JOURNALOFVIROLOGY, Mar. 1975, p.619-635 Copyright01975 AmericanSocietyforMicrobiology
Vol. 15, No. 3 Printed in U.S.A.
Role
of
Simian Virus
40
Gene A Function
in
Maintenance
of
Transformation
JOAN S. BRUGGE MDJANET S. BUTEL*
Department of Virology and Epidemiology, Baylor College of Medicine, Houston, Texas 77025 Received forpublication3September1974
Mouse, hamster, and human cells were transformed at the permissive
temperatureby mutantsfrom simian virus 40(SV40) complementationgroupA
in order to ascertain the role of the geneA function in transformation. The
following parameters of transformation were monitored with the transformed
cells under permissive and nonpermissive conditions: morphology; saturation
density; colony formationonplastic,oncellmonolayers, andinsoftagar; uptake
ofhexose; and the expression ofSV40tumor(T) and surface (S)antigens. Cells
transformed by the temperature-sensitive (ts) mutantsexhibited thephenotype
oftransformed cells at the nonrestrictive temperature for all oftheparameters
studied. However, when grown at the restrictive temperature, they were
phenotypically similar to normal, untransformed cells. Growth curves showed
that the ts A mutant-transformed cells exhibited the growth characteristics of
wild-type virus-transformed cells atthepermissivetemperature andresembled
normal cells when placed under restrictive conditions. Therewere 3- to 51-fold
reductions in the levels of saturation density, colony formation, and uptake of
hexose when the mutant-transformed cellswere grownattheelevated
tempera-ture as compared to when they were grown at the permissive temperature.
Mutant-transformed cells from the nonpermissive temperature were able to
produce transformed foci when shifted downtopermissiveconditions, indicating
that thephenotypically reverted cellswerestill viable andthat thereversionwas
areversibleevent. SV40Tantigenwaspresent in the cells at bothtemperatures,
but S antigen was not detected in cells maintained at the nonpermissive
temperature. All ofthewild-typevirus-transformed cells exhibitedatransformed
phenotype when grown under either restrictive or nonrestrictive conditions.
These results indicate that the SV40 group A mutant-transformed cells are
temperaturesensitiveforthe maintenance ofgrowthproperties characteristic of
transformation. Virus rescued fromthe mutant-transformed cells by the
trans-fection method wasts, suggestingthatthe SV40geneA function, rather thana
cellularone, isresponsible forthetsbehavior ofthe cells.
The mechanism by which oncogenic viruses
mediate cellular transformation is stillobscure.
Furthermore, it remains to be definitively
es-tablished
that the DNA tumor viruses controleventsintransformationby the synthesisof one
or more specific viral gene products. One
ap-proach todissecting this complex phenomenon
involves the use of temperature-sensitive (ts)
mutants of a virus. If cells which have been
transformed
by
a tsmutantdisplayatempera-ture-dependent
transformed phenotype, themutant gene function would be implicated as
being involved in the transformation process.
"Temperature-dependent transformed
pheno-type"
isdefinedas a conditionalstate inwhichcharacteristics oftransformation are expressed
by the transformed cellsat apermissive
temper-ature, but are not manifest at anonpermissive
temperature,thereby causing the cellsto revert
tonormalgrowth behavior.
Since only early viral functions appear tobe
expressed in simian virus 40
(SV40)-trans-formed cells (2, 5, 27), it is mostreasonable to
examine mutants ofthis virus whichare
defec-tiveinanearlygene for theirabilityto maintain
the transformed state. Onlyone group ofearly
SV40mutantshas been isolated
(complementa-tion group A). During infection ofsusceptible
monkey cells at the restrictive temperature,
group A mutants synthesize SV40 tumor (T)
antigen and stimulate host cell DNA
replica-tion, but fail to produce viral DNA or capsid
619
on November 10, 2019 by guest
http://jvi.asm.org/
BRUGGE AND BUTEL
antigens (31). More specifically, the gene A
protein has been found to be required for the
initiation ofviral DNA synthesis (29).
To determine whether the SV40geneA
func-tion is involved in the maintenance of
transfor-mation, wetransformed normal cells withgroup
A mutants and then examined the cells after
growth at the permissive and nonpermissive
temperatures for several of the classic
parame-tersoftransformation: morphology; saturation
density; colony formation on plastic, on
mono-layers of normal cells, and insoftagar; uptake
of 2-deoxy-D-glucose (2-D-G); and the
expres-sion of SV40 T and surface(S) antigens.
Multi-ple parameters were monitored to determine
whether loss of the SV40geneAfunction atthe
nonpermissivetemperatureaffectedall, several,
ornoneof thecharacteristicsofthetransformed
state.Basedonresultsfrom the abovetests,itis
concluded that the continual expression of the
gene A function of SV40 is required for the
maintenance of transformation.
MATERIALS AND METHODS
Viruses. SV40 Group Ats mutantsA7, A28, A30, andA58, aswellas theWT-2strain, weregraciously
providedby PeterTegtmeyer (29, 31).The wild-type (WT) Baylor reference strain which had been plaque-purified three times at 40.5C was also used. Virus
stockswerepreparedat33C by inoculating confluent monolayers of CV-1 cells (obtained from Saul Kit)at
a multiplicity of infection of 0.01 PFU/cell. When cytopathic effects involved75to100%of thecells, the cultures were disrupted by three cycles of quick
freezing and thawing, the cell lystateswere clarified
by low-speed centrifugation, and the supernatant fluids were stored at -70C. Virus assays were
per-formedinBSC-1 cells asdescribedpreviously (4).
Cells. The followingnormal cells wereemployed:
human skin cells derivedfromapatientwith Fanconi
anemia, obtained from P. Glade, and designated human Fanconi (HuF) cells; hamster embryo fibro-blasts (HEF) prepared by trypsinizing minced
em-bryos from Syrian hamsters (Con-Olson Co., Inc., Madison,Wisc.); andmouseBALB-3T3 cells(kindly
provided by Wade Parks). Inaddition, theH-50cell line, derived fromahamstertumorinduced in vivoby SV40 (1, 4),andthe VLMcells, derivedfromBALB/c
mouseembryo cells transformedinvitrobySV40and
obtained from S. S. Tevethia (36), were used. With
theexception ofthe BALB-3T3 cells, allthecelllines
were grown at 37C inEagle medium supplemented with 10% fetal bovineserum(FBS), 100Uof
penicil-lin and 100 jigof streptomycin per ml, and 0.075%
sodiumbicarbonate. TheBALB-3T3 cellswere grown
inDulbecco's modification of Eagle medium (Grand Island Biological Co., Grand Island, N.Y.) supple-mented with10%FBS, 100Uofpenicillin and100gg ofstreptomycinperml, and0.3%sodiumbicarbonate
at33C inanatmosphereof 10%CO2.The derivation
ofcell lines transformedby WT and tsAmutantsof SV40 is described below.
Saturation density experiments. Cellsgrowingat the permissivetemperatureweretrypsinized, and1x
101to 2 x 106 cellswere seeded into 35-mm plastic
petri dishes. Replicatecultures were then incubated
ateither the permissiveorthe nonpermissive temper-ature in an atmosphere of CO2. Cell counts were
performedonduplicate sampleseveryotherday, and
the mediawaschangedeverythirdday. Aplateau in
cellcountwas consideredtorepresent thesaturation density.
Colony formation methodology. (i) On
mono-layers of normal cells. Cells growingatthe permis-sive temperature were trypsinized and plated at
several concentrations, rangingbetween 102 and 109 cells/plate,ontoconfluent monolayers of human Fan-coni cells in35-mmplastic petri dishes. Cultureswere
incubated in 5% CO,at the permissive and
nonper-missive temperatures, and the media were changed
every third day. After8 days at the nonpermissive temperatureand 10 daysat thepermissive tempera-ture, colonies were stained with Giemsa asfollows:
cells were washed three times with Tris-buffered
saline(TBS) (pH 7.4)and fixed for 10mininabsolute methanol. The cultureswerethenexposed for10to30 mintoGiemsa staining solution whichconsisted of 5 ml ofGiemsa stock (Matheson, Coleman, and Bell,
Norwood, Ohio), 3ml of0.1Mcitricacid,3ml of 0.2 M Na2HPO4, and 100 ml ofdistilled water, washed withTBS, and air dried.
(ii) Onplastic. Thesameprocedure wasfollowed as describedabove, exceptthat the cellswereseeded into empty plastic petri dishes rather than onto monolayersofnormalcells.
