BMT revealed that enhanced T-cell recovery following IL-7 treat-ment results from a strong expansion of newly developed naı¨ve T cells [36].
Flt3 ligand
The Flt3 ligand (flt3L) receptor is present on thymic progenitors and flt3L is expressed within thymic CD4þ T cells by diminishing competition for self ligands. Flt3L has potent effects on hematopoietic stem cell populations, which could result in enhanced thymic homing of T-cell progenitor, and flt3L could act to expand common lymphoid progenitors, which may be limiting for T-cell regeneration in vivo. Recently, it has been reported that flt3L is a potent immunorestorative agent that enhances both thymic-dependent and thymic-inde-pendent pathways of T-cell regeneration. Fry et al. showed that following BMT in athymic hosts, both dendritic cells and T cells were profoundly depleted and flt3L therapy restored DC numbers and enhanced thymic-independent homeostatic peri-pheral expansion. Thymus-bearing BMT recipients treated with flt3L regenerated increased numbers of thymic-dependent progeny with increased numbers of sjTRECs-positive T cells [37].
Other factors
Neuroendocrine growth factors have been implicated in the regulation of thymocyte development and overall thymic function through direct effects on developing thymocytes and/or modulation of support-ing thymic stroma. Insulin-like growth factor 1 (IGF-1) is a ubiquitously expressed mediator of cellular anabolic function that has been shown to affect the growth of primary and secondary immune organs. Recombinant human IGF-1 was administered into healthy young adult mice and resulted in significant increases in thymic size, cellularity and sjTRECs numbers by continuous two-week infusion. In peripheral T-cell populations, significant increases in the number of CD4þand CD8þnaı¨ve cells were observed after four weeks of IGF-1 treatment. Significant increases in total peripheral sjTRECs numbers were observed at this time, indicating an increase in the number of RTEs with IGF-1 administration. These findings support the concept of IGF-1 as a potentially useful agent in facilitating T-cell reconstitution following hematopoietic stem cell transplant [41].
Conclusions
Quantitative sjTRECs measurements provide a novel and sensitive molecular tool for identification of RTEs in peripheral blood and detection of T-cell production by the thymus. This method is useful for evaluation of cellular immune status in patients with hematological malignancy, cancer patients and immunodeficiency disease and for comparison of the immune reconstitu-tion after SCT and chemotherapy and for screening of
factors for enhancing thymic output function. How-ever, it is important to recognize that quantitative PCR sjTREC analysis provide only an average sjTREC content. The absolute number of sjTRECs that can be calculated based on this measurement, is affected by other parameters, such as cell division, cell death, longevity of naı¨ve T cells and intracellular degradation. Therefore further research is required on the development of mathematical models that allow quantitative estimations of thymic output with sjTRECs analyses by taking into account the main factors that affect sjTRECs content and sjTRECs numbers [2,42].
Moreover, quantitative ofdRec –cJasjTRECs can directly evaluate the recent thymic output function, and it is unable to analyze the particular thymic output function of different TCR Vbsubfamily naı¨ve T cells, because the complexity of TCR Vb repertoire is an important factor for immune reconstitution. The quantitative analysis of series TCR Vb– DbsjTRECs to evaluate the levels of different Vbsubfamily naı¨ve T cells should be established.
Acknowledgements
The project was sponsored by grants from National Natural Science Foundation of China (No. 30270579), Natural Science Foundation of Guang-dong Province (No.23001) and the Department of Education of Guangdong Province (No. Q 02022).
References
[1] Douek DC, McFarland RD, Keiser PH, Gage EA, Massey JM, Haynes BF, Polis MA, Haase AT, Feinberg MB, Sullivan JL, Jamieson BD, Zack JA, Picker LJ, Koup RA. Changes in thymic function with age and during the treatment of HIV infection. Nature 1998;396:690 – 695.
