0022-538X/91/084255-08$02.00/0
CopyrightC 1991,American Society for Microbiology
Characterization of Variable Regions in
the Envelope and S3 Open
Reading
Frame
of Equine
Infectious
Anemia Virus
SOREN ALEXANDERSEN*ANDSUSAN CARPENTER Department of Veterinary Microbiology andPreventiveMedicine,
IowaState University, Ames, Iowa50011
Received 11 February 1991/Accepted 30April 1991
Thepolymerase chain reactionwasusedtoamplifyand clonepartsofthe envelopegeneandoverlappingS3 open reading frame, thought to encode rev, of the virulent in vivo-derived Th-1 isolateof equine infectious
anemia virus(EIAV).The results indicated that EIAV consists ofaheterogeneousmixture ofgenotypespresent
atthe firstfebrile cycle after initial infection. We showedthat the Th-lisolateapparentlycontains nondefective
genotypes aswellastypes which have transmembrane protein truncationsor are revdeficient. Furthermore,
wecouldconfirmthepresenceofahypervariable region in the gp9Oenvelopeglycoprotein. Taken togetherwith
earlier data on the heterogeneity ofthe regulatory motifs present in the long terminal repeat sequences of
viruses from the same in vivo isolate (S. Carpenter, S. Alexandersen, M. J. Long, S. Perryman, and B.
Chesebro,J. Virol. 65:1605-1610, 1991),ourfindings indicate that EIAVusesacomplexsystemof diversity
inbiological phenotypes togetherwithvariationin regulatory andantigenicmakeuptoevade hostresponseand
tocausepersistent infection and recurrentchronic disease.
Equineinfectious anemia virus (EIAV) causes a persistent
infection in horses. The infection is characterized by a
clinically variable disease course which can be acute,
chronic,orasymptomatic (27). EIAV belongs to the
subfam-ilyLentivirinae of the family Retroviridae and is genetically
and antigenically related to other lentiviruses, including human immunodeficiency virus (HIV) (11, 24, 28, 41, 43, 53, 62).
Lentiviruses vary considerably, and infected individuals
or animals can harbor a genetically complex mixture of
genotypes presentatthe sametimeor overperiods of time
(4, 7, 9, 10, 14, 20, 21, 25, 28, 36, 40, 44, 48, 55, 56, 62, 64). One factor important in lentivirus variant selection is the
specificity ofthe host immune response (13, 32, 38, 42, 44).
Inaddition, recent findings suggest that variation in
biolog-ical phenotype is an important mechanism of lentivirus
pathogenesis. Progression todisease in individuals infected with HIV could be correlated with recovery of virus isolates which were more cytopathic and exhibited a broader in vitro
host range (9, 64). Similarly, HIV isolates recovered from
asymptomatic individuals replicate slowly and to low titers in vitro, whereas isolates recovered from individuals with
more severe clinical signs ofinfection arecharacterized by
rapid replication and production of high levels of reverse
transcriptase activity (4, 20).
Progressin defining moleculardeterminants important in
viral virulence and host response is beginning to emerge.
Recentstudiessuggestthat severalviral genes,aswellasthe
long terminal repeat (LTR), contribute to the biological
diversity of HIV type 1 (23, 65). Also, the presence of
specific enhancer elements in other retrovirus LTRs has
been shown toinfluence cell tropism, virulence,and
leuke-mogenicity(33, 58). We havepreviouslyshown that theLTR
sequences of an avirulent cell culture-adapted isolate of
EIAVdiffered from the invivo parental isolate (6). Thetwo
isolates differed in the target cell tropism, i.e.,the parental
isolate Th-1 grew well inmacrophagesandpoorly inequine
*
Corresponding
author.dermalcells, whereas theoppositewastruefor the
culture-adaptedisolate MA-1 (7). Moreover, thespecific regulatory
sequencemotifs present in the LTRs of thetwoisolateswere
significantly different (6). The retrovirus LTR contains the
sole promoterfor viraltranscriptionandisregulatedbothby
viral trans-acting proteins such as tat (3, 60, 61) and by a
multitude ofcellularproteins involved in DNA binding and
transcription complex assembly, initiation, and elongation
(16, 46). Theseresultssuggested that variation in regulatory
sequences in the LTRmay be important in EIAV host cell
tropismand pathogenesis in vivo.
Instudies of EIAV and visnavirus, genetic mutations in
the viral env gene were associated with the occurrence of
antigenicvariant viruseswhichwerepostulatedtopersist by evasion ofthe host immune response (12, 13, 42, 48). For
HIV, the different genotypes have been shown to vary in
biological phenotype, and this variation, in part, is
deter-minedbysequences in theviral envelope(21,65). Also,the
difference in cell tropism ofHIV has been mapped to the
envelope, particularly sequences near the hypervariable
loop V3(21, 35, 45).
In the presentstudywecharacterizedtheenvelope
region
andoverlapping S3 open reading frame, thoughttoencode
rev, of the Th-1 isolate of EIAV derived directly from
infected horse macrophages after asinglepassage in vitro.
The results indicate thatEIAV,asdescribed aboveforHIV,
consists ofa heterogeneous mixture ofgenotypes
already
present at the first febrile cycle after initial infection. We
could show that the isolateapparently contains nondefective
genotypes as well as types which have transmembrane
protein truncations or are rev deficient. Furthermore, we
were able to confirmthepresence ofa hypervariable region
in the gp9O envelopeglycoprotein previously
suggested by
Payne etal. (48). Togetherwith ourearlier dataon
hetero-geneityin the LTRsequencesfrom the samein vivo isolate
(6),thesefindingsindicate thatEIAVuses a
complex
systemofdiversityinbiological phenotypes
together
withvariationin regulatory andantigenic makeupto evade host response
and to cause
persistent
infection and recurrent chronic disease.4255
on November 10, 2019 by guest
http://jvi.asm.org/
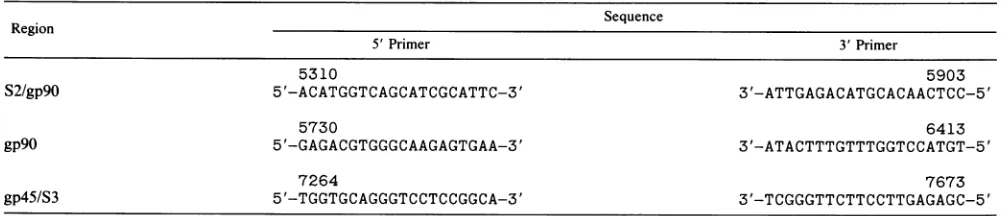
TABLE 1. Nucleotide positions and primersequencesfor variousregions ofthe EIAVgenomea Sequence
Region
5'Primer 3' Primer
5310 5903
S2/gp9O 5'-ACATGGTCAGCATCGCATTC-3' 3'-ATTGAGACATGCACAACTCC-5'
5730 6413
gP9O 5'-GAGACGTGGGCAAGAGTGAA-3' 3'-ATACTTTGTTTGGTCCATGT-5'
7264 7673
gp45/S3 5'-TGGTGCAGGGTCCTCCGGCA-3' 3'-TCGGGTTCTTCCTTGAGAGC-5'
a Thenucleotide positionof the5'endisgivenabove each sequence.