(iii) In soft agar. Thetechnique of Macpherson andMontagnier (19)wasfollowed. Basallayers (2 ml)
of 0.5% agar (Difco) supplemented with Eagle
me-dium, 10% FBS, and0.2% sodium bicarbonate were
formed in 35-mmplastic petri dishes,followedbythe addition of a second layer (1.5 ml) of 0.3% agar
supplemented as above, containingvarious dilutions
of cells (between 102 and 10S cells/plate) which had been growing at the permissive temperature. Plates
were then incubated atthe permissive and the
non-permissive temperatures in atmospheres of5% CO2. Colonycountswereperformedafter7 to10dayswith alowpowermicroscope.
Uptake of hexose. Toassaytheuptakeof2-D-G, cells which had been passed at least twiceat either thepermissiveorthenonpermissivetemperaturewere
trypsinized and seeded into 35-mm plastic petri dishes (1 x 105to 2 x 105cells/culture). Theplates were then incubated atthe appropriatetemperature until the monolayers were 75% confluent. At that time, the cells werewashed withphosphatebuffered
saline (PBS; pH 7.2) at 37C, and the plates were
floatedina37Cwaterbath. PBS (2ml) containing1 uCi of [3H]2-D-G was added to each plate. After 10 min ofincubation, the cells were washed with cold
PBS andscrapedinto1ml ofPBS,and 0.1 ml of this suspensionwascountedusingtoluenecontaining4g
of 2,5-diphenyloxazole and 0.4 g of
1,4-bis-620
J.VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
SIMIANVIRUS 40 GENEA FUNCTION. II.
(5-phenyloxazolyl)-benzene per liter and 10% (vol/
vol) BBS-3 (Beckman Instruments, Fullerton, Calif.), inaliquid scintillation spectrometer(Model LS-250; Beckman Instruments).
Immunofluorescence techniques. Cells grown on
15-mmroundcoverglasseswereharvested,fixed, and
stained asdescribed previously (23). SV40 Tantigen wasdetected withserafromhamstersbearingtumors
induced bySV40-transformed(H-50)cells.The ham-ster serum was followed by fluorescein-labeled
anti-hamster baboon globulin. SV40 S antigen was
de-tected by a method described previously (25).The S
antibody was from hamsters immunized four times
withPARA-adenovirus 7-transformedmarmosetcells (S. S. Layne and F. Rapp, Bacteriol. Proc., p. 155,
1969).
Rescue of SV40 from transformed cells. Cellular DNA was extracted using the modified Hirt (13)
procedure and passedinAfricangreenmonkey kidney
cells following the method described by Boyd and Butel(4).Therescued viruswaspassedasecondtime in CV-1 cells and then assayed by the plaque tech-nique at the permissive and nonpermissive tempera-tures.
RESULTS
Establishment of cell lines transformed by group A mutants of SV40. Cells originating
from three different speciesweretransformedin
vitro with SV40tsmutants to comparethe role
of the gene A function in cells which are
normally nonpermissive (mouse, hamster) with
those whicharesemipermissive (human) forthe
replication of SV40. Cells were transformed as
follows: confluent monolayers were infected
with 5to10PFU of virus percell. After90min
of adsorption at 37 C, the cells were flooded
with media and incubated at the permissive
temperature for 24 h, at which time they were
trypsinized and subcultured. After
approxi-mately six to eight passages, transformed foci
appeared in both the mouse BALB-3T3 cells
and the human Fanconi cells; several passages
later, those cultureswere 100% SV40 T antigen
positive. The normal BALB-3T3 and HuF cells
exhibited a high degree of contact inhibition.
Thispropertymade iteasytodistinguish
trans-formed foci from normal cells. In contrast, the
HEF cellswere not ashighly contact inhibited
and did notdisplay such areadily
distinguisha-ble change in morphology after transformation.
In addition to the lines transformed in tissue
culture, newborn Syrian hamsters were
inocu-lated with 1 x 106 PFU ofSV40tsA7; tumors
which developed approximately 6 months later
wereexcised and trypsinized, and the cellswere
established in culture.
Table 1 is a summary of pertinent
informa-tion onthe cell lines derived for this
investiga-tion. Thedesignation of each cell line indicates
the origin of the cells as well as the strain of
SV40 used for transformation. Thetemperature
TABLE 1. Origin and designation ofSV40-transformed cell lines employed in this study Experimental StrainofSV40 Designationof Currentno. Passage no. temp (C) Hostcell origin used for transformed ofin vitro
when
100%transformation cell line passages Tantigen Per- pNonis-missive sive
In vitro Mouse
BALB-3T3 tsA7 Balb/A7 60 10 33 39.5
BALB/cembryo fibroblasts WT-Baylor Balb/WT-VLM 67 10 33 39.5
Hamster
Hamsterembryofibroblasts WT-2 Ha/WT-2 35 21 37 40.5
ts A28 Ha/A28 30 18 37 40.5
tsA58 Ha/A58 43 23 37 40.5
tsA30 Ha/A30 36 22 37 40.5
Human
Fanconiskincells WT-2 HuF/WT-2 20 8 37 40.5
ts A28 HuF/A28 25 8 37 40.5
In vivo
Syrianhamster(Con-Olsen) WT-Baylor H-50 >100 1 33 39.5
ts A7 HaTu/A7-1 20 1 33 39.5
HaTu/A7-2 20 1 33 39.5
VOL.15, 1975 621
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.504.52.447.409.662.2]BRUGGE AND BUTEL
selected asthe permissive condition reflects the
temperature at which the original normal cell
line was carried. The permissive temperature
for the mutant-induced hamster tumor cells
and the normal BALB-3T3 cells was 33 C. The
nonpermissivetemperatureforthesetwogroups
ofcellswas 39.5C. Withboth the hamster and
human cells transformed invitro, it was
neces-sary to employ 40.5 C for the nonpermissive
temperature to detect phenotypic reversion;
37
C
was used for permissivegrowth.
Thisvariation in the temperature required for
non-permissive behavior is notunprecedented. Graf
andFriis (10) also foundahost-range difference
innonpermissiveconditionswith their
tempera-ture-sensitive Rous sarcoma virus-transformed
cells. It would appear that the biochemical
environment withincellsfromdifferent species
affects the heatsensitivityofproteins to
differ-ent extdiffer-ents. In addition, Kachani and
Sabin
(16)observed that hamster cells transformedin
vitro by SV40 were more heat resistant than
either normal hamster cellsorcells from
ham-ster tumors inducedin vivo.
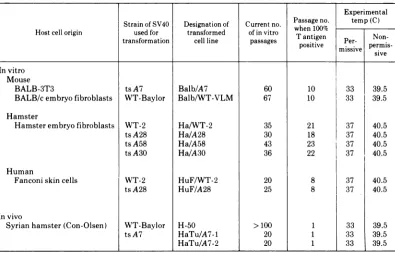
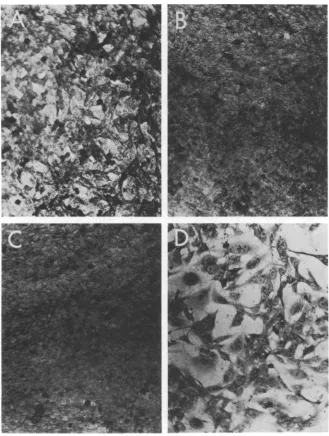
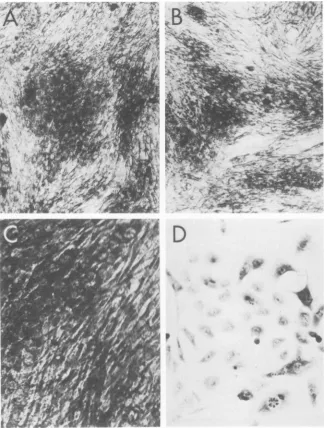
Morphology of transformed cells. The
typi-cal morphology of WT and group A
mutant-transformedcellsatthepermissive and
nonper-missive temperatures isillustratedinFig.1to 3.
The group A mutant transformed cells grown
underpermissive conditions form multiple
lay-ersofcells witharounded
morphology
(Fig.1C,
2C, and 3C), whereas at the nonpermissive
temperaturethe cellsarefibroblastic and
highly
contactinhibited
(Fig. 1D,
2D,
and3D),
resem-bling
the morphology ofnormal cells from thesame species. This difference in
morphology
isnoteasilyrecognizable until the cells have been
passed once atthehightemperature. The cells
can be subcultured many passages at the
non-permissive temperature and will retain the
morphology and
growth
characteristics ofnor-mal cells. Ifthe cells are shifted back to the
permissive temperature,
they
return to theiroriginal
transformedmorphology
within2days.
WT-transformed cells of all three species
ex-hibit the same morphology at both
tempera-tures (Fig. 1A, 1B; 2A, 2B; 3A, and 3B).
