One of the defining characteristics of herpes simplex virus type 1 (HSV-1) infection is the ability of the virus to establish a lifelong latent state in neurons. We previously demonstrated (R. Ramakrishnan, A. J. Fink, G. Jiang, P. Desai, J. C. Glorioso, and M. Levine, J. Virol. 68:1864–1873, 1994) by in situ PCR that many more neurons contain viral genomes than are detected by in situ hybridization for HSV latency-associated tran-scripts (LATs). To determine whether all cells which contain genomes express LATs, we examined trigeminal ganglia for LATs 1 and 8 weeks after corneal scarification with ribonucleotide reductase-deficient HSV-1 by in situ reverse transcriptase PCR. The number of LAT-positive cells detected by in situ reverse transcriptase was substantially greater than the number of cells positive by in situ hybridization and appeared to be similar to the number of cells containing HSV genomes by in situ PCR and the number of ganglionic neurons that project to the cornea as detected by retrograde labeling with Fluorogold. These results demonstrate LAT expression in many neurons containing HSV-1 genomes.
One of the defining characteristics of herpes simplex virus type 1 (HSV-1) infection is the ability of the virus to establish a lifelong latent state in neurons of the sensory ganglia (13, 23). The latent state was initially demonstrated empirically by re-activation of virus from ganglionic neurons after explantation of a ganglion in the absence of recoverable replicating virus in the preparation (6). Subsequently, it was shown that in latency, the HSV genome persists in the nucleus in a circular or con-catemeric form (11, 18) with a chromatin-like organization (9). Expression of the more than 75 genes in the HSV genome that are normally expressed during lytic infection is repressed in latency (12, 23). A single region of the genome is generally accepted to remain transcriptionally active during latency, pro-ducing a family of nonpolyadenylated latency-associated tran-scripts (LATs). LATs are transcribed from the strand opposite
to and overlapping the 39end of the ICP0 gene in the inverted
repeat sequence of the unique long segment of the viral ge-nome (7, 22, 24, 25). LAT expression has become the hallmark of latent infection, detectable by in situ hybridization (ISH) in ganglionic neurons for years after primary infection (although low levels of thymidine kinase and ICP4 may be detectable by reverse transcriptase PCR [RTPCR; 17]).
Use of LAT expression as a measure of the number of genome-containing cells has yielded results which suggest that latency is typically established in only a small fraction of the cells in the ganglion, from 8 to 12 LAT-positive cells per section of ganglion infected with replication-defective viruses with a deletion in the thymidine kinase (5) or ribonucleotide reductase (RR) gene (15, 20). However, we have found that cells expressing LATs detectable by ISH represent only a small proportion of the ganglionic neurons which contain HSV ge-nomes detected by in situ PCR (ISPCR) (20). To determine whether the cells which contain genomes detectable by ISPCR
may nonetheless express LATs at a level below the limit of detection by ISH, we developed an in situ RTPCR (ISRTPCR) to detect the expression of LATs in those cells.
Male Sprague-Dawley rats weighing 225 to 250 g were
inoc-ulated in both eyes following corneal scarification with 2ml of
inoculum containing approximately 4 3 106 PFU of HSV-1
KOS virus RR1CAT/RR2lacZ (8) with the coding sequences of both the large (RR1)- and small (RR2)-subunit genes of the reductase deleted. The RR1 promoter drives chloramphenicol acetyltransferase expression, and the RR2 promoter drives
expression ofb-galactosidase in this recombinant virus. After
perfusion with 4% paraformaldehyde, the ganglia were
re-moved and 4-mm-thick paraffin-embedded sections were
pre-pared as previously described (19). Three animals each were sacrificed at 1 and 8 weeks postinoculation. The ISPCR and ISH findings have been reported previously (20); those exper-iments were repeated for the current study, and additional ISRTPCR experiments were done with the same ganglia.