[2] Hazenberg MD, Verschuren MC, Hamann D, Miedema F, van Dongen JJ. T cell receptor excision circles as markers for recent thymic emigrants: Basic aspects, technical approach, and guidelines for interpretation. J Mol Med 2001;79:631 – 640. [3] De Villartay JP, Hockett RD, Coran D, Korsmeyer SJ, Cohen
DI. Deletion of the human T-cell receptord-gene by a site specific recombination. Nature 1988;335:170 – 174.
[4] Geenen V, Poulin JF, Dion ML, Martens H, Castermans E, Hansenne I, Moutschen M, Sekaly RP, Cheynier R. Quantification of T cell receptor rearrangement excision circles to estimate thymic function: An important new tool for endocrine-immune physiology. J Endocrinol 2003;176: 305 – 311.
[5] Al-Harthi L, Marchetti G, Steffens CM, Poulin J, Sekaly R, Landay A. Detection of T cell receptor circles (TRECs) as biomarkers forde novoT cell synthesis using a quantitative polymerase chain reaction-enzyme linked immuno-sorbent assay (PCR-ELISA). J Immunol Methods 2000;237: 187 – 197.
[6] Markett ML, Boeck A, Hale LP, Kloster AL, McLaughlin TM, Batchvarova MN, Douek DC, Koup RA, Kostyu DD, Ward FE, Rice HE, Mahaffey SM. Transplantation of thymus tissue in complete DiGeorgew syndrome. N Engl J Med 1999;341:1180 – 1189.
[7] Patel DD, Gooding M, Parrott RE, Curtis KM, Haynes BF, Buckley RH. Thymic function after hematopoietic stem-cell transplantation for the treatment of severe combined immunodeficiency. N Engl J Med 2000;342:1325 – 1332. [8] Weinberg K, Blazar BR, Wagner JE, Agura E, Hill BJ,
Smogorzewska M, Koup RA, Betts MR, Collins RH, Douek DC. Factors affecting thymic function after allogeneic hematopoietic stem cell transplantation. Blood 2001;97:1458 – 1466.
[9] Broers AE, Meijerink JP, van Dongen JJ, Posthumus SJ, Lowenberg B, Braakman E, Cornelissen JJ. Quantification of newly developed T cells in mice by real-time quantitative PCR of T-cell receptor rearrangement excision circles. Exp Hematol 2002;30:745 – 750.
[10] Li Y, Yang L, Chen S, Wang Z, Wu X, Przybylski G, Schmidt CA. Recent thymic output function in patients with acute myelogenous leukemia. Blood 2003;102(11):608a.
[11] Li Y, Geng S, Yang L, Chen S, Yin Q. Naı¨ve T cell level and TCR Vb repertoire usage in patients with chronic myelo-genous leukemia. Blood 2004;104(11):246b.
[12] Kimmig S, Przybylski GK, Schmidt CA, Laurisch K, Mowes B, Radbruch A, Thiel A. Two subsets of naı¨ve T helper cells with distinct T cell receptor excision circle content in human adult peripheral blood. J Exp Med 2002;195:789– 794. [13] Pido-Lopez J, Imami N, Aspinall R. Both age and gender affect
thymic output: More recent thymic migrants in females than males as they age. Clin Exp Immunol 2001;125:409 – 413. [14] Steffens CM, Al-Harthi L, Shott S, Yogev R, Landay A.
Evaluation of thymopoiesis using T cell receptor excision circles (TRECs): Differential correlation between adult and pediatric TRECs and naı¨ve phenotypes. Clin Immunol 2000;97:95– 101.
[15] Ye P, Kirschner DE. Reevaluation of T cell receptor excision circles as a measure of human recent thymic emigrants. J Immunol 2002;168:4968 – 4979.
[16] Petridou E, Klimentopoulou AE, Moustaki M, Kostrikis LG, Hatzakis A, Trichopoulos D. Recent thymic emigrants and prognosis in T- and B-cell childhood hematopoietic malig-nancies. Int J Cancer 2002;101:74 – 77.
