Copyright© 1975 AmericanSocietyforMicrobiology Printed in U.S.A.
Replication
of
Bacteriophage M13
IX. Requirement
of the Escherichia coli dnaG Function for M13
Duplex
DNA
Replication
DAN S. RAY,* JEANENE DUEBER, AND SIDNEY SUGGS
MolecularBiology Institute and Department ofBiology,* University of California, LosAngeles, California
90024
Received for publication 3 March 1975
Temperature-shift experimentswithanEscherichia coli dnaG strain indicatea
requirement for the dnaG function for M13 phage production only at an early
stageofinfection. Mutant cells infected atnonpermissive temperature formthe parental RF (SS -- RF) but do not replicate further. A shift to nonpermissive temperature after infection inhibits RF RF replication but not RF SS
synthesis. Thesynthesis ofboth strands of the duplexRF wasinhibited equally
after a temperature shift during RF -- RF replication. We infer that thednaG protein is required for M13 production only during RF replication and that it is required for thesynthesis ofboth strands of the RF.
Bacteriophage M13 contains fewer than 10 genes and consequently utilizes many of its host's functions initsreplication(10). With the
availability of many bacterial mutants defec-tive in their own DNA replication, it is now
possibletodetermine which of the known Esch-erichia coli genes involved in bacterial DNA replication also function in M13 DNA
replication. E. coli polA mutants defective for dnaA, dnaB, dnaC(D), dnaE, or dnaG are all blocked in M13 production at nonpermissive temperature (11). Arecentlydescribed mutant,
dnaZ, also fails to support M13 replication at high temperature (18).
Studieswith dnaB and dnaEmutants(12, 17) suggest that neither of these genes are
abso-lutely essential for theinvivo conversion ofthe
M13 viral DNA to its replicative form
(SS
RF). However, both ofthese gene products are
required for later stages of replication-the dnaBproduct
being
required forRF replication (RF RF) but not for single-strand synthesis (RF SS) and the dnaE product required for RF replication and possible for single-strand synthesis aswell. The dnaA gene appears to be required for oneofthe final steps of replication, the closure of linear viral strands (4). At nonper-missive temperature this mutant accumulates linear rather than circular viral strands.The dnaG gene has been implicated recently inthe initiation of Okazaki fragments (8) and in the formation ofanRNA primer forthe comple-mentary strand during parental RF formation invitroby G4 (J. P. Bouche, K. Zechel, and A.
Kornberg, J. Biol. Chem., in press; K. Zechel, J. P.
Bouche,
and A.Kornberg, J. Biol. Chem., in press), a 4X-related bacteriophage. This stageof 4Xreplicationas well as RFreplication andsingle-strand synthesis are all inhibited in vivo in a dnaG mutant at nonpermissive tem-perature (9).Our studies indicate a requirement for the dnaG geneproductinonlyasingle stage ofM13 replication-the replication ofRF. Neither pa-rental RF formation (SS - RF) nor
single-strand
synthesis
(RF - SS)requires
this host function.MATERIALS AND METHODS Phage and bacterial strains. M13wild-type and M13 5amH3 are from the collection ofDavid Pratt and have been described previously (13, 14). E. coli PC3(F-, leu-, thy-,strR,dnaGt)waskindly provided by Randy Schekman. (We have confirmed the
loca-tion of the temperature-sensitive locus by genetic
mapping ofthe mutation.) E. coli PC3X8 is an F+
derivative of PC3. PC3X8 rev. is a
temperature-resistant revertant ofPC3X8which forms colonies at
41 C; however, the reversion of the dnaGt' locus is
only partial sincePC3X8 rev. neither formscolonies
norsynthesizesDNA at 42C.E.coliK37 (supD) was used as indicator for M13 in plating experiments.
Phage were titered by the agar overlay method
described byAdams (1).
Media. Bacteria were grown in glucose-Casamino
Acids medium (16)containingthymidineat 2
,g/ml.