MATERIALSANDMETHODS
Virus and cells. Th-1 is a field-derived isolate of EIAV
recovered from a horse experimentally inoculated with
whole blood from anEIAV-seropositive, naturally infected
horse (7, 8). The experimentally infected horse was
moni-tored daily after infection, and primary horse macrophage
cultures (HMC) were established from peripheral blood
mononuclearcells aspreviously described (7, 17). The Th-1
virus stock is the cell-free supernatant fluid from these
primary HMCestablished duringthefirstfebrile episode of
equine infectious anemia and grown for 8 to 10 days in
culture (6, 7, 17). Fresh HMC were established from an
EIAV-negative horse, and this culture was inoculatedwith
the Th-1 virus stock. This culture was grown for 6 days,
tested, and foundpositivefor reverse transcriptaseactivity,
and total DNA wasextracted andanalyzedbySouthernblot
hybridization as previously described (6). EIAV sequences
in this DNA preparation represent infectious EIAVafter a
single passage in HMC and, thus, should accurately typify
EIAVgenotypes present in vivo.
Molecular cloning. Amplification and cloning of LTR
se-quences from the DNA preparation ofTh-1-infected HMC
have previously been briefly described (6). In the present study we amplified and cloned segments of the viral
enve-lopeand overlapping S2 and S3 open reading frames using
the same Th-1 DNApreparation and similar techniques. To
clone Th-1 sequences, specific segments were amplified by
thepolymerase chainreaction (PCR) using 20-mer
oligonu-cleotide primers complementaryto sequences atthe 5' and
3' borders of the segment to be amplified. Primers were
synthesizedwith an AppliedBiosystems 380B DNA
synthe-sizer, and the sequence was based on the published
se-quenceof the MA-1 isolate of EIAVderived by cell culture
passage of the Th-1 stock (6). All numbering of nucleotides
in the present report is based on the available GenBank
sequenceof EIAV Wyoming (14, 28), except that a G is put
inafternucleotide 5348 to make the env open reading frame
continuous, aspreviouslydescribed (53). This numbering is in accordance with our previously published sequence of the MA-1isolateof EIAV (6). The nucleotide positions, 3' and 5' ends, and sequences of the primers used for the different areasof the EIAV genome are shown in Table 1.
PCRs were performed in a volume of 100
,u1
containing 200ng of each oligonucleotide primer, 50 mM
KCl,
10 mMTris-HCl (pH 8.4 at 20°C), 1.5 mM
MgCl2,
20p.g
ofgelatin, 0.2 mM each dNTP, 5 U of Taq DNA polymerase (PerkinElmer Cetus, Norwalk, Conn.), and 30 ng of DNA to be
amplified. The reactions were amplified for 25 cycles at an
annealingtemperature of 47°C for 2 min, a synthesis temper-atureof72°C for1.5min, and a heatdenaturation
tempera-tureof94°C for 1min
by using
anautomatedthermalcycler
(JSP
EL-ServiceAPS,
Glostrup, Denmark). Amplified
DNAwas partially purified
by
phenol-chloroform extraction andethanol
precipitation.
Formostof theclonings,
XbaIlinkerswereadded and the DNAwasligatedtopGEM3Z
(Promega
Biotec,
Madison, Wis.) and used to transform competentEscherichia coli JM109 as described previously
(2, 6).
Forcloning
oftheS2/gp9O
segment (clone series 5a), the DNA was cut with BamHI(cuts
at nucleotide 5338) and withHindIII (cuts at nucleotide 5780) and cloned into
pUC19
(GIBCO BRL,
Gaithersburg, Md.). Colonieswere screenedfor the presence of an EIAV insert by hybridization to
EIAV-specific probes
radiolabeled as describedpreviously
(6).
The segment of gp9O amplified with the primers fromnucleotide 5730 to6413 hasaninternal XbaI site at
nucleo-tide6161 andwas thus clonedastwofragments
(designated
clone series 7a and
7b).
Fiveto sevenindependentpositive
colonies from each cloned segment, a total of 26
clones,
were picked, purified by replating, and sequenced by the
dideoxynucleotide chain termination method(57) with
dou-ble-stranded DNA templatesas described previously (2,
5,
6). Each setofclones, representinga specific segment, was
cloned from a single PCR experiment. Cloningofthe
gp9O
segment from another PCR experiment showed the same
distribution of sequence differences (datanot
shown).
Sequences were analyzed by using the Beckman
Micro-genieand the IBI Pustell Sequence Analysis Software
pro-grams. Genetic comparisons were based on sequences of
EIAV MA-1 (6) and EIAVWyoming available in GenBank
orinprevious publications (14, 28, 48, 53, 62).
Nucleotide sequence accession numbers. GenBank
acces-sion numbers for thesequences reported hereareM62655to
M62680.
