0022-538X/95/$04.0010
Copyrightq1995, American Society for Microbiology
Sequences Flanking the Pentanucleotide T-Antigen Binding Sites
in the Polyomavirus Core Origin Help Determine
Selectivity of DNA Replication
L. LI,
1B. L. LI,
1M. HOCK,
1E. WANG,
2AND
W. R. FOLK
1*
Department of Biochemistry, University of Missouri-Columbia, Columbia, Missouri 65211,
1and
Department of Biological Sciences, Columbia University, New York, New York 10027
2Received 12 June 1995/Accepted 16 August 1995
Replication of the genomes of the polyomaviruses requires two virus-specified elements, the
cis
-acting origin
of DNA replication, with its auxiliary DNA elements, and the
trans
-acting viral large tumor antigen (T antigen).
Appropriate interactions between them initiate the assembly of a replication complex which, together with
cellular proteins, is responsible for primer synthesis and DNA chain elongation. The organization of
cis
-acting
elements within the origins of the polyomaviruses which replicate in mammalian cells is conserved; however,
these origins are sufficiently distinct that the T antigen of one virus may function inefficiently or not at all to
initiate replication at the origin of another virus. We have studied the basis for such replication selectivity
between the murine polyomavirus T antigen and the primate lymphotropic polyomavirus origin. The murine
polyomavirus T antigen is capable of carrying out the early steps of the assembly of an initiation complex at
the lymphotropic papovavirus origin, including binding to and deformation of origin sequences in vitro.
However, the T antigen inefficiently unwinds the origin, and unwinding is influenced by sequences flanking the
T antigen pentanucleotide binding sites on the late side of the viral core origin. These same sequences
contribute to the replication selectivity observed in vivo and in vitro, suggesting that the inefficient unwinding
is the cause of the replication defect. These observations suggest a mechanism by which origins of DNA
replication can evolve replication selectivity and by which the function of diverse cellular origins might be
temporally activated during the S phase of the eukaryotic cell cycle.
The process by which DNA replication is initiated at the
simian virus 40 (SV40) origin has been elaborated in
consid-erable detail (28, 29, 41, 58, 101, 102). The viral T antigen binds
to the origin in a sequence-specific and ATP-dependent
man-ner to form a double hexamer (3, 5, 16, 17, 19, 22, 23, 68, 80).
It deforms the origin and then, by acting in concert with a
single-stranded-DNA-binding protein (SSB or RPA) and
topo-isomerase I, induces extensive unwinding of the DNA duplex
(4, 8, 18, 34, 59, 79, 88, 99, 121). Concomitantly, DNA
poly-merase
a
/primase interacts with this complex to initiate
RNA-primed DNA synthesis (13, 27, 39, 76). The nascent initiator
DNA is elongated by the combined action of DNA polymerase
a
/primase and a DNA polymerase
d
(or
ε
) complex (61, 77,
102, 111, 112, 115, 118). Leading-strand synthesis is
subse-quently catalyzed by the DNA polymerase
d
(or
ε
) complex,
while lagging-strand synthesis requires repetitive priming by
the DNA polymerase
a
/primase and elongation by the DNA
polymerase
d
(or
ε
) complex (115, 118).
The series of events required for the assembly of the
initia-tion complex is coordinated by highly specific protein-protein
and protein-DNA interactions. Productive interactions
be-tween the T antigen, single-stranded-DNA-binding protein,
and DNA polymerase
a
/primase are required for primosome
assembly and primer synthesis (13, 35, 36, 59, 72, 93). Specific
interactions between T antigen and DNA polymerase
a
/pri-mase (14, 35, 37, 49, 97) have been suggested to be a basis for
species-specific selectivity of viral DNA replication (9, 40, 74,
75). The highly specific protein-DNA interactions required for
the assembly of an initiation complex occur through the
rec-ognition of the origin by T antigen (3–5, 20, 23, 26, 28, 33, 62,
79, 96, 103). The four pentanucleotide motifs (PuPuGGC) in
the central palindrome of the core origin help organize the T
antigen double hexamers (3, 16, 17, 23, 68, 79, 80). The spatial
organization of these four pentanucleotide motifs is critical,
presumably because the T antigen hexamers must be exactly
positioned relative to each other to unwind the origin and to
accommodate SSB and DNA polymerase
a
/primase (2, 9, 12,
17, 23, 81, 114, 119).
Most polyomavirus origins consist of a core with four
pen-tanucleotide motifs, flanked by a DNA-unwinding element and
an A/T-rich sequence (21, 62, 63, 103). Despite these
con-served features, the T antigen of one virus may not efficiently
replicate another’s genome. Species-specific interactions
be-tween T antigens and cellular proteins such as DNA
poly-merase
a
/primase and differences in the spatial organization of
the four pentanucleotide motifs provide several possible
rea-sons why the origins of the murine polyomavirus (mPy) and
SV40 are not replicated by the other’s T antigens. However,
even among viruses which infect the same species and which
have nearly identical central palindromes, as occurs with the
SV40, BK, and JC viruses, there are substantial differences in
the capacity of one’s T antigen to replicate the other’s origin
(1, 65–67, 75, 98). This provides an opportunity to explore the
selectivity of replication complexes and to assess how diversity
among cellular origins might be used to modulate DNA
initi-ation frequency.
To this end, we have studied the interactions of the T
anti-gen of mPy with its own origin and with the origin of the
primate lymphotropic polyomavirus (LPV). Although the mPy
and LPV origins are very similar and have virtually identical
central palindromes, we find that the mPy T antigen does not
help assemble a functional replication complex at the LPV
origin, even with cellular proteins which productively interact
* Corresponding author. Phone: (314) 4845. Fax: (314) 882-5635. Electronic mail address: [email protected].
7570
on November 9, 2019 by guest
http://jvi.asm.org/
with the mPy T antigen. Our data reveal that the capacity of
mPy T antigen to replicate DNA is strongly influenced by
sequences on the late side of the core origin, which differ
between the mPy and the LPV origins. These sequences help
activate the Tag DNA-unwinding activity. These findings
sug-gest a mechanism by which replication of diverse origins can be
modulated and thereby regulate the DNA replication of a
cellular genome with many origins.
MATERIALS AND METHODS
DNAs.The mPy and LPV DNA constructs used in these experiments were cloned in plasmid pUC19 or in phage M13mp19, and the DNAs were purified by two rounds of centrifugation in CsCl-ethidium bromide gradients.
In vivo DNA replication assays.Swiss mouse 3T6 cells and human P3HR1 lymphoblastoid cells were obtained from the American Type Culture Collection. DNAs were introduced into 3T6 cells following precipitation with calcium phos-phate or DEAE-dextran (70, 113) or into P3HR1 cells by electroporation (106
cells per ml, 1,200 V). The transfected cells were then cultured for 48 h, and the DNAs were extracted. The replication products were analyzed after DpnI diges-tion as previously described (110) with mPy and LPV [32P]DNAs as hybridization
probes. In some experiments, test DNAs were introduced into cells together with origin-defective helper DNAs capable of expressing either mPy T antigen (pSO11 [107]) or LPV T antigen (pLPV-ApaR). These helper DNAs have base
substitutions in the mPy core origin (pSO11) or a small deletion at the ApaI site in the LPV core origin (pLPV-ApaR) which prevents replication of the helper
DNA but permits the expression of viral T antigens required for replication of test DNAs.
Preparation of cell extracts.Murine FM3A cells were purchased from Cellex Biosciences Inc. (Coon Rapids, Minn.). Ten liters of cells harvested at 53105
cells per ml were washed twice with ice-cold phosphate-buffered saline and shipped on ice. The subsequent preparation of FM3A cell extracts was done by the procedure described by Wobbe et al. (120). Cells were disrupted in hypotonic buffer (20 mM HEPES [N-2-hydroxyethylpiperazine-N9-2-ethanesulfonic acid, pH 7.5], 5 mM KCl, 1.5 mM MgCl2, 1 mM dithiothreitol [DTT]) by Dounce
homogenization; the lysate was adjusted to 0.2 M NaCl and centrifuged at 50,000 3g for 30 min, and the supernatant was dialyzed for 3 h against 3 liters of dialysis
buffer (20 mM HEPES [pH 7.5], 1 mM DTT, 0.1 mM EDTA, 50 mM NaCl, 10% glycerol). The resulting extract was clarified by centrifugation at 50,0003g for
30 min and stored in aliquots at2808C.
