Copyrighti 1974 AmericanSocietyforMicrobiology Printed inU.S.A.
Isolation
and
Characterization
of
Sendai
Virus
Temperature-Sensitive
Mutants
ALLEN PORTNER, PRESTON A. MARX, AND D. W. KINGSBURY
Laboratoriesof Virology andImmunology, St. Jude Children's Research Hospital, Memphis, Tennessee38101 Received forpublication 27September1973
Ten temperature-sensitive mutants ofSendai virus, a paramyxovirus, were
isolated and partially characterized. Themutantsreplicatedinchicken embryo
lung cells at 30C, but not at 38C; wild-type virus grew equally well at both
temperatures. Complementation tests divided the mutants into seven groups. Sixgroupssynthesizedneither infectious virusnorRNA when incubatedat38C
fromthebeginningofinfection.Temperature shift-up experimentsdemonstrated
thatthree ofthesecomplementationgroupswereblocked inearlystepsrequired
for RNA synthesis, but these gene functions were not needed throughout the
replicative cycle. In contrast, the other three RNA-negative complementation
groupswere defectivethroughout the replicative cycle infunctions required for
virus-specific RNAsynthesis. Onlyone mutant, whichcomplemented allofthe
above, synthesized RNA but not infectious virus when placed at 38C; the
hemagglutinin ofthis mutantfunctionedonlyatthe permissivetemperature.
Temperature-sensitive (ts) mutants of
ani-mal viruses have been useful instudying virus
replication (6, 7). With respect to
paramyx-oviruses, a number ofNewcastle disease virus
(NDV) nonconditional-lethal mutants as well
as ts mutants havebeenisolated,butthesehave
been used mainly to study genetic interactions
(4, 8, 9). We have isolated andcharacterized a
number of Sendai virus ts mutants to learn
more about biochemical events in
paramyx-ovirusreplication. Preliminary genetic and
bio-chemical analysesof 10 ofthese mutants are the
subject ofthis report.
MATERIALS AND METHODS
Virus and cells. A clone ofwild-typeSendai virus wasplaque purified from the Enders strain of Sendai virus and was free from incomplete virions (17).
Stocksof viruswereprepared byinfecting10-day-old
embryonated hens' eggs with 0.01 hemagglutinating unit and incubating for 72 h at 37C. Methods for preparing chicken embryo lung (CEL) cell cultures, forgrowing Sendai virusinthesecells, andforplaque assay were described (5), but in the present work, Eagleminimalessential medium was used as growth mediumand in theplaque overlay.
Virions were labeled with radioactive amino acids asdescribed (18), except that labeled precursors were added 16h after infection andvirions werecollected 32 h later. Culture fluids containing released virus were centrifuged for 10 min at 3,600 x g toremove cells and debris. Viruswaspelleted (78,000 x g, 5C, 30min)andresuspendedin 0.01Msodiumphosphate (pH 7.2) for fractionation experiments.
RNAextraction, rate-zonalcentrifugation, and
radioactivity determinations. These methods have allbeen fully described (13).
Selection ofts mutants. The methods for muta-genization of wild-type virus and isolation of ts mutants weresimilar to thosedescribed forSindbis virus (2) and vesicular stomatitis virus (14). After mutagenization with
N-methyl-N'-nitro-N-nitrosoguanidine (2), the remaining infectious titer was 10-3 of the initial titer. Mutagenization by
5-fluorouracil (FU)(14)wasdonebytreatingCEL cell
monolayerswith200jgofthecompoundperml for 1
h, by infectingwith10plaque-formingunits(PFU)of Sendai virus per cell, and by allowing the virus to growfor 48 hin the presenceofFU. Aftertreatment with mutagens, the virus stocks were diluted to produceafewplaquesat 30C.Afterincubation for7 to 10 days, well-isolated plaques were picked,
sus-pended in 1 ml ofphosphate-buffered saline (PBS) containing 5% fetal calf serum, and replated at
permissive(P) temperature (30C)andnonpermissive
(NP)temperature(38 C).Tenisolateswere tempera-ture-sensitive, whereas all others tested were not. Stocks of each ts isolate were
grownl
in CEL cell monolayers at 39C for further testing. Virus was harvested when thehemagglutinin titerwas27/mlor greater.Complementation tests. Complementation tests weredoneinCEL cell monolayer cultures containing about106cellsatthe timeofinfection.Sendai virusts mutants were diluted in PBS to give an input multiplicityof5PFU/cell in asingle infectionor2.5
PFU/cell ofeach mutant in mixed infection. Virus wasadsorbed for 30min at25C, the monolayers were washed with PBS, and5 ml ofgrowth medium was added.Afterincubationat 38Cfor 4h, cultureswere washedagain, prewarmedgrowth medium was added, andmonolayerswerereincubatedat38Cfor 48 to 72 298
on November 10, 2019 by guest
http://jvi.asm.org/
SENDAI VIRUSTEMPERATURE-SENSITIVE MUTANTS
h. The mediumwasthen assayedforplaque formation
on CEL cell monolayers at 30C. Complementation levelswerecalculated accordingtoBurge and Pfeffer-korn (3).
