MUTAGENIC SPECIFICITY OF A NOVEL T4 DNA
POLYMERASE MUTANT
.
LINDA J. REHA-KRANTZ AND EVELYN M. LIESNER
Department of Genetics, University of Alberta, Edmonton, Alberta, Canada T6G 2E9
Manuscript received July 10, 1983 Revised copy accepted October 25, 1983
ABSTRACT
The in vivo mutational specificity of a novel T 4 DNA polymerase mutator mutant, tsMI9, was determined. Two genetic tester systems were used to char- acterize the mutant. Results of our studies indicate that tsMI9 promotes tran- sition and transversion mutagenesis and, possibly, frameshift mutagenesis. Cen- tral G C base pairs in runs of three or more consecutive G:C base pairs may
be target sites for tsM19-induced transitions.
TUDIES of the T 4 DNA polymerase have been useful in determining the
S
contributions of the DNA polymerase to the fidelity of DNA replication. In addition to the DNA-synthesizing activity, the T 4 DNA polymerase has a 3’ + 5’ exonuclease activity that “proofreads” newly synthesized DNA by removing misincorporated nucleotides at the primer terminus (BRUTLAG andKORNBERG 1972; MUZYCZKA, POLAND and BESSMAN 1972); however the nu-
clease activity is not the only activity contributing to the fidelity of DNA replication. A novel bacteriophage T 4 mutator DNA polymerase, tsM19, has been isolated by a procedure designed to select mutants with high spontaneous mutation rates (REHA-KRANTZ and BESSMAN 1981). Unlike most other T 4 DNA polymerase mutator mutants, tsMI9 apparently does not have a reduced proofreading-exonuclease activity, but, rather, it is defective in specificity and makes errors by inserting incorrect nucleotides. Before characterizing tsMl9 further biochemically, we wanted to determine the mutational specificity, if any, of the new mutator in vivo. We have used two T 4 genetic tester systems both based on the T 4
rZZ
genes to determine the mutational specificity of tsM19. These tester systems will also be useful in characterizing other T 4 mutator DNA polymerases. The ultimate goal of these studies with tsMI9 and other mutator DNA polymerases is to determine the mechanisms of base se- lection or rejection by DNA polymerases.The first tester system, originally suggested by DRAKE and BALTZ (1976) and further developed for T 4 by RIPLEY (1975), is based on the conversion of an
rZZ
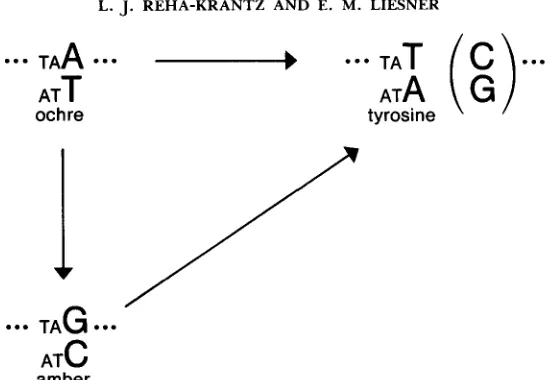
ochre nonsense mutant to the amber nonsense mutant or to wild type. The P7 ochre mutation in the T 4 rIZA gene can revert to either the amber mutant via an A:T 4 G:C transition or the wild-type DNA sequence (codon for tyrosine) via an A:T + T:A or C:G transversion (Figure 1). Thus,336 L. J. REHA-KRANTZ AND E. M. LIESNER
ATT
ochre tyrosine
ATA
\G/
ATC
amberFIGURE 1.-Mutational pathways at the rII P7 site. The P7 ochre mutant can revert from the A:T base pair via an A:T + T:A or C:G transversion to wild type (codon for tyrosine) or to the amber mutant via an A:T + G:C transition. The P7 amber mutant can revert via a G:C + T:A or C:G transversion to wild type.