Saturation density. Transformed cells do
not exhibit density-dependent growth control
and are able to continue dividing in culture
until thenutritional factorshave been depleted
from the media orthe cells slough. As a
conse-quence of this behavior, transformed cells
achieve higher saturation densities than do
normal cells which are contact inhibited. The
saturation densities of the transformed cells
were assayed to determine whether a decrease
in saturation density accompanied the shift of
mutant-transformed cellstothe restrictive
con-dition. Growthcurves of the normal and
trans-formed mouse, hamster, and human cells are
shown in Fig. 4. It can be seen that WT
virus-transformed cells grew more rapidly and
achieved higher saturation density levels than
did the corresponding normal cells from each
species. The growth patterns ofthe WT
virus-transformed cells were very similar at both
permissive and nonpermissive temperatures.
The ts A mutant-transformed cells paralleled
the WT virus-transformed cellsintheir growth
characteristics under permissive conditions. In
contrast, when placed at the nonpermissive
temperature, the mutant-transformed cells
ex-hibited the slower growth rates and lowerfinal
saturationdensities of normal cells. It should be
noted that, although 1 x 105 to 2 x 105 cells
were distributed per culture, only
approxi-mately 10 to 40% of the cells actually plated
(Table 2). Therefore, the saturation density
values of 2 x 105 to 4 x 105 cells per culture
attainedby the normal cells and by the
mutant-transformed cellsatthehightemperature
repre-sents cell growth and not mere attachment of
the cellstoculture dishes.
Several saturation density experiments are
summarized inTable3. Itcan beseen thatthe
maximumcelldensitiesofall the
mutant-trans-formedcell lines arereducedatthe
nonpermis-sive temperature. In general, the saturation
densities of the mutant lines at the elevated
temperature are similarto thevalues attained
by
the normal cells from which they werederived. The ratio of maximum cell density at
the permissive temperature tothatat the
non-permissive temperature
(P/NP)
averagedbe-tween4and7withmost ofthemutantcell lines.
The tumor cell lines,
HaTu/A7-1
andHaTu/
A7-2, were exceptional in that they exhibited
P/NP
ratios of >12.0. Ratiosofthe saturationdensities at thetwo temperatureswith the WT
virus-transformedcells
(P/NP
of 1.0 to 1.4)andthe normal control cells (P/NP of 1.1 to 2.5)
wereall closeto 1.
Theseresults suggest that theinactivation of
thegene Aprotein atthenonpermissive
temper-ature causesthets A mutant-transformedcells
toregainthedensity-dependent growth controls
characteristicofnormalcells.Itshouldbenoted
that the
Balb/WT-VLM
cells are not an idealcontrol in these studies, because they were
derived from
BALB/c
embryo fibroblastsratherthan theirderivative line, BALB-3T3.
Unfortu-nately, the BALB-3T3 cells transformed by WT
virus inparallel with theBalb/A7cellsbecame
622 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
SIMIANVIRUS40GENE A FUNCTION. II.
heavily
contaminated with mycoplasma andwere not
able
to be used in all experiments.However, it is pertinent that Renger and
Basi-lico (24) compared the morphology, the
col-1 ony-forming ability, and thesaturation density
of three lines of SV40 WT-transformed
JBALB-3T3 cells and found no differences in
<jghese
properties
at33and39C./
Colony
formation under variouscondi-tions.
Transformed cells have the ability to formcolonies
under conditions in which the growth of normal cells is inhibited, such as insoft agar or on monolayers of normal cells.
Normal cells are able to plate on plastic or glass
surfaces and form colonies of cells asingle-layer
thick; in contrast, transformed cell colonies
contain multiple cell layers and stain very
deeply. We examined the ability of mutant- and
FiG. 1. Appearance of SV40-transformed BALB-3T3 cellsat thepermissiveand nonpermissive tempera-tures. (A) WT-transformed cells (Balb/WT) atthepermissive temperature, 33 C. (B) Balb/WT cellsat the
nonpermissive temperature, 39.5 C. (C)MutanttsA7-transformed cells(Balb/A7)at33C. (D) Balb/A7cells
at39.5 C. Note the lossoftransformed morphologyatthenonpermissivetemperature. x132.
623
VOL.15,1975
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.504.84.413.188.624.2]624 BRUGGEA'
WTvirus-transformed cells to form colonies at
the permissive and nonpermissive
tempera-turesunderthese threeconditions.Table 4gives
the results of colony formation on plastic
sur-faces. Group A mutant-transformed cells
de-rived from all three speciesformed4- to51-fold
fewer colonies at the nonpermissive
tempera-ture than at the permissive one. The
mutant-transformed hamster and mouse cells
consist-ently demonstrated greater temperature
sensi-tivity than did the comparable transformed
humancell line. TheP/NPratios for theformer
cells were all >15 (15 to 51), whereas the ratio
for the mutant-transformed human cells
aver-aged close to 4 (P/NP of 4.1 + 1.7) from three
separate experiments. WT virus-transformed
cells formed similar numbersofcoloniesatboth
temperatures
(P/NP
of0.8 to 1.5). In thisstudy,only deeply stained colonies were counted to
measure the number of transformed foci, not
'@t ,f'- , r -'r
v-. . ..
?
7' >
-A 7of
.f2et/. y
awF
D
FIG. 2. Appearance of SV40-transformedHEFcellsatthepermissiveandnonpermissivetemperatures.(A)
WT-transformedcells
(Hal
WT-2)atthepermissivetemperature,37C.(B)HalWT-2cellsatthenonpermissivetemperature,40.5 C. (C)MutanttsA30-transformed cells (Ha/A30)at37C. (D) Ha/A30 cellsat40.5 C.Note
thelossoftransformed morphologyat thenonpermissive temperature. x132.
AND BUTEL J. VIROL.
.x IV . ,f
,-.j V,A
1;
)k, r46 vt.,,
I il
II
.a
*SS
V,
4
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.504.101.428.192.620.2]SIMIANVIRUS 40 GENE A FUNCTION.II.
the total number
ofnormal cell colonies. Nor-mal HEF, BALB-3T3, and HuF cells producedsingle-layered,
verylightly
stained colonies at both temperatures which were not counted inthis test. Colonies_were formed at the
nonper-missive temperature by the
mutant-trans--formHEF cells; these
resembled
the lightlystained_colonies formed by normal cells and
were approximately equal in number to the
transformed
fociinitiated by the
samecells
atthe
permissive temperature.The
observed
temperature-dependent
varia-tion in colony formation could have been the
result of the inability of the
mutant-trans-formed cellsto plateatthe restrictive
tempera-ture. Table 2 indicates the plating efficiencies
onplastic surfaces of the cell lines under study.
The normal and mutant-transformed cells
_imnsp
W-- F4
FIG. 3. Appearance of SV40-transformed human Fanconi cells at the permissive and nonpermissive temperatures.(A) WT-transformedhumanFanconi cells(HuFWT-2)atthepermissivetemperature, 37C. (B)
HuF!WT-2 cellsat the nonpermissive temperature, 40.5 C. (C) Mutant tsA28-transformedhuman Fanconi
cells (HuF/A28) at 37C. (D) HuF/A28 cells at 40.5C. Note the loss of transformed morphology at the
nonpermissivetemperature. x132.
625
VOL.15,1975
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.504.85.414.177.618.2]BRUGGEANDBUTEL
showed a twofold or less decreased plating
efficiency atthehightemperature.Therefore,it
is apparent that this cannot account for the
4-to 33-fold variation in colony-forming
ability
described above (Table 4).
Results from experiments assaying colony
20 18
12
10
8
6
4
2
Lii
--J
un
IU-'
-J
x
--Lc
C,
0
20
18
12
10
8
6
4 2
20
18
12
10
8
6
4
2
formation onmonolayersofnormal cells (Table
5) confirm those described above. Group A
mutant-transformed cells formed 4- to 18-fold
fewer colonies at the nonpermissive
tempera-ture than at the permissive one. It should be
notedthat the
P/NP
ratiosforcolonyformationBalb/A7,330C
BalbIWT-VLM,330C
iBalblWT-VLM,39.
50CBalb/A7,39.
50C
,
Balb/3T3,330C
Balb/3T3,39.50C
0 1 2 3 4 5 6 7 8 9 10
DAYS AFTER SEEDING
FIG. 4. Growth curves of normal and SV40-transformed cells at permissive (33 or 37 C) and nonpermissive temperatures (39.5 or 40.5C) wereperformedasdescribed in Materials and Methods for saturation density studies. (A) Mouse cells: normal (BALB-3T3), WT-transformed (Balb/WT-VLM), andmutant ts A7-trans-formed (BalbiA7). (B) Hamster cells: normal (HEF), WT-transformed (HalWT-2), and ts A
mutant-trans-formed (Ha/A28 and Ha/A58). (C) Human cells: normal
(Fanconi),
WT-transformed (HuF/WT-2), and tsA28-transformed(HuF/A28).