RESULTS AND DISCUSSION
Wepreviously identified regions in the EIAVenvregion in
which the MA-1 isolatediffered from the Wyoming isolate of
EIAV(6). In the present studyweused PCRtoamplify and
clone theseregionsfrom the Th-1isolate,theparental stock
ofMA-1 whichbiologically differs in cell tropism and
viru-lence(7).Th-1 sequenceswereamplified and cloned
directly
from HMCestablished fromaninfected horse during the first
febrilecycle after infection. Areas of the env gene,including
major parts of the surface gp9O and transmembrane
gp45
glycoproteins previously described to have a considerable
degree of variability,werecloned andanalyzed (6, 48). One
region of gp9O analyzed overlaps with the S2 openreading
framehavinganas-yet-unknownfunction. The region of the
gp45 analyzed overlaps with the S3 open reading frame
thoughtto encoderev(15, 51,63).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.67.569.80.188.2]0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0
0 '0 0 10 0 t0 0 10 0
T C') C') It It 0
0 0 0 0 0 0
0 0 0 0 0 0
'0 0 0 '0 0
10 10 tN OD
l1111111l 1111ll1l 11l 11l
[image:3.612.60.552.59.334.2]U
LTR
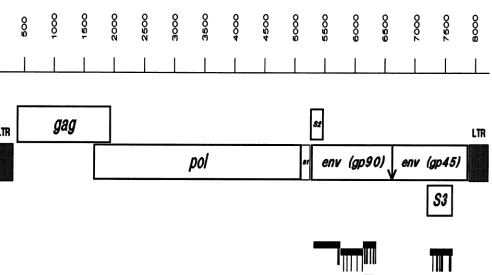
FIG. 1. Schematic representation of theEIAVprovirus. All numbering of nucleotides inthe present report is based onthe available
GenBanksequenceof EIAV Wyoming (14, 28),exceptthataG isputin after nucleotide5348tofixtheenvopenreading frame(53). The total proviralgenome, including bothLTRs,is thus8,229nucleotides. The locations of thetwocomplete LTRs (nucleotides 1to321and7909to
8229) and theopenreading framesgag(nucleotides387to1922),pol (nucleotides1682to5119),env(nucleotides 5303to7888),S1 (nucleotides 5123to5272), S2 (nucleotides 5283to5480), and S3 (nucleotides7234to7638)areindicated. S3 isinreadingframe1;pol, S1, andenvare
inreading frame 2; andgagand S2arein reading frame3. Theputative cleavage siteatnucleotide6643in theenvgeneis indicatedasavertical arrow.Regionsof Th-1 whichwecloned andsequencedareindicatedbythickhorizontal bars. Sequencedifferencesamongthe MA-1(6)and Th-1 clones are shown as thin vertical lines underneath the thick horizontal bars. Fordetails on sequence differences see Fig. 2. The hypervariable region, described previously byPayneetal.(48),spanning nucleotides6176to6343is shownas ahatchedhorizontal bar. The
twostopsignalsin theS3openreadingframe of Th-1 clones 8a51 and8all3areshownashatched circles,and thestopsignalingp45of Th-1 clone8a81isshownas afilled circle.
Nucleotide sequencecomparison betweenthe Th-1 and the
MA-1isolates of EIAV.PCR-generatedclones from the Th-1 isolate of EIAV were cloned and nucleotide sequenced.
Regionsweanalyzedandnumbers of individual cloneswere
as follows: one region in the S2 open reading frame/gp9O
overlap (seven independentclonesdesignatedseries5a),two
regionsinthegp9O area(seven cloneseachofseries 7a and
b), and one region in the gp45/S3 area (five independent
clonesdesignatedseries8a). Oligonucleotide-primer binding sites and regions near the primers which are difficult to
sequence in both orientations were excluded from further
analysis. Thegenomic organization ofEIAV and the
posi-tions of the sequenced segments of the Th-1 isolate are
shown in Fig. 1.
Wecomparedthe nucleotidesequencesof the Th-1 clones
with that of the cellculture-adapted MA-1 isolate of EIAV derived from the Th-1 stock(6) and found severalchanges
(Fig.1 and2).Of the1,029nucleotidessequencedinthegp9O
areaof theenvgene, 16 nucleotides(1.6%)differed between
the Th-l-related clones (Th-1 and MA-1). In the gp45/S3
region,370 nucleotidesweresequencedand 13 bases(3.5%)
differed. AscanbeseenfromFig. 1and2,thechangeswere
notevenlydistributed butweremainlyfound intworegions
of thegp9Oand intwoorthree regionsof thegp45/S3 open
reading frame. No changes were found in the S2 open
readingframe. Of the sevenclones analyzedin the
amino-terminal partofgp9O (series 5a),five cloneswereidenticalto each otherand also identicaltoMA-1. Theothertwoclones in this region had a single nucleotide change each, a
vari-abilitybetweenanypairof the clones between 0 and 2 of444
(Oto0.5%)nucleotides. In the centralregionofgp9O (series
7a) four clones were identical to each other and toMA-1. Theother three clones had 1or2nucleotidechanges each,a
variability ofanypair of clonesbetween 0 and 4 of 373 (0to
1.1%) nucleotides. In the region close to the
carboxy-terminal endofgp9O (series 7b), onlythree ofsevenclones
wereidentical and these three clones hadasinglenucleotide
change comparedwithMA-1 (Fig. 2). Thehighest degreeof
variabilityofanypairof clones in thisregion (series 7b)was
5changesof 212nucleotidesanalyzed,whichisavariability
of 2.4%. Inthegp45/S3 region analyzed (series 8a), onlytwo of the five clones were identical. However, these clones
werehighly differentfrom MA-1 (10 changesof 360 [2.8%]
nucleotidesanalyzed)andwerealsohighlydifferentfrom the other Th-1clones(8to10 nucleotidedifferences).The other three clones in thisregionweremoresimilartoMA-1and to
each other, i.e., 1 to 5 nucleotide differences between any
pair (0.3 to1.4%variability).
Analysisofpotential coding changes. Analysisof thecoding
potential of the Th-1 clones revealed several interesting
features. First,the existence andpositionofahypervariable
region in thegp9O region ofenv, suggested by Payne et al.
LTR
on November 10, 2019 by guest
http://jvi.asm.org/
gp9O region of envelope
Clone
NA-1 5a3 5.8 5.9 5.10 5a11 5.12 5a13
Nucleotide
5743 G (Met) A(lie)
5762 G (Ata) A(Thr*)
MA-1 7a41 7a51 7a71 7.81 7a91 7al16 7al18
Nucteotide
5838 5895 5966
6062
6150
A(Gin)
G (Arg)
T (Phe)
A(Lys) T(lie)
T (Leu)
T(Leu)
C(Leu) G(Gtu)
A (Asn*)
MA-1 7b41 7b51 7b71 7b81 7b91 7b116 7b1i1
Nucleotide
[image:4.612.150.484.81.450.2]6373 G (Met) A (lie)
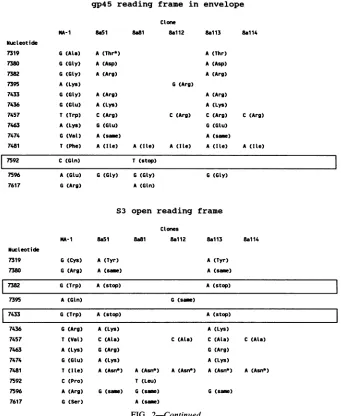
FIG. 2. Characterization of nucleotide and amino acid differences between MA-1 and Th-1 clones. The differencesinnucleotideand amino acid sequences between the MA-1 isolate of EIAV (6) and the sequenced Th-1 clones are indicated. For details on the sequence and numbering of EIAV MA-1, see reference 6. The regions of the Th-1 isolate analyzed are as follows: gp9O/S2 region, clone series 5a, nucleotides 5337to5780; gp9Oregion, clone series 7a, nucleotides 5789to6161, and clone series 7b, nucleotides 6162to6373;gp45/S3 region, cloneseries8a, nucleotides 7284to7643. Nucleotide changesoccurringinthepotentialcysteine-cysteine loop regionfrom nucleotide 6176
to6343suggestedtobehypervariableby Payneetal.(48)areboxed,asarethecoding changes givingprematurestopcodons.*,amino acids potentially involved inglycosylation. Two changed N-linkedglycosylation sitesareunderlined.