it is possible to compare directly transitions and transversions from the A:T base pair at the P7 ochre site. Detection of the reversion of the amber con- vertant to wild
type
allows determination of the frequency of transversions from the G:C base pair. For all pathways, the mutational event is at the third position in the codon, and thus, all pathways share the same DNA environ- ment. This system requires that only a single amino acid be acceptable at the mutant site.T h e second system is based on the elegant studies of BENZER (1961) in the T4
rZZ
genes and the l a d system developed by COULONDRE and MILLER(1977a,b). A number of
rZZ
mutants induced by tsMl9 were isolated and mapped to therZZ
genes. A comparison of the tsM19-inducedrZZ
mutants with the map pattern of spontaneousrZZ
mutants can be used to identify "hot spots" or sites of high mutational activity. Furthermore, comparing tsMl9-induced nonsenserZZ mutants with the DNA sequence
(PRIBNOW et al. 1981) allows determination of the mutational pathways.We report here on the use of these mutational tester systems to characterize the mutational specificity of tsMl9.
MATERIALS AND METHODS
Phage and bacterial strains: T 4 rII mutants were described by BENZER (1961) and were obtained from S. CHAMPE, B. S. SINGER and P. GAUSS. The T 4 tester mutant, P7 ochre, was described by
DRAKE and MCCUIRE (1967) and RIPLEY (1975) and was obtained from L. RIPLEY. The T 4 gene 4 3 mutants (DNA polymerase) have been described (EPSTEIN et al. 1963; REHA-KRANTZ and
BESSMAN 1981). T 4 gene 4 3 rII double mutants were constructed by genetic recombination and selected by the method of DOERMANN and BOEHNER (1970). Double mutants were verified by backcrosses to the parental phages.
Media: Hershey broth, bottom-layer agar and top-layer agar were prepared according to CHASE
and DOERMANN (1958) with the addition of 1 g of sodium citrate/liter in the bottom-layer agar.
All phage and bacterial dilutions were in Hershey broth. Phage stocks were prepared in M 9
medium (ADAMS 1959) with 5 g of Difco casamino acids/liter ( M 9 + ) .
Measurement of rll+ rmersion frequencies: Fluctuation analyses patterned after LURIA and DELBRUCK (1943) were used to determine rll* reversion frequencies. Cultures (10 ml) of CR63 at 2 X lo*
cells/ml were inoculated with individual 7- to 8-hr plaques and aerated by shaking overnight at 30". High titer cultures of 1.2 X 10" to 3.4 X 10" phage/ml were obtained. The number of transversion mutations from P7oc and P7um was determined by plating on CR63(hh). The number of transition mutations from P7oc was determined by detecting the number of P 7 amber conver- tants on CA265(supF) and verified by replica plating on suo or supD h lysogens. The permissive bacterial strain in all cases was C R 6 3 . Ten parallel cultures were grown to determine each reversion frequency, and the data reported represent the median values of each set.
TsM 19-induced rII mutants: Mutator (tsM19)-induced rZZ mutants were isolated by preparing individual cultures of tsMl9 and titering each culture on E. coli B. A single r mutant plaque was picked from each culture and replica plated on a X lysogen to verify the rZZ mutant genotype. From 240 potential rZZ mutants, 114 tsMl9-induced rll mutants were isolated.
Mapping rII mutants: Deletion mapping (BENZER 1961) and two-factor crosses with known rll
mutants were used to position tsM19-induced rZZ mutants.
RESULTS
P7oc and P7am revert via genetically dejned mutational pathways: Possible path- ways of reversion from the P 7 site by single base pair changes were determined by testing the ability of amber and ochre suppressor strains to suppress P7oc and P7am (RIPLEY 1975). Tyrosine was found to be the only amino acid substitution at the P 7 site that produced a functional rIIA protein (Figure 1). All possible amino acid substitutions resulting from a single mutational event at the mutant site were examined except for glutamate, for which there is no suppressor strain (OZEKI et al. 1980); however, since serine, glutamine, leucine, tryptophan or lysine inserted at the mutant site do not restore wild-type activ- ity, it is likely that tyrosine is the only acceptable amino acid at the P 7 site.