[17] Li Y. Leukemia associated clonal expansion TCR Vb
subfamily T cells. Hematology 2004;8:375– 384.
[18] Haining WN, Evans J, Seth N, Callaway G, Wucherpfennig K, Nadler L, Guinan E. Rapid T Cell response to vaccination can occur without antibody response in children post HSCT. Blood 2004;104(11):614a.
[19] Yasunaga JI, Sakai T, Nosaka K, Etoh KI, Tamiya S, Koga S, Mita S, Uchino M, Mitsuya H, Matsuoka M. Impaired production of naı¨ve T lymphocytes in human T-cell leukemia virus type I-infected individuals: Its implications in the immunodeficient state. Blood 2001;97:3177 – 3183.
[20] Svaldi M, Lanthaler AJ, Dugas M, Lohse P, Pescosta N, Straka C, Mitterer M. T-cell receptor excision circles: A novel prognostic parameter for the outcome of transplantation in multiple myeloma patients. Br J Haematol 2003;122: 795 – 801.
[21] Hochberg EP, Chillemi AC, Wu CJ, Neuberg D, Canning C, Hartman K, Alyea EP, Soiffer RJ, Kalams SA, Ritz J. Quantitation of T-cell neogenesis in vivo after allogeneic bone marrow transplantation in adults. Blood 2001;98: 1116 – 1121.
[22] Storek J, Joseph A, Espino G, Dawson MA, Douek DC, Sullivan KM, Flowers ME, Martin P, Mathioudakis G, Nash RA, Storb R, Appelbaum FR, Maloney DG. Immunity of patients surviving 20 to 30 years after allogeneic or syngeneic bone marrow transplantation. Blood 2001;98:3505 – 3512. [23] Lewin SR, Heller G, Zhang L, Rodrigues E, Skulsky E, van
den Brink MR, Small TN, Kernan NA, O’Reilly RJ, Ho DD, Young JW. Direct evidence for new T-cell generation by patients after either T-cell-depleted or unmodified allogeneic
Y. Li
hematopoietic stem cell transplantations. Blood 2002;100:2235 – 2242.
[24] Fallen PR, McGreavey L, Madrigal JA, Potter M, Ethell M, Prentice HG, Guimaraes A, Travers PJ. Factors affecting reconstitution of the T cell compartment in allogeneic haematopoietic cell transplant recipients. Bone Marrow Transplant 2003;32:1001 – 1014.
[25] Chen X, Barfield R, Benaim E, Leung W, Knowles J, Lawrence D, Otto M, Shurtleff SA, Neale GA, Behm FG, Turner V, Handgretinger R. Prediction of T-cell reconstitution by assessment of T-cell receptor excision circle before allogeneic hematopoietic stem cell transplantation in pediatric patients. Blood 2004; Sep 9 [Epub ahead of print].
[26] Poulin JF, Sylvestre M, Champagne P, Dion ML, Kettaf N, Dumont A, Lainesse M, Fontaine P, Roy DC, Perreault C, Sekaly RP, Cheynier R. Evidence for adequate thymic function but impaired naı¨ve T-cell survival following allogeneic hematopoietic stem cell transplantation in the absence of chronic graft-versus-host disease. Blood 2003;102: 4600 – 4607.
[27] Talvensaari K, Clave E, Douay C, Rabian C, Garderet L, Busson M, Garnier F, Douek D, Gluckman E, Charron D, Toubert A. A broad T-cell repertoire diversity and an efficient thymic function indicate a favorable long-term immune reconstitution after cord blood stem cell transplantation. Blood 2002;99:1458 – 1464.
[28] Douek DC, Vescio RA, Betts MR, Brenchley JM, Hill BJ, Zhang L, Berenson JR, Collins RH, Koup RA. Assessment of thymic output in adults after haematopoietic stem-cell transplantation and prediction of T-cell reconstitution. Lancet 2000;355:1875 – 1881.
