0022-538X/06/$08.00⫹0 doi:10.1128/JVI.80.9.4363–4371.2006
Copyright © 2006, American Society for Microbiology. All Rights Reserved.
Cellular Topoisomerase I Modulates Origin Binding
by Bovine Papillomavirus Type 1 E1
Yan Hu, Randolph V. Clower, and Thomas Melendy*
Department of Microbiology and Immunology and Department of Biochemistry, University at Buffalo, and the Witebsky Center for Microbial Pathogenesis and Immunology, 210 Biomedical Research Building,
School of Medicine and Biomedical Sciences, Buffalo, New York 14214
Received 22 December 2005/Accepted 6 February 2006
In addition to viral proteins E1 and E2, bovine papillomavirus type 1 (BPV1) depends heavily on host replication machinery for genome duplication. It was previously shown that E1 binds to and recruits cellular replication proteins to the BPV1 origin of replication, including DNA polymerase ␣-primase, replication protein A (RPA), and more recently, human topoisomerase I (Topo I). Here, we show that Topo I specifically stimulates the origin binding of E1 severalfold but has no effect on nonorigin DNA binding. This is highly specific, as binding to nonorigin DNA is not stimulated, and other cellular proteins that bind E1, such as RPA and polymerase ␣-primase, show no such effect. The stimulation of E1’s origin binding by Topo I is not synergistic with the stimulation by E2. Although the enhanced origin binding of E1 by Topo I requires ATP and Mg2ⴙfor optimal efficiency, ATP hydrolysis is not required. Using an enzyme-linked immunosorbent assay, we
showed that the interaction between E1 and Topo I is decreased in the presence of DNA. Our results suggest that Topo I participates in the initiation of papillomavirus DNA replication by enhancing E1 binding to the BPV1 origin.
Papillomaviruses (PV) are small, noneveloped DNA tumor viruses with double-stranded DNA (dsDNA) genomes. In vivo, PV DNA replication requires two virally encoded proteins, E1 and E2 (44). E1 is the central replication factor and works as the initiator protein in PV DNA replication (45, 47). E2 plays a critical role in PV origin recognition (22, 25, 28, 29, 31, 49). In addition to E1 and E2, PV DNA replication is carried out using host cellular replication machinery (5, 19, 38).
The bovine papillomavirus type 1 (BPV1) minimal origin of replication consists of three elements, an A/T-rich sequence, an 18-bp palindromic sequence that serves as the E1 binding site, and a 12-bp sequence that serves as the E2 binding site (15–17, 43, 45). PV DNA replication is initiated when the origin binding protein, E1, recognizes and binds to the origin DNA. Early in the PV life cycle, host DNA is present in vast excess of PV DNA. Although one might predict otherwise, it has been shown that E1 has fairly low sequence-specific DNA binding (16, 29, 40, 45, 47, 52). In the presence of the second viral protein, E2, E1 and E2 bind cooperatively to adjacent binding sites in the origin through multiple protein-protein interactions, greatly increasing the sequence specificity of E1 for the origin (1, 4, 11, 22, 25, 28, 29, 31, 37, 49). After coor-dinated binding of E1 and E2 to the origin, E2 is displaced in an ATP-dependent manner and E1 forms a double-hexamer complex, which has helicase activity (21, 27, 30, 52). This com-plex initiates PV DNA replication by unwinding double-stranded DNA bidirectionally from the origin (30).
Human Topo I acts as a DNA swivel and is critical for many
cellular processes, such as DNA replication and transcription (3). Topo I is important for both simian virus 40 (SV40) and PV DNA replication (42, 50, 51). The structural and functional homologue of PV E1, SV40 T antigen, has been shown to bind to Topo I (26, 35). It was previously demonstrated that T antigen recruits Topo I to the SV40 origin, and they function together to initiate SV40 DNA replication (10, 34, 36, 41). Topo I has also been shown to bind to PV E1. The E1-Topo I interaction was mapped to two regions within E1, one encom-passing the dsDNA binding domain (DBD) (binding site A) and the other at the C terminus (amino acids [aa] 589 to 649; binding site B) (6). The C-terminal binding site also contains part of the core helicase domain (48). It has been shown that E1 stimulates the enzymatic activity of Topo I (6).
In this study, we analyzed the DNA binding properties of the BPV1 E1 protein in the presence and absence of the cellular protein factor Topo I. We show that Topo I specifically stim-ulates the origin binding of E1. The interaction between Topo I and E1 is important for this stimulation. We also investigated the relative roles of Topo I and E2 in the stimulation of origin binding by E1. These results suggest that Topo I plays a role in the initiation of BPV1 DNA replication by increasing origin recognition by E1.
MATERIALS AND METHODS
Materials.[␥-32
P]ATP was obtained from Amersham Pharmacia. Calf intes-tinal alkaline phosphatase was obtained from Invitrogen. T4 polynucleotide kinase was obtained from GE Healthcare. Mouse Topo I monoclonal anti-body was obtained from Sigma.
Probes and nonspecific competitor DNA.The BPV1 minimal origin (BPV1 coordinates 7914 to 7927) in pUC19 (29) was digested with XbaI and HindIII (58 bp). The digested products were separated using 2% agarose gel electrophoresis. The origin-containing band was excised and gel purified. The origin fragment was dephosphorylated using calf intestinal alkaline phosphatase and labeled using T4 polynucleotide kinase and [␥-32P]ATP. The radiolabeled origin fragment was
* Corresponding author. Mailing address: University at Buffalo, De-partment of Microbiology and Immunology, The School of Medicine and Biomedical Sciences, 213 Biomedical Research Building, 3435 Main Street, Buffalo, NY 14214. Phone: (716) 829-3789. Fax: (716) 829-2158. E-mail: [email protected].
4363
on November 8, 2019 by guest
http://jvi.asm.org/
moiety was removed using thrombin (5 U/mg protein for 12 h on ice). The E1 protein was further purified by SP-Sepharose and Q-Sepharose ion-exchange chromatography. The GST-E1 DBD and the GST-E1 truncation mutant (aa 589 to 649) were purified in a similar manner. BPV1 E2 was purified through SP-Sepharose and Q-Sepharose ion-exchange chromatography. Human Topo I was purified from extracts of baculovirus-infected Sf9 or Hi-5 cells (39). Full-length human replication protein A (RPA) was expressed inEscherichia coliand purified as described previously (14). All proteins were purified to near homo-geneity. Full-length E1 and E2 were tested using in vitro PV DNA replication assays and determined to be fully functional (24).
