JOURNAL OF VIROLOGY,
JUlY
1985, p.206-212 Vol. 55,No. 10022-538X/85/070206-07$02.00/0
CopyrightX 1985,American Societyfor Microbiology
Restricted Changes in the
Adenovirus DNA-Binding Protein That
Lead
to
Extended
Host Range or
Temperature-Sensitive Phenotypes
DOUGLAS E. BROUGH, STEPHEN A. RICE, SUSAN SELL,AND DANIEL F. KLESSIG*
Departmentof Cellular, Viral, and Molecular Biology, UniversityofUtah Schoolof Medicine, Salt Lake City, Utah 84132
Received2January1985/Accepted 22March1985
Humanadenovirusfailstomultiply efficientlyinmonkeycellsowingto ablock to late viral geneexpression. Ad2hr400 through Ad2hr4O3are asetof hostrange(hr)mutantswhichwereselected for their abilitytoreadily
grow in these cells at 37°C. The mutations responsible for this extended host range have previously been
mappedtothe5' portionofthe geneencodingthe72-kilodaltonDNA-bindingprotein (DBP). DNA sequence
analysesindicate thatallfour hrmutantscontain the samealterationatcoding triplet 130, which changes a
histidinecodon to atyrosinecodon. Theseresultsextend thoseofAnderson et al. (J. Virol.48:31-39, 1983), which suggested thatonly this change inthe DBPamino acid sequencecanexpand adenovirushost range to
monkey cells. Thehrphenotypedoesnotappear torequirephosphorylation of this tyrosineresidue, since no
phosphotyrosinewasdetected in DBPisolated fromAd2hr400-infected monkey cells. The hr mutantsAd2hr400 throughAd2hr4O3, however,arecoldsensitive for growthin monkey cells. The mutantAd2ts400, which was
derived from Ad2hr400, represents asecond class of hrmutantswhichcan growefficiently in monkey cells at
32.5°C. Thecold-resistant hr mutation of Ad2ts400 haspreviouslybeen mapped tothe 5' region ofthe DBP
gene(mapunits 63.6 through 66). DNAsequence analysisof this region shows thatthis mutantcontains the
original hr alterationatcoding triplet 130aswell as asecondalterationatcoding triplet 148, which changes an alanine codonto a valine codon. Wesuspect that thealterations atamino acids 130and 148 change the structureof the amino-terminal domain oftheDBP,allowing ittobetterinteractwithmonkey cellcomponents required for late viral geneexpression. Ad2ts400 alsocontains a temperature-sensitive mutation which has
previously been mappedtothe3' portion oftheDBPgene (mapunits61.3 through 63.6).Sequenceanalysis of thisregion indicates that the DBPcoding triplet413 has been altered. Thischangefromaserine codonto a
proline codon is thesamealterationreportedin thepreviouslysequencedDBP mutantsAd5ts125 (W.Kruijer etal., Nucleic Acids Res. 9:4439-4457, 1981) and Ad5tslO7 (W. Kruijeretal., Virology 124:425433, 1983.). Thus itappearsthat onlyaverylimited number of changes in either the 5' orthe3' portion of the DBPgene cangive risetothe hrortemperature-sensitive phenotypes, respectively.
The nonstructural adenovirus 72-kilodalton (kd)
DNA-binding protein (DBP) is a multifunctional phosphoprotein
encoded in early region 2A ofthe adenoviral genome
(co-ordinates 61.6 through 66.5) (10, 13, 30, 41, 55). DBP is
required for viral DNA replication and appears to play a
central rolein regulatingviralgeneexpression. Twoclasses
ofDBPmutants,eachaffecting verydifferentactivities and
mapping in distinctregions oftheearlyregion2Agene,have
been isolated.
One mutant class consists of
temperature-sensitive
(ts)mutants and is typified by the adenovirus type 5 (Ad5)
mutantsAd5ts125(17)andAd5tslO7(50).Studies withthese
mutantssuggest arole fortheDBP in viral DNAreplication,
earlygeneexpression, virus assembly, and cellular
transfor-mation.At the nonpermissivetemperature these ts mutants
areunabletoreplicate viralDNA(17, 29),presumably owing
tothethermolabilenatureofthesingle-stranded DNA
bind-ing activity of the tsDBP (56). Furthermore, in
Ad5tsl25-infected cellsthenormal turnoff of earlygenemRNA
expres-sion does not occur at the nonpermissive temperature (11, 12). The negativeregulation of earlygeneexpression
medi-ated by wild-type (wt) DBP appears to act at the level of
transcription forearly region4 (26, 45) and at the level of
mRNAturnoverforearlyregions 1A and 1B(8). Studies on arevertantofAd5tslO7,
R(ts107)202,
suggest an additional role for DBP in virus assembly (46). R(ts107)202 hasre-* Corresponding author.
gained a wtphenotype in human HeLacells but remains ts
for growth in human 293 cells. This mutant appears to
replicateDNAandexpresslategenesnormallyin293 cellsat
thenonpermissive temperature but failsto assemble virion
particles. Finally, DBPappears toplayarole inmodulating
the efficiency oftransformation ofratcells, since Ad5tsl25
transforms ratcells with three- tofivefold greaterefficiency
thandoes wt virus(24,57).
Thesecondclassofearlyregion2Amutantsarehostrange
(hr) mutants, which have been selected for the ability to
grow efficiently in monkey kidney cells (1, 32, 35). The wt
adenovirusdoesnotgrowwellinmonkey cells,eventhough
virus adsorption (20), earlygeneexpression (3, 20, 23), and
DNA replication(23, 27, 48)all occurnormally. The defect
appears toinvolveablocktolategeneexpression, sincethe levels of late viral proteins arediminished (9, 23, 33). This
reduction can, in most cases, be accounted for by the
decreased levels of late mRNA's(19, 33)which result from reduced transcription (J. Johnston, K. P. Anderson, and D. F. Klessig, manuscript in preparation). An exceptionis
thecapsidfiberprotein, inwhich altered mRNA splicing or
poorutilizationof the message,orboth, also contributesto the block in protein synthesis (4, 5, 34). Mutations in the
early region 2A gene allow adenovirus to overcome the
block to late gene expression and grow
productively
inmonkeycells.