For radioactive labeling of DNA the medium was
supplemented with [3H]thymidineat 10MCi/ml.
Preparationof39P-labeledphage. Forthe
prepa-rationof32P-labeledM13,E. coli K37 wasgrown in 50
348
on November 10, 2019 by guest
http://jvi.asm.org/
resuspended in 0.01 M Tris (pH 8.0) containing 1 mM EDTA and0.5%Sarkosyl and precipitated a second timewithpolyethylene glycol and NaCl. This
precipi-tate wasfinally resuspended in 1 ml of 0.01MTris (pH
8) containing 1 mM EDTA andsedimented for 16 h througha 5 to20%sucrosegradientat24,000 rpmand
5Cin aBeckmanSpinco SW27rotor.Fractions were collected from the centrifuge tube at the end of the
runandassayed for both 32P and PFU.
Harvest and lysis technique. Isotope incorpora-tionwasterminatedbypouring infected cultures into
anequal volume of ice coldTEN-CN buffer (0.01 M Tris, pH 8.0, 1mMEDTA, 50 mM NaCl, and 10 mM NaCN). The cells were harvested and washed by centrifugation for 1 min at 15,000 rpm in aSorvall SS-34rotor at 4C and resuspension in 1 ml ofcold TEN-CN buffer. After washing and resuspending the cellsasecondtime, the cellswerelysedby adding100 Mloflysozyme (4 mg/ml inwater), incubatingfor10
min at 37C, adding 100
Al
of 10% Sarkosyl, and incubatingforanadditional 10min at 37C. Carewastaken not to shear the viscous lysate during the addition andmixingoftheSarkosyl.Thiswas
accom-plished by gently pipetting the detergent into the lysateand slowly rollingthe tubebyhand until well mixed.
Neutral velocity sedimentation oflysates. The viscouslysatecanbegently pouredontothe top ofa5
to 20% sucrose gradient for sedimentation analysis.
Pipetting such lysates should by avoided since this leads to breakage ofthe bacterial DNA and
conse-quently an increased background of bacterial DNA sedimenting in the region of the small phage DNA species. Neutralgradientscontained 0.01 MTris(pH 8), 1mMEDTA, and 1.0 MNaCl. Sedimentationin
an SW27ultracentrifuge rotor wasfor 17hat23,000 rpmand 5C. Fractionswerecollected from the topby pumping 50%sucroseintothe bottom of thegradient and collecting the fractions through a specially de-visedadaptorplacedon topof thecentrifugetube.
Alakline centrifugation techniques. Alkaline equilibrium centrifugation was performed as previ-ouslydescribed (5, 15).
Radioactive counting. Aliquots from neutral
su-crose gradient fractions were spotted directly onto
Whatmanno.3filter papercircles,dried,andcounted by liquid scintillation countinginaBeckman LS230
two-channel counter. Fractions from alkaline CsCl gradientswerecollecteddirectlyontoWhatmanno.3
filter papercircles, dried,andcountedasabove.
proachesthatof theculture heldat 32 Cforthe
entire time (Fig. lb). We infer from this
obser-vationthatthe dnaGfunction isrequiredforSS
-RF
and/or
RF - RFreplication
but not forRF- SSsynthesis, the finalstageofviral DNA
synthesis.
Parental RF formation (SS - RF) in
PC3X8 dnaG cells. The first stage of replica-tion of M13
DNA,
the conversion of thein-fecting viral strandtothe parental RF, doesnot
require the synthesis ofviral proteins. Thisset
of reactions is readily carried out in crude extractsofuninfectedcells andcanbe
reconsti-tuted in vitrousing
purified
bacterial proteins (6). To investigate thepossible role of the dnaG protein in this stage of replication in vivo wehave analyzed RF molecules formed in E. coli PC3X8
dnaG
atdifferenttemperatures. Figure2 shows the neutral velocity sedimentation profiles Jf DNA from lysates of cells infected
at32, 39,and41C. The cellswereinfectedwith 32P-labeled M13 in growth medium containing
[8H ]thymidine,
harvested at5 min after infec-tion,lysed,
and sedimentedthrough
neutralsucrose
gradients.