Expression of LATs was determined first by ISH. Sections were deparaffinized in xylene, rehydrated through graded eth-anol solutions, rinsed with RNase-free phosphate-buffered sa-line (PBS), treated with 1% HCl in PBS for 5 min, and then acetylated with acetic anhydride in triethanolamine for 20 min. After successive washes with 70, 80, and 90% ethanol, the sections were equilibrated in hybridization solution (50%
for-mamide, 10% dextran, 23SSC [13SSC is 0.15 M NaCl plus
0.015 M sodium citrate], 0.01% single-stranded DNA, 0.01% tRNA, 0.02% sodium dodecyl sulfate) and hybridized
over-night at 568C in hybridization buffer with a digoxigenin-labeled
riboprobe antisense to the LAT sequence from a plasmid con-taining BamHI fragment B of HSV-1. After being rinsed with
50% formamide–13SSC at 568C and then 13 SSC at room
temperature, the digoxigenin-labeled probe was localized with an alkaline phosphatase-conjugated antidigoxigenin antibody (1:250; Boehringer Mannheim) and detected with 5-bromo-4-chloro-3-indolylphosphate toluidinium (BCIP)–nitroblue tet-razolium (Vector Laboratories). As we have found and
re-* Corresponding author. Mailing address: 325 Scaife Hall, Univer-sity of Pittsburgh, Pittsburgh, PA 15261. Phone: (412) 648-9793. Fax: (412) 648-1239.
6519
on November 9, 2019 by guest
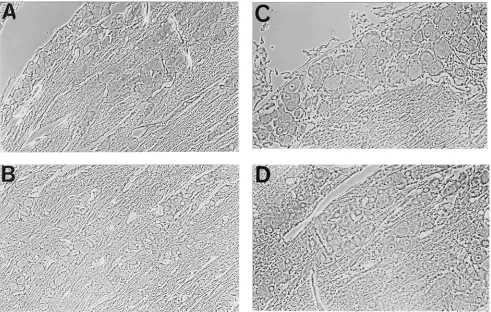
ported previously (20), only 1 to 3 LAT-positive cells per section were detected in ganglia at 1 (Fig. 1A) and 8 (Fig. 2A) weeks postinoculation.
Latent genomes were detected by ISPCR with oligonucleo-tide primers to the HSV glycoprotein B gene as previously
described (20). Briefly, 4-mm-thick sections were
permeabil-ized by treatment with 1% HCl for 5 min; covered with 20ml
of a PCR mixture containing 13 PCR Buffer II
(Perkin-Elmer), 1 mM MgCl2, 13digoxigenin DNA labeling mixture,
100 ng of glycoprotein B primers, 10% glyerol, and 1ml of Taq
DNA polymerase; and sealed under a coverslip with nail pol-ish. Thirty cycles of DNA amplification were carried out in a BioOven III Thermal Cycler (BioTherm Corporation), and the amplified DNA bound by an antidigoxigenin antibody was de-tected with BCIP-nitroblue tetrazolium. As we have reported previously (20), there were 10 to 150 HSV DNA-containing neurons per section at both 1 (Fig. 1B) and 8 (Fig. 2B) weeks postinoculation.
[image:2.612.60.296.69.546.2]To determine whether LATs could be detected by ISRT-PCR, we modified the ISPCR protocol. Sections from the
FIG. 1. Trigeminal ganglion 1 week postinoculation with HSV. (A) LAT-positive nuclei detected by ISH with a digoxigenin-labeled riboprobe. The arrow points to the single positive cell visible. (B) HSV genome containing nuclei detected on sections from the same ganglion by ISPCR with oligonucleotide primers to HSV-1 glycoprotein B and digoxigenin-labeled nucleotides. The ar-row points to 1 of the 72 positive cells visible. (C) LAT-positive nuclei detected by ISRTPCR on sections from the same ganglion with oligonucleotide primers for HSV-1 LAT and digoxigenin-labeled nucleotides. The arrow points to 1 of the 52 positive cells visible.