Isolationof virion glycoproteins. Sendai virions
werefractionated bytreating them (1mg ofprotein
perml) with2%TritionX-100 and1MKClfor20min
at 25C (15, 16). The mixture was centrifuged, at
100,000xgfor 30minat5C, and the glycoproteins in the 100,000 x gsupernatantwereseparatedfromthe
smallest virion polypeptide by dialysis and centrifu-gation (15, 16).
Acrylamide gel electrophoresis.Thismethodwas
previously described(18).
Neuraminidase assay. Fetuin wasused as a
sub-strate. Freesialic acidwas measuredbythe
thiobar-bituric acid procedure(19, 20). Duplicate samplesof virionsor the glycoprotein fraction from virions (15,
16)werediluted with0.2M sodium phosphate buffer (pH5.9)to0.05ml, and0.05ml offetuin (6.25mg/ml) inthesamebufferwasadded. Theincubation temper-ature was30or38C.
RESULTS
Isolation ofts mutants. With nitrosoguani-dine and FU mutagenesis, about 1.5 and 1.0%, respectively, of the plaque isolates contained useful ts mutants. When mutant stocks were
tested at 38 and 30C, ratios of plaques
pro-ducedranged from less than 1.6 x 10-' (ts 271) to 2.5 x 108 (ts 935) (Table 1). Only a few
plaques from the 38 C plateswerenot
tempera-ture-sensitive when picked and tested again at
38 and30C, indicating that themutantshadlow back mutation frequencies and were slightly
"leaky." The frequency of wild-type revertants
in mutant stocks ranged from 3.8 x 10-'to4.5
x 106.
Growth of ts mutants at P and NP temperatures. To examine the growth of ts mutants, cultures were infected with an input
multiplicity of about 5 PFU/cell and incubated
at P and NPtemperatures. Growth curves are
shown forwild-type virus (Fig. 1A) andmutant
TABLE 1. Efficiency of plating of wild-typevirus and
tsmutants
38C/30 C ratio
Virus (PFU/ml)
Wild-type 1.0
ts74 ... 2.4 x 10-'
ts 105 ... 1.2 x 10-'
ts 245 1.2 x 10'
ts271 ... 1.6 x 10'
ts348 ... <2.7 x 10-5
ts557 ... 6.0 x10'
ts595 ... <1.0 x10-7
ts642 ... 1.0 x10-5
ts840 ... <8.0 x10-v
ts935 ... 2.5 x 10-B
-i 2
-0 24 48 72 96 120 0 24 48 72 96 120
HOURS AFTER INFECTION
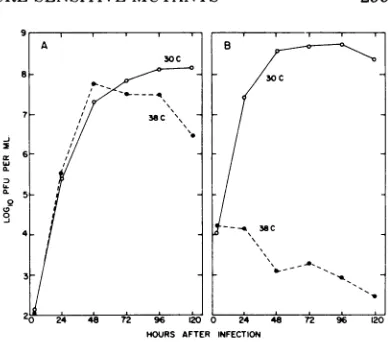
FIG. 1. Growth of wild-type Sendai virus and mu-tant ts271at30and38C. CEL cell monolayerswere
infected with an input multiplicity of 5 PFUIcell. After adsorption (30 min at 24C), monolayers were
washed andincubatedat30or38 C.Atintervals after
adsorption, culture fluids were removed from the
monolayers and assayed forinfectious virusat30C by theplaquemethod. A, Wild-type virus; B, mutant ts
271.0,38 C: 0,30C.