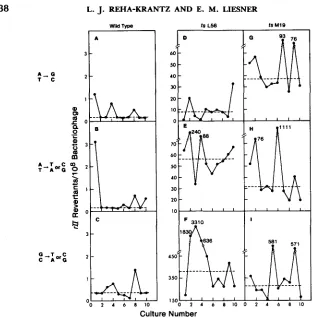
In a wild-type background, the P7oc and P7am sites reverted to rIP rarely at 0.1-0.3 revertants/lO* phage (Figure 2). Reversions via the transition and transversion pathways were approximately equal. T h e tsMl9 mutation en- hanced both transition and transversion pathways; reversions from the A:T base pair via the transition and transversion pathways were both increased
200-
fold in a tsMI9 background, and the number of reversions from the G:C base pair via the transversion pathway was increased >700-fold (Figure2).
Another strong DNA polymerase mutator mutant, tsL56, also increased the reversion of P7oc and P7am via the transversion pathways, but tsL56 was found to be a weaker mutator than tsMl9 for the A:T + G:C transition pathway (Mann- Whitney U test, 0.001 significance level, SIECEL 1956).Isolating and characterizing tsM 19-induced rII mutants: T h e second system is based on isolating forward mutations induced by tsM19 in the T 4 rII genes. Any distinguishing features in the map pattern of tsM19-induced rII mutants may provide additional information on the mutational specificity of tsM19 at a large number of sites.
338 L. J. REHA-KRANTZ AND E. M. LIESNER
:-E
ts L58 ts M19
Culture Number
FIGURE 2.-Mutational specificity of wild-type and DNA polymerase mutants. T h e median val-
ues of the fluctuation tests are indicated by dashed lines. T h e host strain for growing the cultures was CR63. T h e A:T + G C pathway is the P7oc to P7am transition pathway. T h e A:T + T:A or G:C pathway and the G:C + T:A or C:G pathway are the two transversion pathways from either
P7 ochre or P7 amber, respectively. Note that transitions and transversions from the A:T base
pair are determined from the same culture.
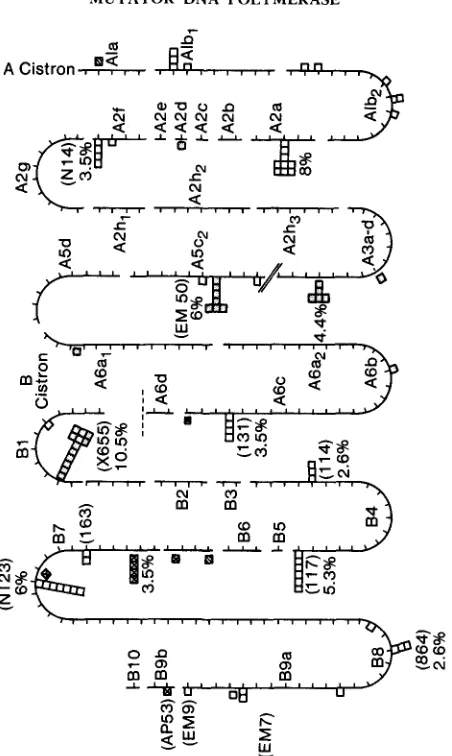
(at 131 in segment A6c and at 117 in segment B4). These sites, which are frameshift sites, represent only 8.8% of the mutants induced in a tsM19 back- ground. The other frameshift hot spot, 114 in segment B4, composed 3.1% of BENZER’S
rZZ
mutants, and a similar proportion (2.6%) was found in the tsM19 collection.A number of sites were unique to tsM19 and were not found in the BENZER
0
A Cistron
-
FIGURE 3.--TsMl9-induced rZZ mutants. Each square indicates a separate isolate; 96 tsM19- induced rlZ mutants from the total of 114 rZZ mutants collected are indicated in this figure. Some rlZ deletion segments in which there were few rZZ mutants are not included, as indicated by slashed, diagonal lines. Shaded squares are opal mutants, crosses indicate ochre mutants, diagonal lines indicate amber mutants and horizontal lines indicate temperature-sensitive mutants. A few of
BENZER’S rZZ sites are shown in parentheses.