EMSAs.Electrophoretic mobility shift assays (EMSAs) were performed by incubating radiolabeled BPV1 origin DNA (approximately 2 fmol DNA) with proteins at the indicated concentrations in buffer containing 20 mM HEPES-K⫹ [pH 7.5], 100 mM NaCl, 10% glycerol, 4 mM ATP, 7 mM MgCl2, 0.1 mg/ml bovine serum albumin, 1 mM dithiothreitol, 1 mM EDTA in the presence of 20 ng of nonspecific competitor DNA for 30 min at 37°C. Glutaraldehyde was added to a final concentration of 0.02% (vol/vol) and incubated for an additional 20 min at 25°C. Without cross-linking, E1 bound to DNA and resulted in a smeared band. Products were resolved by electrophoresis on a 5% native polyacrylamide gel (79:1 acrylamide/bis-acrylamide) or a 1% agarose gel in 0.5⫻ Tris-borate-EDTA buffer (45 mM Tris-borate, 1 mM Tris-borate-EDTA) at 4°C. The gels were dried, and the products were analyzed using a Bio-Rad FX Molecular Imager system with Quantity One software. For antibody supershift assays, polyclonal E1 anti-body (1:20) and monoclonal Topo I antianti-body (1:20) were added to the reaction mixture before glutaraldehyde for an additional 10 min at 25°C. EMSAs using nonorigin fragments were performed in a manner similar to that for origin DNA, except the nonspecific competitor DNA was decreased to 0.4 ng in some reac-tions (see the figure legends).
Protein-protein ELISAs in the presence and absence of DNA.Enzyme-linked immunosorbent assays (ELISAs) were performed in 96-well polyvinyl plates as described previously, with modifications (6, 20). Briefly, the wells were coated for 60 min at 25°C with purified Topo I (2.0 or 2.5 pmol) in 50l of Tris-buffered saline (TBS) (20 mM Tris-HCl [pH 7.4], 150 mM NaCl) as the immobilized protein. The wells were washed (five times) with TBS-T (TBS with 0.1% NP-40 [vol/vol]), blocked using 1% dry milk (wt/vol) and 5% calf serum (vol/vol) in TBS for 60 min at 25°C, and washed again. Increasing concentrations of the second protein were added to the wells and incubated for 60 min at 25°C in TBS-T with 20 U/ml of micrococcal nuclease, 1 mM MgCl2, and 1 mM CaCl2. The wells were washed, a polyclonal anti-GST antibody (1:5,000 in TBS-T with 0.5% dry milk [wt/vol] and 2% calf serum [vol/vol]) was added for 60 min at 25°C, and the wells were washed again. An anti-goat horseradish peroxidase antibody (1:5,000 in TBS-T with 0.5% dry milk [wt/vol] and 2% calf serum [vol/vol]) was applied to the wells for 45 min at 25°C. The wells were washed and incubated with 50l of visualization solution (110 mM sodium acetate [pH 5.5]) containing the chro-mogenic substrate 3,3⬘,5,5⬘-tetramethylbenzidine (0.02 mg/ml) and hydrogen peroxide (0.0075% [vol/vol]). After 5 min, the reaction was stopped with the addition of 50l of 2 M sulfuric acid. The assays were quantified spectropho-tometrically by absorbance at 450 nm. Each assay was performed at least three times. All incubations were performed with rocking at 25°C.
For ELISAs performed in the presence of oligonucleotides, the above proce-dures were modified to include an additional step. The second proteins were preincubated at 25°C for 15 min (with increasing concentrations of a 60-bp double-stranded DNA oligonucleotide) before being added to the blocked wells (see figure legends). Both origin and nonorigin fragments were used as described previously (see below for details). For these assays, micrococcal nuclease was not added.
RESULTS
Topo I specifically stimulates origin binding by E1. We previously reported that papillomavirus E1 protein interacts with the cellular protein Topo I (6). The E1-Topo I interaction was mapped to two regions within E1, one containing the dsDNA binding domain and the other at the C terminus (aa 589 to 649). To further investigate the E1-Topo I interaction, we evaluated whether Topo I could modulate the origin bind-ing activity of E1. A DNA fragment containbind-ing the PV minimal origin of replication was32P end labeled and incubated with
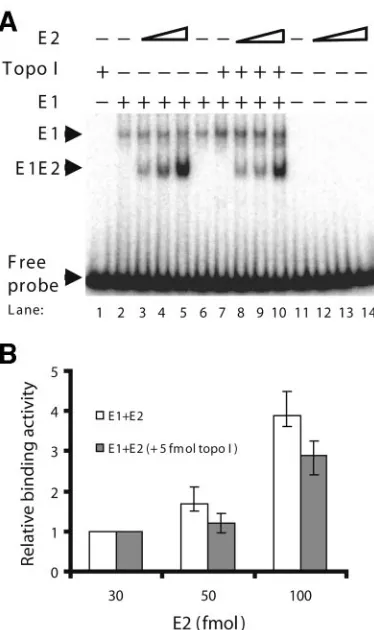
[image:2.585.328.509.68.332.2]purified E1 in the presence and absence of Topo I. The pro-tein-DNA complexes were cross-linked with glutaraldehyde and subjected to gel electrophoresis. The binding of E1 to origin DNA resulted in an E1-DNA complex (Fig. 1A, lane 8). The addition of increasing amounts of Topo I to this complex resulted in increased binding of E1 to origin DNA. At higher levels of Topo I, a Topo I-DNA complex that migrated slightly faster than the E1-DNA complex was seen (Fig. 1A, Topo I, lanes 4 to 7 and 11 to 14). Quantification demonstrated the Topo I stimulation of origin binding activity by E1 to range from three- to sevenfold (Fig. 1B; also see Fig. 3C). To verify
FIG. 1. Topo I stimulates the origin binding activity of E1. (A) A fixed amount of E1 (0.5 pmol) and increasing amounts of Topo I (0 to 200 fmol) were incubated with radiolabeled BPV1 origin DNA for 30 min at 37°C, followed by cross-linking with glutaraldehyde. The result-ing protein-DNA complexes were resolved usresult-ing 5% polyacrylamide gel electrophoresis and analyzed using autoradiography. Migrations of the E1-DNA complex, the Topo I-DNA complex, and the free probe are indicated by arrowheads. (B) Results from panel A were subjected to phosphorimager analysis and quantified using Quantity One soft-ware. The radioactivity of the E1-DNA complex from each lane was quantified and compared to the radioactivity of the E1-DNA complex in the absence of Topo I (lane 8), which was assigned a value of 1. The results represent three separate experiments; the range is depicted by error bars.
