Highly Pathological Influenza A Virus Infection Is Associated with
Augmented Expression of PD-1 by Functionally Compromised
Virus-Specific CD8
ⴙT Cells
John A. Rutigliano, Shalini Sharma, Melissa Y. Morris, Thomas H. Oguin III, Jennifer L. McClaren, Peter C. Doherty, Paul G. Thomas
Department of Immunology, St. Jude Children’s Research Hospital, Memphis, Tennessee, USA
ABSTRACT
One question that continues to challenge influenza A research is why some strains of virus are so devastating compared to their more mild counterparts. We approached this question from an immunological perspective, investigating the CD8ⴙT cell response in a mouse model system comparing high- and low-pathological influenza virus infections. Our findings reveal that the early (day 0 to 5) viral titer was not the determining factor in the outcome of disease. Instead, increased numbers of antigen-specific CD8ⴙT cells and elevated effector function on a per-cell basis were found in the low-pathological infection and correlated with reduced illness and later-time-point (day 6 to 10) viral titer. High-pathological infection was associated with increased PD-1 expression on influenza virus-spe-cific CD8ⴙT cells, and blockade of PD-L1in vivoled to reduced virus titers and increased CD8ⴙT cell numbers in high- but not low-pathological infection, though T cell functionality was not restored. These data show that high-low-pathological acute influenza virus infection is associated with a dysregulated CD8ⴙT cell response, which is likely caused by the more highly inflamed airway microenvi-ronment during the early days of infection. Therapeutic approaches specifically aimed at modulating innate airway inflammation may therefore promote efficient CD8ⴙT cell activity.
IMPORTANCE
We show that during a severe influenza virus infection, one type of immune cell, the CD8 T cell, is less abundant and less func-tional than in a more mild infection. This dysregulated T cell phenotype correlates with a lower rate of virus clearance in the se-vere infection and is partially regulated by the expression of a suppressive coreceptor called PD-1. Treatment with an antibody that blocks PD-1 improves T cell functionality and increases virus clearance.
C
learance of virus-infected cells is the raison d’etre of the CD8⫹ T cell. However, CD8⫹T cells can also contribute significant immunopathology (1), which may partially account for the wide range of disease severity seen during influenza virus infection. Yet in the absence of CD8⫹T cells, virus clearance is delayed, which may lead to exacerbated morbidity and mortality (2,3). Disparate pathologies can be observed when comparing multiple strains of influenza virus and even when comparing the same strain in two different individuals. Identifying the mechanism of CD8⫹T cell-induced immunopathology therefore remains an important area of focus (4).Epithelial cells at the site of influenza virus infection in the airway microenvironment initiate an early and rapid cascade of cytokine production, followed by local inflammation and ensuing responses (5). Nevertheless, some strains are able to subvert these early responses, which may result in prolonged or increased in-flammation and excessive production of cytokines and chemo-kines. This is exemplified by the hypercytokinemia seen in highly pathogenic H5N1 avian influenza infection (6–8). Although highly pathogenic H5N1 avian influenza viruses have been unable to efficiently cross over to the human population thus far, mortal-ity continues to hover around 60% (http://www.who.int/csr /disease/avian_influenza/en/). In addition, the H1N1 swine-ori-gin influenza virus (SOIV) pandemic of 2009 has also been marked by elevated proinflammatory cytokine production in pa-tients who suffered severe infections (9).
It has been established that initiation of the adaptive response is heavily influenced by innate immunity (10). CD8⫹T cell acti-vation in its most general terms requires dendritic cell (DC)
liga-tion and assistance from T helper cells. Other reports have shown that NK cell activity during the immune response can affect CD8⫹ T cells (11) and that even the specific type of target cell can regulate CD8⫹T cell effector function (12). With time, as demonstrated in chronic infections, CD8⫹T cells can become exhausted and lose their functionality (13). This exhaustion is defined by upregula-tion of PD-1 on the cell surface (14). More recently, PD-1 upregu-lation during acute infection has also been described (15). Block-ade of PD-L1, one of two known ligands for PD-1, restores CD8⫹ T cell functionality and wholly improves the response to chronic lymphocytic choriomeningitis virus (LCMV) infection. Little work has investigated the role of PD-1 during influenza virus in-fection. Increased expression of PD-1 on DbNP
366-specific CD8⫹ T cells was briefly shown during acute infection, and cells express-ing PD-1 exhibited impaired gamma interferon (IFN-␥) produc-tion and degranulaproduc-tion at day 14, but not at day 7, postinfecproduc-tion (15). This expression was reversed by anti-PD-L1 treatment. An-other study used a self-antigen system to show that PD-1 was expressed early on hemagglutinin-specific CD8⫹ T cells, and
Received30 September 2013Accepted12 November 2013 Published ahead of print20 November 2013
Address correspondence to Paul G. Thomas, [email protected]. J.A.R. and S.S. contributed equally to this article.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.02851-13
on November 7, 2019 by guest
http://jvi.asm.org/
blockade of PD-L1 at the time of self-antigen encounter led to development of functional CD8⫹T cells (16). However, that work used strains ofListeria monocytogenesthat expressed an epitope from the influenza virus hemagglutinin protein rather than actual influenza virus infection.
Some influenza A viruses are much more virulent than others, and apart from possible early effects of the cytokine storm, we do not fully understand why. To answer this question, we have com-pared the immune responses of mice after high- and low-patho-logical (high- and low-path) influenza virus infections. The airway microenvironment was more highly inflamed early during high-path infection. However, low-high-path infection induced a robust CD8⫹ T cell response characterized by significantly increased numbers of influenza virus-specific CD8⫹T cells that exhibited greater functional activity and were more polyfunctional than those from mice with high-path infection. The attenuated CD8⫹T cell response in high-path infection was associated with increased PD-1 expression, andin vivoblockade of PD-L1 improved CD8⫹ T cell numbers and decreased viral titers but failed to improve overall illness or rescue CD8⫹T cell function. These data suggest that a dysregulated airway microenvironment may abrogate CD8⫹T cell function and thus contribute to the phenotype of high-path influenza virus infection.
MATERIALS AND METHODS
Mice.All mice were cared for under specific-pathogen-free conditions in an approved animal facility at St. Jude Children’s Research Hospital (SJCRH). All animal work was reviewed and approved by the appropriate institutional animal care and use committee at SJCRH (protocol number 098) following guidelines established by the Institute of Laboratory Ani-mal Resources and approved by the Governing Board of the U.S. National Research Council. Severe morbidity in infected animals was assessed daily using a body index score and humane euthanasia applied according to the standards of our approved protocol (greater than 30% weight loss com-bined with a threshold body index score indicating moribundity). Patho-gen-free wild-type (WT) C57BL/6 female mice, B6.129S7-IFNgtm1Ts/J
mice (IFN-␥deficient [IFN-␥⫺/⫺]), B6;129S-Tnftm1Gkl/J mice (tumor
ne-crosis factor [TNF] deficient [TNF⫺/⫺]), and B6.129S2-Il6tm1Kopf/J mice
(interleukin-6 deficient [IL-6⫺/⫺]) were purchased from Jackson Labora-tories (Bar Harbor, ME). Experiments were performed with age-matched groups.
Viruses and infections.All infections were performed with either A/Puerto Rico/8/34 (PR8) or A/HKx31 (x31). The x31 virus contains the six internal genes of PR8 but expresses H3N2 surface proteins, whereas PR8 expresses H1N1 surface proteins (17,18). In one set of experiments, the high-path⌬Vn1203 virus was used as a control for high-path PR8 infection.⌬Vn1203 is a recombinant virus with the six internal PR8 genes and surface H5N1 proteins from A/Vietnam/1203/ 04, constructed using reverse genetics (19,20). The polybasic cleavage site in the H5 was modified to restrict its cleavage to trypsin-like pro-teases. The N1 protein was unchanged. The⌬Vn1203 virus was a gift from Richard J. Webby of SJCRH. Antibody against PD-1L (clone 10F.9G2; BioXCell) was administered intraperitoneally (i.p.) in 200-g doses every other day, beginning the day before infection. Antibody against keyhole limpet hemocyanin (KLH) (clone LTF-2; BioXCell) was used as an isotype control. Antibody against granulo-cyte colony-stimulating factor (G-CSF) (clone 67604; R & D Systems) was administered i.p. in 200-g doses at days 1, 3, and 5 after infection. Mice were chemically restrained with 2,2,2-tribromoethanol (avertin) prior to intranasal (i.n.) challenge with 10450% egg infective doses
(EID50) of either PR8 or x31 in one set of experiments. In other
exper-iments, mice were infected with 103EID
50of PR8 or 106EID50of x31.