FIG. 2. Trigeminal ganglion 8 weeks postinoculation with HSV. (A) LATs detected by ISH. The arrow points to the single positive cell visible. (B) HSV genomes detected by ISPCR. The arrow points to 1 of the 40 positive cells visible. (C) LATs detected by ISRTPCR. The arrow points to 1 of the 21 positive cells visible.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.323.548.71.536.2]same ganglia were deparaffinized, and the cells were perme-abilized with 1% HCl in PBS for 15 min (instead of the 5 min employed in the ISPCR), rinsed in RNase-free PBS, and then treated with 400 U of RNase-free DNase (Worthington
Bio-chemicals) and 10 U of RNasin (Promega) in 200ml of 1 mM
NaCl–10 mM Tris-HCl (pH 7.5) at 378C overnight. After
re-peated rinsing with RNase-free PBS to remove excess DNase,
the samples were reverse transcribed in a 100-ml volume
con-taining 10 U of SuperScript II (Gibco-BRL), 5 U of RNasin, 5
ml of LAT 39 primer
(ACG-AGG-GAA-AAC-AAT-AAG-GG) (100 ng/ml), 10 mM dithiothreitol, and 1 mM
de-oxynucleoside triphosphates in SuperScript II reaction buffer
(Gibco-BRL) at 378C for 60 min. After being rinsed with PBS,
LAT cDNA was amplified for 30 cycles in 20 ml of PCR
mixture sealed under glass coverslips in a BioOven III Thermal Cycler as previously described (19). The PCR mixture
con-tained 13 PCR Buffer II (Perkin-Elmer), 13 digoxigenin
DNA labeling mixture, 100 ng of LAT primers (39 primer
described above, 59primer for LAT DNA
[GAC-AGC-AAA-AAT-CCC-GTC-AG]), 10% glycerol, and 1 ml of Taq DNA
polymerase. The amplified cDNA was detected with an anti-digoxigenin antibody conjugated to alkaline phosphatase and visualized with BCIP-nitroblue tetrazolium. The numbers of LAT-positive cells detected by ISRTPCR were 10 to 75 per section at 1 week (Fig. 1C) and 6 to 54 per section at 8 weeks (Fig. 2C), which are comparable to the number of genome-containing cells and much greater than the number of cells detected by ISH in sections from the same ganglia (20).
Control ISRTPCRs in which reverse transcriptase was not added were conducted on serial sections at the same time (Fig. 3A and B). No signal was obtained when reverse transcriptase was omitted, indicating the absence of viral DNA after DNase treatment. No signal was found in sections pretreated with RNase (Fig. 3C) or when the reaction was run without primers (Fig. 3D). These controls served to confirm that the signal detected was derived from amplification of RNA in the sec-tions. In our experience, the most crucial step in obtaining good amplification of RNA is DNase treatment. The appro-priate time for tissue permeabilization and DNase treatment must be optimized for the specific tissue sample. The amount and source of the DNase I used are very critical to complete degradation of the viral DNA without concomitant RNA deg-radation.
We confirmed the specificity of the ISRTPCR signal as a LAT sequence by Southern blot analysis. Amplified cDNA was extracted from sections of infected ganglia after ISRTPCR with LAT-specific primers and unlabeled nucleotides. After agarose gel electrophoresis and transfer to a nylon membrane (GeneScreen Plus; NEN Research Products), the DNA was
hybridized with a g-32P-59-end-labeled LAT oligonucleotide
[image:3.612.62.553.70.382.2]probe complementary to sequences internal to the PCR primer sequences (CGA CAC CGA TTG GCT GGT GT) (19) by using the Rapid-Hyb system (Amersham Life Sciences). The 195-bp product recognized by hybridization confirmed that the ISRTPCR product from the tissue was LAT specific (Fig. 4A). To determine whether the amplification in the presence of
FIG. 3. Control ISRTPCR. After DNase treatment overnight, a section from the same ganglion (1 week postinoculation) was amplified with HSV-1 LAT oligonucleotide primers and digoxigenin-labeled nucleotides but without reverse transcriptase (A). No positive nuclei are visible. Similar results were obtained at 8 weeks postinoculation (B). Sections pretreated with RNase A prior to RTPCR showed no reaction product (C). Reactions carried out in the absence of primers were also negative (D).
on November 9, 2019 by guest
LAT primers produced other amplimers in addition to the 195-bp product, the same in situ amplification reaction was
carried out on sections in the presence of [32P]dCTP and the
amplified product was extracted and detected after electro-phoresis by autoradiography. A single 195-bp band was found
(Fig. 4B), indicating that amplified LAT is the sole product of the reaction.