626 J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.504.133.401.155.590.2]SIMIANVIRUS 40 GENE A FUNCTION. II.
oncell monolayerswerelower than those found
[image:9.504.49.243.109.309.2]with colony formation on plastic. The frequency
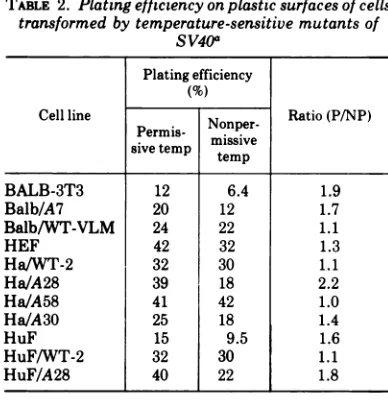
TABLE 2. Plating efficiency on plasticsurfaces of cells
transformed by temperature-sensitive mutants of
SV40a Plating efficiency
(%)
Cellline Nonper- Ratio(P/NP) Permis-misv
sive temp temipve
BALB-3T3 12 6.4 1.9
Balb/A7 20 12 1.7
Balb/WT-VLM 24 22 1.1
HEF 42 32 1.3
Ha/WT-2 32 30 1.1
Ha/A28 39 18 2.2
Ha/A58 41 42 1.0
Ha/A30 25 18 1.4
HuF 15 9.5 1.6
HuF/WT-2 32 30 1.1
HuF/A28 40 22 1.8
aCellsgrowingatthepermissive temperaturewere
trypsinized and seeded at various concentrations
between 102 and 103cells/plate into 35-mm plastic
petridishes containing 2 ml of media with 5% FBS. Onday3thecells were stained and all attachedsingle
cells and smalllightlystained fociwerecounted.
ofcolony formation (the number ofcolonies/103
cells plated) atthepermissivetemperature was
similar under thetwodifferent conditions, soit
appears that the lower P/NP ratios is a
reflec-tion of the greater ability of the transformed
cellstoformcoloniesoncellmonolayers thanon
plastic surfacesatthe hightemperatures.
How-ever, single-layered colonies of normal cells (.
could not be detected in this experiment
be-cause they were masked by the background of
normalFanconi cells. Because of this, itwasnot
possible to determine the plating efficienii-es of
the transformed cell lines on monolayers of
normal cells.
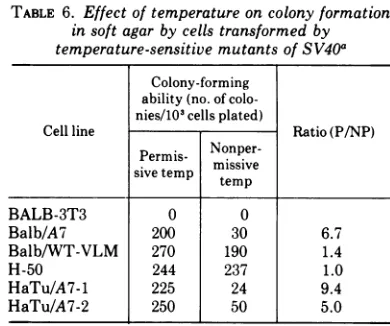
Low P/NP ratios (5.0 to 9.4) were also
ob-tained withtsA mutant-transformed cells from
colony formation experiments performed in soft
agar (Table 6). The WT virus-transformed cells
tested in soft agar (Balb/WT-VLM and H-50)
formed colonies equally well at both
tempera-tures (P/NP of 1.0 to 1.4). It is not known
whether the few colonies initiated by the
mu-tant-transformed cellsatthe restrictive
temper-ature represented revertant cells or whether
they were formed due tofunctional leak of the
mutant geneA protein in those cells.
It is noteworthy that under all three
condi-tions forassaying colony formation, therewas a
TABLE 3. Effect of temperatureon saturationdensity ofcellstransformed by temperature-sensitive
mutantsof SV40a
Avg saturationdensity (no.cells x106/culture)
Cell line No. of expt Ratio(P/NP)
Permissive Nonpermissive
temp temp
BALB-3T3 2 3.5± 1.7 1.4± 0.1 2.5± 0.7
Balb/A7 4 28.0 ±7.0 4.3 1.7 7.1 2.1
Balb/WT-VLM 1 17.7 17.6 1.0
HEF 1 3.7 3.4 1.1
Ha/WT-2 2 16.7± 3.3 13.3± 1.3 1.2± 0.05
Ha/A58 3 34.0±0.9 6.4± 3.2 5.3± 0.7
Ha/A30 1 40.8 8.2 5.0
HuF 2 1.8±0.3 1.7±0.2 1.1 0.05
HuF/WT-2 2 22.0±0.4 18.0±0.3 1.4± 0.4
HuF/A28 1 12.0 3.0 4.0
H-50 1 7.4 7.6 1.0
HaTu/A7-1 3 17.0±4.0 1.5 ± 0.8 12.7±5.4
HaTu/A7-2 2 17.0±7.0 1.5±0.5 12.0± 1.8
aCells growing at the permissive temperaturewere trypsinized, dilutedin media containing5%FBS, and
seeded into35-mm plastic petri dishes at adensityof 1 x 10'to 2 x 102 cells/plate. Replicate cultureswere incubated atthe permissive and nonpermissive temperatures, and cell counts were performed on duplicate
samplesevery otherday. The saturation density ofa cell lineisdefined asTIatnumberofcellspresentin a
culturewhen no rise in cell countsis seen for 3 consecutive days orlonger. The results areexpressedas an average of the cellcounts from two or moreexperiments plusorminus onestandard deviation. Thevalues for
theratio, P/NP, areaverages ofthe ratios from differentexperimentsperformedonthesamecell lineplusor
minusonestandard deviation.
VOL. 15, 1975 627
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 4. Effectof temperatureoncolony formationonplastic surfaces by cells transformed by
temperature-sensitive mutantsofSV40a
Colony-forming ability
(no.colonies/103cellsplated)
Cellline No.ofexperiments Ratio (P/NP)
Permissive Nonpermissive
temp temp
BALB-3T3 2 0 0
Balb/A7 1 196 10 20
Balb/WT-VLM 1 204 136 1.5
HEF 2 0 0
Ha/WT-2 1 264 210 1.3
Ha/A28 2 124 14 5.5 5 23 5
Ha/A58 1 180 8 22
Ha/A30 2 190 34 6 2 33 6
HuF 1 0 0
HuF/WT-2 1 192 244 0.8
HuF/A28 3 212 ± 34 55 ± 14.5 4.1 + 1.7
H-50 1 250 263 1.0
HaTu/A7-1 3 236 ± 43 17 ± 11 15 +8.5
HaTu/A7-2 1 153 3 51
aCellswereplatedasdescribedinthe footnotetoTable2.After 8daysatthenonpermissivetemperature and
[image:10.504.263.458.352.515.2]10days at the permissive temperature, the cells were stained with Giemsa (as described in Materials and Methods), and thenumlberofdeeplystained colonieswascounted. The resultsareexpressedas anaverage of thecolony counts plusorminusonestandard deviation from differentexperiments performed usingthesame cell line. TheP/NPratioswerecalculatedasdescribed in the footnotetoTable3.
TABLE 5. Effect of temperature on colony formation oncell monolayers by cells transformed by
temperature-sensitive mutants of SV40a Colony-forming
ability(no. of colo-nies/103cellsplated)
Cellline Ratio (P/NP)
Permis-
Nonper-Perm
missivesivetemp temp
BALB-3T3 1 1 1.0
Balb/A7 245 31 7.9
HEF 0 0
Ha/WT-2 126 130 1.0
Ha/A28 110 6 18.3
Ha/A58 198 46 4.3
H-50 306 300 1.0
HaTu/A7-1 300 57 5.3
HaTu/A7-2 190 21 9.0
aCells growingatthe permissive temperaturewere
trypsinized and seeded at concentrations ranging between 102 and 109cells/plate ontoconfluent mono-layersofhuman Fanconi cells. After8days of growth at the nonpermissive temperature or 10days atthe permissive temperature, the cells were stained with Giemsa (as describedinMaterials and Methods), and thenumberofdeeply stained colonies was counted.
significant reduction in the ability of the
mu-tant-transformed cells to form colonies at the
restrictive temperature as compared to the
nonrestrictive temperature. No such differences
TABLE 6. Effect of temperature on colony formation insoft agar by cells transformed by temperature-sensitive mutants ofSV40a
Colony-forming ability(no. of colo-nies/103cells plated)
Cell line Ratio(P/NP)
Permis-
Nonper-sivetemp missive
BALB-3T3 0 0
Balb/A7 200 30 6.7
Balb/WT-VLM 270 190 1.4
H-50 244 237 1.0
HaTu/A7-1 225 24 9.4
HaTu/A7-2 250 50 5.0
aCells from the permissive temperature were
tryp-sinized,mixed with a 0.3% agar solution, and placed over a 0.5%agar feeder layer in 35-mm plastic petri dishes. After incubation at the permissive or nonper-missivetemperature for 7 to 10 days, the colonies were counted with a low-power microscope. Only those coloniescontaining greater than10cells were scored.
were found in the values obtained with WT
virus-transformed cells at the two
tempera-tures. These results demonstrate other growth
characteristics of transformed cells which are
temperature sensitive in the group A
mutant-transformed cells.