Illness was monitored by daily weighing after virus challenge.
Plaque assays.Mice were sacrificed and lung tissue removed after collection of bronchoalveolar lavage (BAL) fluid. Lungs were then stored at⫺80°C prior to the assay. Monolayers of Madin-Darbin canine kidney (MDCK) cells in six-well plates were infected with serial dilutions of 1 ml of lung supernatants after homogenization. The infected monolayers were incubated for 1 h at 37°C and then washed with phosphate-buffered saline (PBS). After washing, the cells were treated with 0.8% agarose in minimal essential medium (MEM) containing 1 mg/ml trypsin. The infected cells were incubated at 37°C for 72 h. Plates were then transferred to 4°C for at least 10 min, the agar was gently removed, and the monolayers were stained with crystal violet to visualize influenza virus plaques.
Synthetic peptides and tetramers.All peptides were synthesized by the Hartwell Center at SJCRH. Peptides corresponding to influenza CD8⫹T cell epitopes NP366-374(ASNENMETM) (NP366), PA224-233
(SSLENFRAYV) (PA224), and PB1-F262-70(SSLENFRAYV) (F262) are
all H-2Dbrestricted. PB1
703-711(SSYRRPVGI) (PB1703) is H-2Kb
re-stricted. The OVA257-264peptide (SIINFEKL) (OVA257) is H-2Kb
re-stricted and was used as a negative control in all experiments. Class I major histocompatibility complex (MHC) tetramers were constructed by combining H-2Dbor H-2Kbwith the aforementioned
immuno-genic peptides. Tetramer staining was done as described previously (21).
Flow cytometry and intracellular cytokine staining (ICS).Mice were sacrificed and BAL fluid, mesenteric lymph nodes (MLN), and spleens har-vested after challenge. MLN and spleens were manually disrupted by grinding organ tissue between the frosted ends of two sterile glass microscope slides in sterile PBS containing 2% fetal bovine serum (FBS) (2% PBS). Red cell lysis was performed for spleen cells. Cells were then stained with allophycocyanin (APC)- or phycoerythrin (PE)-conjugated influenza virus tetramers for 1 h at room temperature. After washing, cells were stained with fluorescently la-beled antibodies against CD8␣(clone 53-6.7), CD4 (clone GK1.5), CD27 (clone LG.3A10), the activated isoform of CD43 (clone 1B11), PD-1 (clone J43), PD-1L (clone 10F.9G2), Tim-3 (clone 215008; R&D Systems), PD-L2 (clone TY25), CD45 (clone 104), MHC class II (clone 2G9), CTLA-4 (clone UC10-4B9), and unlabeled anti-CD16/CD32 (clone 2.4G2) to block nonspe-cific Fc receptor-mediated binding. CD8⫹T cells were distinguished from antigen-presenting cell populations by using antibodies against CD45R/B220 (clone RA3-6B2), F4/80 (clone CI:A3-1), CD11b (clone M1/70), and CD11c (clone N418).
For ICS, lymphocytes were cultured in 96-well round bottom plates for 5 h at 37°C in 200l RPMI containing 10% FCS (10% RPMI). To promote antiviral cytokine production, the cells were also supplemented with 1M NP366and PA224peptides, brefeldin A, and CD28 anti-body for costimulation. Positive controls were stimulated with 1g/ml of phorbol myristate acetate (PMA) and ionomycin instead of influenza virus peptides. Afterin vitrostimulation, the cells were washed, fixed, and perme-abilized according to the manufacturer’s protocol (BD PharMingen Cytofix/ Cytoperm kit). Cells were then stained with the antibodies listed above and TNF (clone MP6-XT22), IFN-␥(clone XMG1.2), and CD107a/b (clones 1D4B and ABL-93, respectively). After staining, cells were resus-pended in 2% PBS plus azide and detected using an LSRII flow cytometer (BD Biosciences). Data were analyzed using FlowJo software (Tree Star, San Carlos, CA). Antibodies were purchased from BioLegend, BD PharMingen, and Life Technologies.
Surface staining was performed as described previously. For bro-modeoxyuridine (BrdU) staining, the BrdU kit from BD PharMingen was used according to the manufacturer’s instructions. Briefly, x31 and PR8 (104EID
50)-infected mice received i.p. injection of BrdU (1
mg/mouse) at 14 to 15 h before termination of mice. BAL fluid, drain-ing MLN, and spleen cells were stained first with PA and NP tetramers and then with anti-CD8 and subsequently were labeled with anti-BrdU monoclonal antibody (MAb). Samples were acquired on LSRII flow cytometer (BD Biosciences). Data were analyzed using FlowJo soft-ware (Tree Star, San Carlos, CA).
on November 7, 2019 by guest
http://jvi.asm.org/
Detection of antiviral cytokine production.Cytokine and chemo-kine production in the BAL fluid supernatants of infected mice was quan-tified using Milliplex MAP kits in 96-well assays from Millipore (Mouse Panel I). Plates were analyzed on a Bio-Rad Bioplex HTF system using Luminex xMAP technology to measure the levels of 32 different cytokines and chemokines included in the panel. Additional cytokine production was measured in commercially available enzyme-linked immunosorbent assay (ELISA) kits (R&D Systems).
In vitroactivation.Spleens were harvested from naive WT C57BL/6 mice, and CD8⫹T cells were isolated by magnetic bead separation (Milte-nyi Biotech). CD8⫹T cells were cultured in 96-well plates that were pre-coated with 4g/ml anti-mouse CD3 (clone 17A2, BioLegend). All cells were incubated for 72 h at 37°C in 10% RPMI medium supplemented with 1g/ml anti-mouse CD28 (BD PharMingen) and 1g/ml recombinant human IL-2 (rhIL-2) (R&D Systems). Cells were coincubated in triplicate wells with 5 ng/ml recombinant murine IL-6, recombinant murine G-CSF (both from Peprotech), or both cytokines together. Control wells had no cytokine added. After 72 h, cells were washed and incubated with brefeldin A for an additional 4 h prior to treatment for ICS, as described above. Activation was confirmed by flow cytometry after cells were stained for surface CD27 and CD43 expression and also intracellular production of IFN-␥.
Statistical analysis.The Studentttest or the Mann-Whitney test for unpaired, nonparametric analysis was used to assess statistical signifi-cance in all experiments. Cox proportional hazards survival regression was used to assess statistical significance in survival experiments.Pvalues of⬍0.05 were considered statistically significant.
RESULTS
Clinical profiles of high- and low-path influenza A virus infec-tions. PR8 is a mouse-adapted H1N1 influenza virus that is known to cause severe infection in mice. In contrast, x31 is an H3N2 influenza virus that causes mild to moderate illness in mice, despite containing the six internal genes of PR8. Mice were infected with 104EID
50of PR8 or x31 and were assessed for illness and mortality. The 104-EID
50dose is slightly higher than the 50% minimal lethal dose (MLD50) for PR8. As a con-trol for high-path PR8 infection, a third group of mice was infected with 104 EID
50 of ⌬Vn1203, a genetically modified H5N1 virus known to cause terrible illness in humans as well as mice (22). As expected, mice that were infected with PR8 or
⌬Vn1203 exhibited severe illness that also manifested very early after infection (Fig. 1A). In contrast, x31-infected mice displayed a very mild illness profile. Differences in survival were equally dramatic, with nearly 80% of PR8-infected mice succumbing to infection by day 9, compared to complete sur-vival of the x31-infected group in all experiments (Fig. 1B). All mice infected with⌬Vn1203 were dead by day 7. Viral titers were similar in all three groups through day 6, though titers in the lungs of x31-infected mice were slightly reduced (Fig. 1C). At day 8, virus could no longer be detected in the lungs of x31-infected mice, whereas titers were still measurably high in the PR8-infected mice that had managed to survive the infec-tion to that point. All⌬Vn1203-infected mice had succumbed prior to day 8. These data demonstrate that PR8 and x31 can be used to compare high- and low-path influenza virus infections, and the disparities in the clinical outcomes cannot be suffi-ciently explained by differences in early viral replication in the lungs.
The composition of the airway microenvironment trends to-ward increased inflammation in high-path influenza A virus in-fection.High-path influenza A virus infections are often
de-fined by an overeffulgent cytokine storm (6–8). Because we observed severe illness only in mice infected with PR8, we asked whether hypercytokinemia was contributing to high-path PR8 infection. We investigated cytokine and chemokine levels using multiplex bead assays on supernatants from BAL fluid taken on days 2 through 8 postinfection to obtain a broad snapshot of the airway microenvironment after infection. Overall there was a trend toward increased cytokine levels in the BAL fluid during high-path infection, especially at early time points (Fig. 2). While some of the cytokines typically associated with the cyto-FIG 1Clinical profiles of high- and low-path influenza A virus infection. Mice were infected i.n. with 104EID
50of the PR8, x31, or⌬Vn1203 influenza A virus, and illness (A), survival (B), and virus titers (C) were measured at inter-vals. Panels A and C show results from a representative of five experiments (n⫽4 or 5 mice per group). Panel B shows survival from four combined experiments using between 20 (⌬Vn1203) and 55 total mice per group. *,P⬍ 0.05 for comparisons between PR8 and x31 at days 2, 4, and 8 and between x31 and⌬Vn1203 on day 2.
