iii
...
Indian J. Plant Physiol., Vol. XXXIV No.2, pp. 113-121 (lune, 1991)
PLANT INVERT ASES SENSITIVITY TO CALCIUM IONS
ASHOK MISHRA. M. K. SRIVASTAVA AND S. P. SHUKLA
Sugar Chemistry (Ag.) Division, National Sugar Institute. Kanpur
Received on 7 March, 1989
SUMMARY
Cat+ was found to be an inhibitor of plant invertascs in vitro. But in contrast bacterial invertase (Arthrobacter) was not affected. inhibition of sugarcane leaf sheath invertase was competitive, reversed by dialysis and markedly increased by preincubation of tbe enzyme with Ca"+ before addition of substrate. The apparent inhibition constant (Ki) and the apparent number of Cal+ (n) bound per enzyme molecule were found to be 79 mM and 0.8' respectively. The overall reaction was endothermic. Consequently,
Cal+ is postulated to act by a reversible binding to the enzyme. 'I
INTRODUCTION
Invertases have been known to hold a primary role in the permeation process (Bowen, 1972). Besides it also participate in sucrose utHization and sugar storage process in sugarcane (Fernando, 1977). Candido (1983) reported tbe stimulation in invertase activity by Ca2+ in carrot slices. The present study has been extended
using purer invertase preparation from leaf sheath of sugarclne to investigate the effect of Ca2+ on its activity. Efforts have also been made to compare it with invertase from other sources.
MATERIALS AND METHODS
Sugarcane cultivated at tbe National Sugar Institute experiment farm was used for experimentation. Commercial carrot roots were obtained locally and treated (Quiroga 1977). Saccharomyces cerevisiae and bacterial invertase (Arthrobacter
strain-3) were obtained from the Biochemistry Division of the Institute. BDH yeast invertase was taken for the study. Sugarcane leaf sheath and maize invertases were isolated and purified (prado, 1979).
114 ASHOK MISHRA et 01.
minutes at 35°C. Varying concentration of Caz+ as CaCJz were added to the reac
tion mixture. For kinetic and thermodynamic studies. the enzymes were pre incubated with Ca2+ for 30 minutes at the reaction temperature. The enzyme action was then started by adding buffered substrate solution to the reaction mixture and the reducing sugars were determined by Nelson (1944) and Somogyi (1945) method. One unit of invertase is the amount whicb bydrolyses 11£ mol of sucrose/min. under the conditions of assay.
The nature of inhibition was determined by Lineweaver Burk Plots (line weaver, 1(34). Apparent inhibition constant (Kj ) was obtained by Dixon Plots (Dixon, 1953). The interaction constant (0) was determined by Hill Plot (Sam pietro
et 01., 1980).
RESULTS AND DISCUSSION
Effeet of Ca2+ on invertase activity
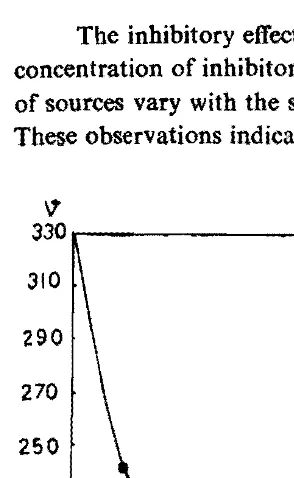
The inhibitory effect tends asymptotically to a minimum value with increasing concentration of inhibitor (Fig. 1). The activity of invertases derived from a variety of sources vary with the source of enzyme and is concentration dependent (Table-I). These observations indicate tbat Ca2+ is able to modify the activity of invertases.
..,
330r---~
f
-
1
Fig. 1. Effect of C1lCItt on the activity of sugarcane leaf sheath invertase.
...,
Jo .
0-1 0-2 0-3 ().4 0-5CaCI2[MJ
310
290
270
250
230
210
190
170
PLANT INVERTASES 115
Table I. Inhibition of invertase by Ca2+*
-Yeast 11 18 25 31 42
Saccharomyces
ccrivisiac 8 17 24 36 48
Maize 6 11 18 23 37
Carrot 9 22 36 48 56
Arthrobacter
o
o
o
0 0r
i
I
Enzyme Cal+concentration (mM) and %inhibition
source
- - -
100 200 300 400 500
• Assay conditions-:were those described in the experimental. Equivalent units of enzyme preparations were used from each source.