Preparation of T antigens.mPy T antigen was purified from extracts of High Five (Invitrogen) insect cells infected with recombinant baculovirus pVL941-LT (a gift of B. Schaffhausen) by immunoaffinity chromatography (75) with the F4 monoclonal antibody (78). At 48 h postinfection, 2.53108
cells in 500 ml of culture were collected, washed, and frozen at2808C. Thawed cells were lysed by sonication in 25 ml of buffer A (50 mM Tris-HCl [pH 8.0], 150 mM NaCl, 1 mM EDTA, 1 mM DTT, 10% glycerol, 1% Nonidet P-40, 1 mM phenylmethylsulfonyl fluoride) and centrifuged at 30,0003g for 30 min, and the supernatant was
mixed at 48C with 4 ml of protein G-Sepharose coupled with F4 antibody for 3 to 15 h. Bound T antigen was washed extensively and eluted from the packed column with a buffer containing 20 mM Tris-HCl (pH 8.5), 1 mM DTT, 1 mM EDTA, 10% glycerol, 500 mM NaCl, and 50% ethylene glycol. The peak frac-tions of T antigen were dialyzed overnight against 2 liters of dialysis buffer (10 mM HEPES [pH 7.5], 5 mM NaCl, 0.1 mM EDTA, 1 mM DTT, 50% glycerol) and stored in aliquots at2808C. Generally, up to 3 mg of T antigen with a purity of over 90% was obtained from 1 liter of infected cells. The purified T antigen retained maximal activity for at least 6 months. As reported by Lorimer et al. (64), different preparations of T antigen purified by this procedure varied in purity and activity.
In vitro DNA replication assay.mPy DNA replication in vitro was assayed by the method of Murakami et al. (75) and Prives et al. (87). Standard reaction mixtures (50ml) contained 80 mM Tris-HCl (pH 7.8); 40 mM creatine phos-phate; 7 mM MgCl2; 0.5 mM DTT; 4 mM ATP; 200mM each CTP, UTP, and
GTP; 100mM each dATP, dGTP, and dTTP; 20mM [a-32P]dCTP (1 to 10
cpm/fmol); 1mg of creatine phosphate kinase; 0.5mg of plasmid DNA; 0.2 to 0.4 mg of FM3A cell extract; and 1 mg of T antigen. Reaction mixtures were incubated for 2 h at 338C, and the reaction was terminated by the addition of an equal volume of 40 mM EDTA–1.0% SDS and digested with 20mg of proteinase K. DNA was purified by phenol-chloroform extraction and ethanol precipitation. After restriction with DpnI, the replication products were fractionated by elec-trophoresis through a 1.0% agarose gel, dried, visualized by autoradiography, and quantitated with a PhosphorImager. The in vitro replication of mPy ori DNA was totally dependent on T antigen. Maximal in vitro DNA synthesis occurred when 0.5 to 1.0mg of DNA template was incubated with 1.0mg of T antigen per reaction.
KMnO4footprinting.The conditions for KMnO4modification were slightly
modified from those used for SV40 DNA by Borowiec and Hurwitz (4). Binding of T antigen to the mPy origin of replication does not require ATP hydrolysis (64); consequently, an ATP analog, 59-adenylyl imidodiphosphate (AMPPNP), was used, as it gave stronger KMnO4signals than did ATP (data not shown).
The standard reaction mixture (30ml) contained 80 mM Tris-HCl (pH 7.8), 40 mM creatine phosphate, 7 mM MgCl2, 0.5 mM DTT, 4 mM AMPPNP, 1.0mg of
T antigen, and 0.5mg of DNA. Reaction mixtures were incubated at 338C for 30 min. Freshly prepared KMnO4was added to a final concentration of 20 mM, and
the reaction was incubated for an additional 30 s at 338C. The reaction was quenched by addingb-mercaptoethanol to 1.0 M. The mixture was diluted to 80 ml with water and desalted on a 1.0-ml Sephadex G-50 spin column. The eluate was divided into two 35-ml portions, and the modified DNA was denatured by incubation for 5 min at room temperature with 0.1 volume of 2 M NaOH–2 mM EDTA. The solution was neutralized with 0.1 volume of 3 M sodium acetate (pH 5.2), and the DNA was precipitated with ethanol. The resultant single-stranded DNA was dissolved in 10ml of 40 mM Tris-HCl (pH 7.5)–20 mM MgCl2–50 mM
NaCl–1 ng of a 59-end-labeled primer complementary to either the top or the bottom strand sequence about 100 bp away from the central palindrome. The primers were annealed to the DNA by heating for 2 min at 658C, followed by cooling slowly over 15 to 30 min. Primer extension was carried out at 378C for 30 min following the addition of 0.5 mM deoxynucleoside triphosphate (dNTP) mix, 10 mM DTT, and 1 U of Klenow fragment of DNA polymerase I. The reaction was terminated with 6ml of stop solution (96% formamide, 20 mM EDTA, 0.05% bromophenol blue, 0.05% xylene cyanol FF), heated at 908C for 2 to 5 min, and loaded onto a 6% polyacrylamide sequencing gel. Sequencing ladders derived with the same end-labeled primers were run beside the footprints.
Duplex DNA-unwinding assay.To prepare duplex DNA substrates, pPyori and pLPVori DNAs were digested with EcoRI and HindIII, and pPyxho and pL29 DNAs were digested with AccI. A fragment lacking an origin was obtained by digestion of pPyxho DNA with NocI and XbaI. These fragments were gel puri-fied, labeled with one [a-32
P]dNTP together with the other three unlabeled dNTPs by the Klenow fragment of DNA polymerase I, and then purified by NucTrap columns (Stratagene). The measurement of T antigen unwinding ac-tivity with duplex DNA fragments was performed essentially as described by Wang and Prives (117). Reaction mixtures (15ml) contained 80 mM Tris-HCl (pH 7.8), 40 mM creatine phosphate, 7 mM MgCl2, 0.5 mM DTT, 4 mM ATP,
1mg of creatine phosphate kinase, 1.0 ng of ori(1) or ori(2) [32P]DNA
frag-ments, 0.5mg of Escherichia coli SSB, 0.2mg of T antigen, and the indicated amounts of linear pBR322 competitor DNA. Mixtures were incubated at 338C with T antigen for 30 min, and the reactions were terminated by the addition of an equal volume of a solution containing 2% sodium dodecyl sulfate (SDS), 50 mM EDTA, and 2 mg of proteinase K per ml. The mixtures were further incubated at 378C for 30 min, treated with 2ml of loading buffer (20% Ficoll, 0.5% xylene cyanol, 0.5% bromophenol blue), and fractionated by electrophore-sis through 5% polyacrylamide gels containing 0.53TBE. DNA bands were visualized by autoradiography and quantitated with a PhosphorImager.
RESULTS
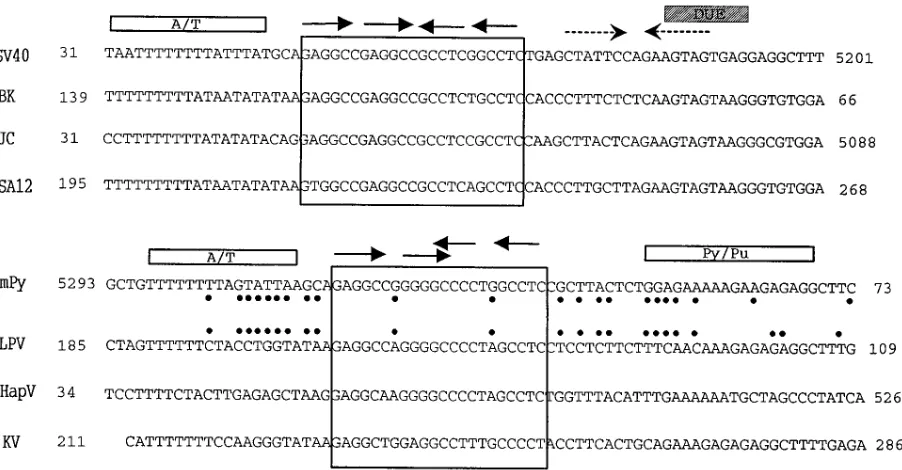
DNA sequence homologies among the core origins of
mam-malian polyomaviruses.
The core origins of replication of the
known primate-specific polyomaviruses SV40, BK virus, JC
virus, SA12, and LPV and the rodent-specific polyomaviruses
mPy, hamster polyomavirus, and K virus are highly
homolo-gous (Fig. 1). Notably, each has a central palindrome with four
PuPuGGC pentanucleotide repeats (the sequence initially
bound by the viral T antigen), an A/T-rich sequence on the late
side, and a sequence on the early side with either an imperfect
inverted repeat or a polypyrimidine-polypurine (Py/Pu) tract
which contains DNA-unwinding elements.