on November 8, 2019 by guest
http://jvi.asm.org/
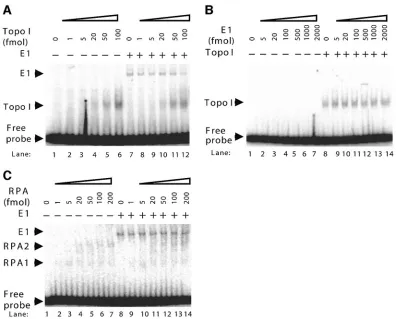
that Topo I stimulation of E1 DNA binding activity is origin specific, we performed EMSAs using a nonorigin DNA frag-ment. The ability of E1 to bind to nonorigin DNA is relative weak compared to its binding to origin DNA (data not shown); therefore, nonspecific competitor DNA was decreased from 20 ng to 0.4 ng in these reactions so that an E1 complex could be detected. E1 bound to the nonorigin DNA probe and resulted in an E1-DNA complex (Fig. 2A, lane 7). With the addition of increasing amounts of Topo I, there was no change in the nonspecific DNA binding activity of E1 (Fig. 2A). These re-sults suggest that Topo I specifically stimulates the origin bind-ing activity of E1.
We also evaluated the ability of E1 to modulate the DNA binding activity of Topo I. Topo I binds DNA with a low degree of specificity (3). The abilities of Topo I to bind to origin and nonorigin DNAs were similar (data not shown). Topo I was incubated with radiolabeled nonorigin DNA in the presence and absence of E1. The results demonstrated that E1 does not affect the DNA binding activity of Topo I (Fig. 2B). PV E1 was shown to interact with several cellular DNA replication proteins, specifically, DNA polymerase␣-primase
(2, 7); the major human single-stranded DNA binding protein, RPA (13); and Topo I (6). To determine whether the stimu-lation of E1 origin binding is specific to Topo I, RPA was also tested for its ability to stimulate the origin binding activity of E1. E1 was incubated with radiolabeled origin DNA in the presence and absence of RPA. The binding of E1 to origin DNA resulted in an E1-DNA complex, and the addition of RPA to the assay did not influence the origin binding activity of E1 (Fig. 2C). As previously reported, RPA also binds to DNA, resulting in a faster-migrating RPA-DNA complex (RPA1) and a slower RPA-DNA complex (RPA2) (20). DNA polymerase␣-primase was also tested, and no stimulation of E1 binding was observed (data not shown). These results fur-ther demonstrate the specificity of the Topo I stimulation of E1 binding, as other cellular proteins that bind to E1 do not stimulate origin recognition.
[image:3.585.95.491.67.386.2]Role of ATP in Topo I stimulation of origin recognition.It was previously reported that ATP and Mg2⫹affect the origin binding activity of E1, specifically, that it is involved in the second part of origin recognition, E1 assembly into a higher-order complex (27, 32). To address this, we first characterized
FIG. 2. Topo I stimulation of E1 binding is highly specific. (A) Topo I does not stimulate the non-origin binding activity of E1. A fixed amount of E1 (1 pmol) and increasing amounts of Topo I (0 to 100 fmol) were incubated with radiolabeled nonorigin fragment for 30 min at 37°C, followed by cross-linking with glutaraldehyde. Migrations of the E1-DNA complex, the Topo I-DNA complex, and the free probe are indicated by arrowheads. The nonspecific competitor DNA was decreased to 0.4 ng in this reaction due to the low nonorigin DNA binding activity of E1. (B) E1 does not stimulate the DNA binding activity of Topo I. A fixed amount of Topo I (0.8 pmol) and increasing amounts of E1 (0 to 2,000 fmol) were incubated with radiolabeled nonorigin fragment for 30 min at 37°C, followed by cross-linking with glutaraldehyde. Migrations of the Topo I-DNA complex and the free probe are indicated by arrowheads. (C) RPA does not stimulate the origin binding activity of E1. A fixed amount of E1 (0.5 pmol) and increasing amounts of RPA (0 to 200 fmol) were incubated with radiolabeled BPV1 origin DNA for 30 min at 37°C, followed by cross-linking with glutaraldehyde. Migrations of the E1-DNA complex, the RPA-DNA complex, and the free probe are indicated by arrowheads.
on November 8, 2019 by guest
http://jvi.asm.org/
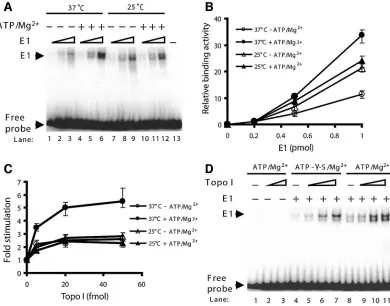
the effects of ATP/Mg2⫹and temperature on the origin bind-ing activity of E1. Increasbind-ing amounts of E1 were incubated with radiolabeled PV origin DNA at 37°C or 25°C in the presence or absence of ATP/Mg2⫹. In these experiments, it could be seen that the binding of E1 to origin DNA resulted in a doublet (Fig. 3A), which has been shown to correspond to hexamers and double hexamers (9, 18). At 37°C, E1 double hexamers were preferentially formed (Fig. 3A and D), similar to what has been reported for SV40 large T antigen (10, 23). The appearance of the E1-DNA complex was dependent upon the E1 concentration. The addition of ATP/Mg2⫹stimulated the binding activity of E1 at 37°C, but not at 25°C (Fig. 3A). Quantification of the E1-DNA complex under all conditions showed optimal E1 binding to the origin to be at 37°C in the presence of ATP/Mg2⫹(Fig. 3B).
We then evaluated whether ATP/Mg2⫹ and temperature affected the stimulation of E1 origin binding by Topo I. Par-allel reactions using a fixed amount of E1 and increasing
amounts of Topo I were performed at 37°C or 25°C in the presence or absence of ATP/Mg2⫹. Under all conditions tested, the addition of Topo I stimulated the origin binding activity of E1. Optimal stimulation of binding of E1 by Topo I to origin DNA occurred at 37°C in the presence of ATP/Mg2⫹. In contrast, the degrees of stimulation were similar at 25°C in the presence and absence of ATP/Mg2⫹and at 37°C in the absence of ATP/Mg2⫹(Fig. 3C). UTP/Mg2⫹was similarly able to support E1 origin binding and optimal Topo I stimulation (data not shown), and based on published E1 origin binding studies (32), we anticipate that the other nucleoside triphos-phates will likewise support E1 origin binding and Topo I stimulation. Mg2⫹alone did not stimulate E1 origin binding (data not shown).