Rutigliano et al.
on November 7, 2019 by guest
http://jvi.asm.org/
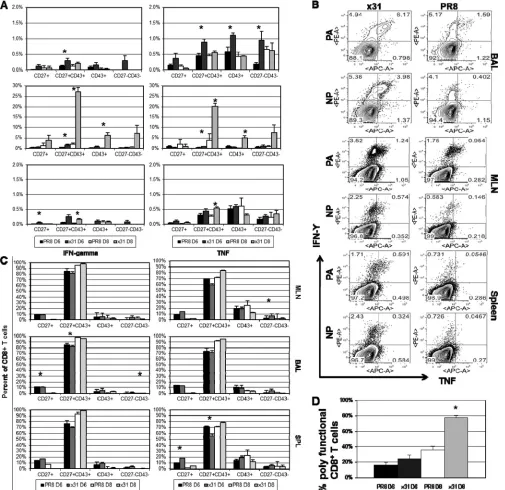
[image:3.585.300.544.67.518.2]kine storm were also upregulated in our experiments, some were not, while others were not consistently elevated across multiple experiments. In this study, IL-6 and G-CSF showed the most robust and consistent responses across multiple ex-periments. G-CSF is not a member of the IL-6 superfamily, but the complex formed by G-CSF and its receptor is similar to that of IL-6, IL-6R␣, and gp130 (23). These data demonstrate that the airway microenvironment is more inflammatory in high-path PR8 infection, although not as inflammatory as the level generally associated with hypercytokinemia reported in other high-path influenza virus studies.
Low-path infection is associated with a more stout CD8ⴙT cell response.Since an innate inflammatory milieu can influence adaptive immunity, we asked if there were differences in the CD8⫹ T cell responses to PR8 and x31 infections. Using tetramer stain-ing to enumerate influenza virus-specific CD8⫹T cells at days 6 and 8 postinfection, we found that mice infected with x31 had significantly increased numbers of PA224- and NP366-specific CD8⫹T cells in the BAL fluid at day 8 postinfection, and this was true in all experiments (Fig. 3A). Differences were not significant at day 6, and numbers were also much lower, which is expected
since the peak of the CD8⫹T cell response in the lung and airways does not begin until days 8 to 10 following influenza virus infec-tion. However, because of the extensive mortality in PR8-infected mice, we could not examine the response beyond day 8. To ensure that the differences seen at day 8 were not biased by the fact that we could only analyze mice that had survived PR8 infection to that point, we also examined the T cell response at day 7 (Fig. 3Band C). Here again we observed a significant increase in the number of tetramer-positive CD8⫹T cells in low-path x31-infected mice compared to high-path PR8-infected mice. These same trends were also recognized in the MLN and spleen at days 6, 7, and 8; however, differences were not as profound or significant, suggest-ing that the local lung airway microenvironment may be the cause of the reduced response during PR8 infection. Additional experi-ments showed that tetramer-positive CD8⫹T cells were incorpo-rating BrdU at similar levels at day 7 during high- and low-path infections (Fig. 3DandE), indicating that there is no difference in proliferation between the two groups. From these data, we con-clude that influenza virus-specific CD8⫹T cells are profoundly and significantly increased in low-path infections.
Given the disparity in the numbers of tetramer-positive cells at FIG 2Cytokine and chemokine production in the airway microenvironment after influenza virus infection. Mice were infected as described forFig. 1. At days 2, 4, 6, and 8 after infection, BAL fluid wash supernatants were collected and analyzed by multiplex cytokine bead analysis. The data set shows cytokines and chemokines that were consistently measured above the limit of detection across multiple experiments. The data show results from a representative experiment of five with 4 or 5 mice per group. *,P⬍0.05 for comparisons between PR8 and x31 at each time point.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:4.585.86.503.66.426.2]on November 7, 2019 by guest
http://jvi.asm.org/
day 8, we next compared cell surface phenotypes that reflect CD8⫹T cell differentiation states. Recent studies have highlighted the distinction of short-lived effector cells (SLEC) (KLRG1high
CD127lo) from memory precursor effector cells (MPEC)
(KLRG1loCD127high) in LCMV andListeriainfections (24–26). However, other research groups, including ours, have found that the KLRG1/CD127 divergence does not comfortably translate to all models, including influenza A virus infection (3,27). It there-fore appears that the MPEC/SLEC classification system may not be feasible in the C57BL/6 murine influenza virus infection model. For this reason, we investigated CD8⫹T cell activation pheno-types based on CD27 and CD43 expression. This classification scheme has been informative in Sendai virus infections and has also been extended to CD8⫹T cell memory in influenza virus infections (28,29). We found that the majority of influenza virus-specific CD8⫹T cells expressed the CD27⫹CD43⫹effector phe-notype (Fig. 3A to C). This was true for both DbPA
224- and DbNP366-specific CD8⫹T cells in the BAL fluid, MLN, and spleen at days 7 and 8 postinfection in all experiments. At day 6, numbers were much lower and differences between PR8 and x31 were small. Not surprisingly, we did not see high numbers of memory precursors (CD27⫹CD43⫺) in the BAL fluid at any time point, and other subpopulations had extremely low numbers as well. These data show that almost all of the tetramer-positive cells fell within the effector subpopulation in the BAL fluid, MLN, and spleen. Although numbers of influenza virus-specific CD8⫹T cells differ significantly in high- and low-path infections, the dif-ferentiation of the cells determined by this surface phenotype schema is unaffected.
Interestingly, we did not notice increased recruitment of cells associated with the innate response, despite the increased inflam-mation that was presented inFig. 2. G-CSF is a known chemoat-tractant for neutrophils (30), but we saw no difference in neutro-phil numbers until day 8 (Fig. 3F). Cells infected with influenza virus are also known to produce proinflammatory cytokines such as MCP-1/CCL-2 (31). CCL-2 recruits dendritic cell (DC) popu-lations to sites of inflammation, but under these specific experi-mental conditions, we did not see significantly elevated TNF/in-ducible nitric oxide synthase (iNOS)-producing DC (TipDC) or DC numbers early in the MLN of PR8-infected animals (Fig. 3G andH). Overall, though the numbers of influenza virus epitope-specific CD8⫹cytotoxic T lymphocytes (CTLs) differ substantially for high-path and low-path infections, the distributions of the various CD27/CD43 phenotypes within those sets are essentially comparable.
CD8ⴙT cells are more activated and polyfunctional in mice after low-path infection.Influenza virus-specific CD8⫹ T cell numbers were significantly elevated during low-path infection with x31. We followed this observation by evaluating the effector activity of influenza virus-specific CD8⫹T cells after infection. Cells were isolated from the indicated organs and stimulatedin
vitrowith PA224and NP366together prior to intracellular cytokine staining. Because CD8⫹T cells do not become activated and infil-trate the airway microenvironment very early after infection, we limited our analysis of intracellular cytokine production to days 6 and 8 postinfection.
We began by determining the magnitude of functionality within each subpopulation. Each CD27/CD43 subpopulation was examined after gating on CD8⫹T cells. We then calculated the percentage of cells that were producing cytokine within each sub-population. At day 8, effector CD8⫹T cells isolated from the BAL fluid of x31-infected mice exhibited impressively robust and sig-nificant increases in IFN-␥and TNF production compared to those from PR8-infected mice (Fig. 4A). We also observed in-creased cytokine production by effectors at day 6, but levels were much lower than at day 8. Again, to ensure that the disparity in CD8⫹T cell effector function at day 8 was not a result of being able to analyze only mice that had survived PR8 infection to that point, we also examined the T cell response at day 7 (Fig. 4B). As ex-pected, CD8⫹T cells isolated from mice with high-path infection were strikingly and significantly depressed in their ability to pro-duce antiviral cytokines compared to CD8⫹T cells isolated from mice with low-path infection. CD8⫹ T cells isolated from the MLN after x31 infection also showed elevated antiviral cytokine production at days 6, 7, and 8 compared to CD8⫹T cells from PR8-infected mice. These trends were also present in other CD27/ CD43 subpopulations but at greatly reduced magnitudes. The overwhelming majority of cytokine-producing CD8⫹T cells were CD27⫹CD43⫹effectors at days 6 and 8 postinfection in all three organs (Fig. 4C). Additionally, there was essentially no difference between low- and high-path infection, demonstrating that the re-sponding cells in high-path infection were not part of an aberrant subpopulation but were normally differentiated effectors. These results suggest that effector function appears to be somewhat di-vorced from surface phenotype classifications.