ts 271 (Fig. 1B). Wild-type virus grew equally
well at 30and 38 C. Mostts mutants werelike ts 271, producing virus at approximately the
same rate at 30C as wild-type virus, but little or no virus at 38 C. Typical amounts of virus released 48 h after infection by each mutantare
shown in Table 2. All mutants synthesized substantialamountsof virus at30 C, andsome
(ts 74, 105, 245, and 271) grew better than
wild-type virus at 30 C. In contrast to growth under permissive conditions, mutant yields at
TABLE 2. Growthof Sendaivirusmutants at30and 38 Ca
Yield(PFU/ml) 38C/30 Cratio
Virus 30 C 38 C (PFU/ml)
Wild-type 2.0x 107 6.0 x 107 3
ts74 3.4x 10' 1.8 x 102 5.3x 10-7
ts105 2.0x 108 <101 <5.0x 10-8 ts245 6.3x 107 <101 <1.6x10-7 ts271 3.6x 10 1.2x 103 3.3x 10-B
ts348 6.3x 105 <101 1.6 x 105
ts557 5.0x 10' <101 <2.0x 10-4 ts595 1.6x 107 <101 <6.3x10-7
ts642 2.5 x 107 5.0x10' 2.0x 10-'
ts840 4.0x 105 <10l <2.5x 10-5
ts935 2.0x 106 <101 <5.0x 10-aCEL cellmonolayers wereinfected withan input
multiplicityof5to10PFU/cell. After 30minat24 C, cultures werewashed and thenwere incubated at30
or38C for 48h.Culture fluidswerecleared of debris
by low-speed centrifugationandthenwereassayedfor
infectivity bytheplaquemethodat30 C.
A
30C
7
8~~~~~~_\-~3 C -_
6_
5-4
2 _ .A
VOL. 13,1974 299
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.493.247.441.63.234.2] [image:2.493.246.440.464.609.2] [image:2.493.44.236.533.666.2]therestrictive temperaturewerelessthan0.2%.
Most of the virus produced atNPtemperature wasofthemutant genotype,unable toproduce virus when tested again at 38 C, indicating slight "leakiness." Additionally, these results confirmed the previous observation that the
mutants havea low reversionfrequency towild type.
Complementation. The results of
com-plementation experimentsareshown in Table 3.
On the basis of thesedata, wehave assigned the mutantstosevencomplementationgroups(Ato
G). Five complementation groups (A and D to
G) arerepresented by single mutants(245, 105, 348, 74, 271). Group B is represented by two mutants (557, 642), and group C by three mutants (595, 840, 935). Within a group, each mutant complemented all other mutants, but
not mutants in the same group. Thus, the
groups arenonoverlapping.
Virus-specific RNA synthesis by ts mutants.Theability of Sendai virusts mutants to synthesize the various species of virus-specific RNA (1, 13) under restrictive conditions
wasexamined. After adsorption of virusat24C for 30 min, infected monolayerswereincubated at 38 C for 48 h. At this time, when viral RNA synthesis is maximal for wild-type virus (13), cultures were treated with actinomycin D and
labeled with [3H]uridine in the presence ofthe
drug. The RNAswereextracted and sedimented
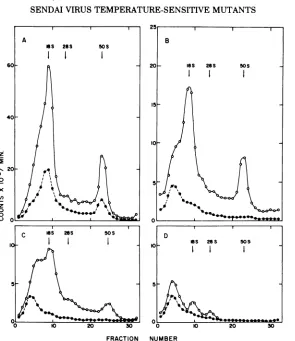
insucrosegradients. Results shown in Fig. 2are
representative of each type of physiological
behaviorobserved, although amember ofevery
complementation group was examined.