Previous studies have documented that the SV40 T antigen
will not replicate the mPy DNA origin in vitro (75, 104), nor
are the SV40 and the mPy core origins replicated in vivo by the
other’s T antigens (1, 51, 107). The topology of T antigen
binding to DNA might be responsible for some of this
repli-cation selectivity, as mutational analysis of the SV40 and mPy
origins has shown that altering the symmetry of the
pen-tanucleotide repeats profoundly affects DNA replication (2,
12, 17, 23, 81, 110, 114). While the central palindromes of the
origins of SV40, BK, JC, and SA12 viruses are nearly perfectly
symmetric about a single base between the second and third
pentanucleotide motifs, the second and third pentanucleotide
motifs of the central palindromes of the mPy, LPV, hamster
polyomavirus, and K viruses overlap (Fig. 1). Nevertheless,
even among the viral origins of SV40, BK, and JC viruses with
similar arrangements of pentanucleotide motifs, there are
pro-found differences in the efficiency with which the origins are
replicated by each other’s T antigens (66, 67, 98), indicating
on November 9, 2019 by guest
http://jvi.asm.org/
that features other than the arrangement of pentanucleotide
motifs in the central palindrome affect the replication
effi-ciency of naturally occurring origins. To further explore such
replication selectivity, we focused on the mPy and the primate
LPV viruses, which replicate in different species but whose
origins and T antigens are highly homologous.
Viral replication selectivity.
mPy infects mice and will
rep-licate to a limited extent in the cells of other rodents. There is
sparse evidence that it productively infects primates: a few
early studies suggested that it can be propagated in monkey
kidney cells (100, 108), and we have observed that it replicates
to a limited extent in certain monkey cells which express SV40
large T antigen (107). LPV was isolated from an African green
monkey and replicates in primate B lymphoblastoid cell lines
but not fibroblastic cells (83, 105). Its genome is more similar
to that of mPy than to those of the other known primate
polyomaviruses (47, 82, 84). The similarity of the mPy and LPV
core origins (Fig. 1) suggested to us that they might be
repli-cated by each other’s T antigen. However, in preliminary
ex-periments, we observed that this was not the case;
conse-quently, we analyzed mPy and LPV DNA replication in greater
detail to define those features important for this replication
selectivity.
Replication of mPy in vivo required a functional enhancer in
cis (31, 54, 91, 92, 106) (Fig. 2A, compare lanes 1 and 2), while
[image:3.612.80.531.70.305.2]the LPV genome introduced into mouse 3T6 fibroblasts did
not replicate (Fig. 2A, compare lanes 3 and 6). In human
lymphoid P3HR1 cells, a reciprocal pattern of replication
se-lectivity occurred, in that LPV replicated efficiently while mPy
did not (Fig. 2B, compare lane 3 to 4). Neither genome lacking
an enhancer replicated in human cells (Fig. 2B, lanes 1 and 2).
This cell type/species selectivity for replication may have
several causes. One might be that the viral enhancer functions
only in certain cell types and thereby restricts replication (30,
91, 92). Another cause might be incompatibility between the
viral T antigen and the replication proteins of the heterologous
FIG. 1. Alignment of the core origins of DNA replication of the known mammalian polyomaviruses. Numbers indicate positions in the physical map of each virus (15, 24, 25, 45, 46, 69, 82, 94, 110). The large boxes enclose the central palindromes containing the four pentanucleotide repeats (shown by arrows). The A/T-rich sequences and the Py/Pu sequences are indicated by rectangles, the imperfect inverted repeats are indicated by the dashed arrows, and the DNA-unwinding element (DUE) is indicated by the shaded box. Core origin sequence differences between mPy and LPV origins are indicated by dots.
FIG. 2. In vivo replication of wild-type and chimeric mPy and LPV DNAs. (A) Replication of viral DNAs in mouse 3T6 cells. Lane 1, pPyE2(mPy DNA lacking enhancer); lane 2, pPyBglII (mPy DNA with BglII linker at late side of core origin at nucleotide [nt] 5293, used as a parent for pPyE2); lane 3, pPy (mPy wild-type DNA); lane 4, pPyxho (mPy DNA with XhoI linker at nt 5293, used as a parent for pL29); lane 5, pL29 (mPy DNA with LPV sequences from nt 185 to 153 substituted for the equivalent mPy sequences between nt 5293 and nt 30); lane 6, pLPV (LPV wild-type DNA); lane 7, control (transfection without DNA). (B) Replication of viral DNAs in human P3HR1 cells. Lane 1, pPyE2; lane 2, pLPVE2(LPV DNA lacking enhancer); lane 3, pPy BglII; lane 4, pLPV. (C) Replication in human P3HR1 cells of viral origins cloned in pUC vectors with LPV T antigen expression. Lane 1, pPyori (mPy DNA from nt 5051 to 204); lane 2, pLPVori9(LPV DNA from nt 5050 to 760); lane 3, pLPVori (LPV DNA from nt 1 to 677); lane 4, pAH (LPV DNA from nt 677 to 394 joined to mPy DNA from nt 30 to 204); lane 5, pAN (mPy DNA from nt 5051 to 30 joined to LPV DNA from nt 153 to 1). (D) Replication of viral origins cloned infM13mp19 vectors in mouse 3T6 cells with mPy T antigen expression. Lane 1,fAH (AH chimeric LPV/mPy DNA); lane 2,fAN (AN chimeric LPV/mPy DNA); lane 3,fM13mp19 (vector control); lane 4,fPyori (mPy origin DNA); lane 5,fLPVori (LPV origin DNA).
on November 9, 2019 by guest
http://jvi.asm.org/
host (1, 9, 40, 74, 75, 93). A third cause might be the presence
of inhibitory proteins such as p53, which might bind one T
antigen but not the other (7, 116).
We attempted to distinguish between these possibilities by
assessing replication of an origin together with helper DNAs
capable of expressing a viral T antigen in cells which provide
the cognate accessory proteins capable of functioning with that
T antigen. When the mPy origin-enhancer DNA was
intro-duced into human P3HR1 cells together with a helper DNA
capable of expressing the LPV large T antigen, replication of
the mPy origin-enhancer was determined to be as efficient as
replication of the LPV origin-enhancer (Fig. 2C, lanes 1, 2, and
3), indicating that the LPV T antigen can form a functional
initiation complex at the mPy origin. However, the LPV
origin-enhancer was not replicated in mouse 3T6 cells when
cotrans-fected together with the mPy T antigen (Fig. 2D, compare
lanes 4 and 5), suggesting either that there is a cis-acting block
upon replication or that the mPy T antigen does not interact
productively with the LPV core origin.
To further analyze the basis for this replication selectivity,
we turned to in vitro assays which permit each step in the
assembly of the replication complex to be analyzed separately.
In vitro replication of mPy and LPV DNAs.
We asked
wheth-er the mPy T antigen togethwheth-er with a murine FM3A cell extract
capable of replicating mPy DNA can replicate an LPV origin
in vitro. This analysis indicated that the LPV origin was
repli-cated only minimally (Fig. 3, compare pPyori with pLPVori),
confirming the in vivo analysis reported above. To define the
DNA sequences that restrict replication of the LPV origin, we
constructed several chimeric DNAs and assessed their capacity
to be replicated by the mPy T antigen and the FM3A cell
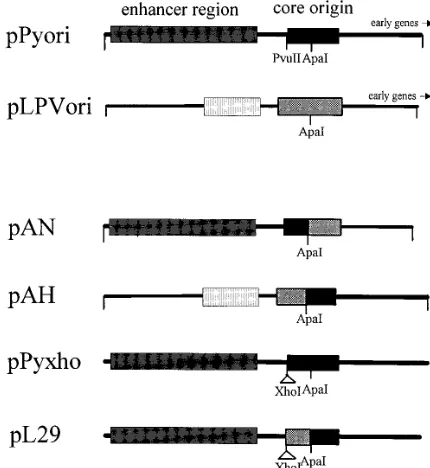
extract. Two chimeric DNAs were prepared by joining
se-quences cleaved at the ApaI sites in the middle of the central
palindromes of the mPy and LPV origins (Fig. 4). pAN DNA
contains the late half of the mPy core origin-enhancer joined to
the early half of the LPV origin, and pAH DNA contains the
late half of the LPV core origin-enhancer joined to the early
half of the mPy origin (Fig. 4). Assays of their replication in
vitro revealed that the pAN chimera supported replication
mediated by Py T antigen (approximately 75% of that of
pPyori DNA), whereas pAH supported only 5% or less of the
replication of pPyori DNA mediated by mPy T antigen (Fig. 3).
To more precisely define the LPV sequences which restrict
replication by the mPy T antigen, an additional DNA construct
(pL29), containing 36 bp of LPV sequence substituted for the
homologous mPy sequences on the late side of the core origin
(between the ApaI and PvuII sites), was prepared. It was not
replicated by the mPy T antigen together with the murine cell
extract, in contrast to the replication of the parent DNA
con-taining the mPy origin (pPyxho) (Fig. 3). This indicates that
sequences on the late side of the core origin determine
repli-cation selectivity.