It was next important to ask whether CD8⫹T cells from x31-infected mice displayed greater polyfunctionality than CD8⫹T cells from PR8-infected mice. It has been shown that during in-fluenza virus infection, CD8⫹T cells begin producing IFN-␥upon activation, followed by TNF production as they become more ac-tivated and thus polyfunctional (32). We therefore calculated the percentage of IFN-␥⫹CD8⫹effector T cells that also produced TNF to determine polyfunctionality in the BAL fluid. At day 6, there was a slight increase in the percentage of IFN-␥⫹CD8⫹T cells producing TNF in the x31-infected mice (Fig. 4D), yet by day 8, this difference became dramatic and significant, with nearly 80% of the IFN-␥⫹CD8⫹T cells also producing TNF, compared to only 35% in PR8-infected mice. These data show that CD8⫹T cells in low-path infection are more polyfunctional and thus are more highly activated than their counterparts in high-path infec-tion. Moreover, cytokine production by the CD8⫹T cell effector population is much more efficient during infection with x31, and
FIG 3Influenza virus-specific effector CD8⫹T cells are more numerous in low-path infection. (A and B) Mice were infected as forFig. 1, and tetramer staining was used to enumerate influenza virus-specific DbPA
224⫹and DbNP366⫹CD8⫹T cells in the BAL fluid, MLN, and spleen at days 6 and 8 postinfection (A), as well as at day 7 (B). After gating on tetramer-positive cells, expression of CD27 and CD43 was examined, and each subpopulation is shown to the right of the tetramer-positive bars. (C) Representative dot plots of the data from panel B. (D) Proliferation of DbPA
224⫹and DbNP366⫹CD8⫹T cells in the BAL fluid, MLN, and spleen at day 7 was measured by incorporation of BrdU. (E) Representative dot plots for panel D. (F) Neutrophils were isolated by gating on CD11b⫹ CD11c⫺cells and then counting MHCII⫺GR1⫹cells. (G) TipDCs were identified as MCHII⫹GR1highcells. (H) Classical DCs in the MLN were identified by gating on CD11b⫹CD11c⫺cells and then counting MHCII⫹GR1⫺cells. The data are from a single representative of more than five experiments with 4 or 5 mice per group. *,P⬍0.05.
on November 7, 2019 by guest
http://jvi.asm.org/
these CD8⫹T cells also produce more cytokines per cell than their PR8 counterparts, suggesting that x31 infection is defined by an incredibly robust yet efficient CD8⫹T cell response that leads to a low-path infection.
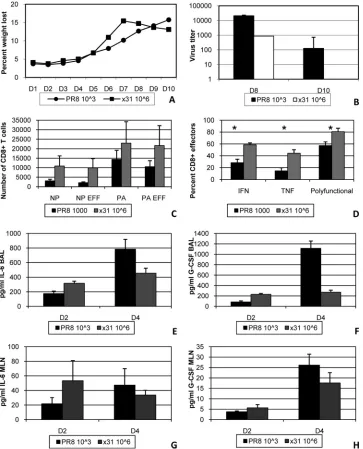
The previous experiments showed that high-path infection was accompanied by reduced numbers of activated, antigen-specific
CD8⫹T cells, which were also defined by reduced effector activity and polyfunctionality. In additional experiments, we changed the infectious doses so that illness would be similar in both groups. This was accomplished by infecting two groups of mice with 103 EID50PR8 or 106EID50 x31 (Fig. 5A). Importantly, all of our assays revealed the same trends in these experiments that we ob-FIG 4Activated influenza virus-specific CD8⫹T cells are almost uniformly CD27⫹CD43⫹effectors. (A) Peptide-induced cytokine production profiles for the different IFN-␥⫹and/or TNF⫹CD27/CD43 subsets shown inFig. 3. Cells were first gated on CD27/CD43 subsets, and the proportion making IFN-␥or TNF was calculated. (B) Representative dot plots of antiviral cytokine production by DbPA
224⫹and DbNP366⫹CD8⫹T cells at day 7 postinfection are shown to confirm that the responses seen at day 8 are under way at day 7. These plots are from the same experiments shown inFig. 3DandE. (C) Cells isolated from the mice sampled forFig. 3were assayed for IFN-␥and TNF production by ICS. Cells were analyzed by gating first on the cytokine-positive CD8⫹sets followed by CD27 and CD43 staining. (D) The percentage of CD8⫹IFN-␥⫹TNF⫹cells was then divided by the percentage of CD8⫹IFN-␥⫹cells to give the percentage of polyfunctional (IFN-␥⫹TNF⫹) cells within the IFN-␥⫹set. The data show results from a representative experiment of five with 4 or 5 mice per group. *,P⬍0.05. Rutigliano et al.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:7.585.43.548.68.558.2]served in our comparisons with 104EID
50. Virus titers were again slightly reduced in x31-infected mice, confirming that virus titer was not the sole factor in determining the severity of influenza virus-associated illness (Fig. 5B). At day 8, increased numbers of DbPA
224- and DbNP366-specific CD8⫹T cells were seen in the BAL fluid during x31 infection, and nearly all of these cells were CD27⫹ CD43⫹effectors (Fig. 5C). In addition, CD8⫹T cells from the BAL fluid of x31-infected mice produced significantly more cyto-kine and were more polyfunctional by ICS (Fig. 5D). When we repeated our multiplex cytokine bead analysis, we again found elevated levels of IL-6 and G-CSF in the airway microenvironment of PR8-infected mice at day 4 (Fig. 5EandF). Cytokine levels were
roughly equivalent in the MLN at the same time point (Fig. 5Gand H). Together, these results clearly show that x31 infection is de-fined by an incredibly robust yet efficient CD8⫹T cell response, regardless of the input virus dose.
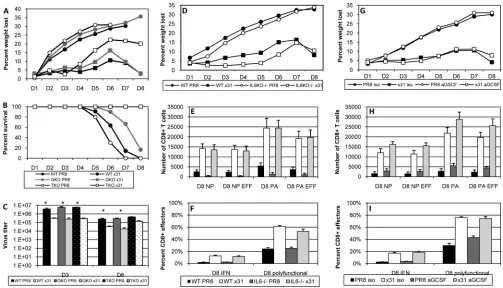
Individual cytokines and the CD8ⴙT cell response.The aug-mented CD8⫹ T cell polyfunctionality we saw in x31-infected mice led us to ask if we could reduce the disparity in pathology of PR8 and x31 infections in the absence of IFN-␥or TNF. IFN-␥⫺/⫺ and TNF⫺/⫺mice were thus infected with 104EID
50of either PR8 or x31 and then monitored for illness and survival. Illness profiles for IFN-␥⫺/⫺and TNF⫺/⫺mice were similar to those for WT controls for both PR8 and x31 infections (Fig. 6A). Interestingly, FIG 5Equalizing the illness outcome does not alter the CD8⫹T cell response to high- and low-path influenza virus infection. (A to D) Mice were infected i.n. with 103EID
50of PR8 or 106EID50of x31 to monitor illness by daily weight change (A) and viral titer (B) and for the analysis of BAL fluid populations taken on day 8 for tetramer-positive CD8⫹CTL numbers (C) and functional activation (D). EFF, tetramer-positive CD27⫹CD43⫹. (E to H) Multiplex cytokine bead analysis was used to assess the concentrations of IL-6 and G-CSF in supernatants from the BAL fluid (E and F) and MLN (G and H) at days 2 and 4 postinfection. The data are from a representative experiment of three with 4 or 5 mice per group. *,P⬍0.05.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:8.585.113.476.67.516.2]illness was slightly augmented in x31-infected TNF⫺/⫺mice, sug-gesting a protective role for TNF in response to x31 infection. However, the increased illness did not correlate with a change in mortality, as all x31-infected groups presented with 100% survival (Fig. 6B). On the other hand, extreme mortality defined all groups infected with PR8. There were slight differences in the kinetics of PR8-induced mortality when comparing WT PR8 infection to in-fection of IFN-␥⫺/⫺and TNF⫺/⫺mice with PR8, and the differ-ential kinetics of survival between IFN-␥⫺/⫺and TNF⫺/⫺mice was significant after PR8 infection. Viral titers in IFN-␥⫺/⫺and TNF⫺/⫺mice also followed the pattern seen in WT mice (Fig. 6C), again indicating that early differences in viral replication were insufficient to explain the disparities in morbidity and mortality. It therefore appears that the quality and efficiency of the response differs in the two infections and is the result of differential regu-lation of the development of CD8⫹T cells by the early responses to PR8 and x31.