Effect of pH on inhibition
The Ca2+ produces a shift of the optimum pH of soluble leaf-sheath invertases from 5 to 6 accompanied by loss in activity (Fig. 2). It has been reported that
0.4
Fig. 2. pH dependence of invertase activity _ without CaQ.
6. - 6.
with9
0·2 CaCh.o~~
____
~__
~~__
~~2 3 .. 5 6 7 8
pH
invertase forms more complex polyacids of molybdenum with lowering of pH
•
(Weigel. 1963). it is possible that at lower pH, Ca2+ also complexes with enzyme116
AlBO&: Y.JIBJtA et01.
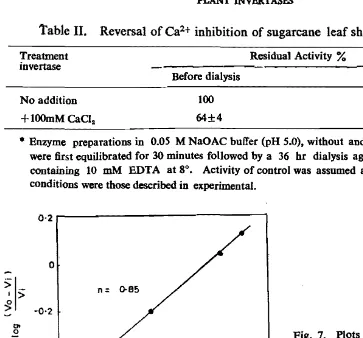
Reversibility of Ca2+ inhibition
The reversibility of sugarcane leaf sheath invertase was examined by dialysing calcium saturated enzyme samples for 36 hrs against 0.05 M NaOAC buffer (pH 5.0), containing 10 mM EDT A. The activity of invertase is restored as soon as excess of
Ca2+ was removed. However, the dialysis of enzyme (wiih('ut Ca2 +) showed an
inhibition of about 12%. It may be due to the loss of less tightly bound calcium of the enzyme while this inhibition could not be restored by the addtion of CaH , indicating the possibility of partial denaturation of enzyme during dialysis or remo val of some of the tightly bound Ca2+ during dialysis causing to an irreversible
inactivation. Fischer (1960) found that the tightly bound calcium holds the enzyme molecule in correct conformation for maximum activity without participating directly in the formation of the enzyme substrate complex and weakly bound calcium stabilizes the enzyme molecule against protease digestion and thermal inac tivation (Rogers, 1969; Greenwood 1970).
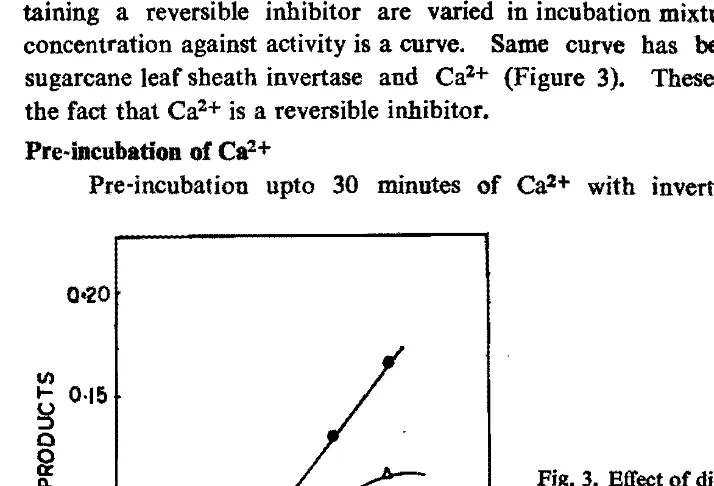
According to Dixon (1960) when the concentration of an enzyme extract con taining a reversible inhibitor are varied in incubation mixture, the plot of enzyme concentration against activity is a curve. Same curve has been obtained with the sugarcane leaf sheath invertase and Ca2+ (Figure 3).
the fact that Ca2+ is a reversible inhibitor.
Pre-iDcubation of Ca2+
Pre-incubation upto 30 minutes of Ca2+ with
These results also support
l
J>-..,
invertase enzyme before the
l..
j ,
i
~
1,
t"
Fig. 3. Effect of dilutions of a mi~ture of 200 1'1 of enzyme and 100 1'1 of
lOOmM CaCI!. control
e-e
and mixture 1::,. - !:, ••
Q<20
I/)
g
O·I!) o ~Q. ~ 0·10
o
30 60 90 12Q 150 leo
l
111
addition of substrate increase the enzyme inhibition while the same for more than 30 minutes did not produce any further increase in the inhibition.. It was also found that pre-incubation of Ca2+ with sucrose did not affect the activity of enzyme.