Similar amounts ofviral RNA were
synthe-sized by ts mutant 271 (group G) atP and NP
temperatures (excluding the adsorption period) (Fig.2A). However, thiswasthreefoldless than
wild-type virus under similar conditions. The distribution of radioactivity was the same as
that found when wild-type viruswasincubated
under the same conditions (13), i.e.,
virus-specific 18S messenger RNA and 50S genomes were made. Thus, we have designated
com-plementationgroupG RNA+. Mutants
belong-ing to complementation groups Ato F synthe-sized little or no RNA when incubated at the restrictive temperature from the beginning of' infection, and we have designated them
RNA-(Fig. 2B, C, and D).
To distinguish viral gene functions required
forRNA synthesis throughout replication from thoserequired only early in the cycle, tempera-ture shift (30 to 38 C) experiments were done.
Infected cultures wereincubated atthe P tem-perature for 48 h and then wereshifted to the NP temperature for 24 h. In all cases, virus
production ceased within 24 h. After the 24 h shift-up, the cultureswerelabeled with [3H
]uri-dine for 1 h in the presence ofactinomycin D,
and the RNAs were extracted and analyzed as
described above. As expected, ts mutant 271
(group G, RNA+) synthesized normal amounts
ofviral RNA after shift-up (Fig. 2A). Both the
mutant and wild-type virus synthesized more
RNA after being shifted from the P to NP
temperature.
Mutants belonging to groups A, B, and C
(RNA-) synthesized substantial amounts of 18
and 50S RNA after being shifted to the NP
temperature for 24 h (Fig. 2B). When the intervalatthe NPtemperaturewasextendedto
[image:3.493.63.460.476.608.2]48 h before labeling (data not shown), the
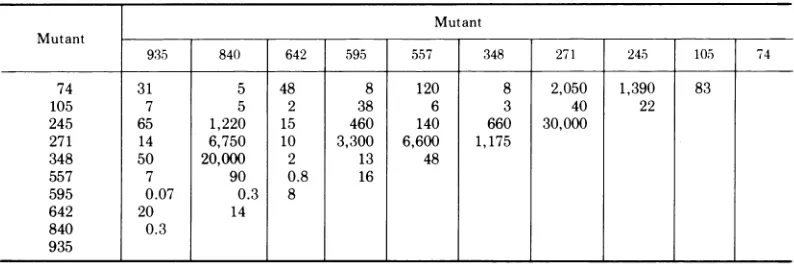
TABLE 3. Complementation levelsa
Mutant Mutant
935 840 642 595 557 348 271 245 105 74
74 31 5 48 8 120 8 2,050 1,390 83
105 7 5 2 38 6 3 40 22
245 65 1,220 15 460 140 660 30,000
271 14 6,750 10 3,300 6,600 1,175
348 50 20,000 2 13 48
557 7 90 0.8 16
595 0.07 0.3 8
642 20 14
840 0.3
935
aComplementation testsweredone as described in Materials and Methods. Complementation levels were calculatedaccordingtoBurgeand Pfefferkorn (3). All valuesgiven are the average of three experiments. These levels weresignificantly greater than 1 by the Student's t test at P < 0.05. Complementation levels divided the mutants intosevencomplementation groups: group A, mutant 245; group B,557and642; group C, 595, 840, and 935; groupD, 105; groupE, 348; group F, 74; and groupG, 271.
300
on November 10, 2019 by guest
http://jvi.asm.org/
SENDAI VIRUSTEMPERATURE-SENSITIVE MUTANTS
z
0 2,
2
I-x
z
0
u
[image:4.493.101.396.49.390.2]FRACTION NUMBER
FIG. 2. Sedimentation ofvirus-specific RNA from cells infected by Sendai virus mutants. CEL cell cultures wereinfected withabout5PFUIcell.After30min at 24 C one group of cultures(0) was incubated for 48 h at 38C. Another group(0)wasincubatedat 30C for 48 h and then was shifted to 38 C for 24 h. Each group then wastreatedfor 1 h with 50 ugof actinomycin D per ml and labeled for 1 h with 50
,Ci
of[3H]uridineper ml in the presence of the drug, all at 38C. RNA was extracted with sodium dodecyl sulfate-phenol (13) andcentrifugedat 20,000 rpmat 20Cfor16h in 34-mllinear 15 to30%osucrose gradients (13). A,Infection with mutant ts 271 (complementation group G); B, mutant ts 557 (complementation group B); C, mutant ts 74
(complementationgroupF);andD,mutant ts 105(complementation group D).
amount of virus-specific RNA made was the
same as after the 24 h shift.