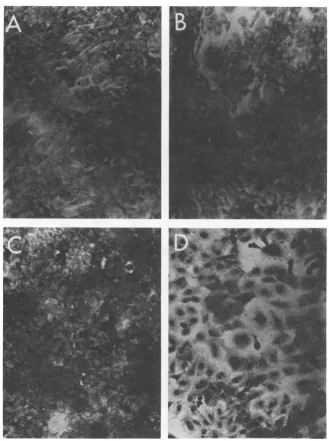
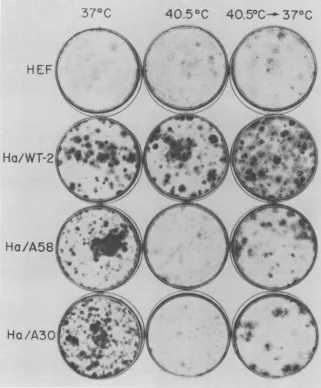
Reversibility of phenotypic changes. To
628 BRUGGE AND BUTEL J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.504.65.258.353.541.2]SIMIAN VIRUS 40 GENE A FUNCTION. II.
demonstrate the
reversibility
of the apparentloss of the transformed phenotype with the
mutant-transformed cells underrestrictive
con-ditions, a shift experimentwas performed with
colonyformation astheassay of transformation.
Normal and transformedhamstercellsfromthe
permissive temperature wereseededintoplastic
petri dishes, and replicate plates were
incu-bated at 37 and 40.5 C. After incubation for 6
days, one-half of the
plates
at 40.5 C wereshifted down to 37
C,
and incubation wascon-tinued. All the plates were stained on
day
11.The results are illustrated in Fig. 5. Plates
370C
HEF
4.C
Ha/WT-2-Ha/A58
.
Ha
/A30
seeded with normal HEF cells contained
nu-merous single-layered, lightly stained colonies
under all three temperature conditions. The
WT-transformedHa/WT-2cells formed
equiva-lentnumbers of colonies, the majority ofwhich
weredensely stainedfoci,under allthree
condi-tions of incubation. The
mutant-transformed
cells, Ha/A58 and
Ha/A30,
alsoformednumer-,,ous transformed foci at 37 C. However,
when"'
those same cells were incubated at 40.5 C
for'k\
theentire 11days, they failedtoform
multilay-ered transformed foci although many lightly"
stained, normal cell colonieswereapparent.It is
40.5
0C
40.50C-4
37°C
FIG. 5. Effect of temperature shift on colony formation by SV40-transformed hamster cells. Cells were plated, and the cultureswereincubatedateither37 or 40.5C. Six days later, part of the cultureswereshifted downfrom40.5 to 37C;all theplateswerestainedonday11.Note theabsenceof transformed focionplates seeded with mutant-transformed cells (Ha/A58andHa/A30) and incubatedcontinuously at 40.5C. Normal, lightly stained colonies were readily visible. Note also that the mutant-transformed cells, when shifted down from40.5 to 37C, initiated foci formation.
VOL.15,1975 629
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.504.90.411.224.612.2]BRUGGE AND BUTEL
significant that plates seeded with Ha/A58 and
Ha/A30 cells and shifted down from 40.5to37 C
contained many deeply stained foci. The
num-ber of colonies found on the shifted plates did
notequal that formed bycontinuous incubation
at 37 C because the plates werestained only 5
days after the shift down to37 C.
These results indicate that the apparent
re-version of ts A mutant-transformed cells to a
normal phenotype isareversibleeventwhenthe
cells are shifted back to permissive conditions.
The results also suggest that the inhibition in
cellgrowth observedwith thesecellsaitthehigh
temperature is not due to cell death caused
either by the elevated temperature or by any
"toxic" effects of the mutated geneA protein.
Uptake of hexose. Cells transformed by
either RNA or DNA tumor viruses possess an
increased ability to transport certain sugars
across the plasma membrane (11). Isselbacher
(14) reported that SV40-transformed
BALB-3T3 cells transported 2.5- to 3.5-fold
greater amounts of 2-D-G than normal
BALB-3T3 cells. Table 7 summarizes assays of
the uptake of [3H]2-D-G by normal, WTvirus-,
and mutant virus-transformed cells at
permis-sive and elevated temperatures. Since the
up-take of 2-D-G by normal cellshas been shown to
be dependentupon celldensity, cultures which
were 75% confluent were used for transport
studies to ensure that the normal and
trans-formed cells were in equivalent growth states.
Both human andmousecellsexhibited a
three-foldorgreaterincrease inuptakeof2-D-G after
transformation by SV40.Thetransformed
ham-ster cell lines were more variable in this
prop-erty. The Ha/A58 cells consistently
demon-strated higher levels of transport than the
normal HEFcells didatthepermissive
temper-ature, but in the one experiment performed
using Ha/A30 cells, less 2-D-G wastransported
by the transformed cells than by normal HEF
cells. However, all the mouse and hamstercell
lines transformed by group A mutants showed
at least a threefold reduction in uptake at the
restrictivetemperaturecompared tothatatthe
nonrestrictive one (P/NP of 3.1 to 10.7). The
levels of uptake for normal cells of all three
species were similar at both temperatures, as
was uptake by WT virus-transformed cells
(P/NP of 0.62to 1.2). Incontrast,theuptake of
2-D-G at theelevated temperature by the HuF
cellstransformed bytsA28wasactually higher
than at the permissive temperature (P/NP of
0.53to 0.83). This parameter isthe onlyonein
which the transformed human cells behaved
differently from the transformed mouse and
TABLE 7. Effectof temperatureonuptake of
[sH]2-D-G by cellstransformed by
temperature-sensitive mutantsofSV40a Uptake of 2-deoxy-glucose (counts per
Cellline Expt m
cells)
Ratiosno. Nonper- (P/NP)
Permis-
missive
sive temp tempBALB-3T3 1 681 730 0.9
Balb/A7 1 4,550 488 9.3
2 10,323 961 10.7
3 10,093 2,148 4.7
Balb/WT-VLM 1 2,900 2,610 1.1
HEF 1 3,067 3,661 0.84
2 1,324 1,058 1.2
Ha/WT-2 1 6,910 11,098 0.62
Ha/A58 1 6,557 1,320 5.0
2 5,409 1,730 3.1
Ha/A30 1 1,207 216 5.6
HuF 1 250 330 0.76
HuF/A28 1 761 1,426 0.53
2 1,935 2,330 0.83
HuF/WT 1 2,908 2,626 1.1
aPetri dishes (35 mm) containing 75% confluent
monolayers of cells werewashed with PBS (pH 7.2) and warmed to 37 C. After the addition of2ml ofPBS containing 1
ACi
of[8H]2-D-G
per ml, the cells were floated in a 37C water bath. After10min, thecells were washedwith coldPBS and scraped into 1 ml of PBS, and 0.1 ml of this suspension was assayed for radioactivity (see Materials andMethods).Triplicatesamples werecounted, and the resultsare expressed
as anaverage ofthe three cultures. Cell counts were determined on replicate cultures which were
incu-batedunderidentical conditionsasthoseassayedfor
uptake of hexose.
hamstercells. Thus, sugaruptakedemonstrates
another property which is
temperature-sensi-tiveinmostgroup Amutant-transformed cells.
Expression of SV40-induced antigens. The
group A protein has not yet been definitively
correlated with any of the SV40-specific
anti-gens found in transformed cells. Since there is
only enough SV40-specific RNA in most
trans-formed cells to code for 1 to 3 proteins, it is
likely that the gene A product is atleast one of
these antigens. Table 8 contains the results
from immunofluorescence tests for two
anti-gens, T andS, in the mutant-transformed cells.
In tests for T antigen, no differenceswere found
in the number ofantigen-positive cells or the
degreeoffluorescence in cells grown at the two
temperatures. In contrast, S antigen was not
detected in cultures of mutant-transformed
cells grown at thenonpermissive temperature.