[image:4.585.99.489.70.376.2]Since the PV E1 protein is an ATPase (52), we addressed whether ATP hydrolysis is required for the stimulation of E1 by Topo I. ATP or a nonhydrolyzable ATP analogue, ATP-␥-S, was tested in the reactions. The level of stimulation in the
FIG. 3. The effects of ATP/Mg2⫹ and temperature on the ability of E1 to bind origin DNA in the presence and absence of Topo I.
(A) Increasing amounts of E1 (0.2, 0.5, and 1 pmol) were incubated with radiolabeled BPV1 origin DNA at 37°C or 25°C in the presence and absence of 4 mM ATP/7 mM Mg2⫹. Reaction products were cross-linked and applied to polyacrylamide gels as described in the legend to Fig. 1A.
(B) Results from panel A were subjected to phosphorimager analysis and quantified. The binding activities of 0.2 pmol of E1 to origin DNA were similar in all cases and were assigned a value of 1. The radioactivity of the E1-DNA complex from each lane was quantified and compared to the E1 activity. All results represent three separate experiments; the range is depicted by the error bars. (C) Optimal Topo I stimulation of E1 origin binding occurs with ATP/Mg2⫹at 37°C. Parallel reactions using a fixed amount of E1 (0.5 pmol) and increasing amounts of Topo I (0, 5, 20, and
50 fmol) were performed at 37°C or 25°C in the presence or absence of 4 mM ATP/7 mM Mg2⫹. Reaction products were cross-linked and applied
to polyacrylamide gels as described in the legend to Fig. 1A. In all cases, the radioactivity of the E1-DNA complex from each lane was quantified and compared to the radioactivity of the E1-DNA complex in the absence of Topo I, which was assigned a value of 1. The results represent three separate experiments; the range is depicted by error bars. (D) ATP hydrolysis is not required for stimulation of E1 by Topo I. Parallel reactions using a fixed amount of E1 (0.5 pmol) and increasing amounts of Topo I (0, 1, 5, and 20 fmol) were performed in the presence of ATP (lanes 5 to 8) or a nonhydrolyzable ATP analogue, ATP-␥-S (lanes 9 to 12). Reaction products were cross-linked and applied to polyacrylamide gels as described in the legend to Fig. 1A.
on November 8, 2019 by guest
http://jvi.asm.org/
presence of ATP-␥-S was similar to that observed with ATP (Fig. 3D). Other nucleotides (AMP-PNP and ADP) were also tested, and similar degrees of stimulation were observed (data not shown). These results indicate that optimal stimulation of E1 origin binding by Topo I occurs at 37°C in the presence of ATP/Mg2⫹ but that ATP hydrolysis is not required for the stimulation.
Role of Topo I in the stimulation of origin binding by E1.
We also examined whether the Topo I-dependent stimulation of origin binding by E1 is affected by the sequence in which these two proteins are added to the assay. When Topo I and E1 were mixed together and simultaneously added to the reaction, full Topo I-dependent stimulation of E1 origin binding was observed (Fig. 4, set 2). In contrast, when either Topo I or E1 was preincubated with DNA for 30 min before the addition of the second protein for 3 min, there was no appreciable Topo I stimulation of E1 origin binding (Fig. 4, sets 3 and 4). These results suggest that it is the interaction between E1 and Topo I that is important for the stimulation of E1 binding to the PV origin.
Topo I stimulated E1 to bind origin DNA, and the complex migrated at the same position as the E1-DNA complex, sug-gesting that Topo I may not be present in the complex (Fig. 1A). The composition of the protein-DNA complex stimulated by the addition of Topo I was examined by the addition of antibodies against E1 or Topo I. The addition of polyclonal anti-E1 antibody (␣E1) further retarded the mobility of the E1-DNA complex (Fig. 5, lane 5) but did not have any observ-able effect on the mobility of the Topo I-DNA complex (Fig. 5, lane 8). Conversely, the addition of monoclonal anti-Topo I
antibody further retarded the mobility of the Topo I-DNA complex (Fig. 5, lane 9) but did not have any observable effect on the mobility of the E1-DNA complex (Fig. 5, lane 6). As previously determined, Topo I stimulated the binding of E1 to origin DNA (Fig. 5, lanes 10 to 12). The addition of the anti-E1 antibody further retarded the mobility of this complex (Fig. 5, lane 13) to the same position as the␣E1-E1–DNA complex (Fig. 5, lane 5). In contrast, the addition of the anti-Topo I antibody did not have any observable effect on the mobility of this stimulated complex. Consistent with this conclusion, im-munoblotting of the complex also showed no detectable Topo I (data not shown). This suggests that Topo I is not present in the E1-DNA complex. It also implies that once E1 binds to DNA, Topo I no longer interacts with E1.
DNA modulates the interaction between E1 and Topo I.To further ascertain whether DNA affects the interaction between E1 and Topo I, ELISA-based protein interaction assays were performed in the presence of increasing amounts of DNA. Topo I was immobilized in ELISA plate wells, and after the plates were blocked, GST-E1 was preincubated with DNA for 15 min at 25°C and applied to the wells. Both origin and nonorigin DNA fragments were tested in the assay. In each case, the results demonstrated a decrease in binding between E1 and Topo I (Fig. 6A and B and data not shown).
To determine which Topo I-interacting domain within E1 is modulated by DNA, GST-E1 truncation mutants were ex-pressed inE. coli, purified, and tested for Topo I binding using ELISAs. The GST-E1 DBD, one of two binding sites within E1 that bind Topo I (6), demonstrated decreased interaction with Topo I in the presence of DNA (Fig. 6C and D). Conversely, the GST-E1 truncation mutant, aa 589 to 649, the second domain that also interacts with Topo I, showed no appreciable decrease in binding to Topo I in the presence of DNA (200 fmol up to 350 fmol) (Fig. 6E and F).