(48), could be confirmed. Second, the data indicated the
presence of another variable region in thegp9O protein and
also provided evidence, as previously suggested byus (6),
that the amino-terminal portion of gp9O and the S2 open
reading frame are highly conserved. Third, clones having
premature stop codons in either gp45 or S3 (rev) were
obtained. Finally,EIAV apparentlyconsistsofamixture of
virusespresentatthe sametime.
Variationof the envelope gp9O glycoprotein and the
over-lapping S2 open reading frame. Rapid sequence variation
followedbyepitope-specific selection of EIAV variants able to escapeimmune surveillance have been implied as
impor-tant factors in the pathogenesis of persistent infection and
disease induced byEIAV (13, 32, 38). Ofthe 16 nucleotide
changeslocated inthe gp9O envelopeglycoprotein regionof
the Th-1 clones analyzed, 15 resulted in changes in the
predicted amino acid sequence, representing4.4%
variabil-ityattheamino acid level (Fig. 2). Furthermore,6of the 15
changes were amino acids potentially involved in
glycosyl-ation (asparagine, serine, and threonine). The changes at nucleotide 5762fromanalanine inMA-1 and the otherTh-1
clones to a threonine in Th-i clone Sall and atnucleotide 6233 fromanasparticacid inMA-1andthe otherTh-1clones
to an asparagine in Th-1 clone 7b41 actually did create
N-linked glycosylation sites. Interestingly, of the 15amino acid differences in the gp9Oregion, 4 are within the
hyper-variable region described by Payne et al. (48). If the area
includingtheflanking cysteinesisincluded(nucleotides 6176
to 6343), makingthis area intoa potential loopresembling
hypervariable loops seen in HIV (21, 35, 45, 48), 7 amino
acids arevariable, givinga heterogeneityashighas7 of56
(12.5%) amino acids. Interestingly, one of the amino acid
changesinvolvedoneofthemultiplecysteines flankingthis
area. Moreover, four of thechanges inthis regionincluded
amino acidspotentially involved inglycosylation, and one
did in factchange anN-linked glycosylationsite. Also, the
silent nucleotide change at position 6331 involved an
N-linkedglycosylation site. Thesefindings indicate, as
pre-6186 A (Lys) C(Thr*)
6195 T (Leu) C (Pro)
6221 G (Val) A (Ile)
6233 G (ASD) A(Asn*)
6284 A(Thr*) C(Pro)
6291 G(Ser*) T(Mle) T (MIe) T( le) T (Mle) T(Ile) T (MIe) T (Ile)
6331 C (Asn*) T (same)
6342 G (Cys) A(Tyr)
on November 10, 2019 by guest
http://jvi.asm.org/
gp45 reading frame in envelope
Clone
MA-I 8a5a 8a81 8a112
A(Thr*)
A(Asp)
A (Arg)
A(Arg)
A(Lys)
C (Arg)
G(Glu)
A (smue)
A (lie) A (tIe)
8a113 8a114
A(Thr)
A (Asp)
A(Arg)
G(Arg)
A (Arg)
A(Lys)
C(Arg) C (Arg)
G (Glu)
A (same) A (Ite) A(Ite)
C (Arg)
A (lLe)
7592 C (Gin) T (stop)
7596 A (Glu) G(Gly) G (Gly) G (Gly)
7617 G(Arg) A(Gin)
S3 open reading frame
Clones
NA-1 8a51 8a81 8112 8a113 8al14
NucLeotide
7319 G(Cys) A (Tyr) A (Tyr)
7380 G(Arg) A (same) A(same)
7382 G (Trp) A (stop) A (stop)
7395 A(Gin) G(sm)
7433 G (Trp) A(stop) A(stop)
7436 G (Arg) A(Lys) A (Lys)
T (Val)
A(Lys) G(Glu)
T (lIe)
C (Pro)
A (Arg)
G (Ser)
C (Ala)
G(Arg)
A(Lys)
A (Asn*)
C (Ala) C (Ala)
G (Arg)
A (Lys) A (Asn*) A(Asn*) A (Asn*)
T (Leu)
G (sam) G (sam)
A (sa)
FIG. 2-Continued.
C (Ala)
A(Asn*)
G(same)
viously suggested by Payne etal. (48, 49), thatvariation of
aminoacidsequenceandchanges in glycosylationpatternin
thisarea of the viral envelope mayplay important roles in
the development of antigenic andbiological variant viruses
and in the pathogenesis of persistent EIAV infection and chronic recurrent disease. Furthermore, our data suggest
that this variation can occur early in infection and can
alreadybedetectedasgenotypic variationin thefirstfebrile
cycle. Another area ofvariability was found in gp9O from
nucleotide 5743 to 5895, inwhich fouramino acid changes
werefound. Thisareacontainsoneofthe changed N-linked
glycosylation sites and has multiple cysteinesatthe
bound-aries, and it is possible that this area represents another
hypervariable loop. In contrast, the amino-terminal part of
gp9Oand theS2openreadingframewerehighlyconserved.
Although the function and significance of the S2 open
reading frameareundetermined, these resultsmayindicate
stringent structural and functional requirements for this
regionof thegenome.