Comparable results were obtained in an analysis of the
rep-lication of these chimeric DNAs in vivo. In mouse 3T6 cells,
the pAN DNA chimera was replicated by mPy T antigen but
the pAH DNA chimera was not (Fig. 2D, lanes 1 and 2), and
the chimeric pL29 DNA did not replicate in mouse 3T6 cells in
contrast to the replication of its parent pPyxho DNA (Fig. 2A,
lanes 4 and 5). Both the pAH and pAN DNAs were replicated
by LPV T antigen in human P3HR1 cells, in agreement with
the observation that the intact mPy origin-enhancer is
repli-cated in human cells by LPV T antigen (Fig. 2C).
The sequence differences between LPV and mPy which must
determine this replication selectivity include one transition in
the central palindrome and nine bases on the late side of the
core origin (Fig. 1). The base substitution in the central
palin-drome has been shown previously not to affect replication of
mPy DNA (110). Therefore, it must be the sequences between
the central palindrome and the late border of the core origin
that determine this replication selectivity. To further assess
how these sequence changes affect replication, we studied the
individual steps leading to the formation of an initiation
com-plex.
mPy T antigen induces structural changes in the origins of
mPy and LPV DNAs.
Early steps in the initiation of SV40 DNA
replication involve the specific binding of large T antigen to the
central palindrome, the assembly of a double hexamer with
extended contacts on the flanking sequences, and the
subse-FIG. 4. Structures of wild-type and chimeric origins of replication. pPyori contains the origin-enhancer of mPy DNA from BclI (nt 5051) to DdeI (nt 204). pLPVori contains the origin-enhancer of LPV DNA from AhaIII (nt 677) to
NcoI (nt 1). pAN consists of the mPy sequences from nt 5051 to nt 30 joined at
the ApaI site to the LPV sequences from nt 153 to nt 1. pAH consists of the LPV sequences from nt 677 to nt 394 joined at the ApaI site to mPy sequences from nt 30 to nt 204. pPyxho contains the entire mPy DNA genome with an 8-bp XhoI linker inserted at the PvuII site (nt 5293). pL29 contains the entire mPy DNA genome with a substitution of LPV sequences from nt 154 to nt 185 between the
[image:4.612.325.540.408.642.2]ApaI and XhoI sites of pPyxho.
FIG. 3. In vitro replication of wild-type and chimeric mPy and LPV origin DNAs mediated by mPy T antigen in murine FM3A extracts. Reaction mixtures containing 0.5 mg of plasmid DNA with cell extract in the absence (2) or presence (1) of 1.0mg of mPy T antigen were incubated at 338C for 2 h and analyzed as described in Materials and Methods.
on November 9, 2019 by guest
http://jvi.asm.org/
quent structural distortion of these flanking elements (3, 4, 16,
52, 68, 79, 119, 121). During this process, the A/T-rich domain
on the late side of the core origin is untwisted, and an
eight-base region in the early palindrome domain on the early side is
melted. The latter site coincides with the DNA-unwinding
el-ement predicted by thermodynamic calculations (63) and with
the origin of bidirectional replication mapped in vivo (28, 55,
56). The degree of structural change induced by T antigen in
the two flanking regions of the SV40 core ori region has been
shown to correlate with the efficiency of initiation of DNA
replication (2). This paradigm is likely to be applicable to all
polymavirus origins, including that of mPy, whose T antigen
binds to the same motifs as does that of SV40 (33, 85, 110).
To determine whether similar structural changes in mPy
DNA are induced by mPy T antigen and whether sequences on
the late side of the core origin affect such changes, we used
KMnO
4modification of DNA. KMnO
4reacts poorly with
nor-mal double-stranded DNA; however, when DNA is melted or
bent sharply (such as by a DNA-binding protein), specific sites
in the DNA become accessible to KMnO
4oxidation (4, 6). The
oxidized nucleotides interfere with elongation by the Klenow
en-zyme, and their location can be determined precisely by
compar-ison with sequence ladders on a denaturing sequence gel.
In the absence of mPy T antigen, minimal levels of KMnO
4modification were detected within the mPy core origin region
(Fig. 5A, lanes 1, 3, 5, 7, 9, and 11). Upon addition of mPy T
antigen, striking enhancements of KMnO
4modification were
observed in the Py/Pu region of the mPy core origin, coinciding
with the Py/Pu region which corresponds with the early
palin-drome (DNA unwinding element) of SV40. Notably, mPy T
antigen mediated similar patterns of KMnO
4modification in
the pPyxho and the pL29 DNAs, even though the latter was not
replicated by mPy T antigen. Similarly, mPy T antigen
dis-torted the pAH, pAN, and pLPVori DNAs even though these
DNAs differed greatly in their replication efficiency with mPy T
antigen. The KMnO
4modification of these DNAs caused by
mPy T antigen generally was much weaker than the KMnO
4modification induced by SV40 T antigen reacting with SV40
DNA (our unpublished observations) and was extremely
diffi-cult to quantify reliably. This may be due to the weaker binding
of mPy T antigen to its core origin sequences. Nevertheless, in
numerous experiments, it was always apparent that mPy T
antigen distorted the early region of the core origin.
The ability of mPy T antigen to induce structural changes in
the opposite strand of these DNA constructs was also
exam-ined. A high background level of KMnO
4oxidation throughout
the core origin was observed in the absence of T antigen (Fig.
5B, lanes 1, 3, 5, 7, 9, and 11). Addition of mPy T antigen
induced distortion of the A/T-rich region in all the DNA
con-structs, and as with the modification of the other strand, the
level of KMnO
4modification did not correlate with the
repli-cation capacity of these DNAs (Fig. 5B, lanes 2, 4, 6, 8, 10, and
12).
In summary, these results show that mPy T antigen binds to
the mPy and LPV core origins and causes structural changes
on both sides of the central palindrome. The pattern and
extent of structural changes do not correlate with the capacity
of the origins to be replicated by mPy T antigen either in vivo
or in vitro. This suggests that steps subsequent to the distortion
of the core origin determine the capacity of these DNAs to be
replicated by the mPy T antigen.
Unwinding of heterologous origins by mPy T antigens.
[image:5.612.336.523.80.641.2]Fol-lowing structural deformation of the viral core origin, the
he-licase activity of SV40 T antigen induces extensive unwinding
of the DNA in concert with a single-stranded-DNA-binding
protein and topoisomerase I (3, 8, 34, 52, 59, 79, 88, 99, 114,
FIG. 5. KMnO4footprints of wild-type and chimeric mPy and LPV origins.
The DNAs (0.5mg) were incubated at 338C for 30 min with or without mPy T antigen in the presence of 4 mM AMPPNP and then incubated with KMnO4for
30 s. Sites of KMnO4modification were determined by primer extension. (A)
Footprints of KMnO4-reactive sites with primers (PyL5190 and LPV520)
com-plementary to the bottom strands (the adenine-rich strand in the A/T region of the core origin). (B) Footprints of KMnO4-reactive sites with primers (PyE167
and LPV0) complementary to the top strands (the thymine-rich strand in the A/T region of the core origin). The locations of domains of the core origin of replication are shown to the left of each set of lanes.
on November 9, 2019 by guest
http://jvi.asm.org/
121). The bidirectional DNA helicase activity of SV40 T
anti-gen is closely correlated with its replication capacity (17, 34, 52,
71, 114).
Previous studies have revealed that the SV40 and mPy T
antigens unwind duplex ori(
1
) and ori(
2
) DNA fragments
with comparable efficiency but that addition of nonspecific
competitor DNA causes the T antigens to unwind duplex
DNAs in an origin-specific manner (52, 117). In agreement
with these observations, we found that the mPy T antigen
unwound a DNA lacking the mPy origin (ori) as well as DNAs
containing the mPy or LPV origin with comparable efficiencies
in the absence of nonspecific competitor DNA (Fig. 6A, lanes
2, 7, and 12); however, with 0.1
m
g or more of linear pBR322
competitor DNA, the mPy T antigen poorly unwound ori(
2
)
DNA and the LPV ori DNA, while unwinding of the mPy ori
DNA fragment was reduced only slightly (Fig. 6A and B).
Similar analyses of the unwinding of pL29 DNA and that of its
parent, pPyXHO, revealed that the pPyxho DNA was
un-wound efficiently, while the pL29 ori fragment was inefficiently
unwound (Fig. 6C and D). These data indicate that the origins
which support replication by mPy T antigen are more readily
unwound in the presence of nonspecific competitor DNA.