Topo I and E2 stimulate the origin binding of E1 indepen-dently.Numerous laboratories have shown that PV E2
[image:5.585.67.263.68.235.2]stimu-FIG. 4. Preincubation of Topo I and E1 stimulates the origin bind-ing activity of E1. (Set 1) E1 (0.5 pmol) and Topo I (0, 5, and 50 fmol) were incubated with origin DNA for 30 min. (Set 2) Initially, E1 (0.5 pmol) was incubated with Topo I (0, 5, and 50 fmol) for 30 min. Origin DNA was then added and incubated for an additional 3 min. (Set 3) Initially, Topo I (0, 5, and 50 fmol) was incubated with origin DNA for 30 min. E1 (0.5 pmol) was then added and incubated for an additional 3 min. (Set 4) Initially, E1 (0.5 pmol) was incubated with origin DNA for 30 min. Topo I (0, 5, and 50 fmol) was then added and incubated for an additional 3 min. Reaction products were cross-linked and applied to polyacrylamide gels as described in Fig. 1A. In each set, the radioactivity of the E1-DNA complex was quantified and compared to the radioactivity of the E1-DNA complex in the absence of Topo I, which was assigned a value of 1. All results represent three separate experiments; the range is depicted by error bars.
FIG. 5. Topo I is not present in the E1-DNA complex. The stim-ulation of E1 origin binding by Topo I was analyzed following addition of anti-E1 (␣E1) and anti-Topo I (␣T) antibodies. Lane 1, free probe; lanes 2 and 3, antibodies added to free probe; lanes 4 to 6, antibodies added to the incubation of E1 (1 pmol) with origin DNA; lanes 7 to 9, antibodies added to the incubation of Topo I (0.5 pmol) with origin DNA; lanes 10 to 14, antibodies added to the incubation of Topo I (50 fmol), E1 (0.5 pmol), and origin DNA. Reaction products were cross-linked, applied to agarose gels, and analyzed using autoradiography. Note that smaller plus signs were used to indicate that less protein was used for formation of the stimulated complex.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:5.585.327.511.70.211.2]FIG. 6. DNA decreases the interaction between E1 and Topo I. (A) ELISAs were performed using Topo I as the immobilized protein. Increasing concentrations of the indicated proteins (GST-E1 or GST-E1 prebound to 200 fmol DNA [60 bp]) were added to the wells. Binding was detected using a polyclonal␣-GST antibody, followed by a secondary horseradish peroxidase antibody. After being developed, the absorbance was measured at 450 nm. The results are representative of three separate experiments; the range is demonstrated by error bars. (B) ELISAs were performed as described for panel A, except that the 60-bp oligonucleotide was added at increasing concentrations. The ordinate represents the percentage of GST-E1 bound to Topo I (where 100% is the amount of GST-E1 bound to Topo I in the absence of DNA), and the abscissa represents increasing molar concentrations of a 60-bp dsDNA fragment. All results represent three separate experiments; the range is depicted by error bars. (C) DNA decreases the interaction between the E1 DBD and Topo I. ELISAs were performed using immobilized Topo I. Second proteins: GST-E1, GST-E1 prebound to 200 fmol DNA (60 bp), GST-E1 DBD, and GST-E1 DBD prebound to 200 fmol DNA (60 bp). All results represent three separate experiments; the range is depicted by error bars. (D) ELISAs were performed as described for panel C, except that the 60-bp oligonucleotide was added at increasing concentrations. The ordinate represents the percentage of GST-E1 DBD bound to Topo I (where 100% is the amount of GST-E1 DBD bound to Topo I in the absence of DNA), and the abscissa represents increasing molar concentrations of a 60-bp dsDNA fragment. All results represent three separate experiments; the range is depicted by error bars. (E) DNA does not affect the
on November 8, 2019 by guest
http://jvi.asm.org/
lates E1 binding to origin DNA through a coordinated mech-anism (21, 25, 28, 29, 31, 32, 37, 49). Here, we show that Topo I stimulates the origin binding activity of E1 in the absence of E2. To investigate how the stimulation effects of E2 and Topo I on E1 are interrelated, we used fixed amounts of E1 and Topo I and added increasing amounts of E2 to an EMSA. As shown previously, the binding of E1 to origin DNA resulted in
the E1-DNA complex (Fig. 7A, lane 2). The addition of E2 resulted in the formation of a faster-migrating E1-E2–DNA complex (Fig. 7A, lanes 3 to 5) (21, 27, 29). Topo I stimulated the binding of E1 to origin DNA, as shown earlier (Fig. 7A, compare the E1-DNA complex in lane 6 with lane 7). In the presence of Topo I, E2 also stimulated the binding of E1 and formed the E1-E2–DNA complex (Fig. 7A, lanes 8 to 10). Interestingly, the quantity of E1-E2–DNA complex was slightly decreased in the presence of Topo I (Fig. 7A and B), possibly resulting from a competition between Topo I and E2 for the fixed amount of E1 in the reaction mixture. The reverse ex-periment with fixed amounts of E1 and E2 and increasing amounts of Topo I produced results consistent with these re-sults (data not shown). These studies appear to indicate that Topo I and E2 stimulate the binding of E1 to origin DNA independently.
DISCUSSION
In this study, we demonstrated that human Topo I specifi-cally stimulates the origin binding activity of PV E1, apparently through a protein-protein interaction. It is still unclear how the interaction between E1 and Topo I stimulates the origin bind-ing activity of E1. We foresee two possibilities. Topo I could bind the PV origin and load E1 through a direct interaction. Conversely, Topo I could interact with E1 prior to DNA bind-ing, evoking a conformational change within E1 and rendering it more efficient for PV origin binding. Topo I is a nonspecific DNA binding protein, and the abilities of Topo I to bind to origin and nonorigin DNAs are similar. It has been demon-strated that Topo I does not stimulate the binding of E1 to nonorigin DNA, so we suspect that the stimulation of E1 origin binding by Topo I may be not caused by direct Topo I loading. Also, we know that when Topo I and E1 were mixed together and simultaneously added to DNA, Topo I stimulation of E1 origin binding was observed, whereas when E1 or Topo I was preincubated alone with DNA before the addition of the other, there was no Topo I stimulation. This suggests that the latter mechanism is more likely—that the interaction between E1 and Topo I imposes a conformational change on E1 that allows E1 to more readily bind to the origin.