Geneticmappingand DNAsequencedeterminationof the
twomutantclasses and their revertantshave
suggested
that 206on November 10, 2019 by guest
http://jvi.asm.org/
the DBPcontainsdistinctfunctionaldomains (2, 36-39, 49), sincethetsmutations andtheir second siterevertantsmapin
the 3' segment of the DBP gene, whereas the hrmutations mapin the 5' segment. Furthermore, analysisof the mutant
Ad2ts400, which contains mutations of both types, has shown that the hr DBPcanovercometheblocktolategene expression in monkey cells even when the viral DNA replication activity of the protein is nonfunctional (49). The twofunctional domains of DBPmay correspond tothetwo physical domains revealed by the sensitivity of DBP to
severalproteolyticenzymes. TheC-terminal 44-kd fragment
retains DNA binding activity (31)and is functional inan in
vitro DNA replication system (6). The smaller, heavily
phosphorylated N-terminal 26-kdfragment (31) presumably
carries the activity required for late gene expression in
monkey cells (49)and probably in humnan cellsas well.
Herewe reportthe DNAsequence analyses of Ad2hr400 through Ad2hr4O3, fourindependently derived hrmutantsof Ad2 thatgrowefficiently inrnonkey kidney cells. Our results
extendthose of Andersonetal. (2), who suggested that only
a specific change in the DBP amino acid sequence can
expand adenovirus host range to monkey cells. However, thesequence analysis of Ad2ts400 shows that anadditional
change inthe N-terminalsegmentof DBPcanallow the virus
to grow even more efficiently in monkey cells than do
Ad2hr400through Ad2hr4O3. Characterization of the altera-tion responsible for the tsphenotype of Ad2ts400, in
con-junction with similar analyses of other DBP ts mutants
(37-39),suggeststhatonlyaverylimitednumber ofchanges can impart thetsphenotype.
MATERIALS ANDMETHODS
Cells, virus, and infections. Propagation of cellsand isola-tion of virus and viral DNA were performed as previously
described(49). Viral infectionswereperformed monolayers aspreviously described (49).
Cloning of adenovirusDNA.TheEcoRIB restriction frag-ments of Ad2hr400 through Ad2hr4O3 and Ad2ts400 were
cloned in the EcoRI restriction site of pBR322. Plasmid DNAwasprepared by the method of Tanaka and Weisblum
(53). The KpnI-SstI fragment (map units [mu] 61.3 through 69.9)wasisolated from theplasmid containing the Ad2ts400
EcoRIB fragment. A1-,ugportion of this DNAwasdigested
with0.05 U of Bal 31 nuclease (Bethesda Research Labora-tories) for 1minat16°C in 100 ,ul(total volume) of the buffer suggested by Maniatisetal.(42). This DNAwasthentreated
with the Klenow fragment of DNA polymerase (42) and cloned into the XbaI site ofM13mpl9 (42) in both orienta-tions by using XbaI linkers (P-L Biochemicals, Inc.). De-letionsof the parent M13 clones were generated with
exo-nuclease III (New England BioLabs) by the method of Henikoff (28), JM105 was transformed with the various
M13mpl9plasmids by the method described in the 1982 P-L Biochemicals handbook (Manual for the M13
Cloning/Sequencing Kit, p. 9). ,B-Galactosidase-negative
cloneswere screened onLB plates supplemented with 0.03
mM
isopropyl-p-D-thiogalactopyranoside
(Sigma Chemical Co.) and 0.03% 5-bromo-4-chloro-3-indolyl-D-galactoside (Boehringer Miannheim Biochemicals) as describedprevi-ously (44). M13 clones were grown in LB medium. The protocol of Messing (44)wasusedtoisolate M13 replicative
form DNA. M13mpl9waskindlyprovided by J. Messing.
DNA sequenceanalysis. The DNAsequence for the DBP geneofAd2hr400wasdeterminedby themethod ofMaxam
andGilbert (43). Restriction enzyme digests (New England
BioLabs) of plasmid DNAwereperformed under the
condi-tions recommended by Maniatis et al. (42). The resultant DNA fragments were separated on agarose gels and
re-covered byelectroelution (42). The isolated fragments were
dephosphorylated with bacterial alkaline phosphatase and
then 5' end labeled with polynucleotide kinase and
[y-32P]ATP (Amersham Corp.) (43). After cleavage withap-propriate restriction enzymnes to yield asymmetrically la-beled fragments, the fragments were fractionated by
acrylamide gel electrophoresis and eluted (43). The isolated fragments were subjected to the chemical degradation
re-actions of Maxam and Gilbert, except that 4% formic acid
was used in place of 4% pyridine formate for the
adenine-plus-guanine reaction. Each treated samplewasanalyzed by
electrophoresis through denaturing 0.4 mM gels (40cmby 20
cm) of three acrylamide concentrations, 20%, 10or8%, and
6%. After electrophoresis, each gel was subjected to auto-radiography at -70°C with Kodak XAR film. In general, about 150 nucleotides of contiguous sequence could be determined foreach labeled fragment. Thegeneral strategy is outlined in Fig. 1A. The BgIII-EcoRI fragment (mu 63.6 through 70.7) from plasmid DNA was first digested with
HpaIIorXhoI,and after5'endlabeling, the second
restric-tion enzyme digestion was done with BstEII. The BglII
fragment (mu 60.2 through 63.6)wasfirstdigested with SstII
or BstEII and then with BstEII and KpnI or BglI,
respec-tively. The KpnI-XhoI fragment (mu 61.3 through 66) was
firstdigested with BstEII and then with HpaII.