Since theparental
RFcon-tains 32p label from the infecting viral
strand,
theamountof3 pfoundin
replicative
forms isadirect measure of the amount of
parental
RF formed. Moreparental
RF is formedatboth39and41
C
thanat32C(Fig. 2). Similar
amountsof
parental
RFwereformed(data
notshown)
ina revertant of the dnaG mutant strain and in twodifferent
wild-type
strains,indicating
alack of a requirement for thednaG
function inparental
RFformation.Replication
beyond
this initialstage ofrepli-cationwasinhibitedat39and41
C
asshownby
the incorporation of 3H label into
only
thecomplementary strand of RF II molecules
(Fig.
3). Extensive RF
replication
would havere-sultedinthe
labeling
ofboth strandsoftheRF.Even asingle roundof
duplex
DNAreplication
would have led to an
easily
detectable amount of 3H in the viral strand ofprogeny RFmole-cules, unless the progeny RF molecule
having
on November 10, 2019 by guest
350 DUEBER, ANDSUGGS
E
dnoG40'
KC8
30 60 90 120 0 30 60 90 120 150
time(min)
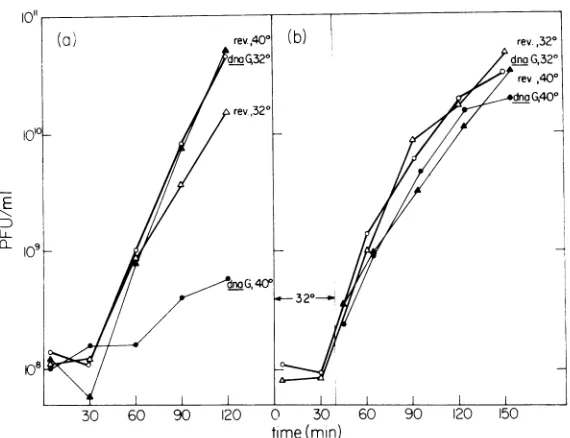
FIG. 1. Requirement for the dnaGfunction early in M13replication.Culturesof E. coli PC3X8 dnaG and E. coliPC3X8 rev. were each grown in glucose-Casamino Acidsmediumat 32Cto acell density of2x 108/mland infected with M13 atamultiplicity of infection of I to 2. (a) Phage productionincultures either held at 32 Cor
shifted to 40 C at 10 minprior to infection; (b) phage production in cultures either held at 32 C or shifted to
40 C at 40 min after infection. Symbols: circles, PC3X8 dnaG; triangles, PC3X8 rev.; open symbols, 32 C; closed symbols, shiftedto 40C.
theparental viral strand isselectively retained
inthe RF II form while the otherdaughter RF is closedtoform anRFI. Should such a
preferen-ial closure occur, it would be possible for a
single
round of RF - RFsynthesis,
butnomore, to go undetected by this method of analysis. Whether or not preferential closure of RF II molecules might occur, extensive RF replication does not take place in the dnaG mutant at nonpermissive temperature.Theobserved blockto RF - RF
synthesis
but not to SS - RFsynthesis indicated that the dnaG function is required only after the initial formation of the
parental RF.
Since parentalRF formationmight
conceiva-blyinvolveresidual activity ofthe dnaG protein it was ofinterest to investigate this reaction in cells preincubated fora longertime at nonper-missive temperature. Figure 4 shows the sedi-mentation profile of DNA from a lysate of PC3X8 cells infected with unlabeled M13 at 40 minafter transfer from 32 to41 C. The amount of3H-labeledRFobserved here is similar to that found in cells infected at only 8 min after transfer(Fig. 2c). These results strongly support the lack ofa requirement forthe dnaG protein in the SS - RF conversion in vivo.