Next, we investigated the roles of IL-6 and G-CSF, as these two cytokines were consistently elevated in our multiplex analyses (Fig. 2). Illness was not dramatically altered in IL-6⫺/⫺mice after infection, although there was a slight improvement that was more obvious in x31-infected IL-6⫺/⫺mice (Fig. 6D). In addition, sur-vival was not consistent across three independent experiments, only one of which was significant by Cox proportional hazards survival regression analysis. As always, no x31-infected mice
suc-cumbed to infection. Cellular analysis showed that there was still a disparity in CD8⫹T cell effector numbers between x31- and PR8-infected mice in the BAL fluid of IL-6⫺/⫺mice at day 8 (Fig. 6E). As with WT mice, CD8⫹effector T cells from the BAL fluid of IL-6⫺/⫺mice exhibited increased cytokine production and poly-functionality after low-path infection (Fig. 6F). In our experi-ments, functionality was always significantly impaired in high-path infection, but the disparity between high- and low-high-path infections was not always as severe in IL-6⫺/⫺mice as in WT mice. While these results suggest a role for the pleiotropic IL-6 during influenza virus infection, they do not demonstrate that IL-6 alone is responsible for the high-path phenotype of PR8 infection.
We also attempted to elucidate the role of G-CSFin vivoduring high- and low-path influenza virus infection. We used a purified neutralizing antibody against mouse G-CSF to deplete the cyto-kine at days 1, 3, and 5 postinfection. Pilot studies showed that administration of this antibody depleted G-CSF levels in the BAL fluid of a PR8-infected mouse by approximately half (data not shown). In subsequent experiments, depletion of G-CSF did not affect the illness profiles after high- and low-path infection (Fig. 6G). On the other hand, our cellular assays revealed that anti-G-CSF treatment did show signs of improving the CD8⫹T cell re-sponse during high-path infection, although the rere-sponses still did not begin to approach the levels seen in low-path x31-infected mice (Fig. 6HandI).
FIG 6High- and low-path infection in IFN-␥⫺/⫺, TNF⫺/⫺, IL-6⫺/⫺, and anti-G-CSF-treated mice. (A to C) WT B6, IFN-␥⫺/⫺, and TNF⫺/⫺mice were infected
i.n. with 104EID
50of PR8 or x31 to determine illness (A), survival (B), and lung virus titers (C) on days 3 and 6. (D to F) WT B6 or IL-6⫺
/⫺mice were infected
i.n. with 104EID
50of PR8 or x31. (G to I) In separate experiments, WT B6 mice were treated with 200g anti-G-CSF antibody or isotype control on days 1, 3, and 5. (D and G) Mice were weighed daily to monitor illness. (E and H) Tetramer-positive CD8⫹CTLs were enumerated on day 8, and the numbers of influenza virus-specific CD27⫹CD43⫹effector CD8⫹T cells were also calculated. (F and I) ICS was used to measure the percentage of CD27 CD43⫹effector CD8⫹T cells that were producing IFN-␥and also to measure polyfunctionality. The data show results from a representative experiment of three (A to F) or two (G to I) with 4 or 5 mice per group. *,P⬍0.05 for all comparisons between PR8-infected groups and their x31-infected counterparts in panels E, F, H, and I.
Rutigliano et al.
on November 7, 2019 by guest
http://jvi.asm.org/
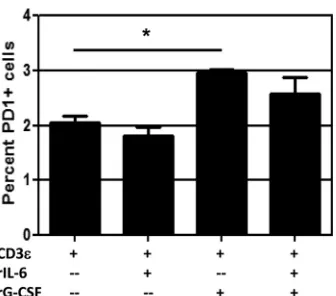
[image:9.585.40.543.65.353.2]These experiments suggested a possible role for IL-6 and G-CSF in influenza virus pathogenesis. Of course, both cytokines are pleiotropic, and other factors can potentially compensate for their absence. We therefore performed a smallin vitroexperiment to isolate the potential effects of IL-6 and G-CSF on the CD8⫹T cell response. CD8⫹T cells from the spleens of naive WT mice were stimulated by plate-bound anti-mouse CD3 as well as anti-CD28 antibody for 72 h in the presence of 5 ng/ml recombinant IL-6, G-CSF, or both. Activation was confirmed by CD27⫹CD43⫹ co-expression and intracellular staining for IFN-␥, and other markers associated with CD8⫹T cell function were examined. Our data revealed that treatment with recombinant G-CSF led to significant upregulation of the activation marker PD-1 (Fig. 7). PD-1 expres-sion was not affected by the addition of IL-6 to the culture during stimulation, nor did IL-6 affect the ability of G-CSF to induce PD-1 expression. Two other markers associated with CD8⫹T cell activation, Tim-3 and CTLA-4, were unchanged by the presence of IL-6 and G-CSFin vitro(data not shown). Although the differ-ences are small, they are significant and were consistently observed across five experiments, suggesting that G-CSF may specifically influence PD-1 expression and thus affect CD8⫹T cell function.
CD8ⴙT cells upregulate PD-1 expression in vivo during high-path infection.Our previous data revealed the possibility that PD-1 expression may be linked to the debilitated CD8⫹T cell response during high-path infection. We revisited our 104EID
50 infection model and found significantly increased PD-1 staining by mean fluorescence intensity (MFI) on CD8⫹T cells in the BAL fluid of PR8-infected mice at day 8 compared to cells isolated from x31-infected mice (Fig. 8A). This was true for CD8⫹T cells spe-cific for all 4 major class I MHC epitopes, indicating that PD-1 expression was not restricted to a subset of influenza virus-specific cells. Furthermore, PD-1 expression was not increased in the BAL fluid at day 6, although there was a trend toward elevated PD-1 staining on high-path CD8⫹T cells in the MLN at day 6. Addi-tional experiments that included day 5 showed that this staining was not present at early time points in the MLN (Fig. 8B),
indicat-ing that PD-1 upregulation was not originatindicat-ing prior to migration into the airway. Further analysis of PD-1 expression at day 7 showed that PD-1 expression in the BAL fluid was significantly increased on DbPA224- and DbNP366-specific CD8⫹T cells after high-path infection (Fig. 8C). Furthermore, this discrepancy was even more pronounced in the lung tissue (Fig. 8D). These data unambiguously demonstrate that PD-1 expression is significantly increased on influenza virus-specific CD8⫹T cells during high-path infection.
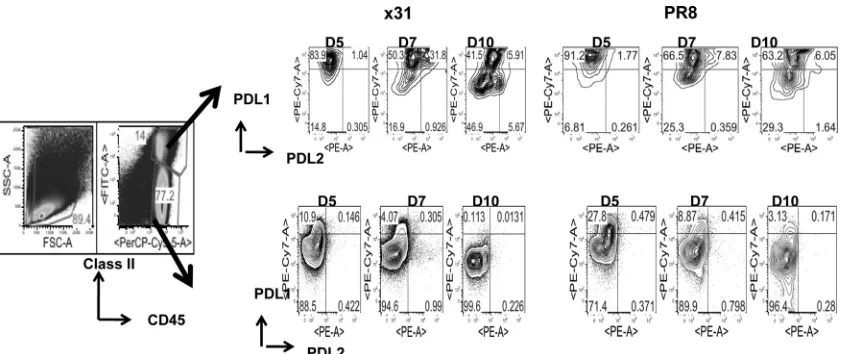
PD-1 has two known ligands, PD-L1 and PD-L2. We repeated our 104EID
50comparison and looked at expression of PD-L1 and PD-L2 on target cells in the airway (i.e., BAL fluid). It has been shownin situthat CD45⫹CD11c⫹DCs that infiltrated the lungs stimulated CD8⫹T cell cytolytic activity and cytokine release, but CD45⫺respiratory epithelium stimulated only cytolytic activity (12). Our data show that in mice infected with 104EID50of PR8, PD-L1 expression on MHCII⫹CD45⫹cells is elevated through-out the infection compared to that in x31-infected mice (Fig. 9). PD-L1 expression is also elevated on CD45⫹MHCII⫺cells during infection with 104EID50of PR8. Interestingly, we found that at day 7, coexpression of PD-L1 and PD-L2 on MHCII⫹CD45⫹cells was spectacularly increased in x31-infected mice compared to PR8-infected mice. However, CD45⫹/MHCII⫺ cells expressed significantly higher PD-L1 levels in the BAL fluid (Fig. 9) at almost all the time points tested after PR8 infection.