The sequence around the coding triplet 130 was
deter-mined in the mutants Ad2hr401 through Ad2hr4O3. The BglII-EcoRI fragment (mu 63.6 through 70.7) from each plasmidwas first digested with HpaII, 5' end labeled, and
A.
SstI[ BstEfl BgIZ BstElE
A. I
Hpafl Xhol
I I
61.6mop units 63.6 66 66.5
EN.
61.6map units 63.6 66 66.5
-
--FIG. 1. Strategy usedtosequencetheDBPgene.Theregion of theadenovirusgenome (mu 61.6through 66.5)that encodestheDBP is depicted as a large open bar. Arrows indicate the direction of sequencing. (A) DNA sequence ofthe Ad2hr400 DBP was deter-minedbythechemical degradation method ofMaxamand Gilbert (43). Purified restriction enzymefragments ofDNA from pBR322 clones containing the EcoRIB fragment (mu 58.5 through70.7) of
Ad2hr400weredigested with various restrictionenzymes,labeledat both ends with polynucleotide kinase, cut with other restriction enzymes to generate asymmetrically labeled fragments, purified, and sequenced. Relevant restriction sites are shown. (B) DNA sequencing of Sangeretal.(51)wasusedtodeterminetheAd2ts400 mutant DBP DNA sequence (mu 61.6through 66). Thick arrows
represent regions sequenced by using a 15-nucleotide universal primerandtemplates from theparentM13mp19clones carrying the KpnI-SstI(mu61.3through 69.9)fragmentordeletions of it gener-ated by exonuclease III treatment (28). Thin arrows represent regions sequenced by the parent template and synthetic 20-nucleotideprimers.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.323.561.421.555.2]208 BROUGH ET AL.
then digested with BstEII to yield asymmetrically labeled
fragments forsequence determination.
The sequence of Ad2ts400 DBP was determined by the
dideoxynucleotide-primer extension method ofSangeretal.
(51), withmodifications described intheOctober 1983New
England Nuclear handbook (Step-by-Step Protocolfor
35S-Sanger Sequencing) for using [a-35S]dATP as radioactive
label. The M13 single-stranded DNA usedastemplate was
isolated by the protocol described in the 1980 Bethesda
ResearchLaboratories handbook(M13Cloning/Dideoxy Se-quencing Instruction Manual, p. 29). A 15-nucleotide uni-versal primer (New England BioLabs) was used for
sequenc-ing the full-length insert and the M13 deletion variants
obtained from exonuclease III treatment. Gaps left in the
DBPsequencebecauseof lack ofappropriateM13 deletions
were filled by using 20-nucleotide primers synthesized and
purified by preparative gel electrophoresis (Applied
Biosystems, Inc.). Regions of DNA sequence containing
putative mutationsor sequence ambiguities were subjected
tosequenceanalysis oftheopposite strand with appropriate
20-nucleotide primers. The general sequencing strategy is
illustrated inFig. 1B.
Phosphoamino acid analysis. Ad2- or Ad2hr400-infected
CV-1 cells were labeledfrom 22 to 24 h postinfection with
100 ,Ci ofH332PO4 in 0.2 ml of Dulbecco modified Eagle
medium minus phosphate supplemented with 2% dialyzed
calf serum. Proteins were extracted at 0°C by a high-salt
Nonidet P-40 extraction (15) in the presence of30 ,ug of
phenylmethylsulfonyl fluoride and 100 ,uM ammonium
metavanadate toinhibit phosphataseactivity(22). DBP was
immunoprecipitated
withapolyclonal rabbit antiserum madeagainst native DBP andelectrophoresed on a7.5% sodium
dodecyl sulfate-polyacrylamide gel. The proteins were
electroeluted from the gel bands, passed over small
Sephadex G-25 columns to desalt, and then subjected to
partial acid hydrolysis (14). The hydrolyzed samples were
lyophilized,
solubilized in water, and dried a secondtime.Thesampleswerethensolubilizedin 10 ,ulofwater
contain-ing phosphoserine, phosphothreonine, and phosphotyrosine
as markers and were spotted onto chromatography paper
(Whatman 3MM). Two-dimensional, high-voltage paper
electrophoresis was performedasdescribed by Clinton and
Huang (14), except thatelectrophoresis was atpH 1.9 for2
hin thefirst dimensionand atpH3.5for40 minin the second
dimension. As acontrol, epidermal growth factor-activated
protein kinase fromhumanepidermoid carcinoma A-431cell
membraneswas used to label anendogenous
membrane-as-sociatedproteinattyrosine
(54).
This membranepreparationwaskindly provided by S. Wiley.
RESULTS
Nucleotidesequence analysis ofAd2hr400toAd2hr4O3.The
mutants Ad2hr400 through Ad2hr4O3 were isolated after
nitrous acid mutagenesis ofAd2 and subsequent selection
for variants which could grow productively in the
estab-lishedmonkey kidneycelllineCV-1 (35).Previously, genetic
mapping ofthe mutants by markerrescue placedthe
muta-tions inthe 5' portion ofthe DBP gene located between mu
63.6 and 66 (36). To determine the precise location and nature of the mutation in each isolate, DNA sequence
analysisof the DBPgenewasperformed.The strategy used
for
determining
thesequenceoftheentire mutantDBPgene(mu 61.6 through 66.5) from Ad2hr400 is
given
in Fig. 1A.Only one nucleotide difference was observed in the mutant
DBP gene compared with the Ad2 DBP DNA sequence
publishedby Kruijer et al. (39). A thymine residue replaces
G
A+G
C+T
G-
_
C
A>C
A-A_
...4..;.