We also used the retrograde axonal tracer Fluorogold (Flu-rochrome Inc.) to define the distribution of trigeminal ganglion
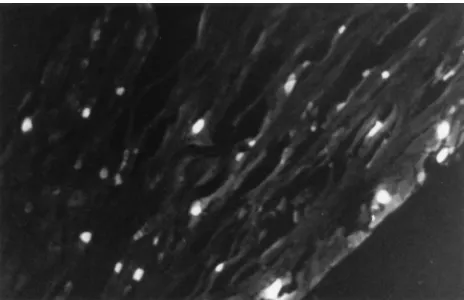
cells that project to the cornea. A 15-ml volume of a 5%
(wt/vol) aqueous solution of Fluorogold was applied, after cor-neal scarification, to both eyes of three rats, and after 1 week, Fluorogold-containing cells were detected in paraffin-imbed-ded sections by fluoroscence microscopy with a wide-band UV filter. The number and distribution of neurons containing Flu-orogold (Fig. 5) were similar to those of the neurons contain-ing HSV-1 DNA, as detected by ISPCR, or LAT, as detected by ISRTPCR.
We had previously demonstrated that use of LAT expression detected by ISH as a measure of the number of genome-containing cells underestimates the number of latent HSV genome-containing cells in a trigeminal ganglion. In this study, we found that by use of ISRTPCR to detect LAT expression, the majority of cells containing HSV genomes can be shown to express LATs, although at levels below the limit of detection by ISH. There are several possible explanations for this observa-tion. Two different LAT promoters, LAP1 and LAP2, have been identified (2, 10, 14), and it has been suggested that high-level expression in some cells reflects LAP1 activity while the low level of expression in other cells reflects LAP2 activity in the absence of LAP1 activity (4). Alternatively, expression of undefined transcriptional regulatory cellular gene products may transactivate high-level LAT expression in a limited subset of ganglionic neurons from only a single viral promoter ele-ment. Responsive elements in the LAT promoter sequence include a TATA box (2), CCAAT box homology, and recog-nition sequences for transcription factors including Sp1, AP-2, YY-1, CREB, LPBF/MLTF/USF, Egr-1, and the HSV-1 trans-activator ICP4 (1, 3, 16, 21, 26). Finally, the level of LAT expression may reflect the number of HSV genomes in neu-rons. High levels of LATs detectable by ISH may be found in cells which contain many HSV genomes, and lower levels may be found in cells containing fewer genomes. The apparent uniformity of the signals obtained with ISPCR and ISRTPCR may result from the asymptotic effect of amplification.
The expression of LATs in many HSV-1 genome-containing cells has important implications regarding the regulation of LAT expression and the role of LATs in latency, as well as for the potential use of HSV-based vectors for gene transfer to the nervous system.
FIG. 4. Specificity of the amplified signal. (A) Southern hybridization of cDNA extracted from single 4-mm-thick sections of ganglia at 1 and 8 weeks postinoculation after in situ RTPCR with LAT primers and unlabeled nucleo-tides. The extracted DNA was electrophoresed on a 1% agarose gel, transferred, and hybridized to a32
P-labeled LAT oligonucleotide probe internal to the PCR primers. The reaction products obtained with (1) and without (2) inclusion of reverse transcriptase are shown. A band corresponding to the 195-bp LAT fragment was visible only in the presence of reverse transcriptase. (B) ISRTPCR was performed on a serial 8-week section in the presence of oligonucleotide primers to LAT and 10mCi of [a-P32
[image:4.612.320.552.69.219.2]]dCTP. The amplified product was extracted and electrophoresed on a 1% agarose gel, which was then exposed to X-ray film. The reaction products obtained with (1) and without (2) reverse transcriptase are shown. A single band corresponding to the 195-bp LAT fragment was visible only in the presence of reverse transcriptase.
FIG. 5. Ganglionic neurons containing Fluorogold 1 week after application of Fluorogold to the cornea. The distribution of labeled cells is similar to that determined for HSV genomes by ISPCR and that determined for LATs by ISRTPCR. Forty-one positive cells are visible in this section.
on November 9, 2019 by guest
http://jvi.asm.org/
J. Virol. 69:7899–7908.
5. Coen, D. M., M. Kosz-Vnenchak, and J. G. Jacobsen. 1989. Thymidine kinase-negative herpes simplex virus mutants establish latency in mouse trigeminal ganglia but do not reactivate. Proc. Natl. Acad. Sci. USA 86:4736– 4740.