Since the nature and identity of S antigen
630 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:12.504.262.456.77.323.2]SIMIAN VIRUS40GENEAFUNCTION. II. TABLE 8. Effect of temperature on synthesis of
virus-induced antigensin cellstransformed by
temperature-sensitivemutantsofSV40
Tantigena Santigen' Cellline Per- Nonper- Per-
Nonper-missive missive missive missive temp temp temp temp
Balb/WT-VLM +C + + +
Balb/A7 + + + 0
Ha/WT-2 + + + +
Ha/A58 + + + 0
H-50 + + + +
HaTu/A7-1 + + + 0
HuF/WT + + + +
HuF/A28 + + + 0
aT (tumor) antiserum was prepared by
inoculat-ingweanlingSyrianhamsterswithSV40-transformed
hamstercells.Animals werebledafterthe appearance of tumors. Cells grown at either the permissive or
nonpermissivetemperature were fixed for 3 min with
acetone and stained by the indirect immunofluor-escencetechniqueasdescribed(23).
bS (surface) antiserum was prepared by
immu-nizing hamsters four times with mycoplasma-free
PARA-transformed marmoset cells. Unfixed cells
were stained by an indirect immunofluorescence
technique described previously (25). The reaction of
theSantiserum withSV40-transformedcells was not diminishedby absorption witheithersheepredblood
cells or herpes simplex virus type 11-transformed
hamstercells.
c+, Positive reaction; 0, negative reaction with
immunofluorescence reagents.
remains
obscure,
it isdifficulttointerprettheseresults. The anti-S sera used in this study
appear to reactwithan
SV40-specific
antigen(s)(see footnote to Table 8), but Hayry and
De-fendi (12)
prepared
asimilarreagentwhichwasable to react with normal cell antigens
uncov-ered
by
trypsin treatment. Therefore, untilmore is known concerning the nature of this
antigen, itcan
only
be concluded that thereis astrong correlation between the presence of S
antigen and SV40 transformation. Thus, these
results indicate another
phenotypic
property ofSV40
transformation which istemperature-dependent
in cells transformedby
group A mutants.Rescue oftransformingvirus. It is critical
to establish that the observed
temperature-dependent
phenotypic
expressions oftransfor-mation inthegroup Amutant-transformedcells
is due to a mutantviral function and is not a
reflection of an altered cellularfunction. Using
a special selection procedure, Renger and
Basi-lico (24) isolatedan SV40-transformedcell line
with properties similartoourgroup A
mutant-transformed lines. However, when virus was
rescued from the former cells, itwasfoundtobe
WT in its growth properties. This indicated to
the authors that the fluctuation in the
expres-sion of the transformed state was due to a
mutant cellular function. Since no selective
conditionswereapplied duringthe derivation of
ourcell linesand sincetheywere notcloned, it
seems highly unlikely that only ts cells would
have been transformed and selected in the
evolution of all the transformed cell lines of all
threespecies, and thenonlyfrom those cultures
exposedto ts mutantvirus.
However,
tocharac-terizethetransformingvirus,SV40wasrescued
fromrepresentative tsA-transformed cell lines
ofeachspecies
by
the DNA transfer method(4),
and the rescued virus was assayed at the
per-missive and nonpermissive temperatures to
determine if it retained its temperature
sensi-tivity (Table 9). The virus rescued from group
A mutant-transformed mouse, hamster, and
humancellswas, in fact, temperaturesensitive.
The relative plating efficiencyat40.5compared
to 33 C ranged from 6.2 x 10-1to 1.3 x 10-2.
Although it cannot be proved that the viruses
recovered in this experiment represent the
genomes
responsible
for the transformationevents, these results suggest that the
tempera-ture-dependent
properties of the group Amutant-transformed cellsare due, inall
proba-bility,
to adefective viral geneproduct and notanalteredcellular function.
TABLE 9. Temperature-sensitive nature of virus rescuedfromSV40 group A mutant-transformed
cellsa
Cellline from Titer of rescued virus
which virus (PFU/ml)
Efficiency
of wasrescued 40.5cc 33 platingbBalb/A7 <1.0 x102 3.0 x 104' <3.3 x 10-a
HaTu/A7-1 <1.0x 102 4.7 x 104 <2.1 x 10-a
Ha/A28 1.1 x 101 8.5 x
10'
1.3 x 10-2Ha/A58 2.3 x 101 3.7 x 103 6.2 x 10-a
HuF/A28 <1.0x 102 4.0 x 104 <2.5 x 10-a
aVirus was rescued from the transformed cells by transfection by theprocedure described in Materials and Methods. The rescued virus, which had been
passed once through African green monkey kidney
cells and once through CV-1 cells, was then assayed by the plaque technique (see Materials and Methods) at thepermissive (33 C) and nonpermissive (40.5 C) temperatures. Under these conditions, WTSV40has
an efficiencyofplating (40.5C/33 C) of 1.0. The ts A
mutantstocks have efficiency of plating values which range from 10-3 to 10-6.
bExpressedas(PFU per ml at 40.5C)/(PFUper ml
at33C).
cAssaytemperature.
VOL.15,1975 631
on November 10, 2019 by guest
http://jvi.asm.org/
[image:13.504.256.449.415.539.2]BRUGGE ANDBUTEL
DISCUSSION
This paper has presented evidence which
suggeststhat cells transformed
by
SV40tsmu-tants from complementation group A exhibit
the phenotypic characteristics of transformed
cells whengrown atthepermissivetemperature
and those of normal cells when cultured under
nonpermissiveconditions. Three differenttypes
ofparameters wereexamined tomonitor
trans-formation: (i) those based on the
degree
ofcontact inhibition ofthe cells
(saturation
den-sity and colony formation), (ii) those based on
enzyme functioninthe
plasma
membrane(up-take of
2-D-G),
and (iii) those based on thesynthesis of SV40 antigens. The
expression
ofall of the above criteria of transformation was
foundto betemperature sensitive.
Therefore,
itis concluded that the SV40 gene A
protein
isessential for themaintenance of
growth
charac-teristics
generally
accepted
to betypical
oftransformation.
Since the apparent reversion to a normal
phenotype ofthetsAmutant-transformed cells
at the elevated temperature was monitored
primarily on the basis of parameters which
measure cell growth, it might be
speculated
that the observed phenomena were
merely
areflection of cell stasis or death at the
high
temperature. Several observations suggest this
explanation is incorrect and
indicate,
rather,
that the cellsare
undergoing
phenotypic
rever-sion. (i) WT virus-transformed cells did not
exhibit any temperature
sensitivity
at there-strictive temperatures used in these
experi-ments, indicating that the temperatures
se-lected for study could be tolerated
by
trans-formed cells. (ii) The
growth
characteristics ofthetsmutant-transformed cells atthe elevated
temperature were very similar to those of
nor-mal cells derivedfrom the samespecies
(Fig.
4),
showing that the cellswereabletodivideatthe
high temperature. (iii) All ofthe
mutant-trans-formed cell
lines,
with theexception
of thetumor cell lines, could be passed repeatedlyat
thenonpermissivetemperature. The cells
main-tained the
morphology
and contact-inhibitedgrowth behavior of normal cellsthroughout the
passages (Fig. 1D, 2D,and3D).If the cellswere
unable to grow under restrictive
conditions,
such continual passage would not be
possible.
(iv) The mutant-transformed cellswereable to
produce single-layered colonies at the
nonper-missive temperature which stained lightly and
resemblednormal cell colonies(Fig.5). (v) The
shift-down experiment (Fig. 5) indicated that
cell death
and/or
toxicity of a mutant geneproductdoes not account for theinabilityofthe
mutant-transformed cellsto formtypical
trans-formed foci at thehightemperature. Ifeitherof
those suggestions were correct, the cells would
beunable to initiate newgrowth andwould not
develop foci when shifted down to the
permis-sive temperature. (vi) The cells were able to
actively transport 2-D-G at the nonpermissive
temperature(Table 7),againshowing the viable
stateofthemutant-transformed cells. (vii)The
preceding evidence is strengthened by the cell
counts performed in the saturation density
experiments. The cell counts were performed
with a vital dye, trypan blue, which readily
differentiates the living from the dead cells;
only the viable cells were counted, and these
increased in number throughout the
experi-ments.
The fact that multiple properties of the cells
fluctuated in unison suggests that the gene A
function is involved in the primary events
controlling the expression of the transformed
state.Mutant-transformedcells of mouse,
ham-ster, andhumanorigin behaved similarly,
indi-cating that the gene A protein interacts in the
same manner with constituents of cells which
are both nonpermissive andsemipermissive for
SV40 replication. The only lack of similarity
observed in the phenotypic properties of the
transformed cells from the three different
spe-cies was the failure of the HuF/A28 cells to
exhibit a temperature-sensitive uptake of
2-D-G. The significanceofthisfindingremains to
be elucidated, but, with this one human cell
line, it appears that increased transport of
hexose can be separated from the other
pheno-typic expressions of transformation. Studies are
in progress to define the role of the gene A
function in transformed monkey cells to
deter-mine whether a similar temperature-sensitive
expression of transformation is seen in cells
from apermissive species following
transforma-tion by SV40ts A mutants.