To see if we could rescue the high-path phenotype and corre-late PD-1 with reduced CD8⫹T cell function, we treated mice with antibody against PD-L1 during infection to block the PD-1/ PD-1L interaction. Treatment with anti-PD-L1 antibody alone has been shown to rescue PD-1-induced exhaustion (14). Our data revealed that anti-PD-L1 treatment had no effect on illness or survival in three separate experiments (Fig. 10A). However, anti-PD-L1 treatment did lead to a significant reduction in virus titers at day 10 in mice with high-path infection (P⫽0.043). In this case, titers in x31-infected mice had already fallen below the level of detection by day 8. Analysis of the CD8⫹T cell response showed that influenza virus-specific CD8⫹T cell numbers were signifi-cantly increased in anti-PD-L1-treated mice with high-path infec-tion at day 8 postinfecinfec-tion compared to isotype control-treated mice (Fig. 10B). This general trend was repeated in three separate experiments, although with a slight delay in one experiment. It was notable that anti-PD-L1 treatment had minimal effect on CD8⫹T cell numbers during x31 infection and that CD8⫹T cell functionality was still diminished in mice with high-path infection after anti-PD-L1 treatment (Fig. 10CtoE). The percentage of CD8⫹T cells producing cytokine was unchanged after anti-PD-L1 treatment in both infections. We also expanded upon our assess-ment of polyfunctionality by examining the percentage of IFN-␥⫹ CD8⫹effectors that also stained positive for the degranulation markers CD107a/b, and we saw no changes with anti-PD-L1 treat-ment (Fig. 10E). When we looked at other phenotypic markers that have been associated with diminished CD8⫹T cell activity, we saw no significant differences between any groups, suggesting that our experimental outcomes are restricted to effects mediated by PD-1 (Fig. 10FtoI). These data clearly demonstrate that upregu-lation of PD-1 during high-path infection correlates with reduced CD8⫹T cell numbers and activity. However, blockade of PD-L1 led to recovery of CD8⫹T cell numbers and reduced viral titers, indicating that although functionality was not rescued during high-path infection, the increased numbers did lead to better con-FIG 7G-CSF induces PD-1 expression on activated CD8⫹T cellsin vitro.
CD8⫹T cells were isolated from spleens of naive WT mice and cultured in 96-well plates precoated with 4g/ml anti-mouse CD3 and 1g/ml anti-mouse CD28 and rhIL-2. Triplicate wells were supplemented with 5 ng/ml recombinant mouse IL-6 or G-CSF as indicated. After 72 h, cells were treated with brefeldin A for 4 h, followed by intracellular cytokine staining for IFN-␥ to confirm that cells had been activated (not shown). Surface staining for PD-1 is shown. The data are from a single representative of more than five experi-ments. *,⫽P⬍0.05 for comparison between CD3εalone and CD3εplus rG-CSF.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:10.585.81.249.64.212.2]FIG 8PD-1 expression is upregulated on effector CD8⫹T cells in the BAL fluid after high-path infection. Mice were infected as described forFig. 1. (A) At days 6 and 8 postinfection, cells isolated from the BAL fluid, MLN, and spleen were assessed by flow cytometry. Influenza virus-specific CD8⫹T cells were identified by tetramer staining for all 4 epitopes recognized during primary infection. PD-1 expression on tetramer-positive/CD8⫹T cells was measured by mean fluorescence intensity. (B) PD-1 expression in the MLN at days 5 and 6 in a separate experiment to assess the likelihood of exhaustion occurring in the MLN before CD8⫹T cells migrated into the airway. (C and D) Representative dot plots and histograms illustrating the percentages of DbPA
224⫹and DbNP366⫹CD8⫹ T cells that express PD-1 at days 7 and 10 in the BAL fluid (C) and lung (D). The data are a single representative of more than five experiments with 4 or 5 mice per group. *,P⬍0.05.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:11.585.43.542.29.638.2]trol of the infection. In summary, blocking the PD-1/PD-L1 inter-action increases the numbers of epitope-specific CD8⫹CTLs that can be recovered from the respiratory tract in high-path versus low-path influenza A virus infection, but this has no effect either on the diminished cytokine response profile for individual re-sponding CD8⫹T cells or on the overall severity of the disease process.
DISCUSSION
The history of influenza is dominated by routine pandemics that devastate the human population (33–36). Tremendous effort has been made to discover why these pandemic viruses are so severe, but the answer remains incomplete. In the current study, we have shown that high-path influenza virus infection is the result of a blunted CD8⫹T cell response characterized by reduced numbers and functionality. The diminished CD8⫹T cell response during high-path infection coincided with increased PD-1 expression. Anti-PD-L1 treatment rescued influenza virus-specific CD8⫹T cell numbers and enhanced virus clearance, but CD8⫹T cell func-tionality was not restored. Other studies have confirmed that CD8⫹T cell exhaustion is a contributing factor to depressed im-munity in chronic infections such as LCMV (13,14) and in some acute lower respiratory tract infections (15). In contrast to these and other studies, we did not observe a full reversal of CD8⫹T cell functionality by blocking the PD-1/PD-L1 interaction.
Several factors may contribute to the depressed CD8⫹T cell response during PR8 infection. We observed a much more proin-flammatory airway microenvironment, suggesting that this milieu promotes abnormal development of the CD8⫹T cell response and high-path infection. Our analysis of the airway microenvironment revealed that several cytokines were dramatically increased at early time points during PR8 infection. Among the elevated cytokines was MCP-1/CCL2, which has been shown to recruit TipDCs to the site of infection and promote CD8⫹expansion and survival (37). It is interesting to note that in our study, the elevated CCL2 re-sponse during high-path infection actually appears to prevent PR8-infected mice from suffering even more severe illness than we observed. IL-12p70 production was also increased in the airway
microenvironment of PR8-infected mice. The role of IL-12 in the induction of T cell responses is well established (38). As with IL-12p70, we saw early, increased production of IL-6 and G-CSF in PR8-infected mice, whereas levels in x31-infected mice started much lower and gradually increased to coincide with the peak of the CD8⫹T cell response. In vitroexperiments suggested that G-CSF may be able to induce PD-1 expression on activated CD8⫹ T cells, just as we observedin vivo. The sum of our data therefore leads us to hypothesize that high-path influenza virus infection is defined by a highly inflammatory airway microenvironment at early time points, and these proinflammatory factors, including G-CSF and others, induce PD-1 expression on CD8⫹T cells as they infiltrate the airway. Although these CD8⫹T cells cannot be said to be exhausted based on the currently accepted definition, it is unambiguously clear that debilitated effector functionality in acute, high-path influenza A virus infection is associated with in-creased PD-1 expression on virus-specific CD8⫹T cells.
Based on these results, depletion of CD8⫹T cells in the low-path infection might be expected to generate the high-low-path phe-notype. However, numerous studies in x31-infected mice have demonstrated that CD8 T cell depletion does not cause severe mortality, though it does delay viral clearance. This is likely due to a highly effective compensating antibody response in these ani-mals (39–41).
Costimulatory roles for PD-1 have been described. Experi-ments on tolerance induction showed that lymphatic endothelial cells upregulate PD-1 in the absence of costimulation through 4-1BB (42). Increased PD-1 expression led to reduced expression of the IL-2 receptor and reduced CD8⫹T cell survival. Another recent study revealed that inhibition of PD-L1 activity compro-mised CTL expansion duringListeriainfection but not during vesicular stomatitis virus infection (43). These studies are consis-tent with a costimulatory role for PD-1 in acute influenza virus infection that leads to the canonical exhaustion phenotype and functionality seen in chronic infections.
Infection of primary cultures of human airway epithelia has shown that PR8 was more infectious than x31 (44). This early, increased replication may be propelling the genesis of a highly FIG 9PD-L1 expression is upregulated on target cells during high-path infection. Mice were infected with 104EID
50of PR8 or x31 and sacrificed at days 5, 7, and 10 to assess expression of PD-L1 and PD-L2 on cells targeted by CD8⫹T cells in the BAL fluid. Target cells were defined by expression of class II MHC, and CD45.2 PD-L1 and PD-L2 expression was examined after gating on CD45⫹class II MHC⫹and CD45⫹class II MHC⫺cells. The data show dot plots from a representative experiment of two with 4 or 5 mice per group.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:12.585.83.506.65.242.2]on November 7, 2019 by guest
http://jvi.asm.org/
proinflammatory milieu that ablates CD8⫹T cell function during acute influenza virus infection. In contrast, the slower replication of x31 during the earliest stages of infection may allow for a more methodical and efficient development of airway inflammation and the ensuing CD8⫹ T cell response. Previously referenced studies of PD-1 have shown that anti-PD-L1 treatment reversed the exhausted phenotype and restored CD8⫹T cell functionality through blockade of the PD-1/PD-L1 interaction. While we saw increased numbers after blocking PD-L1 in vivo, CD8⫹T cell function was not restored in high-path infection, compared to no apparent effect in low-path infection. The inability to rescue ef-fector function suggests that the airway microenvironment in high-path infection may overwhelm the CD8⫹T cell to the point that effector function cannot be rescued, even when PD-L1 -me-diated suppression has been abrogated. The anti-PD-L1 treatment thus rescues CD8⫹T cell numbers in high-path influenza virus infection, but not functionality. Additional studies to address this disparity are under way.