In conclusion, these experiments indicate that the failure of
the mPy T antigen to replicate the LPV origin correlates with
the efficiency with which the origin is unwound and that
se-quences on the late side of the core origin help determine the
capacity of the mPy T antigen to act as an efficient unwinding
protein.
DISCUSSION
The initiation of DNA replication at a specific site on a
circular duplex DNA requires the identification of the origin by
a sequence-specific protein(s) and, subsequently, opening of
the DNA and synthesis of primers from which the leading and
lagging strands can be extended. The polyomaviruses utilize
the T antigen to nucleate the assembly of a replication complex
responsible for this process. The pentanucleotide-binding sites
recognized by the T antigens of the various members of this
family of viruses are highly conserved, yet several viruses have
evolved significant replication selectivity. How can this be
ex-plained?
One reason occurs in the specific interactions between the T
antigens and the cellular proteins which interact to assemble a
functional initiation complex. There is abundant evidence,
al-beit somewhat conflicting, that the T antigens of these viruses
interact differentially with the DNA polymerase
a
/primases of
different species (9, 40, 74, 75, 93). Such protein-protein
inter-actions probably change in concert with evolution of the
rep-lication machinery of the host cell.
Another means by which replication selectivity might occur
is through changes in the cellular proteins capable of activating
or inhibiting origin function. These proteins are believed to
function by preventing nucleosome occlusion or by affecting
assembly or function of the initiation complex (29). The
as-semblage of cellular proteins present in a cell which are
capa-ble of acting upon a specific viral core origin, auxiliary and
enhancer sequences, or the T antigen undoubtedly will dictate
the replication efficiency of that origin.
Our experiments suggest a third basis for replication
selec-tivity, namely, that it can occur because of a deficiency in the
capacity of the viral T antigen to efficiently unwind an origin
and that the sequences in the origin help dictate this process.
Regulation of the T antigen unwinding activity has been
sug-gested to be a key control point for replication (32, 34, 76, 89,
90). Only 2 to 3% of the T antigen-origin complex undergoes
FIG. 6. Unwinding of duplex DNA by mPy T antigen. (A and C) Represen-tative autoradiographs of the unwinding of ori(2), mPy ori, and LPV ori DNAs (A) and pPyxho ori and pL29 ori DNAs (C) by mPy T antigen in the absence of or with increasing amounts of unlabeled linear pBR322 competitor DNA. ss and ds, single- and double-stranded, respectively. (B and D) Plots of duplex DNA unwound as a function of competitor DNA. Incubations with labeled ori(1) or ori(2) DNA fragment were performed as described in Materials and Methods in the absence and presence of the indicated amounts of unlabeled linear pBR322 competitor DNA. Unwinding was quantitated by PhosphorImager and normal-ized to 50% in the absence of competitor DNA. The indicated values are averages of multiple independent experiments. The bars represent the standard error of the mean.
on November 9, 2019 by guest
http://jvi.asm.org/
unwinding in vitro (76). Control of unwinding may occur by
phosphorylation or dephosphorylation of specific residues in T
antigen (11, 43, 71, 73, 86, 114). It is apparent from our results
that sequence differences between origins also can affect the
conversion of T antigen from an origin-binding protein to an
origin-unwinding protein.
The studies of Lynch and Frisque (65) also indicated that
sequences on the late side of a viral origin help determine
replication specificity (between JC and SV40 viruses), and this
was shown to occur at a point beyond T antigen binding. Our
results extend their analysis to additional viruses and focus
upon a specific step in the replication pathway. These
obser-vations expand the significance of the switch in T antigen
activity from an origin-binding protein to an unwinding
activ-ity, to include a means by which the efficiency of DNA
repli-cation from an origin might be constitutively regulated. One
obvious circumstance when this might be advantageous is in
the event that a cell is infected by two distinct viruses; each
virus would benefit were its own T antigen not capable of
activating the other’s origin.
Perhaps another, more profound consequence of such
rep-lication selectivity is in the opportunity it provides to
deter-mine the timing of initiation of cellular DNA replication after
an initiation complex has bound to an origin. For example,
eukaryotic chromosomes contain multiple origins which
repli-cate at distinct times in S phase, but the mechanism for such
temporal control is not understood. Quantitative assessments
show that specific yeast origins have efficiencies of activation
that range from less than 20% to greater than 90% per cell
cycle (38, 42, 44). One means for such control is likely to be
through specific sequences affecting the efficiency (and perhaps
the rate) at which the initiation complexes initiate unwinding
of the DNA.
Recent studies of the interaction of SV40 T antigen with the
SV40 origin have suggested that the generation of structural
changes in the origin is mediated primarily through the
sugar-phosphate backbone and that sequence changes in the flanking
A/T element or early palindrome which diminish replication
do so solely because they cause these sequences to be resistant
to the structural alterations induced by T antigen (95). Our
data are not in accord with this suggestion. We find that the
restriction of mPy T antigen by the LPV origin does not
cor-relate with the capacity of the mPy T antigen to induce
struc-tural deformation but does correlate with its capacity to induce
unwinding. Similar analyses of the interaction of SV40 T
an-tigen with mutant origins and of mutant T anan-tigens acting at
the SV40 origin have led to a similar correlation (71, 81). In a
complementary study Bhattacharyya et al. (1a) demonstrate
that mPy and SV40 T antigens both produce structural
alter-ations in each other’s origins, but nevertheless, these T
anti-gens fail to replicate the respective heterologous origin.
The correlation between replication selectivity and the
se-quences on the late side of the origin and the effect of these
same sequences upon unwinding suggest that the replication
defect is caused at least in part by inefficient unwinding.
Nev-ertheless, the apparent replication defect appears to be greater
than the unwinding defect, which suggests either that the
un-winding assay is less discriminating than the replication assays
or that other factors might also be involved in the replication
selectivity. Both reasons seem likely: the unwinding assay with
SV40 T antigen has previously been found to be less
discrim-inating with origin mutants than are replication assays (17),
and mPy T antigen is much more promiscuous about the DNA
sequences it chooses to unwind than is the SV40 T antigen
(117).
We also note that our in vitro unwinding assays with T
antigens and duplex DNA fragments have been carried out
under artificial conditions and that many proteins which help
participate in the replication complex are absent.
Incorpora-tion of these proteins into our assays might reveal more
spe-cifically how sequences of the core origin affect unwinding. For
example, it has been reported that the A/T-rich flanking motif
is required for the stimulation of T antigen-dependent origin
DNA unwinding by DNA polymerase
a
(76). It is this same
region which we have found to be important in the replication
selectivity by mPy T antigen and which is responsible for the
replication selectivity of JC virus T antigen with its origin (65).
Since the p48 primase subunit helps determine the
species-specific initiation of Py DNA replication (9, 40), we wonder
whether the primosome has a special affinity for sequences in
this region of the core origin, so as to more efficiently initiate
primer synthesis (a suggestion which parallels that proposed by
Schneider et al. [93]). These same sequences induce bending in
DNA and are also known to be capable of binding a variety of
cellular proteins which might affect T antigen and polymerase
a
/primase activity (10, 21, 48, 57, 60, 109). Additionally,
aux-iliary sequences beyond the core origin bind T antigen and
cellular activators and thereby promote DNA unwinding (52a,
53). Mutations of the A/T tract in the core origin can be
compensated for by changes in these auxiliary sequences (50);
hence, there are likely to be specific interactions between
them. Understanding these additional interactions should help
explain how activation of the DNA replication complex is
mod-ulated within the cell.
ACKNOWLEDGMENTS
We thank B. Schaffhausen and D. Pallas for gifts of baculovirus vectors and hybridomas; Carol Prives, J. Dudley, V. Bermudez, Wei-dong Guo, and X. Wang for advice; and Mark Hannink and David Pintel for comments on the manuscript. Sarah Scanlon, as always, helped ensure that everything worked.
Financial support was provided by USPHS grant CA38538.
REFERENCES
1. Bennett, E. R., M. Naujokas, and J. A. Hassell. 1989. Requirements for species-specific papovavirus DNA replication. J. Virol. 63:5371–5385. 1a.Bhattacharyya, S., H. E. Lorimer, and C. Prives. 1995. Murine
polyomavi-rus and simian vipolyomavi-rus 40 large T antigens produce different structural alter-ations in viral origin DNA. J. Virol. 69:7579–7585.
2. Borowiec, J. A. 1992. Inhibition of structural changes in the simian virus 40 core origin of replication by mutation of essential origin sequences. J. Virol. 66:5248–5255.
3. Borowiec, J. A., F. B. Dean, P. A. Bullock, and J. Hurwitz. 1990. Binding and unwinding—how T antigen engages the SV40 origin of DNA replication. Cell 60:181–184.