C-G-
I-.*
...c-
S
W.ikiiii
A-
-I-G-U
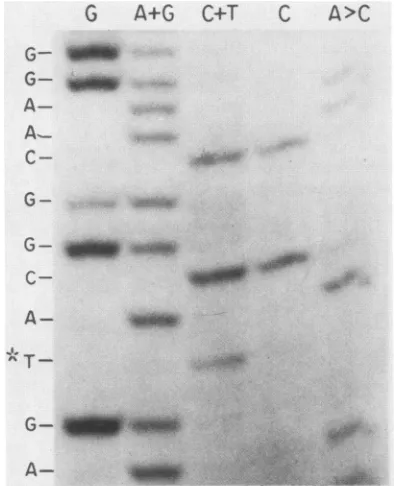
A-FIG. 2. Maxam and Gilbert (43) sequence analysis of the Ad2hr400 DBP gene.Aportion of the sequencing gel autoradiogram thatcontainsthesingle nucleotide change (*),ascompared withwt Ad2, at position 1 ofcoding triplet 130is shown. Thechangeofa
cytosineto athymineincoding triplet130changesthecodonfrom histidinetotyrosine. Thesamechangewasalso observed in the hr mutantsAd2hr401throughAd2hr4O3.
acytosineresidue at thefirstposition of coding triplet130 in
the DBP gene (Fig. 2), changing a histidine codon to a tyrosine codon.
Thenucleotide sequence in theregion foundtobe altered
in Ad2hr400wasdetermined forthethree otherhr mutants,
Ad2hr4O1through Ad2hr4O3. All three mutants werefound
tohave
precisely
thesame nucleotide change asAd2hr400. No other change was observed in theregion
examined,whichencompassed 100 to 150 nucleotides surrounding the
mutation. Thusallfourmutantsexaminedhereaswellasthe
phenotypically
similar mutants Ad5hr4O4 (38) andAd2+NDhr600
throughAd2+NDhr603
(2) haveprecisely
thesame mutation.
Therefore,
all nineindependently
isolatedadenovirus mutants selected for the
ability
to grow inmonkey cells haveacquiredthesame amino acid changein
the DBP, a substitution of
tyrosine
for histidine at aminoacid 130.
Phosphoamino acid analysis ofAd2hr400 DBP. To
deter-minewhether the hrmutantDBP
might
become phosphory-lated on tyrosine in monkeycells, we examined thephos-phorylation patterns of the DBP isolated from Ad2- and
Ad2hr400-infected CV-1 cells. Infected cells were labeled
from22to 24h postinfectionwith [32P]orthophosphate, and
the DBP was isolated by
immunoprecipitation
ofthe cellextracts with polyclonal antiserum directed
against
DBP. Theimmunoprecipitated proteins
wereeluted fromasodiumdodecyl
sulfate-polyacrylamide gel,
acidhydrolyzed,
andsubjected
tohigh-voltage
paperelectrophoresis (Fig.
3A andB).Phosphoserineand
phosphothreonine
werefoundinbothwtand hr DBPs in agreement withprevious analyses ofwt
DBP synthesized in infected HeLa cells (7). However,
phosphotyrosine
was notdetected in either themutantorwtproteins. Phosphotyrosine
wasdetected in thepositive
con-J. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.334.531.74.317.2]MUTATIONS IN THE ADENOVIRUS DNA-BINDING PROTEIN
Origein
I
.P-Tyr
-
'P-Thr
~P-Se
rFIG. 3. Two-dimensional separation ofphosphoamino acids. The positions ofthe origin, phosphotyrosine (P-Tyr), phosphothreonine (P-Thr), andphosphoserine(P-Ser) markers, and [32P]orthophosphate(Pi)areshown.H332P04-labeledDBPfrom Ad2-infected CV-1 cells(A) orAd2hr400-infected CV-1 cells (B) waspurified and subjected to acidhydrolysis. As apositive control, A-431 cell membrane fractions labeled in vitro (C)wereacidhydrolyzed inparallel.Theresultingproducts were separatedbytwo-dimensional high-voltage electrophoresis (seetextfor details).
trol consisting ofin vitrophosphate-labeled A-431cell
mem-braneswhich containepidermal growth factor receptors and
one or moreproteinswhicharephosphorylatedat atyrosine residue (Fig. 3C) (54).
Nucleotide sequence analysis ofAd2ts400. Although thehr
mutantsAd2hr400 throughAd2hr403 grow very efficiently in
monkey cellsat 37°C,theyform relatively small plaques on
CV-1 cell monolayers at 33°C, and the kinetics of plaque
formation is dramatically delayed compared with that in
similar infections of CV-1 cells bythe Ad2-simian virus 40
(SV40) hybrid virus, Ad2+ND1. This cold-sensitive
pheno-type is seenonly in monkey cells, since wt and hr viruses
formplaques with similar kineticsonhumancellmonolayers
at 330C. Ad2ts400, a nitrous acid-induced mutant of
Ad2hr400,wasselectedfor itsabilitytogrowreadilyinCV-1
cellsat33°C(49).Thecold-resistant hr mutationresponsible
for thisphenotypewasmapped bymarkerrescueanalysisto
the 5' portion of the DBP gene (mu 63.6 through 66). However, Ad2ts400 also contains a ts mutation in the 3' segmentofthe DBP gene (mu 61.6through 63.6) (49).Its ts
phenotype is similarto thatofAd5tsl25 in that viral DNA
replication andthe turndownofearly geneexpressionatlate
times areblocked atthe nonpermissive temperature.
The DNA sequence ofAd2ts400 was analyzed between
mu 61.6 and 66 by the strategy outlined in Fig. 1B. This
analysis indicated that Ad2ts400 carries three mutations in
the DBP gene as compared withthe wt DBP sequence. All
three changesreplaced cytosine with thymine.Ad2ts400still
maintains the hr alteration at coding triplet 130 which is
presentinAd2hr400. Anadditionalchange wasfound in the
5'section ofthe DBP gene,which by markerrescuemapping
is known to convey the cold-resistant hr mutation. The
alteration wasat position 2 in coding triplet 148 (Fig. 4A),
changing an alanine codon to a valine codon. The single
alterationin the 3'region,towhichthe ts mutation has been
mapped, isatthefirstposition of codingtriplet 413 (Fig. 4B),
changingaserine codontoaprolinecodon. Thismutation is
the same alteration seen in two other ts DBP mutants, Ad5ts125 (38) and Ad5ts107(37).