[29] Malphettes M, Carcelain G, Saint-Mezard P, Leblond V, Altes HK, Marolleau JP, Debre P, Brouet JC, Fermand JP, Autran B. Evidence for naı¨ve T-cell repopulation despite thymus irradiation after autologous transplantation in adults with multiple myeloma: Role ofex vivoCD34+ selection and age. Blood 2003;101:1891 – 1897.
[30] Bahceci E, Epperson D, Douek DC, Melenhorst JJ, Childs RC, Barrett AJ. Early reconstitution of the T-cell repertoire after non-myeloablative peripheral blood stem cell transplan-tation is from post-thymic T-cell expansion and is unaffected by graft-versus-host disease or mixed chimaerism. Br J Haematol 2003;122:934 – 943.
[31] Chao NJ, Liu CX, Rooney B, Chen BJ, Long GD, Vredenburgh JJ, Morris A, Gasparetto C, Rizzieri DA.
Nonmyeloablative regimen preserves “niches” allowing for peripheral expansion of donor T-cells. Biol Blood Marrow Transplant 2002;8:249– 256.
[32] Bellucci R, Alyea EP, Weller E, Chillemi A, Hochberg E, Wu CJ, Canning C, Schlossman R, Soiffer RJ, Anderson KC, Ritz J. Immunologic effects of prophylactic donor lymphocyte infusion after allogeneic marrow transplantation for multiple myeloma. Blood 2002;99:4610– 4617.
[33] Aquino VM, Douek DC, Berryman B, Johnson M, Jain VK, Collins RH. Evaluation of thymic output by measurement of T-cell-receptor gene rearrangement excisional circles (TREC) in patients who have received fludarabine. Leuk Lymphoma 2003;44:343 – 348.
[34] Marchetti G, Meroni L, Molteni C, Bandera A, Franzetti F, Galli M, Moroni M, Clerici M, Gori A. Interleukin-2 immunotherapy exerts a differential effect on CD4 and CD8 T cell dynamics. AIDS 2004;18:211 – 216.
[35] Okamoto Y, Douek DC, McFarland RD, Koup RA. Effects of exogenous interleukin-7 on human thymus function. Blood 2002;99:2851 – 2858.
[36] Broers AE, Posthumus-van Sluijs SJ, Spits H, van der Holt B, Lowenberg B, Braakman E, Cornelissen JJ. Interleukin-7 improves T-cell recovery after experimental T-cell-depleted bone marrow transplantation in T-cell-deficient mice by strong expansion of recent thymic emigrants. Blood 2003; 102:1534 – 1540.
[37] Fry TJ, Sinha M, Milliron M, Chu YW, Kapoor V, Gress RE, Thomas E, Mackall CL. Flt3 ligand enhances thymic-dependent and thymic-inthymic-dependent immune reconstitution. Blood 2004;104:2794 – 2800.
[38] Henson SM, Pido-Lopez J, Aspinall R. Reversal of thymic atrophy. Exp Gerontol 2004;39:673– 678.
[39] Andrew D, Aspinall R. Age-associated thymic atrophy is linked to a decline in IL-7 production. Exp Gerontol 2002;37: 455 – 463.
[40] Tan JT, Dudl E, LeRoy E, Murray R, Sprent J, Weinberg KI, Surh CD. IL-7 is critical for homeostatic proliferation and survival of naı¨ve T cells. Proc Natl Acad Sci USA 2001;98:8732 – 8737.
[41] Chu YW, Yakar S, LeRoith D, Gress R. Insulin-like growth factor I is a positive regulator of thymic function. Blood 2004;104(11):210a.
[42] Hazenberg MD, Borghans JAM, de Boer RJ, Miedema F. Thynic output: A bad TREC record. Nat Immunol 2003;4:97 – 99.