Thegroup A mutants were originally isolated
onthebasisoflossofinfectivity at the elevated
temperature (31). Therefore, the evidence
pre-sented in this paper and the accompanying
reports (21, 22, 30) reveal that in the SV40
system atleastonefunction is requiredbothfor
replication ofintact virus and for maintenance
oftransformation. SincetheSV40 genome
con-tainsonly alimitedamount of genetic
informa-tion, it is not surprising that a virus-specific
function would serve such adual role. However,
thissituation isapparently not always the case
with RNA tumor viruses. In the avian system,
transformation functions can be separated from
replication functions. Temperature-sensitive
mutantshave been isolatedwhich are defective
632 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
SIMIAN VIRUS40GENEAFUNCTION.II.
only for replication (33), defective only for
transformation (3, 20), or defective for both
(35).
DuringSV40lytic infection, the gene A
prod-ucthasbeenfound tobedirectlyresponsiblefor
the initiation of viral DNA synthesis (29).
However, the function is not essential for the
continuation of viral DNA replication once it
has been initiated. It appears, therefore, that
SV40 carriesits owncontrolsystem, orreplicon,
forthe regulation ofviralDNA synthesis. This
information provides a clue concerning the
possible function of the gene A product in
transformed cells. It is possible that normal
cellular DNAsynthesiscomesunder thecontrol
of the viral initiator function in transformed
cells. This could be achieved by the viral
initiator protein interactingwithcellular
opera-torregionstostimulatecellular DNAsynthesis.
Alternatively, themere initiation ofreplication
of the integrated viral genome at the viral
initiator site could trigger a complete round of
cellular DNA synthesis. Either mechanism
could causenormalcellular controlprocesses to
be superseded by those ofthe integrated virus.
In the absence of normal regulatory controls,
cellular DNAsynthesis could be
triggered
repet-itively by a continuously expressed gene A
function. This modelisdescribedin moredetail
elsewhere (J. S. Butel, J. S. Brugge, and C. A.
Noonan, Cold Spring Harbor Symp.
Quant.
Biol., in press). Models with similar features
have also been proposed recently
by
others (R.G. Martin, J. Y. Chou, J. Avila, and R.
Saral,
Cold
Spring
HarborSymp.
Quant.Biol.,
in press; 18, 29).The fact thattsA-transformed cellsrevertto
a normal phenotype at the elevated
tempera-ture is
probably
due to the heat inactivationofthegene Aprotein.Withthe loss offunctional
viral initiator protein, normal
regulatory
mech-anisms resumecontrol of DNA
replication.
Thereturn to normal patterns of host cell DNA
synthesis
should restore some, ifnotall,
ofthephenotypic propertiesofnormal cells. The loss
of the transformed phenotype observed at the
nonpermissive temperature is afully reversible
phenomenon; the transformed characteristics
reappear afterthe cells areshifted back to the
permissive temperature. This suggests that the
transformed phenotype iscontrolledbya
prod-uct of the SV40 genome and that the viral
DNA itself is notexcised orlost from the cells
when reversion occurs.
Thenature ofthe viral initiatorprotein isnot
known. It is possible that this protein could
function as an endonuclease since an
endonu-cleolytic cut in
superhelical
SV40 DNA wouldbe essential for the initiation of viral DNA
synthesis. Virions of a similar mutant of
poly-oma virus, TS-a (also known to be defective in
viral DNA synthesis), were found to carry a
defectiveendonuclease activity (6). In addition,
Ritzi and Levine (26) found thatSV40group A
mutants cause less fragmentation of cellular
DNA than WT virus during lytic infection at
the nonpermissive temperature. This evidence
alsosuggests a defective endonuclease activity.
Although the synthesis of T antigen does not
appear to be temperature-sensitive in group A
mutant-transformedorinfected cells, the
possi-bility that the gene A protein is actually T
antigen cannot be ruled out. A single amino
acid changein theactivesite of aprotein would
not necessarily affect the integrity of its
anti-genic determinants.The sedimentation
proper-tiesofT antigen from tsA-infected cells
incu-bated at permissive or nonpermissive
tempera-tures have been found to be different (M.
Osborn
and K. Weber, personalcommunica-tion). Thissuggeststhatwhile the antigenic site
of the protein is not affected by temperature,
the total configuration of the polypeptide is
altered enough to cause achange in its S value.
It should also be remembered that all
SV40-infected and transformed cells possess T
anti-gen (2,5), suggestingthat itsfunction, likethat
ofthegene A protein, is essentialbothforSV40
replication and formaintenance of
transforma-tion.
It wouldbe informative to know the status of
the tumor-specific transplantation antigen (5)
in the SV40 tsA-transformed cells. If its
pres-ence were also observed to fluctuate when the
cellswereshifted frompermissive to
nonpermis-sive conditions, that would constitute
sugges-tive evidence that tumor-specific
transplanta-tion antigen is also under the control of SV40
geneA.
Asinglesizeclassof
early
SV40 mRNA whichsediments at
19S
has been found inproduc-tively infected cells (34). This observation
sug-gests that the 19S RNA molecule
might
betranslatedintoa
single, large
polypeptide
whichmight,
subsequently,
be cleaved into smallerpolypeptides
in a manneranalogous
to thepattern of
cleavages
which occurs in thepolio-virus system (15, 28). Either the large primary
translation product or the cleavage
products
may possessmultiple functions inthe
cell
(i.e.,stimulation of host cell DNA
synthesis,
initia-tionofviral DNA
synthesis, helper activity
foradenovirus replication, etc.). If a lesion in one
part of the gene affected the processing ofthe
primary polypeptide, the other functionswould
be affected as well. For
example,
the viralVOL. 15,1975 633
on November 10, 2019 by guest
http://jvi.asm.org/
BRUGGE AND BUTEL
initiatorfunction might be located on a
differ-entcleavageproductfrom thefunction
respon-sible for the induction of cell DNA synthesis,
but both activities would appear to be
inacti-vated under nonpermissiveconditions.It isnot
possibleonthe basisofavailable datato
distin-guish in the SV40 system the situation of a
precursor polypeptide which gets cleaved from
the presence of a single multifunctional early
protein.
One previous theory of the mechanism of
virus-mediated transformation
hypothesized
that the mere integration of a foreign viral
genome into a cellular chromosome was
suffi-cientto causetransformation(37). The
identifi-cation of a viral function
(the
gene A protein)which isessentialforthe maintenanceof
SV40-induced transformation proves thatintegration
aloneis notsufficient forstabletransformation,
but, rather, that the continual expression ofat
leastoneviral proteinisrequired. Thisfactalso
suggeststhat theDNAtumorvirusesare
acting
directly on the cell
during
transformation andnot indirectly by the activation of an RNA
tumor virus as proposed
by
the oncogenehy-pothesis (32).This observation doesnotruleout
the
possibility,
ofcourse, that DNA and RNAtumorvirusesmight interactunder appropriate
conditions andmightevenexhibitanenhanced
transforming
ability
under conditions ofjointinfection.
The only previous suggestion ofviral control
ofthe transformed phenotype inaDNA tumor
virus system was with BHK-21 cells
trans-formed by polyoma virus mutant ts-3
(7,
8).With thosecells, only twoparameters of
trans-formation (topoinhibition and wheat germ
ag-glutination) reverted to normal when the cells
were elevated to the nonpermissive
tempera-ture.Twootherpropertiesof the cells(growthin
soft agar and wound serum requirement)
re-mained transformed. Anothergroup ofpolyoma
ts mutants,
typified
by
TS-a,
appeartoresem-bleSV40 groupA mutants
during lytic
infection(9). However, experiments performed to date
with thepolyomavirus ts A mutants have led to
theconclusion that the function isrequired only
for the initiation and not the maintenance of
transformation. It ispossible that the polyoma
virus ts A function differs somewhat from the ts
Afunction ofSV40. Alternatively, experiments
designed to examine additional parameters of
polyoma ts A mutant-transformed cells (other
than the soft agar assay at 38.5 C) might detect
a similar role for that viral protein in the
maintenance ofthetransformed state.
It has been reported previously that SV40
gene A product isessential for the initiation of
transformation (17, 29). In those studies, cells
were infected and plated atthe
nonpermissive
temperature, and the number of colonies was
counted about 30 dayslater. Thepresent
find-ingthat thegeneA function isrequiredfor the
maintenance of transformation makes the
inter-pretation of those results more difficult, if
initiation is considered to be that initial event
which commits a normal cell to ultimately
expressthe characteristics of a transformed cell.