DISCUSSION
The nitrous acid-induced hr mutants Ad2hr400 through
Ad2hr403, all contain the same histidine-to-tyrosine change
at position 130 in DBP, as do the spontaneously generated
mutants Ad5hr4O4 (38) and Ad2+ND3hr600 through
Ad2+ND3hr603 (2). Thus all nine independently isolated
humanadenovirus mutants, selected forthe abilityto
over-comethe blockto late geneexpression and virusgrowthin monkey cells,haveacquiredthe same mutation. Byasingle
base substitution, the histidine codon can be changed to
codons forseven different amino acids. The substitution of
only atyrosineat
position
130inthe hr DBPs suggests thatthis amino acid plays an importantrole in the hrfunction.
The recent discovery ofthe tyrosine kinase activity of
severaloncogeneproducts and of various membranegrowth
factor receptors or their associated components suggests
thattyrosine
phosphorylation
maybe involvedin mediatingavariety of cellularresponses (18, 54). Sincethe hr
pheno-typedepends on thesubstitution oftyrosine for histidine at
amino acid 130 in DBP, the protein was characterized to
determine whetherphosphorylation of this tyrosine residue
G A J C G A T C
C- ._
>~~~
T- _
C- t
C-j
eA-> ^.__
A. B.
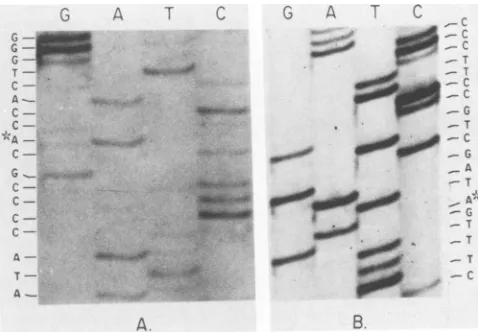
FIG. 4. Dideoxynucleotide sequence analysis of the mutant Ad2ts400DBPgene.Thefigureshowsportionsof thesequencing gel autoradiograms ofthe viral DNA noncoding strand which contain thetwomutational sitesinAd2ts400notfound inparentalAd2hr400.
(A) Alteration in coding triplet 148 which is responsible for the cold-resistant hr phenotype. The altered nucleotide (*, G-to-A; C-to-T on the coding strand) changes codon 148 from analanine codon to a valine codon. (B)Mutation in codingtriplet 413 which impartsthe tsphenotype. The altered nucleotide(*,G-to-A;C-to-T
onthecoding strand)changes codon 413 fromaprolinecodontoa serinecodon.
A.
rigi
.P-Tyr
fP-Th
rP-Ser
Pi
209 VOL.55, 1985
SW
I,Pi
11-1
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.75.552.75.218.2] [image:4.612.322.561.459.625.2]210 BROUGH ET AL.
was importantfor the mutant phenotype. Althoughboth hr and wt DBP contained similar amounts of both
phos-phoserine and phosphothreonine, phosphotyrosine was not
detected in either protein. This suggests that tyrosine phos-phorylation is not involved in the hr phenotype. We cannot, however, exclude the possibility that if onlyasubsetof DBP
molecules are phosphorylated on tyrosine, the level ofthis
phosphorylationmight be belowthesensitivity ofourassay,
which could readily detect phosphotyrosine in a control experiment.
The hrmutants Ad2hr400through Ad2hr4O3, which were
selected to grow well inmonkey cellsat37°C, arerelatively
inefficient at late gene expression and growth in
monkey
cells but not in human cells at 33°C. The mutant virus
Ad2ts400 was selected on the basis of its cold-resistant hr
phenotype. Themutationresponsible forthisphenotypehas
beenmapped to the 5'regionofthe DBP gene. Weknowthat
thetsmutationlocatedin the3' segmentofthe DBP geneis
not involved in the cold-resistant hr phenotype, since we
have obtained a virus, Ad2hr4O5, which contains a wt 3'
portion of the DBP gene but retains the cold-resistant hr
phenotype. Interestingly, Ad2hr4O5 forms larger plaques
than do Ad2hr400 through Ad2hr4O3 at37and at39.5°C(S.
Rice and D. Klessig, unpublished results). DNA sequence
analysis oftheAd2ts4005' segmentindicatedthatin addition
tothealteration ofthe histidinecodontothe tyrosinecodon
at position 130, which is found in all of the hr mutants,
Ad2ts400 contained an alteration at the coding triplet 148.
Since an hr mutant containing only achange at amino acid
148 has neverbeen isolated, it seemslikely thatthischange alone is insufficienttoimpartthe hrphenotype. Morelikely,
the alterationatcoding triplet
148
works inconcertwiththechange atposition130 tofine tune the structure ofthe DBP
so that it can interact more efficiently with monkey cell
factors required forexpression of viral late genes.
The other alteration inthe DBP geneof Ad2ts400 is in the
3'portion ofthe geneandisresponsible forthetsphenotype.
It was previously reported that Ad2ts400 has a phenotype
similar to the mutants Ad5tsl25 and Ad5tslO7 at the
nonpermissive temperature (49). In fact, Ad2ts400 has the
samenucleotide changeas these mutants. The alteration in
coding triplet 413 of Ad2ts400, Ad5tsl25, and Ad5tslO7
changesaproline codonto aserinecodon. Itseemsurprising
that three of the four ts DBP mutants which have been
sequenced containthe samenucleotide alteration. Since we
have used themutants Ad5tsl25 andAdStslO7inour labora-tory, we considered the unlikely possibility that Ad2ts400
arose by a recombination event
involving
Ad5tsl25 orAd5ts1O7. We can exclude this
possibility,
since Ad2ts400containsAd2
serotype-specific
DNA sequencesthroughout
the gene,includingtwoAd2-specific differences(relative
to thewt AdS sequence [38]) locatedwithin 25 nucleotidesof
the ts mutation.