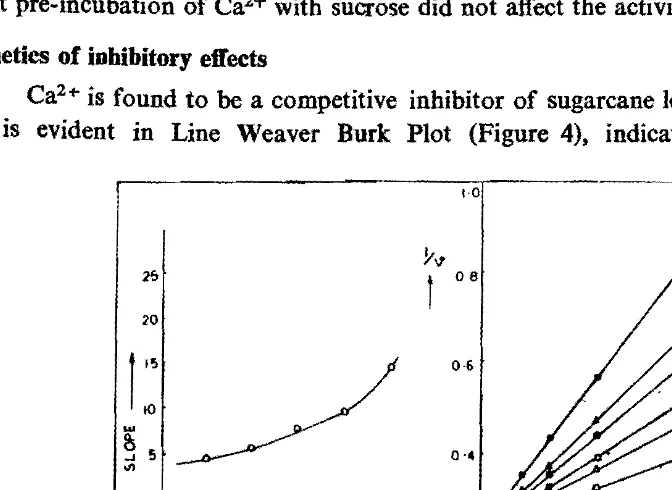
Kinetics of inbibitory effects
Ca2+ is found to be a competitive inhibitor of sugarcane leaf sheath invertase, as is evident in Line Weaver Burk Plot (Figure 4), indicating that Ca2+ when
, ) !
~
!I"I
I!
~
I
t
~
2!i 20
0·1 0·2 0·3 0·4
( CoCf2] M
Y"
) 0 8
o
•
4 II fO-I
( I ISUCROSE] 1.4
Fig. 4. Lineweaver-burk plots showing competitive type of inhibition of sugarcane leaf sheath invertase by Cal +.
(0-0), C~-6) (0-0) ( . - . ) (6-6) (.-B) are with 0, 100 200, 300, 400 and 500 mM Caels respectively.
present in large excess compete with the substrate for the active site of enzyme. The replot of slopes of the double reciprocal plot vs Ca2+ concentration indicates
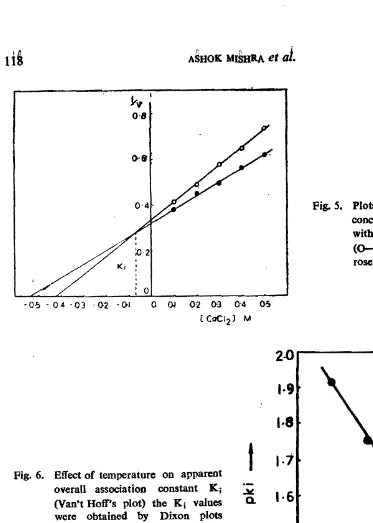
that Ca2+ acts through a complex mechanism, as reported for the action of fructose on invertase (Sampietro et al., 1980), The value of apparent inhibition constant (Kj ) by Dixon plots (Figure 5) was found to be 79 mM. By plotting per cent residual activity vs Ca2+ concentration, a sigmoidal inhibition curve was obtained,
this confirmed the regulatory property of enzyme (Prado, 1979). Furthermore, n calculated from Hill plot (Figure 7) was approximately 0.85, indicating that one Ca2+ atom is bound per enzyme molecule and that the phenomenon of coopera
tivity is not present (Rogers, 1969).
r
tis
K,
: 0 .t-\r I
0-8
Fig. S. Plots of l/v Vs. CaCI. concentrations ( . . ) with 0.2 M sucrose and
(0-0) with 0.1 M suc
rose in assay mixture.
-O~ - 0 4 -03 02 . 01 0 Q.I 0-2 OJ 0-4 05
(CO<:I2) M
2·0..---..
1·9
r~,
1·8
t
,.j
Fig. 6. Effect of temperature on apparentoverall association constant K j
(Van't Hoff's plot) the Ki values
~
'·6
were obtained by Dixon plots(Fig. 5).
1·5
1·3 - - - ' - - -
t.g
3003·' 3-2
~33-4
..!..
X 10"'3
T
& 4'
Table II. Reversal of Ca2+ inhibition of sugarcane leaf sheath invertase·
r
Treatment ResiduaJ Activity %invertase
Before dialysis After dialysis
No addition 100 88±3
+
l00mM CaCla 64±4 89±4• Bnzyme preparations in 0.05 M NaOAC buffer (pH 5.0). without and with CaCI. (100 mM) were first equilibrated for 30 minutes followed by a 36 hr dialysis against the same buffer containing 10 mM BDTA at 8°. Activity of control was assumed as 100 per cent. As~ay
conditions were those described in experimental.