Genetic variation has been shown to occur in HIV and
other lentivirus infections. This variation can occur both
withinanindividualover aperiod of time andas amixture of genotypes present atthesametime(4, 7, 9,10, 20, 21, 25, 36,
40, 55, 56, 64). The different genotypes present have been
shown to vary in biological phenotype, and it has been shown that this variation inpartisdeterminedbysequences
inthe viralenvelope (21, 30, 65). Virulence determinants of
immunosuppressive isolates of feline leukemia virus have
also beenmappedtonucleotidechangesin the viralenvelope
gene (47, 50), and env gene sequences were found to be
important fortheproductionofleukemiainducedbyFriend
and Rauscher murine leukemia viruses (34, 37, 54, 59).
Furthermore,thedifference betweenmacrophage-tropicand
T-cell-tropic HIVhasbeenmappedtotheenvelope,
partic-ularlysequences nearand in thehypervariable loop V3(21,
35). Notably, our isolates, Th-1 and MA-1, differ in their
abilitiestoreplicate inmacrophages (7) and have consider-ablevariation in thehypervariable region, perhaps indicating
asimilar celltropismdeterminantintheenvelopeof EIAV.
Also, it is important to note that the degree ofvariability
found in theTh-1isolate canbe detectedonlywhenisolates
are analyzed before repeated cell culture passage. It has
Nucleotide
7319
7380
7382
7395
7433
7436
7457
7463
7474
7481
G (Ala)
G(Gly)
G(Gly)
A (Lys)
G (Gly)
G(Glu)
T(Trp)
A (Lys)
G (Val)
T (Phe)
7457
7463
7474
7481
7592
7596
7617
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.134.475.76.492.2]been shown for HIV and influenza virus that repeated passage will select for the fastest replicating virus without
thediverse pressure of in vivo biological and immunological
selection (21, 55). We PCR amplified and cloned our Th-1
EIAV after only one cell-free passage in HMC. Thus, our
clones should be representative of EIAV genotypes present
in vivo.
Analysis of the gp45 transmembrane glycoprotein. Of the13 nucleotide changes in the gp45 transmembrane protein re-gion analyzed, 12 resulted in amino acid changes (9.8% variability of the 123 amino acids), of which only one
involved potential glycosylation (Fig. 2). The biological
significance of most of the changes in this reading frame is at
present unclear. However, clone Th-18a81 had a stop codon
instead of a glutamine in this reading frame at nucleotide
7592, suggesting a premature termination of the gp45 at this
point. Interestingly, the stop codon in gp45 of clone Th-1 8a81 occurred in the same carboxy-terminal area of the
protein described for certain variants of EIAV passaged
repeatedly incultured equine kidney cells (52). This trunca-tion occurs close to the proposed cleavage site of gp45 which produces a 32-kDa glycosylated N-terminal segment and a
20-kDa nonglycosylated C-terminal fragment (52).
Trunca-tion at this point would suggest producTrunca-tion of only the
N-terminal segment. Furthermore, it has been shown that viruses having a truncated form of gp45 are more infectious
for cultured cells (52). In cell culture the stop codon was
changed from a glutamine or a glutamic acid codon, and
intriguingly, a stop codon changed from a glutamine codon
hasalsobeenreported in the transmembrane protein of a cell
culture-derived isolate of simian immunodeficiency virus
(31), exactly as reported here for EIAV in vivo. The
trun-cation of the simian immunodeficiency virus transmembrane protein only occurred when the virus was passaged in human
cells and not in monkey cells (31). A truncated transmem-brane protein for certain isolates of HIV types 1 and 2 has also recently been shown (22, 39). Our clones were derived from PCR-amplified DNA extracted from infected horse
monocyte/macrophage
cultures passaged only once. Themonocyte/macrophage lineage of cells is considered to be
the target cell for EIAV infection in vivo, and thus, our data
indicate thatlentiviruses having a truncated transmembrane
protein may play a role during in vivo infection. An exact
role of theC-terminal region of the lentivirus transmembrane
protein has not been defined; it is, however, tempting to speculate that this region is involved in viral replication or
cytopathogenicity in certain cells.
Analysis of theS3 (rev) open reading frame. In the S3 open
readingframe, overlapping the gp45 reading frame, only 7 of
the 13 nucleotide changes resulted in amino acid differences
and only 1 involved potential glycosylation. However, two
identical clones, Th-1 8a51 and 8a113, had two tryptophan
codonschanged to stop codons at nucleotides 7382 and 7433.
TheS3 open reading frame of EIAV is thought to encode a
lentivirustrans-acting regulatory protein termed rev (15, 51,
63). The rev protein of HIV and simian immunodeficiency
virus functions in the stability and transport of specific
incompletely spliced viral RNAs between the nucleus and
cytoplasm and isabsolutely required for expression of viral
structuralproteins. When functional rev is present,
cytoplas-mic RNAs consist primarily of unspliced genocytoplas-mic and
par-tially spliced env mRNAs. Without rev, cytoplasmic RNAs
consist ofmultiply spliced mRNAs (1, 18, 19, 26, 63).
Thus,
rev isimportant in the transition from early to late
transcrip-tion andreplication of thevirus (29). The stop codons in our
Th-1 clones both werechanged from tryptophan codons and
occurred early in the reading frame as described for in
vitro-derived rev-deficient phenotypes of EIAV (63). In the
twocellculture-derived clonespreviouslycharacterized, the
rev variants had either a 2-base insertion followed by a
tryptophan-to-stop codon change or a4-basedeletion.
Inter-estingly, this deletion occurred only 8 bases from the stop
codon in our Th-1 clones. It has been shown that cell
cultures infected with rev-deficient EIAV produce no
infec-tious virus and release few or noparticles from theinfected
cells. Furthermore, such infected cells contain
predomi-nantlymultiplyspliced RNA, and no full-lengthor envRNA
can be detected in the cytoplasm (63).
The Th-1 clones containing the rev stop codons also
contained an upstream nucleotide substitution atnucleotide
7319 which changed oneof the twocysteineresiduesin
S3
toa tyrosine (Fig. 2). This is interesting considering that the
cysteine residues in retroviruses, including EIAV, are
usu-ally highly conserved (48). Taken together, these data
indi-cate that our clones represent sequences from EIAV
ge-nomes which are deficient in rev function (63) and that the
rev-deficient phenotype appears to relieve the pressure on
conservation of the twocysteines in this gene. Furthermore,
rev-deficient clones, whether obtained in vivo or in vitro,
seem to have a similar pattern of truncation, suggesting that
either an underlying basal mechanism for stop codon
intro-duction in this particular area, or, more interestingly, a
truncated
S3
open reading frame product, perhaps havingonly one cysteine residue, may have a biological significant
function. rev-deficient viruses do not express viral proteins
at the cell surface and therefore can remain undetected by
the host immune system. Also, such infected cells do not
cause viral interference and can thus be reinfected with
replication-competent viruses. Therefore, rev-deficient
vi-ruses may play important roles in maintaining
persistent
infection and in recurrent disease. Until a comprehensive
and detailed transcriptional, translational, and functional
map including several isolates and phenotypes of EIAV is prepared, these intriguing questions cannot be answered.