Requirementof the dnaGproteinfor RF RF replication.Toexplorefurther the
involve-ment ofthe dnaG
protein
in RF - RFreplica-tionwe have investigated the temperature
sen-sitivity
of RF - RFsynthesis
in mutant and revertant cells infected with M13 mutants de-fective in gene5.RF RFsynthesis takes placein the absence of the gene 5 DNA-binding
protein
but not that of RF - SS (13).Figure
5 shows the neutral velocity sedimentationpro-files oflysates of PC3X8 and a
partially
tem-perature-resistant revertant ofthe same strain. Both strains were infected with M13
5amH3,
and portions were transferred to separate
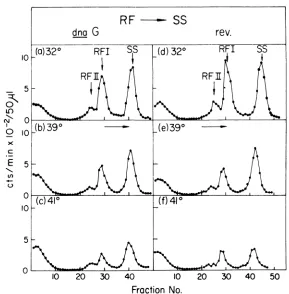
cul-ture tubes at 32, 39, and 41 C at 15 min after infection. Each culture was pulse labeled with [3H]thymidinefrom 25to 30minafterinfection, harvested, lysed, and sedimented through neu-tral sucrose gradients. Labeling of replicative forms wasinhibitedat39and41 Cin thednaG mutantstrainbut less sointhe revertant strain. These results directly implicate the dnaG pro-tein in M13 RF , RF replication.
To investigate the possible selective inhibi-tion ofthesynthesisofthe M13 complementary strand we have analyzed the distribution of label between the two strands of RF II mole-cules formed in the dnaG mutant at 32 and 39 C. Separationofthe twostrands is achieved
by equilibrium centrifugation in alkaline CsCl. Figure 6 shows the 3H label contained in the
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.503.117.402.75.294.2]0
E
-r)
0 20 30
Fraction No.
0
c
0
a-rn~ E
C-)
0
10
20
30
40
50
[image:4.503.261.445.53.313.2]3 Fraction No
FIG. 3. Distribution of 3H label between the viral
~
2 andcomplementarystrandsofRFlIIformedinPC3X8
dnaGat39and41 C. Pooled fractionsfrom theRFII
regions of Fig. 2b andc weredialyzed against0.01M t4
~
Tris(pH8) containing 1 mMEDTA and thensedi-mented to equilibrium in alkaline CsCI gradients.
Densityincreases from righttoleft in these gradients.
0 Thepositions in thedensitygradientof viral strands
40 50 (VS) and complementary strands (CS)areindicated
by verticalarrows.Symbols: 32pJ;0,3H.
FIG. 2. Parental RFformation(SS RF)inPC3X8
dnaG. Three 20-ml cultures of PC3X8 dnaG were
growninglucose-CasaminoAcidsmedium at32Cto
a celldensity of approximately2x 108cells/ml. Two
of the cultures were transferred to39and41 C, and after 7 min [3H]thymidine was added to all three
cultures. At 8 min after the temperature shift each
culturewasinfected with 32P-labeledM13at1010/ml,
and after an additional 5 min all cultures were harvested, lysed, and sedimented through neutral
sucrosegradientsasdescribed. The direction of
sedi-mentation is indicated by the horizontalarrow. The
sedimentationpositions of RF I,RFII,andphageare
indicatedbyverticalarrows.Symbols: ,32p;0,3H.
separated strands of the RF II's. The extentof labeling of the complementary strand relative
tothatoftheviralstrand is thesameat39 Cas
at 32 C. Adifference in the relative labelingof thetwostrandswould have beenexpectedifthe viralstrand hadbeen synthesizedpreferentially
at the nonpermissive temperature. This evi-dence suggests that the dnaG protein is not
required exclusively for the synthesis of the complementary strand of the RF duringRF ,
0
LO
10-
5-
0-10 20 30 40 50
Fraction No.