It is well established that PV E2 stimulates the binding of E1 to the origin through a coordinate mechanism and forms an E1-E2–DNA complex (21, 27–29, 49). Here, we show that Topo I stimulates the origin binding of E1 and forms an E1-DNA complex. Despite the use of glutaraldehyde, no E1-Topo I-DNA complex was ever observed. Using EMSAs with both E2 and Topo I, we also demonstrated that PV E2 and human Topo I independently stimulate the origin binding activity of E1. This suggests that Topo I and E2 may work through
[image:7.585.68.255.68.383.2]dif-interaction between the E1 C terminus (HPV11 GST-E1 aa 589 to 649; binding Site B [BSB]) and Topo I. ELISAs were performed using immobilized Topo I. Second proteins: GST-E1, GST-E1 prebound to DNA (60 bp), GST-E1 binding site B (aa 589 to 649), and GST-E1 binding site B prebound to DNA (60 bp). BPV1 GST-E1 and HPV11 GST-E1 both bind to Topo I (6), and both demonstrated decreased binding to Topo I in the presence of DNA (panel A and data not shown). All results represent three separate experiments; the range is depicted by error bars. (F) ELISAs were performed as described for panel E, except that the 60-bp oligonucleotide was added at increasing concentrations. The ordinate represents the percentage of GST-E1 BSB bound to Topo I (where 100% is the amount of GST-E1 BSB bound to Topo I in the absence of DNA), and the abscissa represents increasing molar concentrations of a 60-bp dsDNA fragment. All results represent three separate experiments; the range is depicted by error bars.
FIG. 7. Topo I and E2 stimulate the origin binding activity of E1 independently. (A) Effect of E2 on E1-origin complex formation in the presence and absence of Topo I. The additions of E1 (0.5 pmol), E2 (0, 30, 50, and 100 fmol), and Topo I (5 fmol) are indicated above each lane. The proteins were incubated with radiolabeled BPV1 origin DNA for 30 min at 37°C, followed by cross-linking with glutaralde-hyde. Migrations of the E1-DNA complex and E1-E2-DNA complex are indicated by arrowheads. (B) The formation of the E1-E2–DNA complex in the presence and absence of Topo I was quantified using Quantity One software. The binding activities of E1 to origin DNA with 30 fmol of E2 were similar in both cases and were assigned a value of 1. The radioactivity of the E1-E2-DNA complex from each lane was quantified and compared to the E1-E2 DNA binding in lanes 3 and 8. All results represent three separate experiments; the range is depicted by error bars.
on November 8, 2019 by guest
http://jvi.asm.org/
recognition is particularly challenging during early stages of infection in vivo, when the level of E1 is low and the cellular DNA is in vast excess over PV DNA. It has been demonstrated that PV E2 plays a major role in enhancing origin recognition by E1. However, E2 is also present at very low levels during the early stages of infection. In the absence of sufficient levels of E2, having a parallel pathway to stimulate E1 binding depen-dent on cellular proteins, which are present at higher levels, may provide another means for stimulation of origin binding by E1. Our in vitro experiments demonstrate that Topo I, but not other E1-interacting cellular proteins, specifically stimu-lates origin binding by E1. This suggests that in vivo, Topo I may provide such an alternate pathway to assist in origin rec-ognition by E1 when E2 levels are low.
Another possibility for why parallel pathways exist to stim-ulate origin recognition by E1 is that this process is so impor-tant that redundant pathways may be evolutionarily favored. Different PVs may have evolved different pathways for stimu-lation of E1 origin recognition. For example, for some PVs, E2 can stimulate E1 binding through two different mechanisms, while the two mechanisms do not appear to come into play for other PVs (8, 37). It has also been shown that the requirement for E2 in PV DNA replication varies for different PVs. In the most extreme case, HPV1A does not require E2 at all for HPV1A DNA replication (12). In such an instance, the role of Topo I may be much more critical, since E2 does not play a stimulatory role. Hence, the role of Topo I in PV origin rec-ognition may vary from PV to PV. Much work remains to address these questions.
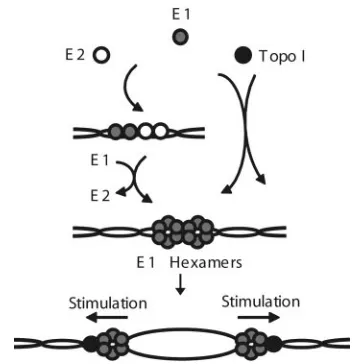
It was previously proposed that E1 recruits Topo I to the PV replication fork through the interaction between the E1 C terminus and Topo I (6). Based on these results, we propose the following model for the role of Topo I in PV DNA repli-cation (Fig. 8). During the initial steps of PV DNA replirepli-cation, E2 and Topo I can independently stimulate origin binding by E1. E2 binds cooperatively with E1 to the origin, resulting in the initial formation of an E1-E2–DNA complex. During sub-sequent steps, more E1 molecules are recruited to the origin of replication through protein-protein and protein-DNA interac-tions in an ATP-dependent fashion to form a double hexamer (9, 18). Concurrently, E2 is displaced from the origin. Our results suggest that there is an alternative pathway, whereby through the interaction with cellular Topo I, E1 takes on a conformation that favors specific binding to origin DNA. Con-comitant with the initial E1 binding to origin DNA, Topo I is released. Topo I stimulation of E1 binding appears to bypass the intermediate complex formed by E1-E2 and takes E1
di-rectly to double-hexamer formation. Upon origin firing, the E1 hexamers act as the replicative helicase (30, 33, 52), and through the interaction between the E1 C terminus and Topo I, E1 recruits Topo I to replication forks. E1 strongly stimu-lates Topo I’s relaxation activity (6). This stimulation is likely important during the elongation stages of PV DNA replica-tion. Together, these results suggest that PV E1 helicase and human Topo I work in concert during multiple phases of PV DNA replication.
Our results suggest that the interaction between E1 and Topo I modulates E1’s ability to bind origin DNA. There are several known cases of cellular chaperone proteins assisting in viral origin recognition by viral recognition proteins. In bacte-riophage P1, theE. colichaperones DnaK and DnaJ facilitate origin-specific DNA binding by RepA and the initiation of DNA replication (46). It has been suggested that interactions between the chaperones and RepA mediate a conformational change in RepA, rendering it more active for origin DNA binding. In human PVs, the human chaperone proteins Hsp70 and Hsp40 independently and additively enhance origin bind-ing by E1 (18). Again, it has been proposed that the interaction between chaperones and E1 alters the conformation of E1 (18). However, the results reported here are the only case we know of where a nonchaperone cellular protein stimulates origin recognition by a viral origin recognition protein. It will be interesting to see whether other viral recognition proteins are similarly stimulated by their interactions with cellular rep-lication factors.