For nonsense mutations in regions in which the DNA sequence is known, it is possible to correlate nonsense sites with specific mutational pathways, as done by COULONDRE and MILLER (1977a) for the lacl gene of E. coli. This analysis can be done for two tsMl9-induced rII hot spots. As shown in Table 1, the DNA sequence of the two hot spots is very similar, and the mutational pathway in both cases is a G:C + A:T transition.
Y,
U
5.
- E ,
44 3
2 %
b $
Y 9 2
w
E &
2
2
I $ U & U
2
k,y
k
2 .Y
s
.Y E 5z 5
z z g a
Y h: .E! b
.s
.- 0 a 0 .- o e , * e ,4 .= VI .=
m * . - U1
c 'j;
m .% > .t:
" 2 2 2
a m m m
.5 b b b Z k 5 k
< b < Y,
. r t T t
+ v i !
, 0 0 0
transition mutations from A:T or G:C base pairs. At least seven to ten of these sites are also in the tsMl9 collection.
DISCUSSION
In order to determine the mutational specificity of a mutagen, in this case a mutator DNA polymerase, tsMl9, it is necessary to develop genetic tester systems that measure the activity of the mutagen in well-defined mutational pathways. We have used two tester systems to determine the mutational spec- ificity of tsM19: a reversion assay system and a forward assay system. Each system alone has strengths and weaknesses. One potential problem with the first system, which depends on detecting revertants from a particular site in the DNA, is that this site could have special properties that could mask mu- tational activity. A second problem is that not all mutational pathways can be determined from the P7 site. Frameshift mutations and G:C + A:T transitions can not be determined, for example. Other tester sites can be used to over- come these problems, but we have used instead a second system based on isolating tsMl9-induced rII mutations. A strength of forward mutation assays is their ability to identify the most frequent types of mutation. These two systems combined provide information on the mutational activity of tsMl9.
Earlier studies on the mutational activity of tsMl9 indicate that tsM19 has strong mutator activity (REHA-KRANTZ and BESSMAN 198 1). T h e reversion frequency of the rII ochre mutant r W 1 9 9 is increased more than 50-fold above wild-type levels by tsM19 (REHA-KRANTZ and BESSMAN 198 l), and reversion frequencies of another rII ochre mutant, r W 1 8 ? , and the amber convertants of r W 1 9 9 and r W 1 8 ? are also increased 50- to 100-fold above wild-type levels
(L. REHA-KRANTZ, unpublished observations). Reversions via the A:T + G:C
transition pathway producing the amber convertants from r W 1 9 9 ochre and
r W 1 8 3 ochre are also increased more than 50-fold by tsM19. T h e rUI7199
mutant is reported to revert by transition from an A:T base pair, and r W 1 8 3
is reported to revert by either transition or transversion from A:T base pairs (DRAKE 1963), but it is difficult to determine the mutator activity for the various mutational pathways quantitatively, because there are many possible pathways at the r W 1 9 9 and rUV183 sites. We concluded from these earlier studies that tsM19 promotes strongly A:T + G:C transitions and may also promote transversions from A:T pairs and transitions from G:C pairs. T h e P7
site described here offers several advantages: the mutational pathways are de- fined, and all mutational events occur at the same position in the DNA, which minimizes DNA environmental effects (Figure 1). Transitions and transversions from the A:T base pair and transversions from the G:C base pair are all increased several hundredfold by tsM19 (Figure 2). This high activity in both transition and transversion pathways is in contrast to tsL56, another mutator DNA polymerase, which has lower activity in the transition pathway (Figure
2). Thus, for these mutational pathways tested, tsMl9 is a more general mu- tator than tsL56.