The slight differences that we saw in virus titers early after infection were not sufficient to wholly explain the mechanism of high-path infection. Because PR8 and x31 express different sur-face hemagglutinin and neuraminidase proteins, there may be any number of effects on attachment, fusion, or release of progeny virions that are magnified early, only to equilibrate later. Some viruses demonstrate a reduced ability to infect certain cell types (45), although recent mathematical modeling studies suggest that infectivity does not affect peak viral load (46). The influenza virus NS1 protein counters innate and adaptive immunity by inhibiting type I IFN production (47), but in our model both viruses contain the same NS1 gene. It is also known that increased cytokine pro-duction is associated with more pathogenic strains, even in cases of similar viral replication (5).
IL-6 and G-CSF both signal through Jak1-mediated phosphor-ylation of STAT3, which then dimerizes and acts as a transcription factor. We observed that infection of IL-6⫺/⫺mice failed to rescue the high-path phenotype. However, all cytokines in the IL-6 su-perfamily employ the gp130 receptor (48). Many of these cyto-kines are pleiotropic and have overlapping functions, making it difficult to discern individual contributions from any one family member to sophisticated networks and responses. To assess any effects from this family of cytokines, be it individually or the fam-ily as a whole, it may be more informative to target the common gp130 receptor. Like that of IL-6, G-CSF-mediated signaling de-pends on Jak1, although Jak2 and Tyk2 can also be used (23,49, 50). In fact, the three-dimensional atomic structures of the human G-CSF/G-CSFR, IL-6/IL-6R␣/gp130, and IL-12/IL-12R com-plexes are all similar (48,51,52). It is therefore not surprising that signaling through the G-CSFR initiates the same signaling path-ways as the IL-6 family of cytokines. Ourin vivodata showed a
slight improvement in effector CD8⫹T cell responses after anti-G-CSF treatment during high-path infection. We therefore spec-ulate that the highly proinflammatory nature of the airway microenvironment during PR8 infection may be the result of dys-regulation of gp130-mediated signaling or the common Jak2/ STAT3 signaling pathway initiated by the IL-6 superfamily recep-tors and the G-CSF receptor.
Suppressor of cytokine signaling 3 (SOCS3) inhibits STAT3-mediated proinflammatory signaling (23,52–54). SOCS3 is up-regulated by several cytokines, including IL-6, IL-10, and G-CSF, all of which induce STAT3 activation. SOCS3 then acts in a nega-tive feedback loop to suppress IL-6 and G-CSF activity (55,56), but does not inhibit IL-10. SOCS3 targets specific residues in gp130 and G-CSFR to disrupt signaling through STAT3 (53,54, 57). This raises the possibility that disruption of SOCS3-mediated inhibition of proinflammatory responses may be a contributing factor to high-path influenza virus infection. It has been suggested that IL-7 can suppress SOCS3 in T cells, leading to IL-6 produc-tion (58). This allowed for extensive expansion of naive and effec-tor T cells, which was at least partially dependent on IL-6. Our multiplex data from the airway microenvironment did not reveal consistent results with IL-7, but we did see expansion of CD8⫹T cell numbers after anti-PD-L1 treatment and reduced viral titers in mice with high-path infection. One might therefore hypothe-size a potential mechanism by which PD-L1 also promotes SOCS3 activity, which may explain the elevated IL-6 and reduced num-bers we saw in high-path infection.
Certainly, any number of proteins associated with immune modification could be contributing to the disparate outcomes of high- and low-path influenza virus infection. One protein of in-terest might be T cell immunoglobulin and mucin protein-3 (Tim-3). Recently, it was shown that blocking the interaction of Tim-3 with its ligand, galectin-9, increased the CD8⫹T cell re-sponse to influenza virus infection (21). Another study has shown that the most severely exhausted CD8⫹T cells during chronic LCMV infection coexpress Tim-3 and PD-1 (59). During acute LCMV infection, Tim-3 was temporally expressed but eventually downregulated by day 30. However, we did not see any difference in Tim-3 expression in any of our experiments. Similarly, we saw no changes in expression of the T cell inhibitor CTLA-4. While it may not be surprising that observations associated with PD-1 and PD-L1 in chronic infection do not necessarily correlate to acute influenza virus infection, it is intriguing that we have found a strong relationship between PD-1 expression and high-path influ-enza virus infection that does not seem to respond to the same treatments that rescue CD8⫹T cell exhaustion in chronic LCMV infection.
Our data suggest that dysregulation of the early immune re-sponse after high-path infection manifests as a highly
proinflam-FIG 10Administration of anti-PD-L1 rescues CD8⫹T cell numbers but not functionality or illness. Mice infected i.n. with a dose of 104EID
50of PR8 or x31 were treated with 200g anti-PD-L1 antibody or isotype control antibody starting the day before infection and continuing every other day thereafter. (A) Illness and survival were assessed daily. Viral titers were measured by plaque assay at the indicated days. (B) Tetramer-positive CD8⫹T cells in the BAL fluid at day 8 postinfection (left graphs). After gating on tetramer-positive populations, activation phenotypes were measured, and the number of CD27⫹CD43⫹influenza virus-specific CD8⫹T cells was calculated (EFF) (right graphs). (C to E) Activation and effector function was measured by ICS afterex vivostimulation with PA224 and NP366peptides. The percentages of IFN-␥⫹effectors (C) and polyfunctional effectors (D) were determined as described forFig. 5. (E) The percentage of CD107a/b⫹effectors was determined by calculating the percentage of effectors that were IFN-␥⫹/CD107a/b⫹and dividing that percentage by the total percentage of IFN-␥⫹effectors. (F to I) Cells from the mice used forFig. 7and8were also assayed for Tim-3 (F and G) and CTLA-4 (H and I) expression by flow cytometry. The day 8 BAL fluid CTLs were analyzed by MFI for Tim-3 and CTLA-4 staining of tetramer-positive CD8⫹TLs. The data show a representative experiment of two with 4 or 5 mice per group. *,P⬍0.05.
on November 7, 2019 by guest
http://jvi.asm.org/
matory airway microenvironment that induces CD8⫹T cell ex-haustion, defined by increased PD-1 expression. Blockade of the PD-1/PD-L1interaction did rescue CD8⫹T cell numbers and im-proved virus clearance, but it failed to restore effector function on a per-cell basis. These data provide insight into the mechanism of high-path influenza virus infection and may identify potential therapeutic targets, should another devastating pandemic on the level of the 1918 Spanish flu assail mankind.
ACKNOWLEDGMENTS
We thank Scott Brown and Vandana Chaturvedi for insightful sugges-tions and invaluable discussion.
This work was supported by NIH grants AI065097 (P.G.T) and RO170251 (P.C.D.) and the NIH/NIAID St. Jude Center of Excellence for Influenza Research and Surveillance (HHSN266200700005C) and Amer-ican Lebanese Syrian Associated Charities at St. Jude Children’s Research Hospital.
We have no financial conflicts of interest or competing interests.
REFERENCES
1.Moskophidis D, Kioussis D.1998. Contribution of virus-specific CD8⫹ cytotoxic T cells to virus clearance or pathologic manifestations of influ-enza virus infection in a T cell receptor transgenic mouse model. J. Exp. Med.188:223–232.http://dx.doi.org/10.1084/jem.188.2.223.
2. Thomas PG, Keating R, Hulse-Post DJ, Doherty PC. 2006. Cell-mediated protection in influenza infection. Emerg. Infect. Dis.12:48 –54. http://dx.doi.org/10.3201/eid1201.051237.
3.Valkenburg SA, Rutigliano JA, Ellebedy AH, Doherty PC, Thomas PG, Kedzierska K.2011. Immunity to seasonal and pandemic influenza A viruses. Microbes Infect. Inst. Pasteur13:489 –501.http://dx.doi.org/10 .1016/j.micinf.2011.01.007.