Thus,
thesemutants are defective inearly functions
essen-tial for viral RNAsynthesis, but these functions
are not required laterin thereplicativecycle.
Mutants ts 348 and 74 (groups E and F,
RNA-)
synthesized
low levels ofvirus-specificRNAaftershift-upfor 24h(Fig.2C). ViralRNA
synthesis decreasedevenfurther (incontrast to
groups A to C) as the interval at the NP temperature was extended to 48 h (data not
shown).
The group D mutant (ts 105, RNA-) was
clearly separated physiologicallyfrom groups E
and
F.
Aftershift-upfor24h,ts105synthesized
little or no virus-specific RNA (Fig. 2D). (Thepeak ofRNAfound atabout 5Swas also found
in uninfected cells.) When the shift intervals
weredecreasedto 2 or 4 h(datanot shown), ts
105still showeda marked declineinviral RNA
synthesis, indicating that this mutant ceases
RNAsynthesis rapidly under restrictive
condi-tions.
(When
examined in the same way,mu-tants
belonging
to groupsE and Fsynthesized
normalamounts ofviralRNA.) From these data
it is apparent that mutants belonging to
com-plementation groupsD, E, and F aredefective
in functions required for virus-specific RNA
synthesis throughout the replicative cycle, but
the defect in group D may be more
intimately
related to the synthetic machinery for viral RNA.Mutant ts 271 has a
hemagglutinin
thatdoes notfunctionatNPtemperature. Mutant ts 271
synthesized
RNA, but not infectious virus, at the NP temperature. When theability
VOL.13,1974 301
on November 10, 2019 by guest
http://jvi.asm.org/
PORTNER, MARX, KINGSBURY
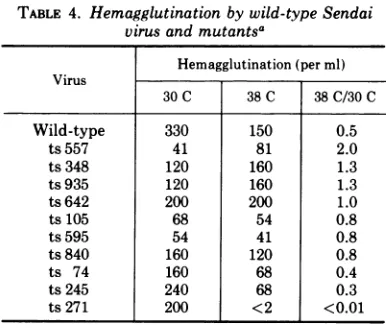
ofthis mutant to agglutinate erythrocytes was
examined, agglutinationoccurredatP, butnot
at NP temperature. Wild-type virus and the
other mutants agglutinated chicken
erythro-cytes atbothtemperatures
(Table
4). When themixture of virus and erythrocytes was shifted
from NP to P temperature, the erythrocytes
became fully agglutinated. When the
ag-glutinated mixture then wasshifted back from P to NP temperature, the hemagglutinin
pat-ternwasagainlost. These resultsindicated that
the hemagglutinin of mutant ts 271 did not
function atNP temperature, but that function
was restored at P temperature. However, the
inability to hemagglutinate chicken
erythro-cytes does not exclude the possibility that the
hemagglutinin was still capable ofbinding to
receptors.
Sendai virions contain at least six
polypep-tides (12). By
analogy
with otherparamyxovi-rions, the largest glycopolypeptide with an ap-parent molecular weight of 70,000 probably
represents the hemagglutinin (15, 16). The
inability of mutant ts 271 toagglutinate chicken
erythrocytesatNPtemperaturemight be dueto
analteration in someothervirionproteinwhich
makesthehemagglutinin nonfunctional.