Since the productionof a focus requires
multi-ple cell divisions, it would appear that the
maintenanceoftransformation is required fora
focus to be formed. To assay the initiation of
transformation at the nonpermissive
tempera-ture, it would be necessary to infect cells,
incubate them foralimitedtime atthe
nonper-missive temperature, and then shift them to
permissive conditions to allow foci to form, if
they had been initiated under restrictive
condi-tions. If ts A-infected cells were left under
nonpermissive conditions for the entirety ofan
experiment, foci could not develop even if
initiated, because the transformedstate would
not be maintained. Because of these
reserva-tionsabout the
interpretation
of thosedata, it isnot possible to conclude yet whether another
SV40 functionexists fortheinitiation of
trans-formation, or whether the gene A protein
ac-complishes both steps in the transformation
process.
ACKNOWLEDGMENTS
The authors would like to acknowledge the continual supportand encouragement provided by Joseph L. Melnick. We especially thank Peter Tegtmeyer for providing the ts A mutantsand for helpful discussions.
Thework was supported by Public Health Service research grantsCA 10,893 and CA 15,149 from the National Cancer Institute andby training grant 5 TI A174 from the National Institute of Allergy and Infectious Diseases. J. S. Butel is the recipient of Faculty Research Award PRA-95 from the Ameri-canCancer Society.
LITERATURE CITED
1. Ashkenazi, A., and J. L. Melnick. 1963. Tumorigenicity ofsimianpapovavirusSV40 and of virus-transformed cells. J.Nat.Cancer Inst. 30:1227-1265.
2. Benyesh-Melnick, M., and J. S. Butel. 1974. Oncogenic viruses, p. 403-485. In H. Busch (ed.), The molecular biologyofcancer.Academic PressInc.,NewYork. 3. Biquard, J. M., and P. Vigier. 1970. Isolement et etude
d'un mutant conditionnel du virus de Rous a capacite transformante thermosensible. C. R. Acad. Sci. Ser. D 271:2430-2433.
4. Boyd, V. A., and J. S. Butel. 1972. Demonstration of infectious deoxyribonucleic acid in transformed cells. I. Recovery of simian virus 40 from yielder and non-yielder transformed cells. J. Virol. 10:399-409. 5. Butel, J. S., S. S. Tevethia, and J. L. Melnick. 1972.
Oncogenicity and cell transformation by papovavirus SV40: therole ofthe viral genome. Adv. CancerRes. 15:1-55.
634 J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
SIMIAN VIRUS40GENE A FUNCTION. II. 6. Cuzin, F., D. Blangy, and P. Rouget. 1971. Activite
endonucleasique de preparations purifeesduvirersdu polyome. C. R. Acad. Sci. Ser.D273:2650-2653. 7. Dulbecco, R., and W. Eckhart. 1970.
Temperature-dependent properties of cells transformed bya
ther-mosensitivemutantofpolyomavirus. Proc. Nat. Acad. Sci. U.S.A. 67:1775-1781.
8. Eckhart, W. R., R. Dulbecco, and M. Burger. 1971. Temperature-dependent surface changes in cells in-fectedortransformedbyathermosensitivemutantof polyoma virus. Proc. Nat. Acad. Sci. U.S.A. 68:283-286.
9. Francke, B., andW.Eckhart. 1973.Polyomagene func-tion required for viral DNA synthesis. Virology 55:127-135.
10. Graf, T., andR. R. Friis. 1973. Differentialexpressionof transformation inratand chicken cells infected withan
aviansarcomavirus tsmutant.Virology56:369-374. 11. Hatanaka, M. 1974. Transport of sugars in tumor cell
membranes. Biochim.Biophys.Acta335:77-104. 12. Hayry, P., and V. Defendi. 1970. Surface antigens of
SV40-transformedtumorcells.Virology41:22-29. 13. Hirt, B. 1967.Selectiveextraction ofpolyomaDNA from
infectedmousecell cultures.J. Mol. Biol. 26:365-369. 14. Isselbacher, K. J. 1972.Increaseduptakeof amino acids and 2-deoxy-D-glucose by virus-transformed cells in culture.Proc. Nat. Acad. Sci. U.S.A. 69:585-589. 15. Jacobson, M. F., and D. Baltimore. 1968. Polypeptide
cleavagesin the formation ofpoliovirus proteins.Proc. Nat. Acad. Sci. U.S.A. 61:77-84.
16. Kachani, Z. F., and A. Sabin.1969.Reproductive
capac-ity and viability at higher temperatures of various transformed hamster cell lines. J. Nat. Cancer Inst. 43:469-480.
17. Kimura, G., andR. Dulbecco. 1973. A temperature-sensi-tivemutantof simian virus 40affecting transforming ability. Virology52:529-534.
18. Levine, A. J. 1973.A model for the maintenance of the transformedstateby polyoma andSV40,p.61-75. In
M. Pollard (ed.), Perspectives in virology, vol. VIII. AcademicPressInc.,N.Y.
19. Macpherson, I., and L. Montagnier. 1964. Agar
suspen-sioncultureforthe selectiveassayofcells transformed
by polyoma virus. Virology 23:291-294.
20. Martin, G. S. 1970. Rous sarcoma virus: A function
required for maintenance of the transformed state.
Nature(London) 227:1021-1023.
21. Martin, R., and J. Y. Chou. 1975. Simian virus 40 functionsrequired for the establishment and
mainte-nance of malignant transformation. J. Virol.
15:599-612.
22. Osborn, M., and K. Weber. 1975. SimianvirusgeneA
function and maintenance of transformation. J. Virol. 15:636-644.
23. Rapp, F., S. Pauluzzi, and J. S. Butel.1969.Variationin properties of plaqueprogenyof PARA (defectivesimian papovavirus40)-adenovirus7.J. Virol. 4:626-631. 24. Renger, H.C.,andC.Basilico. 1972. Mutation causing
temperaturesensitive expression ofcell transformation by a tumor virus. Proc. Nat. Acad. Sci. U.S.A. 69:109-114.
25. Richardson, L. S., andJ. S. Butel. 1971. Properties of transformed hamster cells containing SV40 tumor
antigen in the cytoplasm. Int. J. Cancer 7:75-85. 26. Ritzi, E. M., and A. J. Levine.1973.The fragmentationof
cellular DNA and the formation of pseudovirions during SV40 infection of Africangreenmonkeykidney cells. J. Gen. Virol. 20:353-367.
27. Sambrook, J.1972.Transformation by polyomavirusand simian virus40.Adv.Cancer Res. 16:141-180. 28. Summers, D. F., and J. V. Maizel, Jr.1968.Evidencefor
largeprecursorproteinsin poliovirus synthesis.Proc.
Nat. Acad. Sci.U.S.A.59:966-971.
29. Tegtmeyer, P. 1972. Simian virus 40 deoxyribonucleic acid synthesis: the viral replicon. J. Virol.10:591-598. 30. Tegtmeyer, P.1975.Function ofsimian virus 40geneAin
transforming infection. J. Virol. 15:613-618.
31. Tetgmeyer, P., and H. L.Ozer.1971. Temperature-sensi-tivemutantsofsimian virus40:infection of permissive cells. J. Virol. 8:516-524.
32. Todaro, G., and R. J. Huebner.1972.The viraloncogene
hypothesis:newevidence. Proc. Nat. Acad. Sci.U.S.A.
69:1009-1015.
33. Vogt, P. K., J. A. Wyke,R. A. Weiss, R. R. Friis, E. Katz, and M. Linial. 1974. Avian tumorviruses: mutants,
markers, and genotypic mixing, p. 190-205. In 25th
Annual symposium on fundamental cancer research,
1972, Houston, Texas. William & Wilkins Co.,
Balti-more.
34. Weinberg, R. A., S.0.Warnaar,and E. Winocour.1972.
Isolation andcharacterizationof simian virus 40 ribo-nucleic acid. J.Virol. 10:193-201.
35. Wyke, J. A., and M. Linial.1973.Temperature-sensitive avian sarcoma viruses: aphysiological comparisonof twenty mutants.Virology 53:152-161.
36. Zarling, J. M., and S. S. Tevethia.1973.Transplantation immunitytosimian virus40transformed cellsintumor
bearing mice. I. Developmentof cellular immunityto
simian virus 40 tumor-specific transplantation
anti-gens during tumorigenesis by transplanted cells. J. Nat. Cancer Inst.50:137-147.
37. Zinder, N. 1971. Genetic recombination and viral
on-cogenesis, p. 15-22. In M. Pollard (ed.), Perspectives invirology, vol. VII.Academic Press Inc.,N.Y.
VOL.15, 1975

![TABLE 7. Effect of temperature on uptake of[sH]2-D-G by cells transformed by](https://thumb-us.123doks.com/thumbv2/123dok_us/1575604.110152/12.504.262.456.77.323/table-effect-temperature-uptake-sh-d-cells-transformed.webp)