Our data together with the emerging body of evidence from other lentivirus infections indicate that a heterogeneous population of EIAV genotypes may coexist in vivo and that genotypes of different antigenic makeup, cell tropism, cyto-pathogenicity, and replicative capabilities together initiate and sustain the development of persistent infection and chronic disease. Thus, it may be possible, perhaps by the development of pseudotypes or by protein complementa-tion, that the coexistence of replication-competent and -de-fective viruses infecting the same cell permits the amplifica-tion and transmission of the defective genotype, a scenario well known from avian and murine leukemia viruses and recently described for certain isolates of feline leukemia
virus (47).
ACKNOWLEDGMENTS
We thank Lene Arndrup Poulsen for excellent technical assis-tance.
This work was supported in part by grants to S.A. from the Danish Center for Animal Biotechnology Research, the Danish Agricultural and Veterinary Research Council, and the
Bernhard
Bang Foundation and by a grant to S.C. from the Iowa State University Agricultural Biotechnology Council.
REFERENCES
1. Ahmed, Y. F., S. M. Hanly, M. H. Malim, B. R.Cullen, and W. C. Greene. 1990. Structure-function analyses of the
HTLV-I
Rex and HIV-1 Rev RNA response elements: insights into the mechanism of Rex and Rev action. Genes Dev.4:1014-1022.on November 10, 2019 by guest
http://jvi.asm.org/
2. Alexandersen, S., M. E. Bloom, and S. Perryman. 1988.Detailed transcription map of Aleutian mink disease parvovirus. J.Virol. 62:3684-3694.
3. Arya, S. K., C. Guo, S. F. Josephs, and F. Wong-Staal. 1985. trans-activator gene of human T-lymphotropic virus type III (HTLV-III). Science 229:69-73.
4. Asjo, B., L. Morfeldt-Manson, J. Albert, G. Biberfeld, A. Karisson, and K. Lidman. 1986. Replicative capacity of human immunodeficiency virus from patients withvaryingseverity of HIV infection. Lancetii:660-662.
5. Bloom, M. E., S. Alexandersen, S. Perryman, D. Lechner, and J. B. Wolfinbarger. 1988. Nucleotide sequence and genomic organization of Aleutian mink disease parvovirus (ADV): se-quence comparisons between anonpathogenicand apathogenic strain of ADV. J. Virol. 62:2903-2915.
6. Carpenter, S., S. Alexandersen, M. J. Long, S.Perryman,andB. Chesebro. 1991. Identification of ahypervariable region in the long terminal repeat of equineinfectiousanemiavirus. J. Virol. 65:1605-1610.
7. Carpenter, S., and B. Chesebro. 1989. Change in host cell tropism associated with in vitro replicationof equineinfectious anemia virus. J. Virol. 63:2492-2496.
8. Carpenter, S., L. H. Evans, M.Sevoian,and B.Chesebro.1987. Role of host immune responseinselectionofequineinfectious anemia virus variants. J. Virol.61:3783-3789.
9. Cheng-Mayer, C., D. Seto, M. Tateno, and J. A. Levy. 1988. Biologic features of HIV-1 that correlate withvirulence inthe host. Science240:80-82.
10. Chiodi, F., A. Valentin, B. Keys, S. Schwartz, B. Asjo, S. Gartner, M. Popovic, J. Albert, V.-A. Sundqvist, and E.-M. Fenyo. 1989. Biological characterization ofpaired human immu-nodeficiency virus type1isolates from blood and cerebrospinal fluid. Virology 173:178-187.
11. Chiu,I. M., A. Yaniv, J. E. Dahlberg, A. Gazit,S. F. Skuntz, S. R. Tronick,and S.A.Aaronson. 1985. Nucleotide sequence evidence for relationship of AIDS retrovirus to lentivirus. Nature (London)317:366-368.
12. Clements, J. E.,N.D'Antonio, and0.Narayan. 1982.Genomic changes associated with antigenic variation of visna virus. II. Commonnucleotidesequencechanges detected invariants from independent isolations. J.Mol. Biol. 158:415-434.
13. Clements,J. E.,S.L.Gdovin,R. C.Montelaro,and 0. Narayan. 1988. Antigenic variation in lentiviral diseases. Annu. Rev. Immunol. 6:139-159.
14. Derse, D., P. L. Dorn,L.Levy, R.M.Stephens, N. R. Rice, and J. W. Casey. 1987.Characterizationof equineinfectiousanemia virus long terminal repeat. J.Virol. 61:743-747.
15. Dorn,P., L. DaSilva,L.Martarano,and D. Derse. 1990. Equine infectious anemiavirustat:insights into the structure,function, and evolution of lentivirus trans-activator proteins. J. Virol. 64:1616-1624.
16. Duh, E.J.,W. J.Maury, T. M. Folks, A. S. Fauci, and A. B. Rabson. 1989. Tumornecrosis factor a activates human immu-nodeficiency virus type 1 through induction of nuclear factor binding to theNF-KB sites in the long terminal repeat. Proc. Natl. Acad. Sci. USA86:5974-5978.
17. Evans, K.S.,S.L.Carpenter, and M. Sevoian. 1984. Detection ofequine infectious anemia virus in horse leukocyte cultures derived fromhorsesinvariousstages ofequine infectius anemia viralinfection. Am. J. Vet. Res. 45:20-25.
18. Felber, B. K., C. M. Drysdale, and G. N. Pavlakis. 1990. Feedback regulationofhuman immunodeficiency virus type 1 expressionbythe Revprotein. J.Virol. 64:3734-3741. 19. Felber, B. K., M.Hadzopoulou-Cladaras,C. Cladaras, T.
Cope-land, and G. N. Pavlakis. 1989. revproteinofhuman immuno-deficiency virustype 1 affects the stability and transportofthe viralmRNA.Proc. Natl. Acad. Sci. USA 86:1495-1499. 20. Fenyo, E. M., L. Morfeldt-Manson, F. Chiodi,B. Lind,A. von
Gegerfelt, J. Albert, E. Olausson, and B.Asjo. 1988. Distinct replicative andcytopathic characteristics of human immunode-ficiencyvirus isolates. J. Virol. 62:4414 4419.