4. Borowiec, J. A., and J. Hurwitz. 1988. Localized melting and structural changes in the SV40 origin of replication induced by T antigen. EMBO J. 7:3149–3158.
5. Borowiec, J. A., and J. Hurwitz. 1988. ATP stimulates the binding of SV40 T-antigen to the origin of replication. Proc. Natl. Acad. Sci. USA 85:64–68. 6. Borowiec, J. A., L. Zhang, S. Sasse-Dwight, and J. D. Gralla. 1987. DNA supercoiling promotes formation of a bent repression loop in lac DNA. J. Mol. Biol. 196:101–111.
7. Braithwaite, A. W., H. N. Sturzbecher, C. Addison, C. Palmer, K. Rudge, and J. R. Jenkins.1987. Mouse P53 inhibits SV40 origin-dependent DNA replication. Nature (London) 329:458–460.
8. Brill, S. J., and B. Stillman. 1989. Yeast replication factor-A functions in the unwinding of the SV40 origin of DNA replication. Nature (London) 342:92–95.
9. Bru¨ckner, A., F. Studlbauer, L. A. Guarino, A. Brunahl, C. Schneider, C. Rehfuess, C. Prives, E. Fanning, and H.-P. Nashevu.1995. The mouse DNA polymerasea-primase subunit mediates species-specific replication of polyomavirus DNA in vitro. Mol. Cell. Biol. 15:1716–1724.
10. Carmichael, E. P., J. M. Roome, and A. F. Wahl. 1993. Binding of a sequence-specific single-stranded DNA-binding factor to the simian virus 40 core origin inverted repeat domain is cell cycle regulated. Mol. Cell. Biol. 13:408–420.
11. Cegielska, A., and D. M. Virshup. 1993. Control of simian virus 40 DNA
on November 9, 2019 by guest
http://jvi.asm.org/
replication by HeLa cell nuclear kinase casein kinase 1. Mol. Cell. Biol. 13:1201–1211.
12. Cohen, G. L., P. J. Wright, A. L. Delucia, B. A. Lewton, M. E. Anderson, and P. Tegtmeyer.1984. Critical spatial requirement within the origin of simian virus 40 DNA replication. J. Virol. 51:91–96.
13. Collins, K. L., and T. J. Kelly. 1991. Effects of T antigen and replication protein A on the initiation of DNA synthesis by DNA polymerasea -pri-mase. Mol. Cell. Biol. 11:2108–2115.
14. Collins, K. L., A. S. R. Russo, B. Y. Tseng, and T. J. Kelly. 1993. The role of the 70 kDa subunit of human polymeraseain DNA replication. EMBO J. 12:4555–4566.
15. Cunningham, T. P., and J. M. Pipas. 1985. Simian agent 12 is a BK virus-like papovavirus which replicates in monkey cells. J. Virol. 54:483– 492.
16. Dean, F. B., J. A. Borowiec, T. Eki, and J. Hurwitz. 1992. The simian virus 40 T antigen double hexamer assembles around the DNA at the replication origin. J. Biol. Chem. 267:14129–14137.
17. Dean, F. B., J. A. Borowiec, Y. Ishimi, S. Deb, P. Tegtmeyer, and J. Hurwitz. 1987. Simian virus 40 large tumor antigen requires three core replication origin domains for DNA unwinding and replication in vitro. Proc. Natl. Acad. Sci. USA 84:8267–8271.
18. Dean, F. B., P. Bullock, Y. Murakami, C. R. Wobbe, L. Weissbach, and J. Hurwitz.1987. Simian virus 40 (SV40) DNA replication: SV40 large T antigen unwinds DNA containing the SV40 origin of replication. Proc. Natl. Acad. Sci. USA 84:16–20.
19. Dean, F. B., M. Dodson, H. Echols, and J. Hurwitz. 1987. ATP-dependent formation of a specialized nucleoprotein structure by simian virus 40 (SV40) large tumor antigen at the SV40 replication origin. Proc. Natl. Acad. Sci. USA 84:8981–8985.
20. Deb, S., A. DeLucia, C. Baur, A. Koff, and D. Tegtmeyer. 1986. Domain structure of the simian virus 40 core origin of replication. Mol. Cell. Biol. 6:1663–1670.
21. Deb, S., A. DeLucia, A. Koff, S. Tsui, and P. Tegtmeyer. 1986. The AT domain of the simian virus 40 core origin directs DNA binding and coor-dinately regulated DNA replication. Mol. Cell. Biol. 6:4578–4584. 22. Deb, S., and P. Tegtmeyer. 1987. ATP enhances the binding of simian virus
40 large T antigen to the origin of replication. J. Virol. 61:3649–3654. 23. Deb, S., S. Tsui, A. Koff, A. L. Delucia, R. Parson, and P. Tegtmeyer. 1987.
The T-antigen-binding domain of the simian virus 40 core origin of repli-cation. J. Virol. 61:2143–2149.
24. Deininger, P. L., A. Esty, P. Laporte, H. Hsu, and T. Friedmann. 1980. The nucleotide sequence and restriction enzyme sites of the polyoma genome. Nucleic Acids Res. 8:855–860.
25. Delmas, V., C. Bastien, S. Scherneck, and J. Feunteun. 1985. A new mem-ber of the polyomavirus family: the hamster papovavirus. Complete nucle-otide sequence and transformation properties. EMBO J. 4:1279–1286. 26. DeLucia, A. L., B. A. Lewton, R. Tjian, and P. Tegtmeyer. 1983. Topography
of simian virus 40 protein-DNA complexes: arrangement of pentanucle-otide interaction sites at the origin of replication. J. Virol. 46:143–150. 27. Denis, D., and P. A. Bullock. 1993. Primer-DNA formation during simian
virus 40 DNA replication in vitro. Mol. Cell. Biol. 13:2882–2890. 28. DePamphilis, M. L. 1993. Eukaryotic DNA replication: anatomy of an
origin. Annu. Rev. Biochem. 62:29–63.
29. DePamphilis, M. L. 1993. How transcription factors regulate origins of DNA replication in eukaryotic cell. Trends Cell Biol. 3:161–167. 30. DePolo, N. J., and L. P. Villarreal. 1991. E1A represses wild-type and
F9-selected polyomavirus DNA replication by a mechanism not requiring depression of large tumor antigen transcription. J. Virol. 65:2921–2928. 31. de Villiers, J., W. Schaffner, C. Tyndall, S. Lupton, and R. Kamen. 1984.
Polyoma virus DNA replication requires an enhancer. Nature (London) 312:242–246.
32. Diller, J. D., and M. K. Raghuraman. 1994. Eukaryotic replication origins: control in space and time. Trends Biochem. Sci. 19:320–325.
33. Dilworth, S. M., A. Cowie, R. Kamen, and B. Griffin. 1984. DNA binding activity of polyomavirus large tumor antigen. Proc. Natl. Acad. Sci. USA 81:1941–1945.
34. Dodson, M., F. B. Dean, P. Bullock, H. Echols, and J. Hurwitz. 1987. Unwinding of duplex DNA from the SV40 origin of replication by T anti-gen. Science 238:964–967.
35. Dornreiter, I., W. C. Copeland, and T. S.-F. Wang. 1993. Initiation of simian virus 40 DNA replication requires the interaction of a specific domain of human DNA polymeraseawith large T antigen. Mol. Cell. Biol. 13:809– 820.
36. Dornreiter, I., L. F. Erdile, I. U. Gilbert, D. von Winkler, T. J. Kelly, and E. Fanning.1992. Interaction of DNA polymerasea-primase with cellular replication protein A and SV40 T antigen. EMBO J. 11:769–776. 37. Dornreiter, I., A. Hoss, A. K. Arthur, and E. Fanning. 1990. SV40 T antigen
binds directly to the catalytic subunit of DNA polymerasea. EMBO J. 9:3329–3336.
38. Dubey, D. D., L. R. Davis, S. A. Greenfeder, L. Y. Ong, J. G. Zhu, J. R. Broach, C. S. Newlon, and J. A. Huberman.1991. Evidence suggesting that the ARS elements associated with silencers of the yeast mating-type locus
HML do not function as chromosomal DNA replication origins. Mol. Cell. Biol. 11:5346–5355.
39. Edenberg, H. J., S. Anderson, and M. L. DePamphilis. 1978. Involvement of DNA polymerase alpha in simian virus 40 DNA replication. J. Biol. Chem. 253:3273–3280.
40. Eki, T., T. Enomoto, C. Masutani, A. Miyajima, R. Takada, Y. Murakami, T. Ohno, F. Hanaoka, and M. Ui.1991. Mouse DNA primase plays the principal role in determination of permissiveness for polyomavirus DNA replication. J. Virol. 65:4874–4881.