ACKNOWLEDGMENTS
[image:8.585.330.506.67.249.2]This work was supported by NIH grant R29 GM56406 to T.M. We thank Arne Stenlund for the BPV1 E1 and E2 expression vec-tors, Dennis McCance for HPV11 E1 expression vecvec-tors, and J. J. Champoux for the human Topo I baculovirus vector. We thank Van G. Wilson for the polyclonal E1 antibody and E. Niles for critical reading of the manuscript. We also thank J.-S. Liu, S.-R. Kuo, J. C. Fisk, W. FIG. 8. Proposed roles for Topo I in PV DNA replication. During the initiation of DNA replication, Topo I and E2 independently stim-ulate the binding of E1 to origin DNA. After E1 binds to origin DNA and forms a double-hexamer complex at the origin, Topo I is recruited to the origin, presumably through interaction with the E1 C-terminal region of the helicase domain. E1 stimulates the relaxation activity of topoisomerase I (6) during fork elongation.
on November 8, 2019 by guest
http://jvi.asm.org/
Ruyechan, and members of the Witebsky Center for Microbial Patho-genesis at the University at Buffalo for invaluable scientific discussion.
REFERENCES
1.Berg, M., and A. Stenlund.1997. Functional interactions between papillo-mavirus E1 and E2 proteins. J. Virol.71:3853–3863.
2.Bonne-Andrea, C., S. Santucci, P. Clertant, and F. Tillier.1995. Bovine papillomavirus E1 protein binds specifically DNA polymerase alpha but not replication protein A. J. Virol.69:2341–2350.
3.Champoux, J. J.2001. DNA topoisomerases: structure, function, and mech-anism. Annu. Rev. Biochem.70:369–413.
4.Chen, G., and A. Stenlund. 1998. Characterization of the DNA-binding domain of the bovine papillomavirus replication initiator E1. J. Virol.72: 2567–2576.
5.Chow, L. T., and T. R. Broker.1994. Papillomavirus DNA replication. Inter-virology37:150–158.
6.Clower, R. V., J. C. Fisk, and T. Melendy.2006. Papillomavirus E1 protein binds to and stimulates human topoisomerase I. J. Virol.80:1584–1587. 7.Conger, K. L., J. S. Liu, S. R. Kuo, L. T. Chow, and T. S. Wang.1999. Human
papillomavirus DNA replication. Interactions between the viral E1 protein and two subunits of human DNA polymerase alpha/primase. J. Biol. Chem. 274:2696–2705.
8.Dixon, E. P., G. L. Pahel, W. J. Rocque, J. A. Barnes, D. C. Lobe, M. H. Hanlon, K. A. Alexander, S. F. Chao, K. Lindley, and W. C. Phelps.2000. The E1 helicase of human papillomavirus type 11 binds to the origin of replication with low sequence specificity. Virology270:345–357.
9.Fouts, E. T., X. Yu, E. H. Egelman, and M. R. Botchan.1999. Biochemical and electron microscopic image analysis of the hexameric E1 helicase. J. Biol. Chem.274:4447–4458.
10.Gai, D., R. Roy, C. Wu, and D. T. Simmons.2000. Topoisomerase I associ-ates specifically with simian virus 40 large-T-antigen double hexamer-origin complexes. J. Virol.74:5224–5232.
11.Gillitzer, E., G. Chen, and A. Stenlund.2000. Separate domains in E1 and E2 proteins serve architectural and productive roles for cooperative DNA bind-ing. EMBO J.19:3069–3079.
12.Gopalakrishnan, V., and S. A. Khan.1994. E1 protein of human papilloma-virus type 1a is sufficient for initiation of viral DNA replication. Proc. Natl. Acad. Sci. USA91:9597–9601.
13.Han, Y., Y. M. Loo, K. T. Militello, and T. Melendy.1999. Interactions of the papovavirus DNA replication initiator proteins, bovine papillomavirus type 1 E1 and simian virus 40 large T antigen, with human replication protein A. J. Virol.73:4899–4907.
14.Henricksen, L. A., C. B. Umbricht, and M. S. Wold.1994. Recombinant replication protein A: expression, complex formation, and functional char-acterization. (Erratum,269:16519.) J. Biol. Chem.269:11121–11132. 15.Holt, S. E., G. Schuller, and V. G. Wilson.1994. DNA binding specificity of
the bovine papillomavirus E1 protein is determined by sequences contained within an 18-base-pair inverted repeat element at the origin of replication. J. Virol.68:1094–1102.
16.Holt, S. E., and V. G. Wilson.1995. Mutational analysis of the 18-base-pair inverted repeat element at the bovine papillomavirus origin of replication: identification of critical sequences for E1 binding and in vivo replication. J. Virol.69:6525–6532.
17.Li, R., J. Knight, G. Bream, A. Stenlund, and M. Botchan.1989. Specific recognition nucleotides and their DNA context determine the affinity of E2 protein for 17 binding sites in the BPV-1 genome. Genes Dev.3:510–526. 18.Liu, J. S., S. R. Kuo, A. M. Makhov, D. M. Cyr, J. D. Griffith, T. R. Broker,
and L. T. Chow.1998. Human Hsp70 and Hsp40 chaperone proteins facil-itate human papillomavirus-11 E1 protein binding to the origin and stimulate cell-free DNA replication. J. Biol. Chem.273:30704–30712.
19.Liu, J. S., and T. Melendy.2002. Human papillomavirus DNA replication, p. 53–70.InD. J. McCane (ed.), Human papilloma viruses, 1st ed., vol. 8. Elsevier Science B.V., Amsterdam, The Netherlands.
20.Loo, Y. M., and T. Melendy.2004. Recruitment of replication protein A by the papillomavirus E1 protein and modulation by single-stranded DNA. J. Virol.78:1605–1615.
21.Lusky, M., J. Hurwitz, and Y. S. Seo.1994. The bovine papillomavirus E2 protein modulates the assembly of but is not stably maintained in a replica-tion-competent multimeric E1-replication origin complex. Proc. Natl. Acad. Sci. USA91:8895–8899.
22.Lusky, M., J. Hurwitz, and Y. S. Seo.1993. Cooperative assembly of the bovine papilloma virus E1 and E2 proteins on the replication origin requires an intact E2 binding site. J. Biol. Chem.268:15795–15803.
23.Mastrangelo, I. A., P. V. Hough, J. S. Wall, M. Dodson, F. B. Dean, and J. Hurwitz.1989. ATP-dependent assembly of double hexamers of SV40 T antigen at the viral origin of DNA replication. Nature338:658–662. 24.Melendy, T., J. Sedman, and A. Stenlund.1995. Cellular factors required for
papillomavirus DNA replication. J. Virol.69:7857–7867.
25.Mohr, I. J., R. Clark, S. Sun, E. J. Androphy, P. MacPherson, and M. R. Botchan.1990. Targeting the E1 replication protein to the papillomavirus origin of replication by complex formation with the E2 transactivator. Sci-ence250:1694–1699.