21. Fisher, A.G., B. Ensoli, D. Looney, A. Rose,R.C.Gallo,M.S. Saag, G. M. Shaw, B. H. Hahn, and F. Wong-Staal. 1988.
Biologically diverse molecular variants within a
single
HIV-1isolate. Nature (London)334:444-47.
22. Franchini, G.,and M. L. Bosch.1989.Genetic relatedness ofthe
humanimmunodeficiencyviruses type1 and 2(HIV-1, HIV-2)
andthesimianimmunodeficiencyvirus(SIV).Ann. N.Y.Acad. Sci. 554:81-87.
23. Golub,E.I.,G.Li,and D. L.Volsky.1990. Differencesin basal
activity of the long terminal repeat determine the different
replicative capacitiesoftwoclosely related human
immunode-ficiencyvirustype 1isolates. J. Virol. 64:3654-3660.
24. Gonda,M. A., M. J. Braun, J. E. Clements,J. M.
Pyper,
F.Wong-Staal,R.C.Gallo,and R. V.Gilden.1986.HumanT-cell
lymphotrophic virustype III shares sequence
homology
withafamily ofpathogenic lentiviruses. Proc. Natl. Acad. Sci. USA
83:4007-4011.
25. Hahn,B. H.,G. M. Shaw,M. E.Taylor, R. R.Redfield,P.D.
Markham, S. Z.Salahuddin,F. Wong-Staal,R. C.
Gallo,
E.S.Parks,and W. P. Parks.1986.Genetic variation inHTLV/LAV overtime in patients with AIDS oratrisk for AIDS. Science 232:1548-1553.
26. Hanly,S.M.,L. T.Rimsky,M. H.Malim,J.H.Kim,J.Hauber,
M. D. Dodon,S.-Y. Le,J. V. Maizel,B. R. Cullen,andW. C. Greene. 1989.
Comparative
analysis
of the HTLV-IRex and HIV-1 Rev trans-regulatoryproteins
and theirRNA response elements. GenesDev. 3:1534-1544.27. Issel, C. J., and L. Coggins. 1979.
Equine
infectious anemia:currentknowledge.J. Am. Vet. Med. Assoc. 174:727-733.
28. Kawakami, T., L. Sherman, J. Dahlbert, A. Gazit, A. Yaniv,
S. R. Tronick,and S. A. Aaronson. 1987. Nucleotide sequence
analysis ofequine infectiousanemia
proviral
DNA.Virology
158:300-312.
29. Kim, S., R. Byrn, J. Groopman, and D. Baltimore. 1989.
Temporal aspects ofDNA and RNA
synthesis during
humanimmunodeficiencyvirusinfection: evidence for differentialgene
expression. J.Virol. 63:3708-3713.
30. Kim, S., K. Ikeuchi, J. Groopman, and D. Baltimore. 1990. Factorsaffectingcellular
tropism
ofhumanimmunodeficiency
virus.J. Virol.64:5600-5604.
31. Kodama, T., D. P. Wooley, Y. M. Naidu, H. W. Kestler III,
M.D.Daniel,Y.Li,and R.C. Desrosiers. 1989.
Significance
ofpremature stop codons in env of simian
immunodeficiency
virus.J. Virol. 63:4709-4714.
32. Kono,Y.,K.Kobayashi,and Y.
Fukunaga.
1973.Antigenic
driftofequineinfectiousanemiavirusin
chronically
infectedhorses. Arch. GesamteVirusforsch. 41:1-10.33. Li,Y.,E.Golemis,J.W.
Hartley,
and N.Hopkins.
1987. Diseasespecificity ofnondefective Friend and
Moloney
murine leuke-mia viruses iscontrolledby
asmall number of nucleotides. J.Virol.61:693-700.
34. Linemeyer,D.L.,J.G.Menke,S. K.Ruscetti,L.H.Evans,and
E. M. Scolnick. 1982.
Envelope
gene sequences which encode thegpS2proteinofspleen
focus-forming
virus arerequired
forthe induction of
erythroid
cellproliferation.
J. Virol. 43:223-233.35. Liu, Z. Q., C. Wood, J. A. Levy, and C.
Cheng-Mayer.
1990.Theviralenvelopegeneisinvolved in
macrophage tropism
ofahuman
immunodeficiency
virustype1strain isolatedfrombrain tissue.J. Virol.64:6148-6153.36. Lutley,R.,G.Petursson,P. A.Palsson, G.
Georgsson,
J.Klein,andN.Nathansson. 1983.
Antigenic
drift in visna: virus variationduring long-term infection of Icelandic
sheep.
J. Gen. Virol. 64:1433-1440.37. Machida,C. A.,R. K.Bestwick,andD. Kabat. 1985. Aweakly
pathogenic Rauscher
spleen focus-forming
virus mutant that lacks thecarboxyl-terminal
membrane anchor ofitsenvelope
glycoprotein. J. Virol. 55:990-993.
38. McGuire, T. C., K. O'Rourke, and W. P. Cheevers. 1987. A
reviewof
antigenic
variationby
equine
infectious anemiavirus. Contrib. Microbiol. Immunol.8:77-89.39. McNearney,T.,P.Westervelt,B.J.Thielan, D.B. Trowbridge,
J.Garcia, R.Whittier,andL. Ratner. 1990. Limited sequence
heterogeneity among
biologically
distinct humanimmunodefi-ciency virus type 1 isolates from individuals involved in a
on November 10, 2019 by guest
http://jvi.asm.org/
clustered infectious outbreak. Proc. Natl. Acad. Sci. USA 87:1917-1921.
40. Meyerhans, A., R. Cheynier, J. Albert, M. Seth, J. Kwok, L. Sninsky, L. Morfeldt-Manson, B. Asjo, and S. Wain-Hobson. 1989.Temporalfluctuationsin HIVquasispeciesin vivo are not reflected bysequential HIV isolation. Cell 58:901-910. 41. Montagnier,L., C. Dauguet, C. Axler,S.Chamaret, J. Gruest,
M. T.Nugeyre,R. Rey, F.Barr*-Sinoussi, and J. C. Chermann. 1984. A new typeof retrovirus isolated frompatientspresenting with lymphadenopathy and acquired immune deficiency syn-drome: structural andantigenic relatedness with equine infec-tiousanemiavirus. Ann. Virol. 135E:119-134.