I
f
0'2~---j
0
~I>
~ -0'2
0>
~
-0·14
Fig. 7. Plots of (V~Vi)/Vi Vs log (CaCI.) Where CaCls & Vi is the activity in the presence of CaCl••
-0·6 L..._-,,_ _~_ _ _ _ _~----'
1·9 2·' 2·3 2·'" 2·7
log CaCI 2
inactive enzyme-calcium complex is a function of temperature, decreasing sharply with the increasing temperature. Enthalpy .6.H was calculated from Vant Hoff's plot (Figure 6), as described by Taketa (1965). The AH was found +6.39 Kcal/ mol, indicating the endothermic nature of the reaction between Ca2+ and sugarcane
leaf sheath invertase. From the Ki values, the free energy change (.6. G) values and the entropy change (.6.S) values of the calcium comple~ were calculated by the ,6,G = -RT in K; and .6.G =.6.H T .6.S. At 35°.6.G was found -l.17 Kcal/ mol and ,6,S = -24.58 eu.
The present study has revealed that activities of higher and lower plant inver tases are inhibited by Ca,2+ in vitro. Sugarcane leaf sheath invertase showed maxi mum inhibition, while the invertase from bacteria] source was not affected by Ca2+
120
AsUok MisBRA etai.
Table
In.
Effect of temperature on the sugar cane leaf sheath invertase inhibition-by Ca2+.
Tempe-
ca
s+concentration Ki*rature (mM) and inhibition (%) (mM)
-
100 200 300 400 500...~....
5' 37 51 61 64 69 29±4
15° 33 48 54 00 64 42±4
25° 29 40 46 54 60 57±4
35· 27 32 40 47 51 83±4
"" Apparent inhibition constant (Ki) values were obtained from Dixon Plots (1953). The values are the mean of triplicate estimations.
from that of bacterial source. However. higher activity of sugarcane teaf sheath invertase may be due to higher purification of the eJlZ)'JE.e.
REFERENCES
Bowen,
J.:a,
Hooter, James B. (1972). Sugar transport in immature internodal tissue of sugar cane. II. Mechanism of sucrose transport Plant Physiol. 49,789-793.Candido, P. P. Ricardo, and Jose A.P. Passarinho (1983); Effect of cations on invertase activity and sugar leakage of carrot slices. Phytochemistry, 22, 1701-1706.
Dixon, M. (1953). The Determination of enzyme inhibitor constant. Biochem. J : 55, 170.
t
Dixon, M. and Webb, E.C. (1966). Enzymes, p. 59. Longman, Essex. i~
t
Fernando, B. P.; Marta A. V. and Antonio, R. S. (1977). Sugarcane g1ycosidases. A new I
f
bound invertase from leaf sheaths. ISSCT. 1683-1691. """"': Fischer, B. H. and Stein, B. A. (1960). The enzymes (Boyer P.D., ed.) Vol. IV, 2nd Edn.
p. 313, Academic Press, New York.
Greenwood, C. T. (1970). The Carbohydrates (Pigman, W. & Horton, D. eds.) Vol. IlB, 2nd Bdn. p. 471, Academic Press, New York.
Lineweaver, H. and Dean Burlc (1934). Determinati0!l of enzyme dissociation constants, J.
Am. Chem. Soc. 56, 658-66.
Nelson, N. (1944). A photometric adaptation of the Somogyi method for the determination of glucose. J. Bioi. Chem., 153 : 375-380.
Prado. E.; Sampietro, A. R.; Vattuone, M.A. (1979). Ammonium heptamolybdate, an inhibitor of plant invertases. Phytochemistry, 18, 1799-1802.
Quiroga, E. N.; De Maxud, T. R.; Vattuone, M. A.; Prado, F. E.; Sampietro. A. R. (1977). Sugarcane glycosidases. A general view of the g1yC<"Sidases from stalk, Plant Sci.
I~·
!~ .
..
PLANT INVERTASI!S t21 Rogers, K. S. and Yusko, S. C. (1969). Sodium dodecyl sulfate inactivation of bovine liver
glutamate debydrogenase. J. Bioi. Chem. 244, 6690-6695.
Sampietro, A.R.; Vattuone, M.A. and Prado F.E. (1980). A regulatory invertase from sugarcane leaf sbeaths. Phytochemistry, 19, 1637-42.
Somogyi, M. (1945). A new reagent for the determination of sugars, J. Bioi. Chem •. 160; 61-68. Taketa, K. and Pogell B. M. (1965). Allosteric inbibition of Rat liver Fructose-l,6-Diphos
pbatase by adenosine 5'-monophospbate J. Bioi. Chem. 240, 651·662.