There-fore, we tested isolated glycopolypeptides (15,
16) of ts 271forthetsphenotype. The prepara-tions contained all of the hemagglutinating
activity, and mainly virion glycopolypeptides
wereseen inpolyacrylamide gels (Fig.3,peaks2
and 5). When this material was tested for ts
hemagglutinin activity, the resultsweresimilar
tothose obtained withintact virions.Wild-type
TABLE 4. Hemagglutination by wild-type Sendai virus andmutantsa
Hemagglutination(perml) Virus
30C 38C 38C/30C
Wild-type 330 150 0.5
ts557 41 81 2.0
ts348 120 160 1.3
ts935 120 160 1.3
ts642 200 200 1.0
ts105 68 54 0.8
ts595 54 41 0.8
ts840 160 120 0.8
ts 74 160 68 0.4
ts245 240 68 0.3
ts271 200 <2 <0.01
aChicken erythrocytes (0.5% inPBS, pH 7.2) and
viruseswereseparatelywarmed in a water bath at 30 or 38C, mixed, and maintained at the respective temperatures. The test was performed in tubes, and titers representthereciprocal of the highest dilution
showinghemagglutination at 30 min after mixing.
[image:5.493.264.451.58.216.2]DiSTANCEMOVED (mm)
FIG. 3. Polyacrylamidegel electrophoresis of Sen-dai mutant ts 271
[3Hlglycopolypeptides
prepared as described in Materials andMethods.['4C]polypep-tides from wholewild-typeSendaivirions were added beforeelectrophoresis. Thenumberingsystem is that
ofMountcastle et. al. (12). Migration isfrom leftto
right.a 3H;
O,
4C.virion glycoproteins isolated the same way
ag-glutinated chicken erythrocytesat30and38
C,
whereas ts 271 glycoproteins were unable to
agglutinate
erythrocytes
at 38 C(data
wereidentical to those shown in Table 4). We
at-temptedto
identify
which of thetwoglycopoly-peptides from
wild-type
or mutant ts 271 wasthe
hemagglutinin,
but we were unable to separate theglycopolypeptides
by
centrifuga-tionin sucrose
gradients (15, 16). However,
it isclear that thets
hemagglutinin
resides inoneofthesolubilized viral
glycopolypeptides,
andnotinanother vironcomponentwhich
might
affecthemagglutinating activity
inintactvirions.Ifthesame
glycopolypeptide
hasneuramini-dase and
hemagglutinin
activities(15,
16),
thenmutant ts 271
might
have a tsneuraminidase. To test thispossibility, equivalent
amounts ofglycopolypeptideswere isolated from
wild-type
virus and ts 271, and neuraminidase activity
was measured at the P and NP temperatures
(Fig. 4). Therateofsialic acid releasebyts271
enzyme was about the same at 30 and 38C,
indicating that the neuraminidase was not
tem-perature-sensitive. Increasing the temperature
to 46C did not change the results. However,
comparedwithwild-type, the mutant
neurami-nidase functionedmuch lessefficiently (Fig. 4),
even at theP temperature.Additionally, when
ts 271andwild-typevirions weretestedforheat
stability at 54C, it wasfound that the mutant
hemagglutinin and neuraminidase activities
were more heat labile than wild-type virus.
After 90 s at54C, wild-typevirionshad35% of their original hemagglutinin and 30% of their
original neuraminidase activities, but mutant
302
x9
L)
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.493.55.248.436.600.2]SENDAI VIRUSTEMPERATURE-SENSITIVE MUTANTS
2.
2.0k
0.5-60 120
M NUTES
FIG. 4. Theeffect of30 and 38 Conneuraminidase activitiesoftheglycoprotein fraction from wild-type Sendai virus andts271.Replicatetubescontaining50
hemagglutinatingunitswereplacedat30or38 C. The
wild-typeviruspreparationcontained 5 ugof protein
perml,andts271contained 10 Mgof proteinperml.
Atintervals, onetube ofeach wasremoved, rapidly
chilled, andassayed forreleasedsialic acid(19, 20).
virions retainedonly3% of theirhemagglutinin andless than 1%of their neuraminidase
activi-ties. These data are consistent with the idea
that a single glycopolypeptide has
neuramini-dase and hemagglutinin activities, but other
explanationsare notruledout.
DISCUSSION
The Sendai virus genome has a molecular
weightof 6 x 106 (10). Thevirionpolypeptides have molecular weights from 40,000 to 80,000 (12). Thus, the genome may contain enough informationforeightto ten polypeptides.