42. Montelaro, R.,J. Ball, P. Rwambo, and C. Issel. 1990. Antigenic variationduring persistent lentivirus infectionsand its implica-tions for vaccinedevelopment. Adv. Exp. Med.Biol. 251:251-272.
43. Montelaro, R. C., W. G. Robey, M. D. West, C. J. Issel, and P.J. Fischinger. 1988.Characterization oftheserological cross-reactivity between glycoproteins of the human immunodefi-ciencyvirusandequineinfectious anemia virus. J. Gen. Virol. 69:1711-1717.
44. Narayan, O., D. E. Griffin, and J. Chase. 1977.Antigenic shift of visnavirus inpersistently infected sheep. Science197:376-378. 45. Neurath, A.R., and N. Strick. 1990.Confronting the hypervari-ability ofanimmunodominant epitope eliciting virus neutraliz-ing antibodies from the envelope glycoprotein of the human immunodeficiency virustype 1(HIV-1). Mol. Immunol. 27:539-549.
46. Osborn, L., S. Kunkel, and G. J. Nabel. 1989. Tumornecrosis factor a and interleukin 1 stimulate the human immunodefi-ciency virus enhancer by activation of the nuclear factor KB. Proc. Natl.Acad. Sci. USA 86:2336-2340.
47. Overbaugh, J., P. R. Donahue, S. L. Quackenbush, E. A. Hoover, and J. I.Mullins. 1988. Molecularcloning ofafeline leukemia virus that induces fatal immunodeficiencydisease in cats. Science239:906-910.
48. Payne, S. L., F.-D. Fang, C.-P. Liu, B. R. Dhruva, P.Rwambo, C. J. Issel,and R. C. Montelaro. 1987. Antigenic variationand lentivirus persistence: variations in envelope gene sequences duringEIAVinfectionresemblechanges reportedforsequential isolatesof HIV.Virology 161:321-331.
49. Payne, S. L., K. Rushlow, B. R. Dhruva, C. J. Issel, and R. C. Montelaro. 1989. Localization of conserved and variable anti-genic domains of equine infectious anemia virus envelope glycoproteins usingrecombinantenv-encoded protein fragments produced inEscherichiacoli. Virology 172:609-615.
50. Poss, M. L., J. I. Mullins, and E. A. Hoover. 1989. Posttransla-tionalmodifications distinguish the envelope glycoprotein of the immunodeficiency disease-inducing felineleukemia virus retro-virus. J. Virol. 63:189-195.
51. Rasty,S., B. R. Dhruva, R. L. Schlitz,D. S. Shih, C. J. Issel, and R.C. Montelaro. 1990.Proviral DNA integration and transcrip-tionpatterns of equine infectious anemia virusduring persistent andcytopathic infections. J.Virol.64:86-95.
52. Rice, N. R., L. E. Henderson, R. C.Swoder, T. D. Copeland, S. Oroszlan,andJ.F.Edwards. 1990.Synthesisandprocessing of
the transmembrane envelope protein of equine infectious ane-mia virus. J. Virol. 64:3770-3778.
53. Rushlow, K., K. Olsen, G. Steigler, S. L. Payne, R. C. Mon-telaro, and C.J. Issel. 1986. Lentivirus genome organization: the complete nucleotide sequence of the env gene region ofequine infectious anemia virus. Virology155:309-321.
54. Ruta, M., R.Bestwick, C.Machida, and D. Kabat. 1983. Loss of leukemogenicity caused by mutations in the membrane glyco-protein structural gene of Friend spleen focus-forming virus. Proc.Natl. Acad. Sci. USA 80:4704-4708.
55. Saag, M. S., B. H. Hahn, J. Gibbons, Y. Li, E. S. Parks, W. P. Parks, and G. M. Shaw. 1988. Extensive variation of human immunodeficiency virus type-1 in vivo. Nature (London) 334: 440-444.
56. Sakai, K., S. Dewhurst, X. Ma, and D. J. Volsky. 1988. Differ-ences incytopathogenicity and host cell range among infectious molecular clones of human immunodeficiency virus type 1 simultaneously isolated from an individual. J. Virol. 62:4078-4085.
57. Sanger, F., S.Nicklen, and A. R. Coulson. 1977. DNA sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA74:5463-5467.
58. Short, M. K., S. A. Okenquist, and J. Lenz. 1987. Correlation of leukemogenic potential of murine retroviruses with transcrip-tional tissue preference of the viral long terminal repeat. J. Virol. 61:1067-1072.
59. Sitbon, M., B. Sola, L. Evans, J. Nishio, S. F. Hayes, K. Nathanson, C. F. Garon, and B. Chesebro. 1986. Hemolytic anemia anderythroleukemia, two distinct pathogenic effects of Friend MuLV:mapping of the effects to different regions of the viralgenome.Cell47:851-859.
60. Sodroski, J., R. Patarca, C. A. Rosen, F.Wong-Staal,and W. A. Haseltine. 1985. Location of the trans-acting region on the genome of human T-cell lymphotropic virus type III. Science 229:74-77.
61. Sodroski,J., C. A. Rosen, F.Wong-Staal, S. Z. Salahuddin, M. Popovic, S. Arya, R. C. Gallo, and W. A. Haseltine. 1985. Transacting transcriptional regulation of human T-cellleukemia virustype IIIlongterminalrepeat. Science227:171-173. 62. Stephens, R. M., J. W. Casey, and N. R. Rice. 1986. Equine
infectious anemiavirusgag andpol genes: relatedness tovisna andAIDS virus. Science 231:589-594.
63. Stephens, R.M., D. Derse, and N. R. Rice. 1990. Cloningand characterization of cDNAs encoding equine infectious anemia virusTat andputative Revproteins.J. Virol.64:3716-3725. 64. Tersmette, M., R. A. Gruters, F. DeWolf, R. W. Y. DeGoede,
J. M. A. Lange, P. T. A. Scheliekens, J. Goudsmit, H. G. Huisman, and F. Miedema. 1989.Evidence forarole ofvirulent humanimmunodeficiency virus (HIV)variants in the pathogen-esis of acquired immunodeficiency syndrome: studies on se-quentialHIVisolates. J. Virol.63:2118-2125.
65. York-Higgins, D., C. Cheng-Mayer, D. Bauer, J. A. Levy, and D. Dina.1990.Humanimmunodeficiency virustype 1cellular host range,replication, andcytopathicityarelinked to theenvelope regionof the viral genome. J. Virol.64:4016-4020.