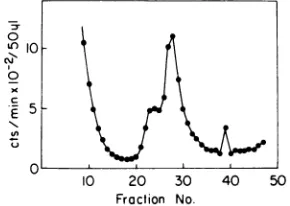
FIG. 4. Parental RF formation in PC3X8 dnaG
after extendedincubationat41 C.A40-ml culture of
PC3X8 dnaGwasgrownat2 x 108cells/mlasinthe
legendtoFig.2.The culturewasshiftedto41 Cfor40
min before infection byunlabeledwild-typeM13. One
minutepriortoinfection [3H]thymidinewasaddedto
the cultureto 10 MCi/ml. At 5minafter infection the
culturewasharvested, lysed,and sedimentedthrough
aneutralsucrosegradientasdescribed.The direction
ofsedimentation isfrom lefttoright.
RF replication.
Single-strand synthesis
(RF
SS) inPC3X8 dnaG. The final stage of M13 DNA
2
on November 10, 2019 by guest
[image:4.503.53.238.62.416.2] [image:4.503.277.422.421.524.2]352 RAY, DUEBER, ANDSUGGS
0
F
>(b)39
--b
(e)
390
E
'N'
(c)410
(f)410
5
l0 20 30 40
l0
20 30 40 [image:5.503.108.410.70.439.2]Fraction
No.FIG. 5. RFreplication(RF , RF) inPC3X8 dnaG andPC3X8rev. Three 30-mlculturesof PC3X8dnaG
andthree 30-ml cultures ofPC3X8rev. wereeach grownat 32 C ingiucose-CasaminoAcids mediumtocell densitiesof approximately2 x 108/mland theninfectedwith M13 5amH3 at1010/ml.At 15min after infection
twoculturesofeachsetweretransferredto39and41 C,andafter10min each culturewaspulselabeled with [8H]thymidinefor5min. All cultures wereharvested, lysed,and sedimented through neutralsucrose gradients
asdescribed. The direction of sedimentation is indicated by the horizontal arrows.
replication, the asymmetric
synthesis
of viralstrands, requires two viral gene functions in addition to host functions
(13).
Toinvestigate
the potential role of dnaG in this stage of
replicationwehavepulselabeled both thednaG
mutantand itsrevertantaftershiftingto39and 41 C at 45 min after infection. The cells were
pulse labeled with [3H]thymidine for 5 min at
10 min after the temperature shift. Figure 7 shows the velocity sedimentation pattern of
lysates fromeach of the infected cultures.
Sub-stantial amounts of viral single strands were
synthesized even at 41 C in the dnaG mutant (Fig. 7c). Comparison with the amount of
syn-thesisobservedintherevertantstrainshows no significant differenceinsingle-strand synthesis. We infer from thisthatasymmetricviralstrand
synthesis does not require a functional dnaG
protein for continued synthesis.
DISCUSSION
The observations presented here indicate that the E. colidnaGfunction is required for M13 RF replication but not for parental RF formation
(SS , RF) or
single-strand synthesis
(RFSS). A point of concern with regard to the significance of these results for the SS - RF reaction is the need for only a single round of
on November 10, 2019 by guest
http://jvi.asm.org/
c
10
20
3
0E",
N~~~~~~~~
C)
10 20 30 40
Froction
No.
FIG. 6. Distribution of 3H label between the viral and complementary strands ofM13 5amH3 RF II formed at 32 and 39 C in PC3X8 dnaG. Pooled fractions from the RF II regions of Fig.5aand bwere dialyzed and sedimented to equilibrium in alkaline CsCI as in the legend to Fig. 3. 32P-labeled viral DNA was added as a density marker. Density
in-creases from right to left. The positions of viral
strands (VS) and complementary strands (CS) are
indicatedby verticalarrows.Symbols: 0,3p;0,3H.
replication for parental RF formation tooccur.