41. Erdile, L. F., K. L. Collins, A. Russo, P. Simancek, D. Small, C. Umbricht, D. Virshup, L. Cheng, S. Randall, D. Weinberg, I. Moarefi, E. Fanning, and T. Kelly.1991. Initiation of SV40 DNA replication: mechanism and control. Cold Spring Harbor Symp. Quant. Biol. 56:303–313.
42. Fangman, W. L., and B. J. Brewer. 1992. A question of time: replication origins of eukaryotic chromosomes. Cell 71:363–366.
43. Fanning, E. 1994. Control of SV40 DNA replication by protein phosphor-ylation: a model for cellular DNA replication? Trends Cell Biol. 4:250–255. 44. Ferguson, B. M., and W. L. Fangman. 1992. A position effect on the time
of replication origin activation in yeast. Cell 68:333–339.
45. Fiers, W., R. Contreras, G. Haegemann, R. Rogiers, A. van de Voorde, H. van Heuverswyn, J. van Herreweghe, G. Volchaert, and M. Ysebaert.1978. Complete nucleotide sequence of SV40 DNA. Nature (London) 273:113– 120.
46. Frisque, R. J., G. L. Bream, and M. T. Cannella. 1984. Human polyoma-virus JC polyoma-virus genome. J. Virol. 51:458–469.
47. Furuno, A., T. Miyamura, and K. Yoshiike. 1984. Monkey B-lymphotropic papovavirus DNA: nucleotide sequence of the region around the origin of replication. J. Virol. 50:451–456.
48. Galli, I., S. M. Iguchi-Ariga, and H. Ariga. 1992. The AT-rich tract of the SV40 ori core: negative synergism and specific recognition by single stranded and duplex DNA binding proteins. Nucleic Acids Res. 20:3333– 3339.
49. Gannon, J., and D. P. Lane. 1990. P53 and DNA polymeraseacompete for binding to SV40 T antigens. Nature (London) 329:456–458.
50. Gerard, R., and Y. Gluzman. 1986. Functional analysis of the role of the A1T-rich region and upstream flanking sequences in simian virus 40 DNA replication. Mol. Cell. Biol. 6:4570–4577.
51. Gerard, R. D., R. A. Guggenheimer, and Y. Gluzman. 1987. Analysis of nonpermissivity in mouse cells overexpressing simian virus 40 T antigen. J. Virol. 61:851–857.
52. Goetz, G. S., F. B. Dean, J. Hurwitz, and S. W. Matson. 1988. The unwind-ing of duplex regions in DNA by the simian virus 40 large tumor antigen-associated DNA helicase activity. J. Biol. Chem. 263:383–392.
52a.Guo, W., and W. Folk. Unpublished data.
53. Gutierrez, C., Z. S. Guo, J. Roberts, and M. L. DePamphilis. 1990. Simian virus 40 origin auxiliary sequences weakly facilitate T-antigen binding but strongly facilitate DNA unwinding. Mol. Cell. Biol. 10:1719–1728. 54. Hassell, J. A., W. J. Muller, and C. R. Muller. 1986. The dual role of the
polyomavirus enhancer in transcription and DNA replication. Cancer Cells 4:561–569.
55. Hay, R. T., and M. L. DePamphilis. 1982. Initiation of SV40 DNA repli-cation in vivo: lorepli-cation and structure of the 59ends of DNA synthesized in the ori region. Cell 28:767–779.
56. Hendrickson, E. A., C. E. Fritze, W. R. Folk, and M. L. DePamphilis. 1987. The origin of bidirectional DNA replication in polyomavirus. EMBO J. 6:2011–2018.
57. Hertz, G. Z., M. R. Young, and J. R. Mertz. 1987. The AT-rich sequence of the simian virus 40 origin is essential for replication and is involved in bending of the viral DNA. J. Virol. 81:2322–2325.
58. Hurwitz, J., F. B. Dean, A. D. Kwong, and S.-H. Lee. 1990. The in vitro replication of DNA containing the SV40 origin. J. Biol. Chem. 265:18043– 18046.
59. Kenny, M. K., S. H. Lee, and J. Hurwitz. 1989. Multiple functions of human single-stranded-DNA binding protein in simian virus 40 DNA replication: single-strand stabilization and stimulation of DNA polymeraseaandd. Proc. Natl. Acad. Sci. USA 86:9757–9761.
60. Kilwinski, J., M. Baack, S. Herland, and R. Knippers. 1995. Transcription factor Oct1 binds to the AT-rich segment of the simian virus 40 replication origin. J. Virol. 69:575–578.
61. Lee, S. H., T. Eki, and J. Hurwitz. 1989. Synthesis of DNA containing the simian virus 40 origin of replication by the combined action of DNA polymerases alpha and delta. Proc. Natl. Acad. Sci. USA 86:7361–7365. 62. Li, J. J., K. W. Peden, R. A. Dixon, and T. J. Kelly. 1986. Functional
organization of the simian virus 40 origin of DNA replication. Mol. Cell. Biol. 6:1117–1128.
63. Lin, S., and D. Kowalski. 1994. DNA helical instability facilitates initiation at the SV40 replication origin. J. Mol. Biol. 235:496–507.
64. Lorimer, H. E., E. H. Wang, and C. Prives. 1991. The DNA-binding prop-erties of polyomavirus large T antigen are altered by ATP and other nucle-otides. J. Virol. 65:687–699.
65. Lynch, K. J., and R. J. Frisque. 1990. Identification of critical elements within the JC virus DNA replication origin. J. Virol. 64:5812–5822.
on November 9, 2019 by guest
http://jvi.asm.org/
66. Lynch, K. J., and R. J. Frisque. 1991. Factors contributing to the restricted DNA replicating activity of JC virus. Virology 180:306–317.
67. Lynch, K. L., S. Haggarty, and R. J. Frisque. 1994. DNA replication of chimeric JC virus-SV40 genomes. Virology 204:819–822.
68. Mastrangelo, I. A., P. V. Hough, J. S. Wall, M. Dodson, F. B. Dean, and J. Hurwitz.1989. ATP-dependent assembly of double hexamers of SV40 T antigen at the viral origin of DNA replication. Nature (London) 338:658– 662.
69. Mayer, M., and K. Doerries. 1991. Nucleotide sequence and genome or-ganization of the murine polyomavirus, Kilham strain. Virology 181:469– 480.
70. McCutchan, J. H., and J. S. Pagano. 1968. Enhancement of the infectivity of simian virus 40 deoxyribonucleic acid with diethylaminoethyl-dextran. J. Natl. Cancer Inst. 41:351–357.
71. McVey, D., S. Ray, Y. Gluzman, L. Berger, A. G. Wildeman, D. R. Marshak, and P. Tegtmeyer.1993. cdc2 phosphorylation of threonine 124 activates the origin-unwinding functions of simian virus 40 T antigen. J. Virol. 67: 5206–5215.
72. Melendy, T., and B. Stillman. 1993. An interaction between replication protein A and SV40 T antigen appears essential for primosome assembly during SV40 DNA replication. J. Biol. Chem. 268:3389–3395.
73. Moarefi, I. F., D. Small, I. Gilbert, M. Hopfner, S. K. Randall, C. Schnei-der, A. A. Russo, U. Ramsperger, A. K. Arthur, H. Stahl, T. J. Kelly, and E. Fanning.1993. Mutation of the cyclin-dependent kinase phosphorylation site in simian virus 40 (SV40) large T antigen specifically blocks SV40 origin DNA unwinding. J. Virol. 67:4992–5002.
74. Moses, K., and C. Prives. 1994. A unique subpopulation of murine DNA polymerasea/primase specifically interacts with polyomavirus T antigen and stimulates DNA replication. Mol. Cell. Biol. 14:2767–2776. 75. Murakami, Y., T. Eki, M. Yamada, C. Prives, and J. Hurwitz. 1986.
Species-specific in vitro synthesis of DNA containing the polyoma virus origin of replication. Proc. Natl. Acad. Sci. USA 83:6347–6351.
76. Murakami, Y., and J. Hurwitz. 1993. DNA polymerase alpha stimulates the ATP-dependent binding of simian virus tumor T antigen to the SV40 origin of replication. J. Biol. Chem. 268:11018–11027.
77. Nethanel, T., and G. Kaufmann. 1990. Two DNA polymerases may be required for synthesis of the lagging DNA strand of simian virus 40. J. Virol. 64:5912–5918.
78. Pallas, D. C., C. Schley, M. Mahoney, E. Harlow, B. S. Schaffhausen, and T. M. Roberts.1986. Polyomavirus small t antigen: overproduction in bac-teria, purification, and utilization for monoclonal and polyclonal antibody production. J. Virol. 60:1075–1084.