26.Roy, R., P. Trowbridge, Z. Yang, J. J. Champoux, and D. T. Simmons.2003. The cap region of topoisomerase I binds to sites near both ends of simian virus 40 T antigen. J. Virol.77:9809–9816.
27.Sanders, C. M., and A. Stenlund.1998. Recruitment and loading of the E1 initiator protein: an ATP-dependent process catalysed by a transcription factor. EMBO J.17:7044–7055.
28.Sarafi, T. R., and A. A. McBride.1995. Domains of the BPV-1 E1 replication protein required for origin-specific DNA binding and interaction with the E2 transactivator. Virology211:385–396.
29.Sedman, J., and A. Stenlund.1995. Co-operative interaction between the initiator E1 and the transcriptional activator E2 is required for replicator specific DNA replication of bovine papillomavirus in vivo and in vitro. EMBO J.14:6218–6228.
30.Sedman, J., and A. Stenlund.1998. The papillomavirus E1 protein forms a DNA-dependent hexameric complex with ATPase and DNA helicase activ-ities. J. Virol.72:6893–6897.
31.Sedman, T., J. Sedman, and A. Stenlund.1997. Binding of the E1 and E2 proteins to the origin of replication of bovine papillomavirus. J. Virol.71: 2887–2896.
32.Seo, Y. S., F. Muller, M. Lusky, E. Gibbs, H. Y. Kim, B. Phillips, and J. Hurwitz.1993. Bovine papilloma virus (BPV)-encoded E2 protein enhances binding of E1 protein to the BPV replication origin. Proc. Natl. Acad. Sci. USA90:2865–2869. (Erratum,90:5878.)
33.Seo, Y. S., F. Muller, M. Lusky, and J. Hurwitz.1993. Bovine papilloma virus (BPV)-encoded E1 protein contains multiple activities required for BPV DNA replication. Proc. Natl. Acad. Sci. USA90:702–706.
34.Simmons, D. T., D. Gai, R. Parsons, A. Debes, and R. Roy.2004. Assembly of the replication initiation complex on SV40 origin DNA. Nucleic Acids Res.32:1103–1112.
35.Simmons, D. T., T. Melendy, D. Usher, and B. Stillman.1996. Simian virus 40 large T antigen binds to topoisomerase I. Virology222:365–374. 36.Simmons, D. T., P. W. Trowbridge, and R. Roy.1998. Topoisomerase I
stimulates SV40 T antigen-mediated DNA replication and inhibits T anti-gen’s ability to unwind DNA at nonorigin sites. Virology242:435–443. 37.Stenlund, A.2003. E1 initiator DNA binding specificity is unmasked by
selective inhibition of non-specific DNA binding. EMBO J.22:954–963. 38.Stenlund, A.1996. Papillomavirus DNA replication, p. 679–697.InM. L.
DePamphilis (ed.), DNA replication in eukaryotic cells. Cold Spring Harbor Laboratory Press, Plainview, N.Y.
39.Stewart, L., G. C. Ireton, L. H. Parker, K. R. Madden, and J. J. Champoux. 1996. Biochemical and biophysical analyses of recombinant forms of human topoisomerase I. J. Biol. Chem.271:7593–7601.
40.Thorner, L. K., D. A. Lim, and M. R. Botchan.1993. DNA-binding domain of bovine papillomavirus type 1 E1 helicase: structural and functional as-pects. J. Virol.67:6000–6014.
41.Trowbridge, P. W., R. Roy, and D. T. Simmons.1999. Human topoisomerase I promotes initiation of simian virus 40 DNA replication in vitro. Mol. Cell. Biol.19:1686–1694.
42.Tsurimoto, T., T. Melendy, and B. Stillman.1990. Sequential initiation of lagging and leading strand synthesis by two different polymerase complexes at the SV40 DNA replication origin. Nature346:534–539.
43.Ustav, E., M. Ustav, P. Szymanski, and A. Stenlund.1993. The bovine papillomavirus origin of replication requires a binding site for the E2 tran-scriptional activator. Proc. Natl. Acad. Sci. USA90:898–902.
44.Ustav, M., and A. Stenlund.1991. Transient replication of BPV-1 requires two viral polypeptides encoded by the E1 and E2 open reading frames. EMBO J.10:449–457.
45.Ustav, M., E. Ustav, P. Szymanski, and A. Stenlund.1991. Identification of the origin of replication of bovine papillomavirus and characterization of the viral origin recognition factor E1. EMBO J.10:4321–4329.
46.Wickner, S., J. Hoskins, and K. McKenney.1991. Function of DnaJ and DnaK as chaperones in origin-specific DNA binding by RepA. Nature350: 165–167.
47.Wilson, V. G., and J. Ludes-Meyers.1991. A bovine papillomavirus E1-related protein binds specifically to bovine papillomavirus DNA. J. Virol. 65:5314–5322.
48.Wilson, V. G., M. West, K. Woytek, and D. Rangasamy.2002. Papillomavirus E1 proteins: form, function, and features. Virus Genes24:275–290. 49.Winokur, P. L., and A. A. McBride.1996. The transactivation and DNA
binding domains of the BPV-1 E2 protein have different roles in cooperative origin binding with the E1 protein. Virology221:44–53.
50.Wobbe, C. R., L. Weissbach, J. A. Borowiec, F. B. Dean, Y. Murakami, P. Bullock, and J. Hurwitz.1987. Replication of simian virus 40 origin-contain-ing DNA in vitro with purified proteins. Proc. Natl. Acad. Sci. USA84:1834– 1838.
51.Wold, M. S., D. H. Weinberg, D. M. Virshup, J. J. Li, and T. J. Kelly.1989. Identification of cellular proteins required for simian virus 40 DNA replica-tion. J. Biol. Chem.264:2801–2809.
52.Yang, L., I. Mohr, E. Fouts, D. A. Lim, M. Nohaile, and M. Botchan.1993. The E1 protein of bovine papilloma virus 1 is an ATP-dependent DNA helicase. Proc. Natl. Acad. Sci. USA90:5086–5090.


![FIG. 6. DNA decreases the interaction between E1 and Topo I. (A) ELISAs were performed using Topo I as the immobilized protein.Increasing concentrations of the indicated proteins (GST-E1 or GST-E1 prebound to 200 fmol DNA [60 bp]) were added to the wells](https://thumb-us.123doks.com/thumbv2/123dok_us/172529.47544/6.585.114.472.73.602/decreases-interaction-performed-immobilized-increasing-concentrations-indicated-prebound.webp)