This small amount of synthesis might readily takeplace duetoeitherresidual enzyme
activ-ityatthenonpermissivetemperatureor reratu-ration of the enzyme during some stage of processing of the infected cells prior to lysis. Thestrongest argumentagainst suchanartifact
isthe clean inhibition ofRF RFreplication in cells infected at nonpermissive temperature. Had there been a similar requirement for the dnaG function for the SS RF reaction then thereshould have beenasignificant decreasein
the amount of parental RF formed at high temperature. Yet, just the opposite was
ob-served. There was an actual stimulation of
parental RF formation at temperatures of 39 and 41 C. This lack of a requirement for the
dnaG protein for the conversion ofM13 SS to
RF agrees with the observations made on in
vitro systems (6, 21). This reaction has now
been reconstituted from purified proteins and requires only RNA polymerase, aDNA
polym-erase III holoenzyme, and DNA unwinding
protein (7).
strandsofthe RFindicatedarequirement of the
dnaG function for thesynthesisofboth strands. Residual labeling ofRF molecules in the dnaG
mutant at high temperature labeled both strands equally. In parallel experiments with a
dnaB mutant (unpublisheddata) we alsofound equal inhibition ofsynthesis of bothstrands of
M13 RF at nonpermissive temperature. We therefore infer that neither the dnaB nor the dnaG proteins are required solely for
comple-mentarystrandsynthesis during RF replication.
ThednaGproteinhasbeen shownrecently to
be a rifampicin-resistant RNA polymerase
ca-pable offorming anoligoribonucleotide primer on single-stranded G4 DNAcoated withthe E.
coli unwinding protein (Bouche et al., J. Biol.
Chem., inpress).Atranscriptof aunique region
of the G4 DNA can serve as a primer for the
synthesis
ofthe G4 complementary strand dur-ingSS
_RFsynthesis. This function is similartothatofRNApolymeraseininitiatingthe M13 complementary strand (4, 6, 20). Ittherefore is
likely that the dnaG protein might prime the synthesis of oneof the strands of the M13 RF. However, should the
dnaG,
protein be involved in the initiation of either strand of the RFduring
RF - RFreplication,
the mechanism of initiation would needtobeunique tothatstage of replication since neither complementary strand initiationduring SS - RFsynthesis
norviralstrandinitiationduring RF - SS synthesis require the dnaG function. It is unlikely that thednaGproteinisdirectly involvedin
comple-mentary strand initiation since rifampin
selec-tively
inhibitsM13complementary
strandsyn-thesis during RF - RF
synthesis
(4). Thisobservation suggests
that,
asfor M13 SS- RFsynthesis, the
complementary
strandprimer
is formed by RNA polymerase.We are thus left with the
possibility
thatthednaG protein initiates viral strand
synthesis.
Inhibition ofviral strand synthesis
might
also prevent complementary strand initiation since RF - RF replication appears to occurby
arolling-circle mechanism (7) inwhich the viral
on November 10, 2019 by guest
[image:6.503.52.242.59.305.2]354 RAY, DUEBER, AND SUGGS
C'j0
o
lO0(b)
39
00(e)39
E5
N-0
C(c)
410
(f)
41
010
5-10
20
30
40
l0
20
30
40
50 [image:7.503.114.403.71.365.2]Fraction No.
FIG. 7. Single-strandsynthesis (RF SS)in PC3X8 dnaG and PC3X8rev. Three30-ml culturesof PC3X8 dnaGandthree 30-ml culturesofPC3X8rev.wereeach grownat32C inglucose-CasaminoAcids mediumto a
celldensity ofapproximately2x 108/ml.At45minafter infectiontwoculturesofeachset weretransferredto39
and41 C, andafter 10mineach culturewaspulse labeled with [3H]thymidinefor5min. All cultures were
harvested, lysed, andsedimentedthroughneutralsucrosegradientsasdescribed. The directionof sedimenta-tion is indicatedbythehorizontalarrows. Thesedimentationpositions ofRFI,RFII, and SSareindicatedby
verticalarrows.
strandis theelongatedstrand(19).Inthiscase,
the templateforcomplementarystrand synthe-sis, the elongatedtail of the viralstrand, would
notbeformed and thesynthesisofboth strands would be inhibited equally. The experiments presented herewouldthereforenotdiscriminate between a direct requirement of the dnaG
protein for the synthesis of both strands of the RForofonlythe viral strand. Should the dnaG
protein prove to be directly involved only in
viral strand synthesis during RF RF
synthe-sis, it will be of interestto comparethe
mecha-nism ofinitiation of viral strands during both
RF RF and RF SSsynthesis. From results
presented here, we infer that the dnaG protein
is notrequired forviralstrandsynthesis during
RF SS synthesis.