79. Parsons, R., M. E. Anderson, and P. Tegtmeyer. 1990. Three domains in the simian virus 40 core origin orchestrate the binding, melting, and DNA helicase activities of T-antigen. J. Virol. 64:509–518.
80. Parsons, R. E., J. E. Stenger, S. Ray, R. Welker, M. E. Anderson, and P. Tegtmeyer.1991. Cooperative assembly of simian virus 40 T-antigen hex-amers on functional halves of the replication origin. J. Virol. 65:2798–2806. 81. Parsons, R., and P. Tegtmeyer. 1992. Spacing is crucial for coordination of domain functions within the simian virus 40 core origin of replication. J. Virol. 66:1933–1942.
82. Pawlita, M., A. Glad, and H. zur Hausen. 1985. Complete DNA sequence of lymphotropic papovavirus: prototype of a new species of the polyoma-virus genus. Virology 143:196–211.
83. Pawlita, M., L. Mosthaf, A. Clad, and P. Gruss. 1984. Genome structure and host range restriction of the lymphotropic papovavirus (LPV): identi-fication of a viral lymphocyte specific enhancer element. Curr. Top. Micro-biol. Immunol. 113:26–30.
84. Pipas, J. M. 1992. Common and unique features of T antigens encoded by the polyomavirus group. J. Virol. 66:3979–3985.
85. Pomerantz, B. J., and J. A. Hassell. 1984. Polyomavirus and simian virus 40 large T antigens bind to common DNA sequences. J. Virol. 49:925–937. 86. Prives, C. 1990. The replication functions of SV40 T antigen are regulated
by phosphorylation. Cell 61:735–738.
87. Prives, C., Y. Murakami, F. G. Kern, W. Folk, C. Basilica, and J. Hurwitz. 1987. DNA sequence requirements for replication of polyomavirus DNA in vivo and in vitro. Mol. Cell. Biol. 7:3694–3704.
88. Roberts, J. M. 1989. Simian virus 40 (SV40) large tumor antigen causes stepwise changes in SV40 origin structure during initiation of DNA repli-cation. Proc. Natl. Acad. Sci. USA 86:3939–3943.
89. Roberts, J. M. 1993. Turning DNA replication on and off. Curr. Opin. Cell Biol. 5:201–206.
90. Roberts, J. M., and G. D’Urso. 1989. Cellular and viral control of the initiation of DNA replication. J. Cell Sci. Suppl. 12:171–182.
91. Rochford, R., B. A. Campbell, and L. P. Villarreal. 1987. A pancreas specificity results from the combination of polyomavirus and Moloney mu-rine leukemia virus enhancer. Proc. Natl. Acad. Sci. USA 84:449–453. 92. Rochford, R., B. A. Campbell, and L. P. Villarreal. 1990. Genetic analysis
of the enhancer requirements for polyomavirus DNA replication in mice. J. Virol. 64:476–485.
93. Schneider, C., K. Weibhart, L. A. Guarino, I. Dornreiter, and E. Fanning.
1994. Species-specific functional interactions of DNA polymerasea -pri-mase with simian virus 40 (SV40) T antigen require SV40 origin DNA. Mol. Cell. Biol. 14:3176–3185.
94. Seif, I., G. Khoury, and R. Dhar. 1979. The genome of human papovavirus BKV. Cell 18:963–977.
95. SenGupta, D. J., and J. A. Borowiec. 1994. Strand and face: the topography of interactions between the SV40 origin of replication and T antigen during the initiation of replication. EMBO J. 13:982–992.
96. Shortle, D. R., R. F. Margolskee, and D. Nathans. 1979. Mutational analysis of the simian virus 40 replicon: pseudorevertants of mutants with a defec-tive replication origin. Proc. Natl. Acad. Sci. USA 76:6128–6131. 97. Smale, S. T., and T. Tjian. 1986. T-antigen DNA polymeraseacomplex
implicated in simian virus 40 DNA replication. Mol. Cell. Biol. 6:4077– 4087.
98. Sock, E., M. Wegner, E. A. Fortunato, and F. Grummt. 1993. Large T-antigen and sequences within the regulatory region of JC virus both con-tribute to the features of JC virus DNA. Virology 197:537–548. 99. Stahl, H., P. Droge, and R. Knippers. 1986. DNA helicase activity of SV40
large tumor antigen. EMBO J. 5:1939–1944.
100. Stewart, S. E., B. E. Eddy, A. M. Gochenoun, N. G. Bongese, and G. E. Gnubbs.1957. The induction of neoplasms with a substance released from mouse tumors by tissue culture. Virology 3:380–400.
101. Stillman, B. 1989. Initiation of eukaryotic DNA replication in vitro. Annu. Rev. Cell Biol. 5:197–245.
102. Stillman, B. 1994. Smart machines at the DNA replication fork. Cell 78: 725–728.
103. Stillman, B., R. D. Gerard, R. A. Guggenheimer, and Y. Gluzman. 1985. T antigen and template requirements for SV40 DNA replication in vitro. EMBO J. 4:2933–2939.
104. Sugasawa, K., Y. Murakami, N. Miyamoto, F. Hanaoka, and M. Ui. 1990. Assembly of nascent DNA into nucleosome structures in simian virus 40 chromosomes by HeLa cell extract. J. Virol. 64:4820–4829.
105. Takemoto, K. K., A. Furuno, K. Kato, and Y. Yoshiike. 1982. Biological and biochemical studies of African green monkey lymphotropic papovavirus. J. Virol. 72:502–509.
106. Tang, W. J., S. L. Berger, S. J. Triezenberg, and W. R. Folk. 1987. Nucle-otides in the polyomavirus enhancer that control viral transcription and DNA replication. Mol. Cell. Biol. 7:1681–1690.
107. Tang, W. J., and W. R. Folk. 1989. Constitutive expression of simian virus 40 large T antigen in monkey cells activates their capacity to support polyomavirus replication. J. Virol. 63:5478–5482.
108. Tooze, J. 1980. The molecular biology of tumor viruses. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
109. Traut, W., and E. Fanning. 1988. Sequence-specific interactions between a cellular DNA-binding protein and the simian virus 40 origin of DNA rep-lication. Mol. Cell. Biol. 8:903–911.
110. Triezenberg, S. J., and W. R. Folk. 1984. Essential nucleotides in the polyomavirus origin region. J. Virol. 51:437–444.
111. Tsurimoto, T., T. Melendy, and B. Stillman. 1990. Sequential initiation of lagging and leading strand synthesis by two different polymerase complexes at the SV40 DNA replication origin. Nature (London) 346:534–539. 112. Tsurimoto, T., and B. Stillman. 1991. Replication factors required for SV40
DNA replication in vitro. II. Switching of DNA polymerase alpha and delta during initiation of leading and lagging strand synthesis. J. Biol. Chem. 266:1961–1968.
113. van der Eb, A. J., and F. L. Graham. 1980. Assay of transforming activity of tumor virus DNA. Methods Enzymol. 65:826–839.
114. Virshup, D. M., A. A. Russo, and T. J. Kelly. 1992. Mechanism of activation of simian virus 40 DNA replication by protein phosphatase 2A. Mol. Cell. Biol. 12:4883–4895.
115. Waga, S., and B. Stillman. 1994. Anatomy of a DNA replication fork revealed by reconstitution of SV40 DNA replication in vitro. Nature (Lon-don) 369:207–212.
116. Wang, E. H., P. N. Friedman, and C. Prives. 1989. The murine p53 protein blocks replication of SV40 DNA in vitro by inhibiting the initiation func-tions of SV40 large T antigen. Cell 57:379–392.
117. Wang, E. H., and C. Prives. 1991. DNA helicase and duplex DNA fragment unwinding activities of polyoma and SV40 large T antigen display similar-ities and differences. J. Biol. Chem. 266:12668–12675.
118. Weinberg, D. H., and T. J. Kelly. 1989. Requirement for two DNA poly-merases in the replication of simian virus 40 DNA in vitro. Proc. Natl. Acad. Sci. USA 86:9742–9746.
119. Wessel, R., J. Schweizer, and H. Stahl. 1992. Simian virus 40 T-antigen DNA helicase is a hexamer which forms a binary complex during bidirectional un-winding from the viral origin of DNA replication. J. Virol. 66:804–815. 120. Wobbe, C. R., F. Dean, L. Weissbach, and J. Hurwitz. 1985. In vitro
replication of duplex circular DNA containing the simian virus 40 DNA origin site. Proc. Natl. Acad. Sci. USA 82:5710–5714.
121. Wold, M. S., J. J. Li, and T. J. Kelly. 1987. Initiation of simian virus 40 DNA replication in vitro: large-tumor-antigen- and origin-dependent unwinding of the template. Proc. Natl. Acad. Sci. USA 84:3643–3647.