4.Kim TS, Sun J, Braciale TJ. 2011. T cell responses during influenza infection: getting and keeping control. Trends Immunol. 32:225–231. http://dx.doi.org/10.1016/j.it.2011.02.006.
5.Sanders CJ, Doherty PC, Thomas PG.2011. Respiratory epithelial cells in innate immunity to influenza virus infection. Cell Tissue Res.343:13– 21.http://dx.doi.org/10.1007/s00441-010-1043-z.
6.Chan MC, Cheung CY, Chui WH, Tsao SW, Nicholls JM, Chan YO, Chan RW, Long HT, Poon LL, Guan Y, Peiris JS.2005. Proinflamma-tory cytokine responses induced by influenza A (H5N1) viruses in primary human alveolar and bronchial epithelial cells. Respir. Res.6:135.http://dx .doi.org/10.1186/1465-9921-6-135.
7.Cheung CY, Poon LL, Lau AS, Luk W, Lau YL, Shortridge KF, Gordon S, Guan Y, Peiris JS.2002. Induction of proinflammatory cytokines in human macrophages by influenza A (H5N1) viruses: a mechanism for the unusual severity of human disease? Lancet360:1831–1837.http://dx.doi .org/10.1016/S0140-6736(02)11772-7.
8.Lipatov AS, Andreansky S, Webby RJ, Hulse DJ, Rehg JE, Krauss S, Perez DR, Doherty PC, Webster RG, Sangster MY.2005. Pathogenesis of Hong Kong H5N1 influenza virus NS gene reassortants in mice: the role of cytokines and B- and T-cell responses. J. Gen. Virol.86:1121–1130. http://dx.doi.org/10.1099/vir.0.80663-0.
9.To KKW, Hung IFN, Li IWS, Lee K-L, Koo C-K, Yan W-W, Liu R, Ho K-Y, Chu K-H, Watt C-L, Luk W-K, Lai K-Y, Chow F-L, Mok T, Buckley T, Chan JFW, Wong SSY, Zheng B, Chen H, Lau CCY, Tse H, Cheng VCC, Chan K-H, Yuen K-Y.2010. Delayed clearance of viral load and marked cytokine activation in severe cases of pandemic H1N1 2009 influenza virus infection. Clin. Infect. Dis.50:850 – 859.http://dx.doi.org /10.1086/650581.
10. Iwasaki A, Medzhitov R.2004. Toll-like receptor control of the adaptive immune responses. Nat. Immunol.5:987–995.http://dx.doi.org/10.1038 /ni1112.
11. Vankayalapati R, Garg A, Porgador A, Griffith DE, Klucar P, Safi H, Girard WM, Cosman D, Spies T, Barnes PF.2005. Role of NK cell-activating receptors and their ligands in the lysis of mononuclear phago-cytes infected with an intracellular bacterium. J. Immunol.175:4611– 4617.
12. Hufford MM, Kim TS, Sun J, Braciale TJ.2011. Antiviral CD8⫹T cell effector activities in situ are regulated by target cell type. J. Exp. Med. 208:167–180.http://dx.doi.org/10.1084/jem.20101850.
13. Zajac AJ, Blattman JN, Murali-Krishna K, Sourdive DJ, Suresh M, Altman JD, Ahmed R.1998. Viral immune evasion due to persistence of activated T cells without effector function. J. Exp. Med.188:2205–2213. http://dx.doi.org/10.1084/jem.188.12.2205.
14. Barber DL, Wherry EJ, Masopust D, Zhu B, Allison JP, Sharpe AH, Freeman GJ, Ahmed R.2006. Restoring function in exhausted CD8 T cells during chronic viral infection. Nature439:682– 687.http://dx.doi .org/10.1038/nature04444.
15. Erickson JJ, Gilchuk P, Hastings AK, Tollefson SJ, Johnson M, Down-ing MB, Boyd KL, Johnson JE, Kim AS, Joyce S, Williams JV.2012. Viral acute lower respiratory infections impair CD8⫹T cells through PD-1. J. Clin. Invest.122:2967–2982.http://dx.doi.org/10.1172/JCI62860. 16. Goldberg MV, Maris CH, Hipkiss EL, Flies AS, Zhen L, Tuder RM, Grosso JF, Harris TJ, Getnet D, Whartenby KA, Brockstedt DG, Dubensky TW, Jr, Chen L, Pardoll DM, Drake CG.2007. Role of PD-1 and its ligand, B7-H1, in early fate decisions of CD8 T cells. Blood110: 186 –192.http://dx.doi.org/10.1182/blood-2006-12-062422.
17. Kilbourne ED.1969. Future influenza vaccines and the use of genetic recombinants. Bull. World Health Organ.41:643– 645.
18. Lamb RA, Krug RM.1996. Orthomyxoviridae: the viruses and their replication. Lippincott-Raven Publishers, Philadelphia, PA.
19. Hoffmann E, Krauss S, Perez D, Webby R, Webster RG.2002. Eight-plasmid system for rapid generation of influenza virus vaccines. Vaccine 20:3165–3170.http://dx.doi.org/10.1016/S0264-410X(02)00268-2. 20. Webby RJ, Andreansky S, Stambas J, Rehg JE, Webster RG, Doherty
PC, Turner SJ.2003. Protection and compensation in the influenza virus-specific CD8⫹T cell response. Proc. Natl. Acad. Sci. U S.100:7235–7240. http://dx.doi.org/10.1073/pnas.1232449100.
21. Sharma S, Sundararajan A, Suryawanshi A, Kumar N, Veiga-Parga T, Kuchroo VK, Thomas PG, Sangster MY, Rouse BT.2011. T cell immu-noglobulin and mucin protein-3 (Tim-3)/Galectin-9 interaction regulates influenza A virus-specific humoral and CD8 T-cell responses. Proc. Natl. Acad. Sci. U. S. A. 108:19001–19006. http://dx.doi.org/10.1073/pnas .1107087108.
22. Rutigliano JA, Morris MY, Yue W, Keating R, Webby RJ, Thomas PG, Doherty PC.2010. Protective memory responses are modulated by prim-ing events prior to challenge. J. Virol.84:1047–1056.http://dx.doi.org/10 .1128/JVI.01535-09.
23. Panopoulos AD, Watowich SS.2008. Granulocyte colony-stimulating factor: molecular mechanisms of action during steady state and “emer-gency” hematopoiesis. Cytokine42:277–288.http://dx.doi.org/10.1016/j .cyto.2008.03.002.
24. Hand TW, Morre M, Kaech SM.2007. Expression of IL-7 receptor␣is necessary but not sufficient for the formation of memory CD8 T cells during viral infection. Proc. Natl. Acad. Sci. U. S. A.104:11730 –11735. http://dx.doi.org/10.1073/pnas.0705007104.
25. Joshi NS, Cui W, Chandele A, Lee HK, Urso DR, Hagman J, Gapin L, Kaech SM.2007. Inflammation directs memory precursor and short-lived effector CD8(⫹) T cell fates via the graded expression of T-bet transcrip-tion factor. Immunity27:281–295.http://dx.doi.org/10.1016/j.immuni .2007.07.010.
26. Sarkar S, Kalia V, Haining WN, Konieczny BT, Subramaniam S, Ahmed R.2008. Functional and genomic profiling of effector CD8 T cell subsets with distinct memory fates. J. Exp. Med.205:625– 640.http://dx .doi.org/10.1084/jem.20071641.
27. Trobaugh DW, Yang L, Ennis FA, Green S. 2010. Altered effector functions of virus-specific and virus cross-reactive CD8⫹T cells in mice immunized with related flaviviruses. Eur. J. Immunol.40:1315–1327. http://dx.doi.org/10.1002/eji.200839108.
28. Hikono H, Kohlmeier JE, Takamura S, Wittmer ST, Roberts AD, Woodland DL.2007. Activation phenotype, rather than central- or effector-memory phenotype, predicts the recall efficacy of memory CD8⫹T cells. J. Exp. Med.204:1625–1636.http://dx.doi.org/10.1084 /jem.20070322.
29. Roberts AD, Ely KH, Woodland DL.2005. Differential contributions of central and effector memory T cells to recall responses. J. Exp. Med.202: 123–133.http://dx.doi.org/10.1084/jem.20050137.
30. Lieschke GJ, Stanley E, Grail D, Hodgson G, Sinickas V, Gall JA, Sinclair RA, Dunn AR.1994. Mice lacking both macrophage- and gran-ulocyte-macrophage colony-stimulating factor have macrophages and co-existent osteopetrosis and severe lung disease. Blood84:27–35. 31. Hofmann P, Sprenger H, Kaufmann A, Bender A, Hasse C, Nain M,
Gemsa D.1997. Susceptibility of mononuclear phagocytes to influenza A Rutigliano et al.