ACKNOWLEDGMENTS
This research was supported by Public Health Service
grant AI-10752 from the National Institute ofAllergy and
InfectiousDiseases.
LITERATURE CITED
1. Adams, M. H. 1959. Bacteriophages. Interscience Pub-lishers, NewYork.
2. Bouvier,F., andN.D. Zinder. 1974.Effects ofthe dnaA thermosensitive mutation ofEscherichia coli on bac-teriophage fl growth and DNA synthesis. Virology 60:139-150.
3. Brutlag, D., R. Schekman, and A. Kornberg. 1971. A possible roleforRNA polymerase inthe initiation of M13 DNAsynthesis. Proc. Natl. Acad. Sci. U. S.A. 68:2826-2829.
4. Fidaniain, H. M., and D. S. Ray. 1974. Replication of bacteriophage M13. VIII. Differential effects of rif-ampicinand nalidixic acid onthesynthesisofthetwo strands of M13duplexDNA. J. Mol.Biol.83:63-82. 5. Francke, B. F., and D. S. Ray. 1971. Formation of the
parental replicativeform DNA ofbacteriophage
OX-174and initial events in itsreplication.J Mol. Biol. 61:565-586.
6. Geider, K., and A. Kornberg. 1974. Conversion of M13 viralstrand to the double-stranded replicative forms by purified proteins. J. Biol. Chem. 249:3999-4005. 7. Gilbert, W.,and D.Dressler. 1968. DNAreplication:the
rolling circle model. Cold Spring Harbor Symp. Quant. Biol. 33:824-835.
8. Lark,K.G.1972.Geneticcontrol over the initiation ofthe
on November 10, 2019 by guest
http://jvi.asm.org/
Acad. Sci. U.S.A. 69:2570-2573.
13. Pratt, D., and W. S. Erdahl. 1968. Genetic control of bacteriophage M13 DNA synthesis. J. Mol. Biol. 37:181-200.
14. Pratt, D., H. Tzagoloff, and W. S. Erdahl. 1966. Condi-tional lethal mutants of the small filamentous- coli-phage M13.I.Isolation, complementation, cell killing,
time of cistron action. Virology 30:397-410.
15. Ray, D. S. 1969. Replication ofbacteriophage M13. II.
The roleofreplicative formsinsingle-strand synthe-sis. J. Mol.Biol. 43:631-643.
16. Ray, D. S., and R. W. Schekman. 1969. Replicationof
RNA polymerase from Escherichia coli. Proc. Natl. Acad. Sci. U.S.A. 71:4425-4428.
21. Wickner, R.B.,M. Wright, S. Wickner, and J. Hurwitz.
1972. Conversion of 6X174 and fd single-stranded DNAtoreplicativeforms inextractsofEscherichia coli. Proc. Natl.Acad. Sci. U.S.A. 69:3233-3237.
22. Yamamoto, K. R., B. M. Alberts, R. Benzinger, L. Lawhorne, and G. Treiber. 1970.Rapidbacteriophage sedimentation in thepresence ofpolyethylene glycol and its application to large-scale virus purification. Virology 40:734-744.
![FIG. 5.anddensitiesastwo[8H]thymidine described. RF replication (RF, RF) in PC3X8 dnaG and PC3X8 rev](https://thumb-us.123doks.com/thumbv2/123dok_us/1570536.109650/5.503.108.410.70.439/fig-anddensitiesastwo-thymidine-described-rf-replication-rf-dnag.webp)