342 L. J. REHA-KRANTZ AND E. M. LIESNER
frameshift hot spots observed by BENZER (1961) are not strong hot spots in a tsM19 background. These two hot spots were not found in 55 rZZ mutants isolated each from two other mutator DNA polymerase mutants, tsL56 and tsL88 (SPEYER, KARAM and LENNY 1966); however, RIPLEY and SHOEMAKER (1983) have shown that tsL88 and tsL56 enhance the reversion of one frame- shift mutant, r l 3 l in segment A6c, 8- to 50-fold above wild
type.
T h e isolation of frameshift mutants in a tsM19 background, at least 13 independent isolates at the rl31, r117 and r114 sites from a total of 114 tsMl9-inducedrZZ
mutants, suggests that tsMl9 may act in frameshift mutagenesis; however, it appears that tsMl9 is more active in other pathways. For two tsM19 hot spots, EM50 and X655, the mutational pathway can be determined from the DNA sequence (Table 1). At both sites, the mutational lesion is due to a G:C + A:T transition. Thus tsMl9 enhances both A:T + G:C (Figure2)
and G:C + A:T transitions. At several other sites, it is possible to deduce the mutational pathway by using mutagens specific to certain pathways. Comparing the map patterns of muta- gen-inducedrZZ
mutants (Figure 3) indicates that, at seven to ten sites, the tsM19-induced mutations were due “likely” to G:C + A:T or A:T+
G:C transitions. Although these mutational pathways cannot be verified without DNA sequence data, it appears that many rZZ mutants induced by tsMl9 are transition mutants.Because both transitions and transversions were elevated in a tsMl9 back- ground at the P 7 site, it is surprising that none of the tsM19-induced mutants in which the mutational pathway can be verified are transversion mutants. There are a number of nonsense condons, for example, in segment B1 that can be generated by transversions (PRIBNOW et al. 1981; NELSON et al. 1981); howevqr, the only tsM19-induced nonsense mutants isolated in B1 are at a single site, X655 (Figure 3, Table 1). Similarly, there are numerous possible nonsense sites in segments A5c2 through A6d, yet there are only two tsMl9- induced nonsense mutant sites: the EM50 site, a tsMl9 hot spot, and a new opal mutant site in segment A6al (Figure 3). Thus, although there are nu- merous sites in which nonsense rZI mutants can be produced via transition or transversion pathways, only a few sites are mutated by tsMl9.
tsMl9 (6%). T h e adjacent amber and ochre nonsense mutations have not been observed. Thus, a target site for tsMl9 mutational activity may be at central sites within runs of three or more consecutive G:C base pairs.
As a first test of this model, the DNA sequence from segment A5c2 through B4 (PRIBNOW et al. 1981; A. SUGINO, personal communication) was inspected for similar DNA sequences. There are several sites at which there are runs of three or more consecutive G:C base pairs; however, mutations at these sites cannot be observed unless they produce nonfunctional rZIA or rZZB proteins. Thus, most missense mutations can not be detected. Nonsense mutations can be detected, and a search for DNA sequences similar to the tsMl9 hot spots that could mutate via transition or transversion mutagenesis to generate non- sense codons revealed two potential sequences: TGC* CAA and TGC* CTG. A transversion at the (*) position creates the UGA opal codon. T h e first sequence may be in segment A6al at the site of the tsMl9-induced opal mutant (Figure 3). T h e second sequence is in segment B1.These sequences, like the tsM19 hot spot sequences in Table 1, have runs of three consecutive G:C base pairs, but these sites do not appear to be preferential sites for tsM19 mutational activity (Figure 3). These sites differ from the hot spots in that a transversion, rather than a transition, is required to generate the opal mutation.