The restriction in the number of mutational sites seen in the N-terminalportion of the DBP is perhaps in retrospect reasonable, given the possible constraints in
protein
struc-ture imposed by the strong selection forgrowth in monkey
cells. In contrast, the conserved changesin the C-terminal
portionareverysurprising. Threeoutof fivetsmutations in
this domain have the same alteration at
position
413. Theexceptions areAd2+NDlts23, which contains an alteration
at coding triplet 282 (39) and Ad2tslll,which is
thought
tobe located between coding triplets 200 and 300
(16,
52).
Perhaps onlyverylimitedchangesintheC-terminal
domain
ofDBPwill allow this
protein
tofunctionat32.5°C
butnot at39.5°C. However, this may not be the entire
explanation,
MUTANTS:hr(IO)
Ad2hr400-403 (a.a. 130) Ad2+ND3hr6OO-6O3(a.a. 130)
Ad5hr4O4 (a.a.130) Ad2ts400 (a.a.130,a. o.148) FUNCTIONS: Lotegeneexpression
ts(4)
Ad2ts400
Ad5ts 125 Ad5ts107 Ad2+NDlts23
(a.a.413) (a.a. 413) (a.a. 413) (a.a.282)
Earlygeneexpression
DNAreplication DNA binding
Virus assembly
Morphologicoltransformation FIG. 5. Schematicrepresentationofthe separatefunctional do-mainsof the adenovirusDBP.TheN-terminal 26-kdfragmentof the
proteincontains the hr mutation which relieves the blockto viral lategeneexpressioninmonkeykidneycells. All of10 independently
derived hrmutantscontain thesamealterationatcodon130,which results inthe replacement ofhistidine by tyrosine. Ad2ts400
con-tainsanadditional hrmutation, locatedatamino acid148,changing
alaninetovaline. This mutationfurther enhances thegrowthofthe
virus inmonkeycells,particularlyatlow
temnperatures.
Thefourts mutants that affect earlygeneexpression, DNAreplication, DNA binding, virus assembly, and morphological transformation arelocated inthe44-kdC-terminal domain. Three of these fourmutants have an identical alteration at codon 413, which results in the substitution of serine forproline. Thefourth ts mutationchanges
amino acid282from leucinetophenylalanineinAd2+NDlts23.The two domains may be separated by a hydrophilic hinge region
represented by theconnectingline.
sinceAd2ts400wasisolated afterselection for cold-resistant
hr mutants.
Surprisingly,
one out ofthe two mutantsob-tained, namely Ad2ts400,
simultaneously
hadacquired
a tsalteration at amino acid 413 in addition to the alteration at
amino acid 148 which
imparts
the cold-resistant hrpheno-type. Thus the
repeated
isolation of mutations atcoding
triplet
413might
suggestthehigh
susceptibility
of thisregion
of the gene to
mutagenesis.
The sequence
analysis
ofAd2hr400through Ad2hr4O3
and Ad2ts400 further supports the model that DBP contains at least two functional domains. The hr mutations at aminoacids130and148 and thetsmutationsat aminoacids413 and
282 are
separated
by
anextremely
hydrophilic
region
(cen-tered around amino acid
160)
that could act as apossible
hinge
between the twodomains(Fig.
5).
Thelarger
domaincontains the sites of the ts
alterations,
which affectearly
gene
expression,
DNAreplication,
viralassembly,
and theefficiency
ofmorphological
transformation. The smallerdo-maincontainsthe hr
alterations,
whichallownormalexpres-sion oflate genes in
monkey
cells. Coinfection ofmonkey
cells with wt adenovirus
plus
SV40also allows normal late geneexpression (47),
as does infection withAd2-SV40
hybrid
viruses thatcontainonly
theSV40sequencesencod-ingtheC-terminal
segment
ofTantigen (21, 25,
40).
ThustheC-terminal segment ofSV40 T
antigen
and the N-terminaldomainof adenovirus DBP appeartosharesimilar functions.
ACKNOWLEDGMENTS
Thisworkwas supported by PublicHealth Servicegrantno. Al
17315fromthe National InstitutesofHealth andaSearle
Scholar-ship to D.F.K. from the Chicago
Community
Trust. D.F.K. is supportedbyaFacultyResearchAward(no.270)fromtheAmeri-can Cancer Society, and S.A.R. was
supported
by
apredoctoral
traininggrant(no.T32GM07464-08)fromtheNationalInstitutes of Health.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.318.549.60.209.2]LITERATURE CITED
1. Anderson, C. W. 1981. Spontaneousmutantsof the adenovirus-simian virus 40 hybrid, Ad2+ND3, that grows efficiently in monkey cells. Virology 111:263-269.
2. Anderson, C. W., M. M. Hardy, J. J. Dunn, and D. F. Klessig. 1983. Independent, spontaneous mutants of adenovirus type
2-simian virus 40 hybrid Ad2+ND3 that grow efficiently in
monkey cellspossessidentical mutations in the adenovirustype
2 DNAbinding proteingene. J. Virol. 48:31-39.
3. Anderson, K. P., and D. F. Klessig. 1982. Synthesis of human adenovirus early RNA species is similar in productive and abortive infections of monkey and human cells. J. Virol. 42:748-754.