Com-plementation experiments (Table 3) indicated
thatourSendai virusts mutantsweredefective
in seven different cistrons; thus, this small
seriesofmutantsmayalreadyrepresentmost of the viral gene functions. However, the number
of complementation groups may be high, be-causelowcomplementationvalues could reflect
intracistronic complementation or
complemen-tationbetweenmutants with multiple
tempera-ture-sensitivelesions.
The complementation groupings were
con-firmed in part by the physiological properties of
the mutants. We were able to classify the mutants into four categories according to their
ability to synthesize virus-specific RNA under
restrictive conditions. Complementation group
Gwas RNA+. Complementation groups A to F
synthesized neither viral RNA nor infectious
virus when incubated at the NP temperature
from the beginningof infection. However, after
shift-up late in infection, groups A, B, and C
synthesized substantial amounts of
virus-specific RNA, demonstrating that three gene
functionswererequired transientlyin the
repli-cative cycle for RNA synthesis to be initiated.
Groups D, E, and F appearedtobe defective in
functions required continuously for viral RNA
synthesis. The reason for the slow decline in
RNA synthesis with groups E and F after
shift-up is notapparent. One
possibility
isthatat NP temperature the temperature-sensitive
defect prevents the formation of a protein
required for RNA synthesis, but once the
pro-tein is formed it remains functional aftershift from 30 to 38 C. Such mutants have been
described forSemliki forestvirus (11). Mutant
ts105 (groupD) showedarapid decline inviral
RNA synthesis after shift-up, suggesting that
itsgene productturns overrapidlyor isunable
to maintain a functional configuration after
shift toNP conditions.
Itmay seemsurprising thataviruswithsuch
a small amount of genetic information should
possess six functions related to RNA synthesis.
But the three
early
functions represented bycomplementationgroupsA,B, and Care
proba-bly involvedinpreparing the viraltranscriptive
machinery for productive interaction with the
cell (entry,
"uncoating,"
transport, etc.),be-cause
they
are notrequired
later ininfection. Itshould be
emphasized
that thesefunctions areonly "early" with respect to RNA synthesis:
with respect to virus production they are also
"late," according to the results of shift-up
experiments. This suggests that theyrepresent
virion structural components.
One structural elementofvirionswhose
func-tion maynotberequired forviral RNA
synthe-sis is the hemagglutinin, asexemplified bythe
RNA+ phenotype of mutant ts 271. But it is
possible thatother mutants of thispolypeptide
will be discovered which will affect RNA
syn-thesis.
Recently,
we showed that virionenve-lope glycoproteins (12) and the smallest virion
polypeptide, also thought to be an
envelope
38C
30C WT
38C 271 ,,
n
30C0 A \
Al
/
0X
/ lll
303
VOL.13,1974
1.0~
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.493.44.236.55.365.2]component (10), inhibit virion transcriptase in vitro (P. A. Marx, A. Portner, and D. W.
Kingsbury, manuscript submitted for
publica-tion). Thus, these polypeptides are all
candi-dates for aroleinregulating viral RNA
synthe-S1S.
RNAsynthesis functions continually required
during infection, for which complementation
groups D, E, and F are candidates, would
include transcription and genome replication.
Transcriptive complexes isolated from virions
andinfected cells containtwopolypeptides, the
nucleocapsid structure unit (molecular weight
60,000) and thelargest virion polypeptide
(mo-lecular weight 75,000) (18; P. A. Marx, A.
Portner, and D. W. Kingsbury, manuscript
submitted for
publication).
Bothofthese maybe needed for transcriptase activity.
Polypep-tides involved in genome
replication
have notbeen identified.
It isclear thatwe cannot exclude any
virus-specified
polypeptide
from an involvement inparamyxovirus RNA
synthesis
at this stage inour
understanding. Hopefully,
additional workonthebiochemistry ofSendaivirus ts mutants
will contribute useful answers to these
ques-tions.
ACKNOWLEDGMENTS
Ruth Ann Scroggs and Andrew W. Moseley provided experttechnical assistance.