6. Cook, M. L., V. B. Bastone, and J. G. Stevens. 1974. Evidence that neurons harbor latent herpes simplex virus. Infect. Immun. 9:946–951.
7. Deatly, A. M., J. G. Spivack, E. Lavi, D. R. O’Boyle, and N. W. Fraser. 1988. Latent herpes simplex virus type 1 transcripts in peripheral and central nervous system tissues of mice map to similar regions of the viral genome. J. Virol. 62:749–756.
8. Desai, P., R. Ramakrishnan, Z. W. Lin, B. Osak, J. C. Glorioso, and M. Levine.1993. The RR1 gene of herpes simplex virus type 1 is uniquely trans activated by ICP0 during infection. J. Virol. 67:6125–6135.
9. Deshmane, S. L., and N. W. Fraser. 1989. During latency, herpes simplex virus type 1 DNA is associated with nucleosomes in a chromatin structure. J. Virol. 63:943–947.
10. Dobson, A. T., F. Sederati, G. Devi-Rao, W. M. Flanagan, M. J. Farrell, J. G. Stevens, E. K. Wagner, and L. T. Feldman.1989. Identification of the latency-associated transcript promoter by expression of rabbit beta-globin mRNA in mouse sensory nerve ganglia latently infected with a recombinant herpes simplex virus. J. Virol. 63:3844–3851.
11. Efstathiou, S., A. C. Minson, H. J. Field, J. R. Anderson, and P. Wildy. 1986. Detection of herpes simplex virus-specific DNA sequences in latently in-fected mice and in humans. J. Virol. 57:446–455.
12. Fawl, R. L., and B. Roizman. 1994. The molecular basis of herpes simplex virus pathogenicity. Semin. Virol. 5:261–271.
13. Fraser, N. W., T. M. Block, and J. G. Spivack. 1992. The latency-associated transcripts of herpes simplex virus: RNA in search of a function. Virology 191:1–8.
simplex virus genome in a mouse model system: evidence suggesting an episomal state. Virology 158:265–275.
19. Ramakrishnan, R., D. J. Fink, G. Jiang, P. Desai, J. C. Glorioso, and M. Levine.1994. Competitive quantitative PCR analysis of herpes simplex virus type 1 DNA and latency-associated transcript RNA in latently infected cells of the rat brain. J. Virol. 68:1864–1873.
20. Ramakrishnan, R., M. Levine, and D. J. Fink. 1994. PCR-based analysis of herpes simplex virus type 1 latency in the rat trigeminal ganglion established with a ribonucleotide reductase-deficient mutant. J. Virol. 68:7083–7091. 21. Rivera-Gonzalez, R., A. N. Imbalzano, B. Gu, and N. A. DeLuca. 1994. The
role of ICP4 repressor activity in temporal expression of the IE-3 and latency-associated transcript promoters during HSV-1 infection. Virology 202:550–564.
22. Rock, D. L., A. B. Nesbern, H. Ghiasi, J. Ong, T. L. Lewis, J. R. Lokensgard, and S. Wechsler.1987. Detection of latency-related viral RNAs in trigeminal ganglia of rabbits latently infected with herpes simplex virus type 1. J. Virol. 61:3820–3826.
23. Stevens, J. G. 1989. Human herpesviruses: a consideration of the latent state. Microbiol. Rev. 53:318–332.
24. Stevens, J. G., E. K. Wagner, G. B. Devi-Rao, M. L. Cook, and L. T. Feldman. 1987. RNA complementary to a herpesvirusagene mRNA is prominent in latently infected neurons. Science 235:1056–1059.
25. Wagner, E. K., G. Devi-Rao, L. T. Feldman, A. T. Dobson, Y. Zhang, W. M. Flanagan, and J. G. Stevens.1988. Physical characterization of the herpes simplex virus latency-associated transcript in neurons. J. Virol. 62:1194– 1202.
26. Zwaagstra, J. C., H. Ghiasi, A. B. Nesburn, and S. L. Wechsler. 1991. Identification of a major regulatory sequence in the latency associated tran-script LAT promoter of herpes simplex virus type 1 (HSV-1). Virology 182:287–297.