Earlier studies have predicted that G:C-rich sequences may be more mutable. KOCH (1971) demonstrated that changing an A:T base pair to a G:C base pair increased the mutation frequency at a neighboring site 8-fold. BESSMAN and REHA-KRANTZ (1977) have also postulated that high mutabiity may be associ- ated with G:C-rich DNA sequences, which have a stabilizing effect on inserted incorrect bases. Furthermore, nearest neighbor interactions, including purine- purine base-stacking interactions between incoming dNTPs and the primer terminus, have been shown to be important in misincorporation (TOPAL, DI-
GUISEPPI and SINHA 1980). In addition, the base analogue 2-aminopurine deox-
yribonucleotide (2AP) is incorporated in vitro preferentially after deoxyguan- ylate residues (Gua) by some T 4 DNA polymerases (PLESS, LEVITT and BESS-
MAN 1981). This bias in incorporation of 2AP after Gua has also been inter-
preted as due to stabilizing, stacking interactions. It is interesting that the EM50 and X655 hot spots are preceded by 3' Gua; however, the ochre mu- tations at these sites are also preceded by 3' Gua, but the ochre mutants are not detected (Table 1). T h e tsMI9 DNA polymerase has been shown to mis- incorporate nucleotides (REHA-KRANTZ and BESSMAN 198 1); these incorrect nucleotides are more likely to be misincorporated and not removed by the 3'
+ 5' exonuclease-proofreading activity of the DNA polymerase if the misin- serted nucleotides can be stabilized, such as in a G:C-rich DNA microenviron- ment. Further biochemical characterization of the tsM19 mutant DNA polym- erase will have to examine misincorporation activity in G:C-rich DNA se- quences, specifically at sites where G:C base pairs precede and follow the mutation site.
We wish to thank JENNIFER K. J. LAMBERT for isolating some of the tsM19-induced rII mutants and SUKRAN PARMAKSIZOGLU for technical assistance. We extend special thanks to S. CHAMPE, P.
344 L . J. REHA-KRANTZ AND E. M. LIESNER
provided by M. HOLM= and G. ARRAIZ. This research was supported by an establishment grant from the Alberta Heritage Foundation for Medical Research, by an operating grant from the Alberta Cancer Board-Alberta Heritage Savings Trust Fund (project H-246) and by an operating grant from the Natural Sciences and Engineering Research Council Canada (A8406). L. REHA- KRANTZ is a Scholar of the Alberta Heritage Foundation for Medical Research.
LITERATURE CITED
ADAMS, M . H., 1959 Bnc.[r,-io~~/itibrrs, p. 445. Interscience Publishers, New York.
~ N Z E R , S., 1961 47: 403-415.
0 1 1 the topography of the genetic fine structure. Proc. Natl. Acad. Sci. USA
Studies on the biochemical basis of spontaneous mutation. V. Effect of temperature on mutation frequency. J. Mol. Biol. 116: 115-123.
Enzymatic synthesis of deoxyribonucleic acid. XXXVI. A proofreading function for the 3' --* 5' exonuclease activity in deoxyribonucleic acid polym-
erases. J. Biol. Chem. 247: 241-248.
CHASE, M. and A. H. DOERMANN, 1958 High negative interference over short segments of the genetic structure of bacteriophage T4. Genetics 43: 332-353.
COULONDRE, C. and J. H. MILLER, 1977a Genetic studies of the Inc. repressor. 111. Additional correlation of mutational sites with specific amino acid residues. J. Mol. Biol. 117: 525-575. COULONDRE, C. and J. H. MILLER, 1977b Genetic studies of the lnc repressor. IV. Mutagenic
specificity i n the lncI gene of fik/wric.h/o col,. J . Mol. Biol. 117: 557-606.
DOERMANN, A. H. and I,. BOEHNER, 1970 T h e identification of complex genotypes in bacterio- phage T 4 . I. Methods. Genetics 66: 417-428.
DRAKE, J. W., 1963 Properties of ultraviolet-induced rII mutants of bacteriophage T 4 . J. Mol. Biol. 6 268-283.
DRAKE, J. W. and R. H. BALTZ, 1976 T h e biochemistry of mutagenesis. Annu. Rev. Biochem.