4. Anderson, K. P., and D. F. Klessig. 1983. Posttranscriptional block to synthesis of a human adenovirus capsid protein in abortively infected monkey cells. J. Mol.Appl.Genet. 2:31-43. 5. Anderson, K. P., and D. F. Klessig. 1984. Altered mRNA splicinginmonkey cells abortively infectedwith human adeno-virusmaybe responsible for inefficient synthesis of the virion
fiberpolypeptide. Proc. Natl. Acad. Sci. U.S.A. 81:4023-4027. 6. Ariga, H., H. Klein, A. Levine, and M. Horwitz. 1980. A
cleavage product of the adenoviral DNA-binding protein is
active in DNAreplication invitro. Virology 101:307-310. 7. Axelrod, N. 1978. Phosphoproteins of adenovirus 2. Virology
87:366-383.
8. Babich, A., and J. R. Nevins. 1981. The stability of early
adenovirus mRNA is controlledbythe viral 72 kdDNA-binding protein. Cell26:371-379.
9. Baum, S. G., M. S. Horwitz, and J.V.Maizel, Jr.1972. Studies of the mechanism of enhancement of human adenovirus infec-tion inmonkeycellsby simian virus 40. J. Virol. 10:211-219.
10. Berk, A. J., and P. A. Sharp. 1978. Structure of adenovirus 2
early mRNAs. Cell 14:695-711.
11. Carter, T. H., and R. A. Blanton. 1978. Autoregulation of adenovirus type5 earlygeneexpression. II. Effect of
tempera-ture-sensitive early mutations on virus RNAaccumulation. J. Virol. 28:450-456.
12. Carter, T. H., and R. A. Blanton. 1978. Possible role ofthe
72,000-dalton DNA-binding proteininregulationof adenovirus typeSearlygeneexpression. J. Virol. 25:664-674.
13. Chow, L. T., T. R. Broker, and J. B. Lewis. 1979. Complex splicingpatternsof RNAs fromtheearly regionsof adenovirus-2. J. Mol. Biol. 134:265-303.
14. Clinton, G. M., and A. S. Huang. 1981. Distribution of phosphoserine, phosphothreonine and phosphotyrosine in
pro-teinsofvesicular stomatitis virus. Virology 108:510-514. 15. Crawford, L., K. Leppard, D. Lane, and E. Harlow. 1982.
Cellular proteins reactive with monoclonal antibodies directed
againstsimian virus 40T-antigen. J. Virol. 42:612-620. 16. D'Halluin, J. C., C. Cousin, C. Niel, and P. Boulanger. 1984.
Characterizationofanearly temperature-sensitiveandcytocidal
double mutant of adenovirus type 2. J. Gen. Virol. 65:1305-1317.
17. Ensinger, M. J., and H. S. Ginsberg. 1972. Selection and preliminary characterization oftemperature-sensitive mutants
of type5adenovirus. J. Virol. 10:328-339.
18. Erikson,R.L.,and A. F. Purchio.1982.Aviansarcomaviruses,
protein kinases, and celltransformation, p. 43-57. In G. Klein (ed.), Advances in viral oncology, vol. 1. Raven Press, New York.
19. Farber, M.S., and S. G. Baum. 1978. Transcription of adeno-virusRNA inpermissive and nonpermissive infections. J. Virol.
27:136-148.
20. Feldman,L. A.,J.S.Butel, and F. Rapp. 1966. Interaction ofa
simianpapovavirusand adenoviruses. I. Induction of
adenovi-rus tumor antigen duringabortive infection of simian cells. J.
Bacteriol. 91:813-818.
21. Fey, G., J. B. Lewis, T. Grodzicker, and A. Bothwell. 1979.
Characterization ofafusedprotein specified by the adenovirus
2-simianvirus 40hybrid Ad2+ND1 dp2. J. Virol. 30:201-217.
22. Frackelton, A. R., Jr., A. H. Ross, and H. N. Eisen. 1983.
Characterization anduseof monoclonal antibodies forisolation of phosphotyrosyl proteins from retrovirus-transformed cells and growth factor-stimulated cells. Mol. Cell. Biol. 3:1343-1352.
23. Friedman, M. P., M. J. Lyons, and H. S. Ginsberg. 1970. Biochemical consequences of type 2 adenovirus and simian virus 40 doubleinfections of African green monkey kidney cells. J. Virol. 5:586-597.
24. Ginsberg, H.S., J. J. Ensinger, R. S. Kauffman, A.J. Mayer, andU. Lundholm. 1974.Celltransformation:astudy of regula-tion with types 5 and 12 adenovirus temperature-sensitive mutants.ColdSpringHarborSymp. Quant. Biol.39:419-426. 25. Grodzicker, T., C. Anderson, P. A. Sharp, and J. Sambrook.
1974.Conditional lethalmutantsof adenovirus 2-simian virus40 hybrids. I. Host range mutants of Ad2+ND1. J. Virol. 13: 1237-1244.
26. Handa,H., R. E.Kingston,and P. A.Sharp.1983. Inhibition of adenovirusearly region IV transcriptionin vitro byapurified DNAbinding protein. Nature(London) 302:545-547.
27. Hashimoto, K., K. Nakajima, K. Oda, and H. Shimojo. 1973. Complementation oftranslational defect for growth ofhuman adenovirustype2 in simian cellsbyasimian virus40-induced factor.J. Mol. Biol. 81:207-223.
28. Henikoff, S. 1984. One-way exonucleolytic digestionto target detectionbreakpointforDNAsequencing. Gene28:351-359. 29. Horwitz, M. S. 1978. Temperature-sensitive replication of
HSts125 adenovirus DNA in vitro. Proc. Natl. Acad. Sci. U.S.A. 75:4291-4295.
30. Kitchingman, G. R., S. P. Lai, and H. Westphal. 1977. Loop structuresinhybrids of earlyRNAand theseparatedstrands of adenovirusDNA. Proc. Natl. Acad.Sci. U.S.A.74:4392-4395. 31. Klein, H.,W.Maltzman,and A.J.Levine.1979. Structure-func-tion relationships of the adenovirus DNA-binding protein. J. Biol. Chem. 254:11051-11060.