This research was supported by Public Health Service research grant AI-05343fromtheNational InstituteofAllergy and InfectiousDiseases,by traininggrantT01-CA-05176 and Childhood Cancer Center research grant CA-08480 from the National Cancer Institute, andby ALSAC.D. W.Kingsbury received Career Development Award HD-14,491 from the National InstituteofChild Health and HumanDevelopment.
LITERATURE CITED
1. Blair, C. D., and W. S. Robinson. 1968. Replicationof
Sendai virus. I. Comparison ofthe viral RNA and virus-specific RNA synthesis with Newcastle disease virus.Virology35i537-549.
2. Burge, B. W.,and E. R. Pfefferkorn. 1966.Isolationand characterizationofconditional-lethal mutantsof Sind-bis virus.Virology30:204-213.
3. Burge,B.W., andE. R. Pfefferkorn.1966. Complementa-tionbetweentemperature-sensitive mutants of Sindbis virus.Virology30:214-223.
4. Dahlberg, J. E., and E. H. Simon. 1969. Recombination in Newcastle disease virus (NDV): the problem of complementing heterozygot-s. Virology 38:490-492. 5. Darlington, R. W., A. Portner, and D. W. Kingsbury.
1970.Sendai virus replication: an ultrastructural com-parison of productive and abortive infections inavian cells. J. Gen. Virol. 9:169-177.
6. Fenner, F. 1969. Current topics in microbiology and immunology, p. 1-28, vol. 48. Springer-Verlag, New York.
7. Ghendon, Y. Z.1972.Conditional-lethal mutant of animal viruses. Progr.Med. Virol. 14:68-122.
8. Granoff, A. 1959. Studies on mixed infection with New-castle disease virus. I. Isolation of NewNew-castle disease virus mutants and tests for genetic recombination between them.Virology9:636-648.
9. Granoff, A. 1959. Studies on mixed infection with New-castle disease virus. II. The occurrence of NewNew-castle disease virus heterozygotes and study of phenotypic mixing involving serotype and thermal stability. Virol-ogy 9:649-670.
10. Kingsbury, D. W. 1972. Paramyxovirus replication, p. 1-33. In W. Arber et al. (ed.), Current topics in microbiology and immunology, vol. 59. Springer-Verlag, New York.
11. Martin, E. M. 1969. Studies on the RNA polymerase of sometemperature-sensitive mutants of Semliki forest virus.Virology 39:107-117.
12. Mountcastle, W. E., R. W. Compans, and P. W. Choppin. 1971.Proteins andglycoproteins of paramyxoviruses: a comparisonofsimian virus5,Newcastle disease virus, and Sendai virus. J. Virol. 7:47-52.
13. Portner, A., and D. W. Kingsbury. 1972. Identification of transcriptive and replicative intermediates in Sendai virus-intected cells. Virology 47:711-725.
14. Pringle, C. R. 1970. Genetic characteristics of condi-tional-lethal mutants ofvesicular stomatits virus in-duced by 5-fluorouracil, 5-azacytidine, and ethyl meth-anesulfonate. J. Virol. 5:559-567.
15. Scheid, A.,L. A.Caliguiri, R. W. Compans, and P. W. Choppin. 1972. Isolation of paramyxovirus glyco-proteins: association of both hemagglutinating and neuraminidase activities with thelarger SV 5 glycopro-tein.Virology 50:640-652.
16. Scheid, A., and P. W. Choppin. 1973. Isolation and purification of the enveloped proteins of Newcastle disease virus. J. Virol. 11:263-271.
17. Stone, H. O., A. Portner, and D. W. Kingsburg. 1971. Ribonucleic acid transcriptasesinSendai virions and infected cells. J. Virol. 8:174-180.
18. Stone, H. O., D. W. Kingsburg, and R. W. Darlington. 1972.Sendai virus-induced transcriptase frominfected cells: polypeptides in the transcriptive complex. J. Virol. 10:1037-1043.
19. Warren, L. 1959. The thiobarbituric acid assay of sialic acids. J. Biol. Chem. 234:1971-1975.
20. Webster, R.G., and C. H. Campbell. 1972. An inhibition test for identifying the neuraminidase antigen on influenza viruses. Avian Dis. 16:1057-1066.
304