DRAKE, J. W. and J. MCGUIRE, 1967 Properties of r mutants of bacteriophage T 4 photodynam- ically induced in the presence of thiopyronin and psoralen. J. Virol. 1: 260-267.
EPSTEIN, R. H., A. BOLLE, C. M. STEINBERG, E. KELLENBERGER, E. BOY DE LA TOUR, R. CHEV-
ALLEY, R. S. EDGAR, M. SUSMAN, G. H. DENHARDT and A. LIELAUSIS, 1963 Physiological studies of conditional lethal mutants of bacteriophage T4D. Cold Spring Harbor Symp. Quant. Biol. 28: 375-394.
T h e influence o f neighboring base pairs upon base-pair substitution mutation
Mutations of bacteria from virus-sensitivity to virus-resistance.
Studies on the biochemical basis of mutation. I. A comparison of the deoxyribonucleic acid polymerases of mutator, antimutator, and wild type strains of bacteriophage T 4 . J. Biol. Chem. 247: 7 1 16-7122.
Mutations that detoxify an aberrant T 4 membrane protein. J. Mol. Biol. 1 4 9 377-403.
OZEKI, H., H. INOKUCHI, F. YAMOA, M. KODAIRA, H. SAKANO, T. IKENURA and Y. SHIMURA, 1980 Genetics of nonsense suppressor tRNAs in Esrhrrichici coli. pp. 341-362. In: Tmnsfbr
Rh'A: Biologiccil A.TI)Pc/s, Edited by D. SOLL, J. N. ABELSON and P. R. SCHIMMEL. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
Non random substitution of 2-aminopurine during deoxyribonucleic acid synthesis in vitro. Biochemistry 20: 6235-6244.
BESSMAN, M. J. and L. J. REHA-KRANTZ, 1977
BRUTLAG, D. and A. KORNBERG, 1972
4 6 11-37.
KOCH, R. E., 1971
rates. Proc. Natl. Acad. Sci. USA 68: 773-776. I.URIA, S. and M. DELBRUCK, 1943
Genetics 28: 49 1-5 1 1.
MUZYCZKA, N., R. L. POLAND and M. J. BESSMAN, 1972
NELSON, M. A., B. S. SINGER, L. GOLD and D. PRIBNOW, 1981
PRIRNOW, D., D. C. SIGURDSON, L. GOLD, B. S . SINGER, C. NAPOLI, J. BROSIUS, T . J. DULL and H . F. NOLLER, 1981 rll cistrons of bacteriophage T4: D N A sequence around the intercis- tronic divide and positions of genetic landmarks. J. Mol. Biol. 1 4 9 337-376.
Studies on the biochemical basis of mutation. VI. Selection and characterization of a new bacteriophage T 4 mutator D N A polymerase, J. Mol.
Biol. 145: 677-695.
Transversion mutagenesis in bacteriophage T 4. Mol. Gen. Genet. 141: 23-
A major role for bacteriophage T 4 DNA polymerase
,\IOIIPN).(o)Irt).if Strctis/ics f . r thr Brhcmiorcrl Scirncrs, pp. 1 16-1 27. McGraw-Hill,
REHA-KRANTZ, I,. J. and M. J. BESSMAN, 1981
KIPLEY, L. S., 1975
RIPLEY, L. S. and N. B. SHOEMAKER, 1983 40.
in frameshift mutagenesis. Genetics 103: 353-366.
New York.
SIEGEL, S., 1956
SPEYER, J. F., J. D. KARAM and A . B. LENNY, 1966
TESSMAN, I , 1965
TOPAI., M., S . R. DIGUISEPPI and N . K. SINHA, 1980
On the role of DNA polymerase in base selection. Cold Spring Harbor Symp. Quant. Biol. 31: 693-697.
Genetic ultrafine structure in the T 4 r l l region. Genetics 51: 63-75.
Molecular basis for substitution mutations: effect of primer terminal and template residues on nucleotide selection by phage T 4 D N A
polymerase i l l vitro. J. Biol. Chem. 255: 1 17 17-1 1724.