32. Klessig,D. F. 1977.Two adenovirus mRNAs havea common5' terminal leadersequence encodedatleast 10kbupstreamfrom their main coding regions. Cell12:9-21.
33. Klessig, D.F., and C. W. Anderson. 1975. Blockto
multiplica-tion of adenovirus serotype 2 in monkey cells. J. Virol. 16:1650-1688.
34. Klessig, D. F., and L. T. Chow. 1980. Incomplete splicingand deficient accumulation of the fibermessenger RNAin monkey
cells infected by human adenovirus type 2. J. Mol. Biol. 139:221-242.
35. Klessig, D. F., and T. Grodzicker. 1979. Mutations that allow humanAd2andAdStoexpresslategenes inmonkey cellsmap in the viralgeneencoding the 72K DNAbinding protein. Cell 17:957-966.
36. Klessig,D. F., and M. P. Quinlan. 1982. Genetic evidence for separatefunctional domainsonthehumanadenovirusspecified, 72kd, DNAbinding protein. J. Mol.Appl. Genet. 1:263-272. 37. Kruijer, W., J. C. Nicolas, F. M. A. van Schaik, and J. S.
Sussenbach. 1983. Structure and function of DNA binding proteins from revertants of adenovirus type 5 mutants with a temperature-sensitive DNAreplication. Virology 124:425-433. 38. Kruijer, W.,F. M. A. vanSchaik,andJ. S. Sussenbach. 1981.
Structure and organization of the gene coding for the DNA binding protein of adenovirus type 5. Nucleic Acids Res. 9:4439-4457.
39. Kruijer, W.,F. M. A. vanSchaik,andJ. S. Sussenbach. 1982. Nucleotide sequence ofthe gene encoding adenovirus type 2 DNAbinding protein. Nucleic AcidsRes. 10:4493-4500. 40. Lewis,A.M., Jr., J. J. Levin,W. H.Weise,C. S.Crumpacker,
and P. H.Henry. 1969. A nondefective(competent) adenovirus-SV40 hybrid isolated from the Ad2-SV40 hybrid population. Proc. Natl. Acad. Sci. U.S.A.63:1128-1135.
41. Lewis, J. B., C.W.Anderson, and J. F. Atkins. 1977. Further mapping of late adenovirus genes by cell-free translation of RNA selectedby hybridizationtospecificDNAfragments.Cell 12:37-44.
42. Maniatis, T.,E. F.Fritsch, andJ. Sambrook. 1982. Molecular cloning, alaboratory manual. ColdSpringHarborLaboratory, Cold SpringHarbor,N.Y.
on November 10, 2019 by guest
http://jvi.asm.org/
212 BROUGH ET AL.
43. Maxam, A. M., and W. Gilbert. 1980. Sequencingend labeled DNAwithbase-specific chemical cleavages. Methods Enzymol. 65:499-560.
44. Messing, J. 1983. New M13 vectors for cloning. Methods Enzymol. 101:20-78.
45. Nevins, J. R., and J. J. Jensen-Winkler. 1980. Regulation of earlyadenovirus transcription:aprotein product of early region 2specifically represses region 4 transcription. Proc. Natl. Acad. Sci. U.S.A. 77:1893-1897.
46. Nicolas, J. C., P. Sarnow, M. Girard, and A. J.Levine. 1983. Hostrange temperature conditional mutants intheadenovirus DNAbinding protein are defective in the assembly of infectious virus. Virology 126:228-239.
47. Rabson,A.S., G. T.O'Connor, I. K. Berezesky, andF.J.Paul. 1964. Enhancement of adenovirus growth in African green monkey kidney cell cultures by SV40. Proc. Soc. Exp. Biol. Med. 116:187-190.
48. Reich,P.R., S. G. Baum, J. A. Rose,W.0. Rowe, and S. M. Weissman. 1966. Nucleic acid homology studies of adenovirus type 7 - SV40 interactions. Proc. Natl. Acad. Sci. U.S.A. 55:336-341.
49. Rice,S.A., andD. F.Klessig. 1984. Thefunction(s) provided by the adenovirus-specified, DNA-binding protein required for viral late gene expression is independent of the role of the protein inviral DNA replication. J. Virol. 49:35-49.
50. Rubenstein, F. E., and H. S. Ginsberg. 1974. Transformation
characteristics of temperature-sensitive mutants of type 12 adenovirus. Intervirology 3:170-174.
51. Sanger,F.,S.Nicklen,and A.R.Coulsen. 1977. DNA
sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. U.S.A.74:5463-5467.
52. Stillman,B. W.,E. White,andT. Grodzicker. 1984. Indepen-dentmutationsinAd2tslll causedegradation of cellular DNA anddefective viral DNAreplication. J. Virol.50:598-605. 53. Tanaka,T., and B.Weisblum. 1975. Construction ofacolicin
E1-R factorcomposite plasmid in vitro:meansforamplification
ofdeoxyribonucleic acid. J. Bacteriol. 121:354-362.
54. Ushiro, H., and S. Cohen. 1980. Identification of phospho-tyrosine as a product of epidermal growth factor-activated protein kinase in A-431 cell membranes. J. Biol. Chem. 255:8363-8365.
55. Van der Vliet, P. C., and A. J. Levine. 1973. DNA binding
proteins specificforcells infectedbyadenovirus. Nature
(Lon-don)NewBiol.246:170-173.
56. Van derVliet, P. C., A. J. Levine, M.J. Ensinger, and H. S. Ginsberg. 1975. Thermolabile DNA binding proteins from cells infected with a temperature-sensitive mutant of adenovirus defective in viral DNAsynthesis. J. Virol. 15:348-354. 57. Williams, J. F.,C.S. H.Young, andP.E. Austin.1974.Genetic
analysis of human adenovirus type 5 in permissive and nonpermissive cells. Cold Spring Harbor Symp. Quant. Biol. 39:427-437.
J. VIROL.